
Abstract
This review paper is aimed at presenting and discussing the effects of rumen digesta (RD) as an alternative protein source on nutrient utilization, rumen fermentation characteristics, growth performances, and milk production in ruminants. RD is a by-product of abattoir; it is a partially digested feed that mainly originates from rumen of ruminants. RD consisted of 13.5–46.1% crude protein and some essential nutrients (microbial cells, amino acids, minerals, and volatile fatty acids [VFAs]), which beneficially affect the ruminant production. Recent studies have shown that transfer of RD from buffalo to cattle improved nutrient digestibility. Ensiling RD with agro-industrial by-products improved the nutritional value of agro-industrial by-products, such as by improving protein contents and increasing their digestibility. In addition, dried RD could enhance ruminal fermentation, diet utilization, and rumen microorganisms in in vitro, beef cattle and buffalo, which has no adverse effect on production performance. Therefore, utilization of RD is suggested because it improves nutrient utilization, reduces feed costs, and controls waste disposal to the environment.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The growing livestock industry is producing an enormous volume of waste, which increasingly burdens urban and rural areas with improper disposal that poses problems to both humans and the environment (Cherdthong and Wanapat 2013; Uddin et al. 2018). The attitude of humans toward overcoming these problems is usually quite negative, either because nothing is done to remedy the damages or because it is motivated only by sheer necessity that is undertaken with reluctance (Uddin et al. 2018). However, researchers have recently made some farmers realize that livestock waste can be converted into animal feed. This necessitates the recycling of various wastes into livestock feed ingredients to arrest the competition between man and animal for food. Feeding livestock with abattoir waste (e.g., rumen digesta (RD)) results in reduced feed cost and lower prices of animal production (Agbabiaka et al. 2012; Elfaki and Abdelatti 2016). It contributes to self-sufficiency in protein and makes possible the integration of animal products, which can in turn solve some problems arising from improper waste disposal (Esonu et al. 2010; Uddin et al. 2018). Therefore, this review paper is aimed at presenting and discussing the effects of RD as an alternative protein source on nutrient utilization, ruminal fermentation characteristics, and blood metabolites in ruminants.
Rumen digesta
One of such is from abattoir, namely, RD, which includes fermented and non-fermented feed at various stages of digestion in the rumen. RD is not uniform but rather stratified into liquid, gas, and various particle sizes. RD causes water pollution by entering into the rivers, streams, and local water sources. The abattoir in Thailand annually produces RD of more than 41,000 tons of dry matter from 1.2 million ruminants (FAOSTAT 2018). Thus, conversion of RD waste into animal feed will enhance the flexibility of feed formulation and reduce environmental pollution. In addition, RD with animal basal feed can save the environment reducing fermentation mitigating the greenhouse gas (CH4) production (Uddin et al. 2018).
Dried RD (DRD) consists of 13.5 to 46.15% crude protein (CP), 6.3 to 34.9% crude fiber (CF), 40.5% neutral detergent fiber (NDF), and 8.9 to 31.8% nitrogen free extract (NFE) (Odunsi 2003; Okpanachi et al. 2010; Agbabiaka et al. 2012; Cherdthong and Wanapat 2013; Ojebiyi and Saliu 2014; Yitbarek et al. 2016) (Table 1). However, chemical composition might be different among areas influenced by the (1) feeding regimen, (2) length of the holding time between feeding and slaughter, (3) season, (4) feed resource diversity, and (5) selection of pastures by various animals in various areas (Cherdthong et al. 2014). In addition, it contains potential end products that beneficially affect rumen microbial activity (e.g., microorganism cells, essential amino acids, minerals, and volatile fatty acids [VFAs]) and did not contain any anti-nutritional substances (Okpanachi et al. 2010). Previous works have elucidated that substituting soybean with DRD in animal feed enhances nutrient digestibility and reduces the cost of diet in tilapia, (Agbabiaka et al. 2012), broiler (Esonu et al. 2010), and rabbit (Mohammed et al. 2013). Our reports have indicated that utilization of DRD could improve rumen fermentation, feed utilization, and rumen microorganisms in in vitro trail (Cherdthong and Wanapat 2013), beef cattle (Cherdthong et al. 2014, 2015) and buffalo (Seankamsorn et al. 2017). Thus, the results of the RD study in ruminant animals will be addressed below.
