
Abstract
In this study, we conducted an investigation to determine the true prevalence of bovine and ovine brucellosis in central-eastern Tunisia. A total of 1134 veterinary samples taken from 130 ruminant herds were screened for brucellosis using IS711-based real-time PCR assay. Sera collected from the ruminants were tested using the Rose Bengal test and indirect enzyme-linked immunosorbent assay. Based on serological and molecular results, the true adjusted animal population level prevalence was 23.5 % in cattle, against 13.5 % in sheep. In addition, the true adjusted herd level prevalence of brucellosis was 55.6 % in cattle and 21.8 % in sheep. A statistically significant association was found between vaginal and milk shedding for ruminants. In addition, our results showed that Brucella abortus could be responsible for bovine and ovine brucellosis. Multivariable logistic regression analysis at the animal population level indicated that age and origin variables were important risk factors for cattle. However, age and abortion variables were found to be associated with ovine brucellosis. At the herd level, risk factors for Brucella positivity were as follows: abortion and herd composition for cattle against herd composition, mortality rates, and hygiene for sheep. Animal hygiene, food quality, and sanitary practices on the farm should be applied as strategies to control brucellosis in herds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Like in all African countries, breeding of ruminants plays an important role in the Tunisian national economy (Zaibet et al. 2009). Despite its importance, this sector suffers from certain fragility in terms of balance between food supply and livestock population (Zaibet et al. 2009). Moreover, ruminants are exposed in Tunisia to different health problems, such as respiratory and gastrointestinal parasitic infections and principally abortive diseases (Akkari et al. 2013). Several pathogens, including fungal, protozoan, viral, and bacterial agents, are known to affect directly the reproductive health of ruminants. Among these infectious agents, we note Brucella which is one of the most important abortifacient agents, causing brucellosis in ruminants.
Brucellosis is a major infectious disease of livestock with a global occurrence (OIE 2009). It is mainly caused in cattle by Brucella abortus. However, certain cases of bovine brucellosis have been related to other Brucella species such as Brucella melitensis and Brucella suis (OIE 2009). After abortion, several factors cause the transmission of bovine brucellosis like the contact of susceptible animals with contaminated fetuses, fetal membranes, uterine secretions, vaginal discharges, and milk (Mai et al. 2013). Brucellosis in sheep is principally caused by B. melitensis, although this bacterium may also infect other ruminants (Ducrotoy et al. 2014). Infection of ewes by B. melitensis is characterized by reduction of milk production and abortion or birth of weak lamb and can be transmitted by the same ways given in cattle (Megid et al. 2010). About two thirds of B. melitensis infections of pregnant ewes lead to infection of the udder and excretion of the bacteria in milk during the subsequent lactation (Xavier et al. 2010). B. ovis can occasionally cause abortion of ewes.
Many previous investigations were conducted in some regions of Tunisia and demonstrated the presence of antibodies against Brucella spp. (Chakroun and Bouzouaia 2007; Zaibet et al. 2009). Nevertheless, there is still a gap of information for many parts of the country. Indeed, in the governorate of Sfax, annual investigations made by state veterinarians during vaccination campaigns are not very effective. The present study aimed at (i) determining the true prevalence (TP) of Brucella infection among cattle and sheep in the central-eastern Tunisia and (ii) identifying potential risk factors associated with brucellosis positivity at animal and herd levels.
Materials and methods
Ethical considerations
The study protocol was assessed and approved by the institutional review board (IRB) of the Regional Office of Agricultural Development of Sfax in collaboration with the Veterinary Research Center of Sfax in Tunisia. Samples were collected by authorized veterinarians of the Veterinary Research Center of Sfax.
Study area
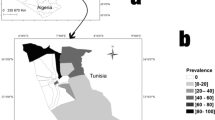
The present study was conducted in Sfax town (average altitude of 13 m), which is located in the East center of Tunisia, at 270 km from the capital Tunis. During this study, many geographical regions of Sfax have been visited (Fig. 1).

Location of the study area in Tunisia
Animals and samples
The number of herds to be studied was calculated using WinEpiscope program version 2.0, and it was based on the formula n = [t2 Pexp (1-Pesp)]/d2 (Thrusfield 2005). For that purpose, an expected herd prevalence of 7 % (Zaibet et al. 2009), a desired absolute precision of 10, and a 95 % confidence interval were considered. However, a contingency of 20 % was added (Boukary et al. 2013), and sample size was adjusted to 130 herds to increase allocations to agro-ecological strata. In total, 20 Holsteins bovine herds and 110 ovine flocks were visited. A total of 1134 samples (378 blood, 378 vaginal swab, and 378 milk samples) were collected by veterinarians and sent to the laboratory. These samples were collected as described by Barkallah et al. (2016).
