
Abstract
The β-glucosidase, which hydrolyzes the β(1–4) glucosidic linkage of disaccharides, oligosaccharides and glucose-substituted molecules, has been used in many biotechnological applications. The current commercial source of β-glucosidase is mainly microbial fermentation. Plants have been developed as bioreactors to produce various kinds of proteins including β-glucosidase because of the potential low cost. Sulfolobus solfataricus is a thermoacidophilic archaeon that can grow optimally at high temperature, around 80 °C, and pH 2–4. We overexpressed the β-glucosidase gene from S. solfataricus in transgenic tobacco via Agrobacteria-mediated transformation. Three transgenic tobacco lines with β-glucosidase gene expression driven by the rbcS promoter were obtained, and the recombinant proteins were accumulated in chloroplasts, endoplasmic reticulum and vacuoles up to 1%, 0.6% and 0.3% of total soluble protein, respectively. By stacking the transgenes via crossing distinct transgenic events, the level of β-glucosidase in plants could further increase. The plant-expressed β-glucosidase had optimal activity at 80 °C and pH 5–6. In addition, the plant-expressed β-glucosidase showed high thermostability; on heat pre-treatment at 80 °C for 2 h, approximately 70% residual activity remained. Furthermore, wind-dried leaf tissues of transgenic plants showed good stability in short-term storage at room temperature, with β-glucosidase activity of about 80% still remaining after 1 week of storage as compared with fresh leaf. Furthermore, we demonstrated the possibility of using the archaebacterial β-glucosidase gene as a reporter in plants based on alternative β-galactosidase activity.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The β-glucosidase can hydrolyze the β(1–4) glycosidic linkage of disaccharides, oligosaccharides and glucose-substituted molecules and also catalyze the synthetic reactions via reverse hydrolytic reaction or transglycosylation (Ahmed et al. 2017). β-Glucosidase has been used in many biotechnological processes. For example, it is among the major hydrolytic enzymes required for the efficient biotransformation of lignocellulosic biomass into biofuels (Park et al. 2016). In addition, in the food industry, β-glucosidase can be used to improve the aroma in wine and release of aromatic compounds in fruit juices (Gueguen et al. 1996). Additionally, β-glucosidase can be used to hydrolyze isoflavone glycosides to increase their absorption by the small intestine in humans (Kim et al. 2012). Furthermore, β-glucosidase has been used for detoxification of cassava during food processing and de-inking of waste paper in the recycling process (Ahmed et al. 2017). On the basis of its synthetic activity, β-glucosidase can be used in the biosynthesis of oligosaccharides and alkyl glycosides, which might have diverse applications such as growth promoters for probiotic bacteria (Diez‐Municio et al. 2014), antimicrobial agents (Otto et al. 1998) and therapeutic agents (Perugino et al. 2004). Currently, the commercial source of β-glucosidases is mainly microbial fermentation (Ahmed et al. 2017).
Transgenic plants have been extensively studied as bioreactors to produce various useful bioactive metabolites or recombinant proteins including enzymes used in biofuel production because of the potential low cost. For instance, transgenic maize overexpressing α-amylase, called Enogen corn enzyme technology, developed by Syngenta, could significantly enhance ethanol production in fermentation (https://www.syngenta-us.com/corn/enogen). In addition, many hydrolytic enzymes from very diverse sources that could be potentially used in the hydrolytic process of lignocellulosic ethanol production have been overexpressed in transgenic plants. For example, endoglucanase and exoglucanase of Acidothermus cellulolyticus have been expressed in tobacco and rice (Jiang et al. 2017). Leaves of transgenic rice can be stored at room temperature for a month and still retain about 80% of the initial activity (Jiang et al. 2017). The β-glucosidase and/or the endoglucanase gene from Thermotoga maritima was expressed in tobacco with the transgene driven by the rbcS or 35S promoter and showed various expression levels when the recombinant proteins were accumulated in distinct sub-cellular compartments such as apoplast, endoplasmic reticulum, chloroplast and vacuole (Jung et al. 2010, 2013; Kim et al. 2010; Nguyen et al. 2015). Furthermore, distinct transit peptides used for targeting the enzyme to chloroplasts might result in various levels of recombinant protein accumulation (Kim et al. 2010). Alternatively, β-glucosidase from the fungus Aspergillus niger, thermophilic bacterium Thermobifida fusca or hyperthermophilic Archaea Pyrococcus furiosus have been expressed in transplastomic tobacco with levels ranging from 2 to 60% of total soluble protein (TSP) without any deleterious phenotypic effect (Petersen and Bock 2011; Yu et al. 2007; Gray et al. 2011; Espinoza-Sánchez et al. 2016; Castiglia et al. 2016). These studies demonstrate that transgenic plants could be used as bioreactors to produce abundant recombinant enzymes at low cost.