Utilization of RD for small animals
It has been demonstrated that RD contains no anti-nutritional factors (Agbabiaka et al. 2012). This has prompted some researchers to use the material to replace one protein source or another. Odunsi (2003) combined RD with blood meal to replace groundnut cake or fish meal in a layered diet and reported low values for the parameters monitored, namely, feed intake, weight gain, hen daily egg production, weight of eggs, and shell thickness for the laying birds. It was attributed to the unpalatable nature of the diet due to the influence of the blood meal, which agreed with the findings of Adeniji and Balogun (2002) in a study carried out on pullets.
Mohammed et al. (2013) used RD as a replacement for soybean in a rabbit diet and demonstrated that there was a linear enhancement in feed intake and average daily gain of rabbits when the level of RD was increased. Feed intake was reported to increase up to 40% based on the level of inclusion of RD, confirming the earlier report of Whyte and Wadak (2002). Rabbits fed diets containing RD compared well in terms of performance with those on conventional feeds (Esonu et al. 2006). Esonu et al. (2010) suggested that the enhanced animal efficiency could be attributed to the higher protein of the undigested starchy and fibrous carbohydrates, long-chain fatty acids, and partially digested diet protein material due to the influence of the microbial protein in the RD. In terms of carcass characteristic evaluation, Esonu et al. (2006) reported that the values of the dressed weight, gizzard, and heart were similar to those of the control group, and the values indicated that the RD was well utilized by the birds. The reasons advanced were that the enhanced performance could be due to sufficient dietary fiber level, which may have activated the intestine, as well as higher occurrence of peristaltic movement and more enzyme activity, resulting in efficient digestion and utilization of the feed.
Rumen digesta transfer study
Winugroho et al. (1993) demonstrated the RD transfer technique from ruminants consumed in suitable feed to target ruminants in order to improve digestibility of nutrients by stimulation of ruminal bacteria. RD transfer could enhance utilization of dried Calliandra calothyrsus in sheep, which has been previously found to be of low nutrient value compared with the fresh form. Widiawati and Winugroho (1996) found that RD fill transfer could enhance the use of a diet supplemented with dried Calliandra calothyrsus, and native cattle were the significant animal donors for sheep.
In addition, a comparative experiment between buffalo and cattle in potential transfer of buffalo RD into cattle was elucidated by Wanapat et al. (2003). The RD (about 50% by weight of total digesta) from each rumen of fistulated buffalo was transferred to the rumen of fistulated cattle. These transfers were done as rapidly as possible to reduce extended exposure of digesta to the air. After the transfer was completed, all lids of fistulae were closed. Monitoring of RD for 14 days after transfer demonstrated enhanced microbial population in cattle as compared with that of original cattle. This indicates that buffalo RD could be transferred (Wanapat et al. 2003).
Ensiled rumen digesta with roughages
Silage can be made from plant materials with suitable moisture content depending on the methods of storage, degree of compression, and water content of feed materials. Ensiling of RD with rice straw helps to improve the nutritional value, such as by improving protein contents and reducing pH, fiber contents, and lignin (Maigandi et al. 2002). Furthermore, the ensilation of RD with agro-industrial by-products increases their digestibility. Thus, ensilation of rumen with crop residue can save the environment and at same time fulfill the nutritional requirements of the ruminants (Elfaki and Abdelatti 2016). The acceptability and feeding value of RD could be improved by ensiling, a controlled ruminal fermentation used to enhance the feeding value of feedstuffs, as reported by Gerald and Thomas (2006). Ensiling aids in masking the odor of feed materials, including RD, because odor is one of the factors affecting its efficient utilization (Maigandi et al. 2002). Ensiling of RD was found to improve its nutrient value, although researchers further stated that if the process exceeds 4 weeks, the protein in the material may be converted to NH3 and could hinder the supply of the needed protein to the animal for normal growth (Maigandi et al. 2002). Muhammad et al. (2016) concluded that ensiling RD with cowpea hay improves feed intake and digestibility, as well as increasing average daily gain from 71.43 to 90.77 g/day when compared with the no-RD group.