Serological assays
The Rose Bengal test (RBT) (Institut Pourquier, France) was conducted according to the OIE protocols (2009). In addition, sera were tested using ID Screen Brucellosis Serum Indirect Multi-Species ELISA Kit (ID VET diagnostics, France) for the presence of IgG antibodies against B. abortus, B. melitensis, and B. suis. As recommended by the manufacturer, all sera were considered to be negative when % S/P < 110 %, suspicious when 110 % ≤ % S/P < 120 %, and positive when % S/P ≥ 120 %. The diagnostic sensitivity and specificity of the ELISA Kit were assumed to be 99 and 98 %, respectively. Sera producing positive results in both tests were considered as positive for Brucella spp.
DNA extraction
All vaginal swab, blood, and milk samples collected from animals were extracted by ZR Fungal/Bacterial DNA MiniPrep™ D6005 Kit (Zymo Research) as described by Barkallah et al. (2016).
Rt-PCR assays
Rt-PCR targeting the IS711 gene of Brucella spp. was performed as described by Hinic et al. (2008). PCR positive samples for Brucella spp. were tested by SYBR Green rt-PCR assays specific for B. abortus, B. melitensis, and B. ovis, using the primers described elsewhere (Hinic et al. 2008). Each reaction was run in a mastermix containing 12.5 μl of 2× SYBR Permix Ex Taq Tli RNaseH Plus (TaKaRa), 0.2 μM of each primer (Table 1), and 5 μl of purified DNA to a final volume of 25 μl using nuclease-free water. All amplifications were conducted by incubating the samples at 95 °C for 30 s, followed by 40 cycles of 5 s at 95 °C, and 30 s at 60 °C. Melt-curve analysis was performed immediately after the amplification protocol (60–95 °C). DEPC-treated H2O was used as a negative PCR control. Positive samples were sent to GATC Biotech SARL (France) for sequencing.
Statistical analysis
The TP of brucellosis at the animal level was estimated using the following formula: TP = (AP + Sp-1)/(Se + Sp−1) (Rogan and Gladen 1978). Herd level prevalence was calculated as the number of herds with at least one positive animal divided by the total number of herds tested. The true herd prevalence (THP) was estimated from distributions of herd sensitivity (HSe = 1−(1−AP)n) and specificity (HSp = Spn) (Musallam et al. 2015). The risk factor analysis was performed separately at the individual animal and herd levels for sheep as described by Barkallah et al. (2016). Statistical analysis was performed using the Statistical Package for Social Sciences (SPSS) version 19 software.
Results
Detection of Brucella spp.
Among 214 bovine sera, 52 (24.3 %) and 43 (20.09 %) were RBT and i-ELISA positive, respectively (Table 2). Our analysis also showed that 24 (31.3 %) and 14 (8.53 %) out of 164 investigated ovine sera were positive for Brucella using the RBT and i-ELISA, respectively (Table 2).
For bovine samples, Brucella DNA was detected in 46 (21.49 %) vaginal swab, 21 (9.8 %) milk, and 5 (2.3 %) blood samples (Table 2). For ovine samples, Brucella DNA was detected in 19 (11.58 %) vaginal swab, 12 (7.3 %) milk, and 4 (2.43 %) blood samples (Table 2). The rt-PCR melt curve data identified only one peak with a melting temperature of 81.5 °C for all positive samples for Brucella spp. Based on species-specific rt-PCR assays, all bovine and ovine positive samples were due to the presence of B. abortus DNA (Tm = 83.5 °C). The amplicons of these positive samples were found to be 100 % identical to the B. abortus gene in the database. Only one ewe was found to be positive for B. melitensis (Tm = 78.5 °C). The amplicon of this positive sample was 100 % identical to the target gene.
Brucella prevalence results
The apparent individual animal prevalence of brucellosis in cattle (21.5 %) was significantly higher than in sheep (12.8 %) (Z-test: P = 0.014) (Table 3). Twelve of 20 dairy herds had at least one positive cow for Brucella spp., resulting in a herd prevalence of 60 %. Compared with sheep, an overall 17.3 % herd level prevalence of brucellosis was found (Z-test: P < 0.0001) (Table 3).