Sulfolobus solfataricus, an extreme thermoacidophilic archaeon, grows optimally at 80 °C and pH 2–4 under sufficient metabolizing sulfur environments. It is one of the most widely studied organisms in Archaea, and its genome sequence has been determined and exploited extensively (She et al. 2001). The β-glucosidase (sso3019) from S. solfataricus, a tetramer with molecular weight 60 kD for each subunit, has been purified to homogeneity and overexpressed in Escherichia coli, and the biochemical properties have been extensively characterized (Moracci et al. 1995; Aguilar et al. 1997). The β-glucosidases from S. solfataricus, including β-galactosidase and xylanase activities, show broad substrate specificity (Moracci et al. 1995; D'Auria et al. 1999b). In addition, the crystal structure of β-glucosidase from S. solfataricus showed resilience as a key factor in thermostability (Aguilar et al. 1997). Alcohol, particularly 1-butanol, has been found to enhance enzymatic activity by modifying the protein microenvironment and resulting in a more flexible structure (D'Auria et al. 1999a). A large ion pair network on the surface stabilizes β-glucosidase, and its stability is strongly affected by pH 10, which is correlated with the perturbed ionic interactions as compared with the protein at neutral pH (D'Auria et al. 1998).
Previously, a β-glucosidase (lacS) gene derived from the archaeon S. solfataricus MT-4 strain were overexpressed in the cytoplasm of transgenic tobacco, which could result in the autohydrolysis of plant polysaccharides into free sugar in the plant extracts (Montalvo-Rodriguez et al. 2000). The plant-expressed recombinant proteins in distinct subcellular compartments had a profound effect on the accumulation (Wei et al. 2004; Jiang et al. 2011). In the present study, we used transgenic tobacco overexpressing archaeal hyperthermostable glycosyl hydrolase (sso3019) from the S. solfataricus P2 strain accumulated in three distinct cellular compartments—chloroplasts, endoplasmic reticulum (ER) and vacuole—to test the feasibility of using plants as a bioreactor for production of this recombinant enzyme. Plant-expressed β-glucosidase with accumulation in chloroplasts could reach up to 1% of total soluble protein (TSP) and showed higher activity than that expressed in ER and vacuole. We also explored additional β-galactosidase and β-glucuronidase activity for this archaebacterial multi-functional enzyme in plants. Our results demonstrated that plant-expressed β-glucosidase conferred high β-galactosidase activity but no β-glucuronidase activity. The β-galactosidase is commonly used as a reporter system in mammalian cells, insects and yeast (Naylor 1999), but not plants because of high endogenous activity. However, by heat-pretreatment of plant tissues prior to enzymatic reaction or increasing the reaction temperature, the endogenous β-galactosidase activity could be eliminated. Therefore, from the β-galactosidase activity findings, in this study, we also demonstrated that the archaebacterial β-glucosidase (sso3019) might provide an alternative source of reporter gene in plants.