Feeding rumen digesta in ruminants
Today, manipulation of the rumen using dietary resources is increasingly attractive. Feeding of RD to ruminant animals as a protein source involves not only manipulating rumen fermentation but also recycling waste generated from abattoir. In addition, replacing high-quality protein sources with RD could improve rumen fermentation and digestibility of nutrient in ruminants (Elfaki and Abdelatti 2016). Thus, use of RD in animal diets will enhance the flexibility of feed and reduce pollution (Cherdthong et al. 2015). Inclusion of RD could improve digestibility of nutrients in ruminants. Furthermore, digestibility of NDF enhanced (55.6 to 59.9%) with DRD supplementation in the ration was greatest when RD was added for beef cattle (Cherdthong et al. 2014). Similarly, addition of RD to 8% of substrate increased in vitro true digestibility by 11.5% when compared with the no-RD group (Cherdthong and Wanapat 2013). Furthermore, Seankamsorn et al. (2017) reported that buffalo fed 150 g of RD pellets exhibited the greatest CP and fiber digestibility. The high potential for the proliferation of ruminal microorganisms and improvement of intake and digestion of nutrients should be noted. Similarly, rabbits fed with 20–40% RD were better able to utilize fiber than those on a no-RD diet, which could possibly be due to the presence of microorganisms in the caecum that act on fiber digestion (Mohammed et al. 2013).
Effect of dried rumen digesta on the rumen fermentation
Effect of various levels of rumen digesta on concentrations of total volatile fatty acid (TVFA) and VFA profiles is shown in Table 2. Ruminal fermentation of substrate by ruminal microbes results in production of microbial protein cells, NH3-N, and VFAs. Feeding of DRD did not adversely affect ruminal pH, temperature, and ruminal NH3-N (Cherdthong and Wanapat 2013). Ruminal total VFA concentration was not changed by DRD, and the mean values ranged from 110 to 130 mmol/L (Cherdthong et al. 2014). It was indicated that the DRD could replace soybean meal up to 100% DM in a ruminant diet without negatively affecting rumen fermentation parameters. Furthermore, Seankamsorn et al. (2017) revealed that VFA concentration profiles were not affected by DRD pellet (DRDP)–level supplementation except propionate, which was greatest when DRDP was added at 150 g/day. The enhancement of propionate concentration could possibly be due to the improvement of feed intakes, nutrient digestibility, and ruminal bacterial activity. In agreement with our previous study, in vitro propionate concentration was greater when DRD was included compared with no-DRD supplementation (Cherdthong and Wanapat 2013).
Effect of dried rumen digesta on rumen microorganisms
Effect of DRD on microbial population diversity is shown in Table 3. The rumen is a dynamic system, in which resident microorganisms must adapt consistently to changes in feed component, quantity, and consumption frequency. Cherdthong et al. (2015) used a molecular technique (real-time PCR) to determine the population of fibrolytic microorganisms. The study indicated that population of R. flavefaciens was increased with high DRD levels in the ration. Population of R. flavefaciens was greatest when replacing soybean meal by DRD at 100% in the concentrate diet. It could possibly be that the considerable numbers of essential nutrients in DRD (e.g., microbial cells, vitamin) improved the ruminal bacteria population (Liu et al. 2002). Okpanachi et al. (2010) revealed that the biological value of bacterial cells, minerals, and essential by-products in the DRD was found to be high, therefore resulting in efficient microbial cell production related to nutrient digestibility in animals. Seankamsorn et al. (2017) demonstrated that the fungal zoospores were enhanced when DRDP level increased and were significantly greatest when DRDP was added at 150 g/day. Moreover, supplementing of DRDP could provide a continuous ammonia-nitrogen and energy supply for fungal cell synthesis and enhance fungal activities in the rumen of ruminants. Therefore, increasing the fungal population could increase fiber digestibility and roughage intake. Similarly, Cherdthong and Wanapat (2013) elucidated that addition of 8 mg/5 g substrate of DRD could enhance in vitro microbial mass by 23.4 mg.