The apparent prevalence of brucellosis on both individual animal and herd levels were adjusted to the test sensitivities (Se (RBT × ELISA × rt−PCR) = 0.9 × 0.99 × 0.99 = 0.882) and specificities (Sp = 1−(1−Sp RBT) × (1−Sp ELISA) × (1−Sp rt−PCR) = 1−(1–0.75) × (1−0.98) × (1−0.99) = 0.99). In cattle, the estimated overall animal population-level TP was 23.5 %, against 13.53 % in ewes (Table 3). The average number of tested animals in each herd of cattle was 10, while for sheep, the number was shown to be between 1 and 5 animals according to the herd sizes. Therefore, the HSp was 0.9 for cattle and 0.95 for sheep. For cattle, the HSe was 0.999, whereas for sheep was 0.613. The THP of brucellosis was 55.6 % for cattle and 21.8 % for sheep (Table 3).
Univariable analysis of related risk factors
For cattle, the chi-square univariable analysis revealed five variables with P values ≤0.05 (Table 1-S). All these significant variables were considered as risk factors of Brucella infection. For sheep, only two variables (age range and history of abortion) were significantly associated with brucellosis at the individual animal level (P < 0.05) (Table 1-S).
The total number of herds for all the study regions and the herd level risk factors assumed to be associated with brucellosis in cattle and sheep are presented in Table 2-S. All significant and no significant variables (with P < 0.2) were then entered to the multivariable logistic analysis.
Multivariable logistic regression analysis
The Hosmer–Lemeshow goodness-of-fit test showed adequate fit for both the cattle (P = 0.996) and the sheep (P = 0.907) models. From the final model for cattle (Table 4), it can be seen that the odds of brucellosis were significantly higher in cows older than 5 years as compared to cows between 3 and 5 years, with an OR of 26. The odds of brucellosis positivity were significantly higher in homebred cows compared to purchased cows with an OR of 139. For sheep between 2 and 3 years old, the odds of brucellosis positivity were 15.62 times higher compared with those that are 1 year of age. In addition, ewes that had aborted a fetus during the previous year had an OR of being Brucella positive that was 62 times higher than those that had not aborted.
At the herd level, risk factors associated for Brucella positivity in the multivariable logistic regression analysis were as follows: abortion and herd composition for cattle against herd composition, mortality, and hygiene/floor type for sheep. The Hosmer–Lemeshow goodness of fit test showed that the two models fit well the data for the cattle (P = 0.994) and sheep (P = 0.645) herds (Table 5).
Discussion
In the routine diagnosis of brucellosis, which is connected with monitoring investigations of the disease in animals, different kinds of methods are used. In our study, the RBT was chosen for animals screening because is sensitive, cheap, and rapid (Lolli et al. 2016). However, it is characterized by its low specificity. Then, the RBT was associated with i-ELISA in a parallel interpretation scheme for this study. The combination of the two serological tests is expected (i) to reduce the occurrence of misclassification and (ii) to improve the testing sensitivity by allowing the detection of the antibodies produced during acute and chronic infections (Musallam et al. 2015). However, it should not be forgotten that if there is suspicion of infection or shedding of Brucella spp. despite the absence of serological response, the test for pathogen detection (rt-PCR) should be performed. Indeed, a highly significant correlation was found between the amounts of DNA (Ct values) obtained by using the rt-PCR and the reaction intensity produced by i-ELISA. This finding can be explained by the invasion of the genital tracts of animals by Brucella organisms, which is known to produce a large and persistent rise of antibodies (Abdalla and Hamid 2012). In contrast, Brucella DNA was detected by means rt-PCR in vaginal secretion and/or milk of some seronegative ewes. This could be due to the delay between infection and seroconversion. On the other hand, when the disease becomes chronic, the antibody titer may drop to undetectable levels (Tittarelli et al. 2007). Furthermore, there was no dominant route of bacterial shedding, which is in agreement with data recently obtained for bovine and ovine herds (Díaz Aparicio 2013; Wareth et al. 2014). Thus, vaginal and milk routes need to be investigated to select Brucella shedders in ruminant herds. The low rate of Brucella in blood samples can be explained by the low bacterial load especially in cases of chronic and focal brucellosis (Mangalgi and Sajjan 2014).