Material and method
Cloning of β-glucosidase gene from S. solfataricus
The archaebaceria S. solfataricus P2 (ATCC 35,092) was obtained from the BioSource Collection and Research Center (BCRC), Taiwan. Bacterial DNA was isolated as described (Hoffman and Winston 1987) with slight modification. The bacterial genome sequence (accession no. AE006641) was downloaded from GenBank. The β-glucosidase (sso3019) gene was PCR-amplified from the total DNA of S. solfataricus P2 with the specific primer pair 3019-F and 3019-R (Supplementary Table 1), which also resulted in the c-Myc tag sequence in the 5′end of DNA fragments. Then the DNA fragments were cloned into the pT&A cloning vector (Real Biotech Corp., Taiwan) and checked with restriction enzyme digestion as well as further Sanger sequencing to obtain the pBglb plasmid.
Construction of plant nuclear expression vectors
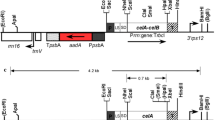
Archaebacterial β-glucosidase was accumulated in three distinct subcellular compartments of tobacco cells—chloroplasts, ER and vacuoles. AFVY located in the C-terminal of the vacuolar storage glycoprotein phaseolin was a sorting signal to direct the protein to the vacuole (Frigerio et al. 2001). To accumulate the β-glucosidase in the vacuole, the DNA fragment encoding AFVY was annealed by the 3019AFVY-F and 3019AFVY-R primer pair (Supplementary Table 1). Then, the annealed DNA fragment was ligated with the 1.5-kb DNA fragment from the NotI/SacI double-digested pBglb plasmid and the recombinant DNA fragment was cloned into the NcoI/SacI site of pImpactVector 1.2 to obtain the pBgl-Va plasmid. To accumulate the protein in ER and chloroplast, the 1.5-kb DNA fragment from the NcoI/NotI-digested pBglb plasmid was cloned into pImpactVector 1.3 and pImpactVector 1.4, respectively (Tu et al. 2014), to obtain pBgl-Er and pBgl-Cp vectors. The 3.5-, 3.5-, and 3.7-kb DNA fragments conferring sso3019 gene expression cassettes were AscI/PacI double-digested from pBgl-Cp, pBgl-Er and pBgl-Va plasmids, respectively, then cloned into the AscI/PacI site of pBINplus binary vectors to obtain the plant nuclear expression binary vectors pBin-Cp, pBin-Er and pBin-Va, respectively (Fig. 1).

Schematic representation of plant nuclear expression vectors and bacterial expression vector. The pBin-Cp, pBin-Er and pBin-Va are plant nuclear expression binary vectors with their backbone derived from pBINplus. sso3019, β-glucosidase gene of S. solfataricus P2. PNos, Nos (nopaline synthase gene) promoter; TNos, Nos terminator; NptII, Kanamycin resistance gene; PRbcS, RbcS (ribulose 1, 5-bisphosphate carboxylase/oxygenase small subunit) gene promoter; TRbcS, RbcS terminator; Sp, signal peptide of secretory pathway; Tp, chloroplast transit peptide; KDEL, ER retention signal; AFVY, vacuole sorting signal; c-Myc, c-Myc tag; RB, right border of T-DNA; LB, left border of T-DNA. The arrows indicate the gene-specific primer pair used for PCR analysis. The pET-Bglb is a bacterial expression vector with the backbone plasmid pET24a. PT7, T7 promoter; Olac, Lac operator; His6, six His tag; TT7, T7 terminator
Plant transformation, selection and screening
Agrobacteria-mediated leaf disc transformation of tobacco (Nicotiana tabacum L. cv. Petit Havana) was performed as described (Chang et al. 2007) with slight modification. In brief, leaf discs infected with A. tumefaciens strain GV3101 carrying individual pBin-Cp, pBin-Er or pBin-Va binary vectors were incubated on TSM medium [0.44% Murashige and Skoog (MS) medium with vitamin, 3% sucrose, 0.1 mg/L naphthalene acetic acid (NAA), 1.0 mg/L benzyladenine (BA) and 0.3% phytagel, pH 5.7] containing both timentin (250 mg/L) and kanamycin (100 mg/L) for selection. Regenerated shoots were transferred to TRM medium (0.44% MS with vitamin, 3% sucrose, 0.1 mg/L NAA, and 0.3% phytagel, pH 5.7) containing kanamycin (100 mg/L) for rooting. T0 plants were self-crossed, and T1 progenies were selected in MS medium containing kanamycin to check for Mendelian segregation. The transgenic lines with the segregation ratio of approximately 3–1 between survived (green) versus dead (albino) seedlings were taken as single copy transgene insertion in the genome. In addition, β-glucosidase activity assay was performed as described in the following section to screen T1 progenies. The selected transgenic plants with relatively high enzymatic activity were used to further confirm the transgene integration by PCR analysis with Bglb-F forward and Bglb-R reverse primers (Supplementary Table 1). The selected T1 transgenic lines that were kanamycin-resistant, PCR-positive and with high enzymatic activities were further self-crossed, and then all progenies resistant to kanamycin were selected for putative homozygous T2 plants.