The effect of DRD supplementation on nitrogen (N) utilization is shown in Table 4. Microbial protein synthesis affects the demand for protein in ruminant animals (NRC 2001). Cherdthong et al. (2014) elucidated that DRD replacing soybean meal (SBM) at 100% increased ruminal microbial CP (MCP) and the efficiency of microbial N synthesis, which is reflected in improved microbial concentration in the rumen fluid. From the study above, MCP and efficiency microbial N synthesis may depend on substances contained in DRD, such as bacterial cells, essential amino acids, and mineral content, rather than the CP quantity from soybean meal.
Nitrogen consumed by ruminants ultimately appears in either the urine or feces or is interchanged with the body’s N reserves. Cherdthong et al. (2014) reviewed that N intake and N excretion from urine and feces were not altered with DRD replacing SBM at 100% in the ration. Thus, substituting SBM with DRD could be suggested for suitable feed sources without adversely affecting N utilization.
Effect of dried rumen digesta on growth performance and milk production
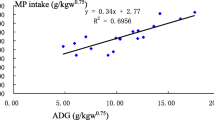
Mondal et al. (2012) demonstrate that there is no adverse impact of RD on the digestibility of nutrients, growth, and feed efficiency and thus may be considered absolutely safe for animal consumption and can be included in the diet of goats without any deleterious influence on the nutrient utilization. In addition, feeding Awassi lambs diets containing RD up to 10% improved average daily gain (ADG), feed conversion ratio (FCR), and feed efficacy (DMI:ADG) which has been revealed by Al-Wazeer (2016). These results agree with Osman et al. (2015) who found that final BW, total weight gain, and daily weight gain were highest in Shugor desert lambs fed 10% RD and least in animals fed no RD. Furthermore, Khan et al. (2014) reported that replacement of RD at 50% of conventional total mixed ration (TMR) could improve ADG at 0.71 kg which was not altered when compared with the no-RD fed group (0.73 kg of ADG). The improved daily weight gain with increased levels of RD in rations was mainly due to rumen contents’ high nutritive value (Uddin et al. 2018). Abouheif et al. (1999) found no significant difference in the growth performance, hot carcass weight, and dressing percentage of animals fed with a dietary mixture of RD and barley mixture at ratio 4:1 compared with the control group.
Feeding goats with enzyme-treated RD as replacement for Berseem clover (BC) on milk production was elucidated by Khattab et al. (2011). In this regard, partial replacement of BC by 50% treated RD increased milk yield and 4% FCM compared with untreated RC ration. The diets containing RD positively influenced fat content compared with the control. Also, milk fat, total solids, total protein, and lactose percent were increased with treated RD compared with the untreated group (Khattab et al. 2011).
Limitations of rumen digesta utilized as protein source
Ruminant diet production increasingly relies on the global acquisition of diet material, increasing the risk of bacterial and chemical contaminants being transferred into feed-producing ruminants (Wanapat 2004). There are some concerns when RD is introduced as a protein source in animal feed. Firstly, the contamination of pathogens from the animal host may influence the quality of RD. Thus, sources of RD should be considered, and it should be confirmed that the animal host was not infected by pathogens. In addition, preparation process of RD before being fed to the animals, such as being heat-treated or sun-dried, and pellets could reduce the pathogen contamination in RD. Secondly, the quality of RD may not be stable, which depends on many factors, such as feeding regimen, the type of feed resources, and selectivity of pastures in various areas (Agbabiaka et al. 2012). Thirdly, the drying time of RD before being used in animal rations is important. Two to 3 days are required for sun-drying RD until it reaches 95% DM. Lastly, the high fiber content in RD might affect the digestibility in animals; thus, optimum levels of supplementation should be considered.
Conclusions
Feeding of RD in animal diets might enhance nutrient utilization without adverse effects to the ruminants. Thus, conversion of RD to a ruminant animal diet will enhance the flexibility of the diet recipe and reduce the amount of waste to the environment. However, there remain some concerns with pathogenic contamination when RD is introduced as a protein source in animal feeds; therefore, before incorporating RD in the ration, safety should be verified.