At the individual level, our results concerning both bovine and ovine brucellosis are comparable to values of other investigations released in Africa (Bertu et al. 2010; Muma et al. 2013) and in other continents (Muflihanah et al. 2013). At the herd level, our AP values are similar to those observed among bovine and ovine herds in Nigeria (Mai et al. 2013), Egypt (Wareth et al. 2014), and Jordan (Musallam et al. 2015). The adjustment of prevalence according to test performance gave TP values of 23.5 % in cattle and 13.53 % in sheep. These TP values were higher than those obtained in other studies conducted in Tunisia (Zaibet et al. 2009) and other African countries (Boukary et al. 2013; Teklue et al. 2013). In fact, all these studies were based only on serological tests for the diagnosis of brucellosis, and therefore, new infected animals and seronegative shedders were not considered. In addition, the imperfection, the choice of sampling strategy, the changing of techniques used for brucellosis screening between studies are important factors that contribute to the variability of results among studies (Rajala et al. 2016; Lolli et al. 2016). The signaled high prevalence in the other works might be due to the lack of specificity of used tests (Mai et al. 2013).
The fact that the risk of brucellosis transmission in animals at the population and herd level varied significantly depending on the strata is in agreement with the findings of several authors who demonstrated variations in the prevalence of brucellosis related to the production systems (Chimana et al. 2010; Boukary et al. 2013). In cattle, we found that the risk of brucellosis positivity was higher in urban and peri-urban areas compared to rural areas. This could be explained by the fact that most breeders practicing the intensive dairy farming are localized in urban and peri-urban areas where there is a high demand for milk (Jaouad 2004). On the contrary, the risk of infection with Brucella in sheep was equal in different types of areas, where keeping sheep is a way of saving money. The observed relationships between Brucella status and age and abortion were consistent with what is generally known about the biology of the infection (Boukary et al. 2013; Anka et al. 2014). Similarly, the observed relationship between Brucella positivity and vaccination is consistent with what has generally been observed (Musallam et al. 2015). Indeed, our results corroborate those of Zaibet et al. (2009), who observed that vaccination campaigns against the brucellosis for ewes were not very effective, and many efforts should be consolidated in order to reach an acceptable coverage that can protect animals in Tunisia. Considering origin, this study showed an evidence of association between positivity for Brucella and homebred cows, which confirms the role of vertical transmission in Brucella propagation (Muflihanah et al. 2013).
As expected, a number of herd/flock characteristics were associated with the likelihood of a herd/flock being positive. It is reasonable that a larger herd size increases the risk of infection by increasing the contact rate between susceptible and infected animals (Boukary et al. 2013; Musallam et al. 2015). Larger herds might be expected to be associated with intensive management practices that are typically more difficult to control and allow for closer contact between animals and their environment which increase the potential for exposure to infectious excretions (Teklue et al. 2013). Similarly, our findings showed that herding of ruminant species together, which is specially the normal activity of the sheep breeders in Tunisia, is an important factor to increase transmission of brucellosis between different animal species (Matope et al. 2010; Ducrotoy et al. 2014). This finding may explain the infection of studied ewes mainly by B. abortus.
The remaining factors highlight the importance of management and hygiene practices to mitigate the risk of introduction and/or persistence of Brucella spp. in a herd. Specifically, the implementation of biosecurity and quarantine measures (Ibrahim et al. 2010) and appropriate management of cases of abortion (Anka et al. 2014) could make a significant contribution to any control strategy for ruminant brucellosis. In addition to abortion, exposed herds to brucellosis showed high mortality rates of animals. The reduction of these rates in ruminant herds requires the fight against brucellosis through different practices such as the use of artificial insemination, the choice of nutritious food, and the installation of obligatory prophylactic measures. These practices could contribute to keep healthy immune status of the herds and may also give an indication of better herd management.
Brucellosis is endemic at high levels in domestic ruminant species in Tunisia. The infection is heterogeneously distributed, with some farms at high risk as a result of practices such as exchanging rams for service and introducing new animals without quarantine. Mass vaccination of sheep, in addition to the adoption of hygiene and biosecurity practices, is recommended as a control strategy in Tunisia.
References
Abdalla, A., Hamid, M.E., 2012. Comparaison of conventional and nonconventional techniques for diagnosis of bovine brucellosis in Sudan. Tropical Animal and Health Production 44, 1151–1155.