Bacterial expression and protein purification
To express the β-glucosidase gene of S. solfataricus P2 in E. coli, the 1496-bp DNA fragments BamHI/NotI-digested from pBin-Va plasmid (see above) were cloned into the BamHI/NotI site of the bacterial expression vector pET24a to obtain the expression construct pET-Bglb, which could generate a C-terminal histidine-tagged fusion protein in E. coli BL21 (DE3) (Fig. 1). The transformed BL21 cells were grown at 37 °C to OD600 approximately 0.5, then the recombinant β-glucosidase proteins were induced with 1 mM IPTG at 37 °C for 5 h. The recombinant proteins were purified under a native condition by using a Ni-nitrilo-tri-acetic acid (Ni-NTA) agarose resin according to the manufacturer’s protocol (Qiagen, Germany). The protein concentration was determined by using the Bio-Rad Protein Assay Kit (Bio-Rad, Hercules, CA).
β-Glucosidase activity assay
Approximately 150 mg leaf tissue of T1 transgenic tobacco at various developmental stages as indicated was ground into powder in liquid nitrogen, and 300 µl extraction buffer (50 mM Tris–HCl [pH 8.0], 3 mM PMSF, 1% DMSO, 1% 2-mercaptoethanol, 0.2% Triton X-100, 0.3 M NaCl, 0.3 M sucrose, 10 mM ascorbic acid, 0.1% proteinase inhibitor) was added and further homogenized. After centrifugation, the soluble fraction was heat pre-treated or not at 65 °C for 30 min, then centrifuged at 10,000×g for 20 min at 4 °C to remove the insoluble materials. The supernatant was stored at − 80 °C. In addition, aliquots of plant extracts without heat pre-treatment were used to measure the concentration of TSP by using bovine serum albumin as a standard as described above.
The enzymatic activity of tobacco-expressed β-glucosidase was performed in 150 μl reaction mixture containing the indicated amount of protein extracts, 5 mM p-nitrophenyl-β-D-glucoside (pNPG) substrate and 50 mM sodium phosphate, pH 6. The reaction mixture was incubated at 65 °C for 30 min or as indicated. The enzymatic reaction was terminated by the addition of 150 μl of 0.4 M glycine–NaOH. Enzymatic activity was determined by measuring the released p-nitrophenol at OD405nm. In parallel, the reaction mixture without protein extracts was a blank control. The reaction mixture with different concentrations (25–800 µM) of p-nitrophenol was used to establish a standard curve for the quantitation of unknown sample.
SDS-PAGE and immunoblot analysis
Different amounts of bacteria-expressed proteins and plant protein extracts from transgenic tobacco lines or untransformed plant as indicated were separated on 10% SDS–polyacrylamide gels, then stained with Coomassie brilliant blue, de-stained with 15% methanol plus 7.5% acetic acid, and quantified by using ImageJ. Alternatively, protein gels were transferred to PVDF membranes. Blocking was performed in NET buffer (0.25% gelatin, 50 mM Tris–HCl pH 7.5, 50 mM NaCl, 5 mM EDTA and 0.05% Tween-20). Blots were incubated with a monoclonal antibody against c-Myc (1–1000 dilution) (Hopegen Biotechnology, Taiwan) or polyclonal antibody against His6 (1–1000 dilution) (Amersham, USA) in NET buffer at room temperature for 1 h, washed three times with NET buffer, and reactive bands were detected with horseradish peroxidase-conjugated secondary antibody (Amersham, USA).