References
Abouheif MA, Kraidees MS, Al-Selbood BA. 1999. The utilization of rumen content-barley meal in diets of growing lambs. Asian-Australasian Journal of Animal Sciences, 12, 1234–1240.
Adeniji AA, Balogun OO. 2002. Utilization of flavor treated blood-rumen content mixture in the diets of laying hens. Nigerian Journal of Animal Production, 29, 34–39.
Agbabiaka LA, Madubuike FN, Amadi SA. 2012. Studies on nutrients and anti nutrients of rumen digesta from three most domesticated ruminants in Nigeria. Pakistan Journal of Nutrition, 11, 580–582.
Al-Wazeer AAM. 2016. Effect of different levels of dried rumen content on nutrient intake, digestibility and growth performance of Awassi lamps. International Journal of Advanced Research, 4, 2106–2113.
Cherdthong A, Wanapat M. 2013. Manipulation of in vitro ruminal fermentation and digestibility by dried rumen digesta. Livestock Science, 153, 94–100.
Cherdthong A, Wanapat M, Saenkamsorn A, Waraphila N, Khota W, Rakwongrit D, Anantasook N, Gunun P. 2014. Effects of replacing soybean meal with dried rumen digesta on feed intake, digestibility of nutrients, rumen fermentation and nitrogen use efficiency in Thai cattle fed on rice straw. Livestock Science, 169, 71–77.
Cherdthong A, Wanapat M, Saenkamsorn A, Supapong C, Anantasook N, Gunun P. 2015. Improving rumen ecology and microbial population by dried rumen digesta in beef cattle. Tropical Animal Health and Production, 47, 921–926.
Elfaki MOA, Abdelatti KA. 2016. Rumen content as animal feed: A review. Journal of Veterinary Medicine and Animal Production, 7, 80–88.
Esonu BO, Ogbonna UD, Anyanwu GA, Emenalom OO. 2006. Evaluation of performance, organ characteristics and economic analysis of broiler finisher fed dried rumen digesta. International Journal of Poultry Science, 5, 1116–1118.
Esonu BO, Azubuike JC, Udedibie ABI, Emenalom OO, Iwuji TC, Odoemenam V. 2010. Evaluation of the nutritive value of mixture of fermented bovine blood and rumen digesta for broiler finisher. Journal of Natural Sciences Research, 1, 1–8.
FAOSTAT. FAO Statistics Divisions. 2018. http://faostat3.fao.org/home/index.html#HOME [accessed 12 Aug 2018].
Gerald MW. 2006. Thomas M. Processing Cattle Waste for recycling as animal feed. World Animal Review. http:www.fao.org/ag/aga.com [accessed 15 Aug 2017]
Khan MW, Pasha TN, Koga A, Anwar S, Abdullah M, Iqbal Z. 2014. Evaluation and utilization of rumen content for fattening of nili-ravi male calves. The Journal of Animal and Plant Sciences, 24(Suppl. 1), 40–43.
Khattab HM, Gado HM, Kholif AE, Mansour AM, Kholif AM. 2011. The potential of feeding goats sun dried rumen contents with or without bacterial inoculums as replacement for Berseem clover and the effects on milk production and animal health. International Journal of Dairy Science, 6, 267–277.
Liu JX, Susenbeth A, Südekum KH. 2002. In vitro gas production measurements to evaluate interactions between untreated and chemically treated rice straws, grass hay, and mulberry leaves. Journal of Animal Science, 80, 517–524.
Maigandi SA, Tukur HM, Daneji AI. 2002. Fore-stomach digesta in the diet of growing sheep 1. Performance and economics of production. Sokoto Journal of Veterinary Sciences, 4, 16–21.
Mohammed G, Adamu SB, Igwebuike JU, Alade NK, Asheikh LG. 2013. Nutrient digestibility and carcass measurement of growing rabbits fed graded levels of bovine blood-rumen content mixture. Pakistan Journal of Nutrition, 12, 929–933.