Akkari, H., Jebali, J., Gharbi, M., Mhadhbi, M., Awadi, S., Darghouth, M.A., 2013. Epidemiological study of sympatric Haemonchus species and genetic characterization of Haemonchus contortus in domestic ruminants in Tunisia. Veterinary Parasitology, 193, 118–125.
Anka, M.S., Hassan, L., Khairani-Bejo, S., Zainal, M.A., Mohamad, R.B., Salleh, A., Adzhar, A., 2014. A case–control study of risk factors for bovine brucellosis seropositivity in Peninsular Malaysia. PLoS One, 9, e108673.
Barkallah, M., Gharbi, Y., Hmani, M., Mallek, Z., Gautier, M., Gdoura, R., Fendri, I., 2016. Locked Nucleic Acid Probe-Based Real-Time PCR for the diagnosis of Listeria monocytogenes in ruminants. Molecular and Cellular Probes, doi:10.1016/j.mcp.2016.02.010.
Bertu W.J., Ajogi, I., Bale, J.O.O., Kwaga, J.K.P., Ocholi, R.A., 2010. Sero-epidemiology of brucellosis in small ruminants in Plateau State, Nigeria. African Journal of Microbiology Research, 4(19), 1935–1938.
Boukary A.R., Saegerman, C., Abatih, E., Fretin, D., Alambedji Bada, R., Deken, R.D., Harouna, H.A., Yenikoye, A., Thys, E., 2013. Seroprevalence and potential risk factors for Brucella spp. Infection in traditional cattle, sheep and goats reared in urban, periurban and rural areas of niger. PLoS One 8, e83175.
Chakroun, M., Bouzouaia, N., 2007. Brucellosis: a topical zoonosis. Revue Tunisienne d’Infectiologie, 1, 1–10.
Chimana, H.M., Muma, J.B., Samui, K.L., Hangombe, B.M., Munyeme, M., Matope, G., Phiri, A.M., Godfroid, J., Skjerve, E., Tryland, M., 2010. A comparative study of the seroprevalence of brucellosis in commercial and small-scale mixed dairy-beef cattle enterprises of Lusaka province and Chibombo district, Zambia. Tropical Animal Health and Production, 42(7), 1541–1545.
Díaz Aparicio, E., 2013. Epidemiology of brucellosis in domestic animals caused by Brucella melitensis, Brucella suis and Brucella abortus. Revue Scientifique et technique, 32(1), 53–60.
Ducrotoy, M.J., Bertu, W.J., Ocholi, R.A., Gusi, A.M., Bryssinckx, W., Welburn, S., Moriyon, I., 2014. Brucellosis asan emerging threat in developing economies: lessons from Nigeria. PLoS Neglected Tropical Diseases, 8, e3008.
Hinic, V., Brodard, I., Thomann, A., Cvetnić, Z., Makaya, P.V., Frey, J., Abril, C., 2008. Novel identification and differentiation of Brucella melitensis, B. abortus, B. suis, B. ovis, B. canis, and B. neotomae suitable for both conventional and real-time PCR systems. Journal of Microbiological Methods, 75(2), 375–378.
Ibrahim, N., Belihu, K., Lobago, F., Bekana, M., 2010. Sero-prevalence of bovine brucellosis and risk factors in Jimma zone of Oromia region, South-Western Ethiopia. Tropical Animal and Health Production, 42, 141–144.
Jaouad, M., 2004. Dynamique des cheptels bovins en Tunisie et contraintes alimentaires et fourragères. In : Ferchichi A. (comp.), Ferchichi A. (collab.). Réhabilitation des pâturages et des parcours en milieux méditerranéens. Zaragoza : CIHEAM. p 421–424.
Lolli, C., Marenzoni, M.L., Strona, P., Lappo, P.G., Etiang, P., Diverio, S., 2016. Infections and risk factors for livestock with species of Anaplasma, Babesia and Brucella under semi-nomadic rearing in Karamoja Region, Uganda. Tropical Animal Health and Production, 48(3), 603–611.
Mai, H.M., Irons, P.C., Kabir, J., Thompson, P.N., 2013. Herd-level risk factors for Campylobacter fetus infection, Brucella seropositivity and within-herd seroprevalence of brucellosis in cattle in northern Nigeria. Preventive Veterinary Medicine, 111, 256–267.