Effect of temperature and pH on enzymatic activity
To test the effect of temperature on enzymatic activity for tobacco-expressed β-glucosidase, leaf extracts isolated from the transgenic pBin-Cp line were incubated with pNPG substrate at 20–100 °C at the indicated interval for 10 min before measuring enzymatic activity. To test the effect of pH on the enzymatic activity for tobacco-expressed β-glucosidase, leaf protein extracts were reacted with pNPG substrate at 80 °C for 10 min in buffer with pH 1–10. The following buffers at a final concentration of 50 mM were used for testing the effect of pH on activity: KCl–HCl pH 1 and 2; Na-citrate pH 3–6; Na-acetate pH 4–5.5; K-phosphate pH 6, 7 and 8; Na-tetraborate pH 9 and 10.
Thermostability and storage stability
To test the thermostability of tobacco-expressed β-glucosidase, leaf extracts of the transgenic pBin-Cp line were heat pre-treated under 70, 80 and 90 °C for 0–120 min with a 30-min interval, then the residual enzymatic activity of each sample was assayed for 10 min by adding pNPG as a substrate as described above. To investigate the storage stability of tobacco-expressed β-glucosidase in isolated liquid extracts, protein extracts were stored at − 80 °C, − 20 °C, 4 °C or room temperature for 3–30 days at the indicated interval, and then the residual level of enzymatic activity was measured. To investigate the storage stability of tobacco-expressed β-glucosidase in preserved leaf tissues, mature leaves from the 4-month-old transgenic pBin-Cp line were harvested, wind-dried and stored for 1–5 weeks at weekly intervals and at room temperature. Protein extracts were isolated, the concentration of TSP was measured, and the residual level of enzymatic activity was assayed and compared to that of initially harvested leaf tissues.
Measurement of glucose release from cellobiose
Approximately 4.3 g leaf tissue of T2 transgenic tobacco and wild type plant was ground into powder in liquid nitrogen, and 5 ml extraction buffer (50 mM sodium acetate [pH 5.0], 1 mM EDTA, 10 mM ascorbic acid, and 12 mM β-mercaptoethanol) was added and further homogenized. After centrifugation, the soluble fraction was heat pre-treated or not at 65 °C for 30 min, then centrifuged at 10,000×g for 20 min at 4 °C to remove the insoluble materials. The protein was precipitated by slowly adding ammonium sulfate to supernatant until saturation (60%). The precipitated protein was collected after centrifugation at 18,000×g for 25 min at 4 °C. In addition, aliquots of plant extracts prior heat pre-treatment and purified protein were used to measure the protein concentration by using bovine serum albumin as a standard. The indicated concentration of cellobiose (Acros Organics, USA) as substrate was mixed with protein (or without protein as a blank control) which was heat-denatured at 100 °C for 10 min or not. Then the samples were incubated at 70 °C for indicated time to release glucose, and followed by boiled the samples at 100 °C for 10 min to terminate reaction. The released glucose was measured by using a glucose (GO) assay kit (Sigma, USA) according to the manufacturer’s procedure. One unit of enzyme activity was defined as the amount of enzyme required to release 1 μmol of glucose per min.
Assay of β-galactosidase and β-glucuronidase activity
The β-galactosidase and β-glucuronidase activity of plant protein extracts isolated from transgenic tobacco or the wild type were tested by using the chemiluminescent substrates Galacton-star and Glucuron (Tropix, USA), respectively, according to the manufacturer’s procedure. In brief, for measuring β-galactosidase activity, Galacto-Star substrates were incubated with or without (negative control) different amounts of plant protein extracts under the indicated temperature for 10 min before measuring light emission by using the Minilumat LB 9506 Luminometer (Berthold Technologies, Germany). For measuring β-glucuronidase activity, Glucuron substrates were incubated with different amounts of plant protein extracts under the indicated temperature for 10 min, and then the light emission accelerator was added before measurement by luminometry.