Mondal S, Haldar S, Samanta I, Samanta G, Ghosh TK. 2012. Exploring nutritive potential of undigested rumen contents as an ingredient in feeding of goats. Animal Feed Science and Technology, 13, 79–88.
Muhammad N, Bello MM, Maigandi SA, Magaji AA, Garba MG, Yusuf A. 2016. Evaluation of ensiled for stomach digesta (FSD) with cowpea hay on the performance of fattening sheep in semi-arid environment. European Journal of Basic and Applied Sciences, 3, 20–31.
National Research Counci. 2001. Nutrient Requirements of Beef Cattle, 7th ed. National Academy of Sciences: Washington.
Odunsi A. 2003. A blend of bovine blood and dried bovine rumen digesta as a replacement for fishmeal and groundnut cake in layer diets. International Journal of Poultry Science, 2, 58–61.
Ojebiyi OO, Saliu AS. 2014. Effects of feeding bovine rumen content-blood meal (50:50) mixtures on performance and slaughter characteristics of growing rabbits. Journal of Animal and Plant Sciences, 2, 430–434.
Okpanachi U, Aribido SO, Daikwo IS. 2010. Growth and haematological response of growing rabbits to diets containing graded levels of sun dried bovine rumen content. African Journal of Food, Agriculture, Nutrition and Development, 10, 4444–4457.
Osman AAB, Hamed AHM, Elimam ME. 2015. Effects of dried rumen contents level in rations on the performance of Shugor desert sheep in Halfa Elgadeda, Kassala state, Sudan. Animal Review, 2, 81–86.
Seankamsorn A, Cherdthong A, Wanapat M, Supapong C, Khonkhaeng B, Uriyapongson S, Gunun N, Gunun P, Chanjula P. 2017. Effect of dried rumen digesta pellet levels on feed use, rumen ecology, and blood metabolite in swamp buffalo. Tropical Animal Health and Production, 48, 79–86.
Uddin MJ, Hossainand MN, Kawsar MH. 2018. Recycling of rumen digesta: A substitute of goat feed and means of decreasing environmental pollution. IOSR Journal of Agriculture and veterinary Science, 11, 1–7.
Wanapat M. 2004. Current livestock production and protein sources as animal feeds in Thailand. In Protein Sources for the Animal Feed Industry. FAO, Rome, Italy. 199–214.
Wanapat M, Nontaso N, Yuangklang C, Wora-anu S, Ngarmsang A, Wachirapakorn C, Rowlinson P. 2003. Comparative study between swamp buffalo and native cattle in feed digestibility and potential transfer of buffalo rumen digesta into cattle. Asian-Australasian Journal of Animal Sciences, 16, 504–510.
Whyte EP, Wadak L. 2002. Evaluation of rumen content in the growth performance of rabbits. Proceedings of 7th Annual Conference of Animal science Association of Nigeria. (ASAN). Sept. 16–19 University Of Abeokuta. 300–303.
Widiawati Y, Winugroho M. 1996. Rumen fill transfer to improve digestibility of dried Calliandra (Calliaindra calothyrsus) supplementation diet in sheep. Proceedings of the Australian Society of Animal Production, 21, 421.
Winugroho M, Sabrani M, Punarbowo P, Widiati Y, Thalib A. 1993. Non-genetic approach for selecting rumen fluid contain specific microorganism (Balitnak method). llmu dan Peternakan, 6, 5–9.
Yitbarek MB, Mersso BT, Wosen AM. 2016. Effect of dried blood-rumen content mixture (DBRCM) on feed intake, body weight gain, feed conversion ratio and mortality rate of SASSO C44 broiler chicks. Journal of Livestock Science, 7, 139–149.
Acknowledgments
The authors would like to express our sincere thanks to the Increase Production Efficiency and Meat Quality of Native Beef and Buffalo Research Group, Khon Kaen University (KKU), Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, KKU.
Funding
This work was supported by the Thailand Research Fund (TRF) contract grant IRG5980010.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Cherdthong, A. Potential use of rumen digesta as ruminant diet—a review. Trop Anim Health Prod 52, 1–6 (2020). https://doi.org/10.1007/s11250-019-02018-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11250-019-02018-6