Mangalgi, S., Sajjan, A., 2014. Comparison of three blood culture techniques in the diagnosis of human brucellosis. Journal of Laboratory Physicians, 6, 14–27.
Matope, G., Bhebhe, E., Muma, J.B., Lund, A., Skjerve, E., 2010. Herd-level factors for Brucella seropositivity in cattle reared in smallholder dairy farms of Zimbabwe. Preventive Veterinary Medicine, 94, 213–221.
Megid J., Mathias L.A., Robles C.A., 2010. Clinical manifestations of brucellosis in domestic animals and humans. Open Veterinary Science Journal, 4, 119–126.
Muflihanah, H., Hatta, M., Rood, E., Scheelbeek, P., Abdoel, T.H., Smits, H.L., 2013. Brucellosis seroprevalence in Bali cattle with reproductive failure in South Sulawesi and Brucella abortus biovar 1 genotypes in the Eastern Indonesian archipelago. BMC Veterinary Research, 9, 233.
Muma, J.B., Syakalima, M., Munyeme, M., Zulu, V.C., Simuunza, M., Kurata, M., 2013. Bovine Tuberculosis and Brucellosis in Traditionally Managed Livestock in Selected Districts of Southern Province of Zambia. Veterinary Medicine International, doi: 10.1155/2013/730367.
Musallam, I.I., Abo-Shehada, M., Omar, M., Guitian, J., 2015. Cross-sectional study of brucellosis in Jordan: Prevalence, risk factors and spatial distribution in small ruminants and cattle. Preventive Veterinary Medicine, 118(4), 387–396
Office International des Epizooties (OIE) (2009). Chapter 2.4.3. Bovine Brucellosis. In: Manual of Diagnostic Tests and vaccines for Terrestial Animals. OIE, Paris. pp 1–35.
Rajala, EL., Grahn, C., Ljung, I., Sattorov, N., Boqvist, S., Magnusson, U., 2016. Prevalence and risk factors for Brucella seropositivity among sheep and goats in a peri-urban region of Tajikistan. Tropical Animal Health and Production, 48(3), 553–558.
Rogan, W.J., Gladen, B., 1978. Estimating prevalence from the results of a screening test. American Journal of Epidemiology, 107, 71–76.
Teklue, T., Tolosa, T., Tuli, G., Beyene, B., Hailu, B., 2013. Sero-prevalence and risk factors study of brucellosis in small ruminants in Southern Zone of Tigray Region, Northern Ethiopia. Tropical Animal Health and Production, 45(8), 1809–1815.
Thrusfield, M., 2005. Surveys. In Thrusfield M (ed.) Veterinary epidemiology. Blackwell, Oxford: 228–242.
Tittarelli M., Di Ventura M., De Massis F., Petrini A., Giovannini A., Nannini D., Caporale V. 2007. Kinetics of the antibody response in ewes experimentally infected with Brucella melitensis biovar 3. Veterinaria Italiana, 40(2), 5–10.
Wareth G., Hikal A., Refai M., Melzer F., Roesler U., Neubauer H., 2014. Animal brucellosis in Egypt. Journal of Infection in Developing Countries, 8(11), 1365–1373.
Xavier, M.N., Paixão, T.A., den Hartigh, A.B., Tsolis, R.M., Santos, R.L., 2010. Pathogenesis of Brucella spp. Open Veterinary Science Journal, 4, 109–118.
Zaibet, L., Hammami, S., Jabbar, M., 2009. Sustainability of small ruminant production systems in Tunisia: A health marketing approach. Discussion Paper No. 17. ILRI (International Livestock Research Institute), Nairobi, Kenya. pp 138.
Acknowledgments
This project received financial support from “Ministère de l’enseignement supérieur et de la recherche scientifique” and the ISESCO Organization. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Statement on the welfare of animals
“All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.”
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table 1-S
Potential risk/indicator factors associated with individual animal-level brucellosis positivity among 214 cows and 164 sheep (DOC 58 kb)
Table 2-S
Analysis of potential risk factors associated with herd-level prevalence of brucellosis in ruminants from Tunisia (DOC 114 kb)
Rights and permissions
About this article

Cite this article
Barkallah, M., Gharbi, Y., Zormati, S. et al. A mixed methods study of ruminant brucellosis in central-eastern Tunisia. Trop Anim Health Prod 49, 39–45 (2017). https://doi.org/10.1007/s11250-016-1155-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11250-016-1155-x