Zymography analysis
For zymography analysis of plant-expressed recombinant enzymes with β-galactosidase activity, equal amounts of protein extracts with or without heat pre-treatment at 70, 80 and 90 °C for 2 h were separated by 8% native PAGE with 100 V at 4 °C for 90 min. The gel was immersed in 50 mM Na-phosphate (pH 6.0) buffer containing 0.2% X-gal at 70 °C for 45 min to develop indigo bands. The reactive bands showing β-galactosidase activity were quantified by using ImageJ.
Histochemical staining
The 3-week-old seedlings from transgenic or wild-type tobacco underwent histochemical X-gal staining for 4 h and GUS staining overnight, respectively, under 60 °C or room temperature as described (Huang et al. 2006; Jefferson et al. 1987) with slight modification.
Statistic method
The assays of enzymatic activity were performed in triplicate with the exception as indicated, and the data was presented as mean ± standard deviation (SD). In addition, the Dunnett's test was applied to statistically analyze the root length of seedlings.
Results
Construction of plant nuclear expression vectors and plant transformation
In the present study, we constructed three plant nuclear gene expression cassettes with β-glucosidase (sso3019) gene from archaebacteria S. solfataricus P2 based on the backbone plasmid of pBINplus (Fig. 1). Binary vectors of pBin-Cp, pBin-Er and pBin-Va expressing sso3019 gene were driven by the leaf-specific rbcS promoter. In addition, to accumulate plant-expressed recombinant proteins in chloroplasts, for the pBin-Cp construct, the rbcS transit peptide was 5′ in frame-fused with sso3019. For protein synthesis via a secretory pathway, the 5′ end of sso3019 was fused with a secretory signal for the pBin-Er and pBin-Va constructs. Also, the ER-retention signal KDEL was fused to the 3′ end of sso3019 for pBin-Er, so the expressed protein accumulated in the ER. The vacuolar sorting signal AFVY was fused to the 3′ end of sso3019 in the pBin-Va construct, so the plant-expressed recombinant proteins could accumulate in the vacuole (Fig. 1). The signal sequences used in this study have been demonstrated to be able to successfully direct the heterologous proteins to correct subcompartments of plant cell (Frigerio et al. 2001; Tu et al. 2014). Furthermore, to facilitate the detection of recombinant protein, c-Myc tag was fused to the N-terminal of β-glucosidase in the expression cassettes (Fig. 1).
In total, 54, 44 and 32 kanamycin-resistant putative T0 transgenic events were selected from pBin-Cp–, pBin-Er– and pBin-Va–transformed tobacco lines, respectively. T0 transgenic lines were further self-crossed, and the segregation ratio of T1 progenies was analyzed by germinating seeds in 1/2 MS medium containing kanamycin. Approximately 24 pBin-Cp, 12 pBin-Er and 6 pBin-Va transgenic events followed Mendelian segregation, and were possible single transgene integration events in the genome (data not shown).
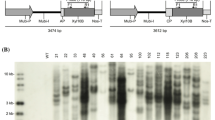
The integration of sso3019 into the nuclear genome of selected transgenic T1 tobacco events was further checked by PCR assay with a primer pair from sso3019; 749 bp was the expected size. Overall, 21 of 22 T1 transgenic events from the pBin-Cp expression line, all 11 T1 transgenic events from the pBin-Er line, and 12 of 13 T1 transgenic events from the pBin-Va line were PCR-positive and might contain the transgene in the chromosome (Supplementary Fig. 1).
β-Glucosidase activity among transgenic plants
Previous studies have shown that plant-expressed recombinant proteins in distinct subcellular compartments might have a profound effect on the accumulation (Wei et al. 2004; Jiang et al. 2011). In this study, the level of β-glucosidase might be distinct among different transgenic lines with recombinant proteins targeted to chloroplasts, ER and vacuoles. In addition, the positional effects might also result in a differential level of transgene expression among events even in the same transgenic line (Chang et al. 2007; Lu et al. 2011). On the basis of the conversion of the pNPG substrate into glucose and p-nitrophenol, β-glucosidase activity was measured by the appearance of p-nitrophenol at OD405nm (Nguyen et al. 2015). The activity assay was carried out in T1 progenies of 3-week-old seedlings to estimate the level of β-glucosidase in transgenic tobacco. At least 43 of 50 transgenic events in pBin-Cp line showed higher enzymatic activity than in transgenic seedlings carrying only empty vector and untransformed plants (Supplementary Fig. 2a). In particular, transgenic events of Cp34, Cp35, Cp37, Cp42 and Cp54 from pBin-Cp–expressed tobacco showed the highest enzymatic activity (Supplementary Fig. 2a). However, only 13 of 35 transgenic events in pBin-Er showed higher level of enzymatic activity than control plants (Supplementary Fig. 2b). Additionally, transgenic events of Er09, Er32 and Er34 from pBin-Er–expressed tobacco showed the highest enzymatic activity (Supplementary Fig. 2b). At least 21 of 28 transgenic events in pBin-Va showed higher enzymatic activity than control plants (Supplementary Fig. 2c). In addition, transgenic events of Va01, Va11 and Va23 from the pBin-Va–expressed tobacco line showed the highest enzymatic activity (Supplementary Fig. 2c). Overall, transgenic pBin-Cp line conferred higher enzymatic activity than that of pBin-Er or pBin-Va. Therefore, the plant-expressed β-glucosidase from transgenic pBin-Cp line was mainly used in subsequent experiments.
To quantify the level of recombinant β-glucosidase in transgenic plants, recombinant β-glucosidase was also overexpressed in bacteria, and then purified with Ni–NTA resin. Different amounts (150 ng to 1.2 µg) of bacteria-expressed β-glucosidase were used to establish a standard curve for measuring the level of β-glucosidase in 2-month-old transgenic plants via SDS-PAGE, followed by staining and quantification by ImageJ (Fig. 2a). The β-glucosidase could accumulate up to approximately 1%, 0.6% and 0.3% of TSP in transgenic pBin-Cp, pBin-Er and pBin-Va lines, respectively (Fig. 2a). In addition, we measured β-glucosidase activity based on an equal amount of TSP among Cp42, Cp54, ER32, Va11 and wild-type plants. Cp54 conferred the highest activity (Fig. 2b), which is consistent with the high expression of β-glucosidase (Fig. 2a). Because the plant-expressed recombinant β-glucosidase contains a c-Myc tag in the N-terminal and bacteria-expressed β-glucosidase contains a His6 tag in the C-terminal (Fig. 1), the expression of recombinant β-glucosidase in transgenic plants or bacteria was further confirmed by immunoblot analysis with anti-Myc or anti-His6 antibody. With anti-Myc antibody, β-glucosidase was detected in transgenic plants rather than untransformed tobacco or bacteria (Fig. 2c). In contrast, when anti-His6 antibody was used, only bacteria expressed β-glucosidase was detected rather than plants (Fig. 2c). In addition, to further increase the level of β-glucosidase in plants, we performed hybridization between transgenic pBin-Cp34 and pBin-Cp54 events, and their progenies showed increased amount of β-glucosidase up to 1.1% of TSP (Supplementary Fig. 3).

Expression level of recombinant β-glucosidase in transgenic tobacco lines. a Plant protein extracts (equivalent to 110 μg of TSP) isolated from leaves of 8-week-old plants and bacteria-expressed and purified β-glucosidase protein (150, 300, 600 and 1200 ng) were subjected to 10% SDS-PAGE and then stained with Coomassie brilliant blue. The intensity of indicated bands (arrow head) with predicted size about 60 kD was quantified by using ImageJ to estimate the amount of plant-expressed β-glucosidase as shown in the percentage of TSP (bottom). Cp42 and Cp54 are distinct transgenic events in pBin-Cp line. Er32 and Va11 are transgenic events in pBin-Er and pBin-Va lines, respectively. Wt, untransformed tobacco. M, protein marker. b The β-glucosidase activity based on equal amount of protein (equivalent to 2 μg of TSP) isolated from distinct transgenic tobacco lines measured by using pNPG as a substrate at 80 °C for 10 min. Data are mean ± SD (n = 3). c Detection of plant- and bacteria-expressed β-glucosidase by immunoblot analysis of TSP (25 μg) isolated from transgenic and untransformed tobacco plants with anti-c-Myc (right panel) or anti-His6 (left panel) antibody. The indicated amount of purified bacteria-expressed β-glucosidase fusion protein was a control. Wt, untransformed tobacco; Cp34, pBin-Cp34 transgenic event; Cp54, pBin-Cp54 transgenic event; Cp54 × 34, progenies from the cross between Cp34 and Cp54 transgenic events
In the present study, the expression of recombinant β-glucosidase gene was driven by the leaf-specific rbcS promoter (Fig. 1). To investigate the expression of β-glucosidase during leaf development, leaves from the developing phase (top) to senescent phase (bottom) in 4-month-old transgenic pBin-Cp34 plants were used to assay enzymatic activity. The β-glucosidase activity was about two- to threefold higher in mature leaves than developing and senescent leaves (Fig. 3).

Activity of plant-expressed β-glucosidase at different stages of leaf development. The leaves of 4-month-old pBin-Cp54 transgenic plants at different developmental stages starting from the top third leaf as first stage down to the 14th leaf as twelfth stage were used to measure enzymatic activity. Four leaf discs each 1 cm in diameter were punched from the middle of each leaf and used to extract proteins for activity assay. The bars indicate the relative enzymatic activity, with the highest activity at stage 5 set to 100%. Data are mean ± SD (n = 3). The line indicates the relative amount of TSP, with the highest amount of protein at stage 2 set to 100%
Optimal temperature and pH
A previous study overexpressing β-glucosidase of the S. solfataricus strain MT-4 in E. coli showed that the optimal temperature for bacteria-expressed β-glucosidase activity was 80 °C (Moracci et al. 1995). The recombinant protein expressed from different organisms might have distinct properties. In the present study, tobacco-expressed recombinant β-glucosidase remained active from 40 to 100 °C, with optimal temperature at 80 °C (Supplementary Fig. 4a). In contrast, enzymatic activity was extremely low (< 3%) at temperatures below 40 °C. This result is consistent with the previous study (Moracci et al. 1995).
The effect of pH on enzymatic activity was also tested as described in materials and methods. Plant-expressed β-glucosidase had a broad pH range from 4 to 10 (Supplementary Fig. 4b). However, the optimal pH for plant-expressed β-glucosidase was 5.5 with Na-citrate or Na-acetate buffer and 6 with K-phosphate buffer in weak acidic condition (Supplementary Fig. 4b).
Thermostability
To test the thermostability of tobacco-expressed β-glucosidase, leaf protein extract was heat pre-treated at 70, 80 and 90 °C from 30 to 120 min with 30-min intervals before measuring its activity. Heat pre-treatment of plant-expressed β-glucosidase at 90 °C for 30 min sharply decreased enzymatic activity to approximately 53%, which continued to decrease to 37%, 20% and 16% after 1, 1.5 and 2 h treatment, respectively (Supplementary Fig. 5). Heat pre-treatment of plant-expressed enzyme at 80 °C for 30 min resulted in approximately 94% residual activity, gradually reduced to about 86%, 87% and 71% after 0.5, 1 and 2 h treatment, respectively (Supplementary Fig. 5). In contrast, heat pre-treatment of plant-expressed enzyme at 70 °C for up to 2 h had almost no effect on enzymatic activity (Supplementary Fig. 5). Therefore, plant-expressed β-glucosidase conferred high thermostability.
Storage stability
The storage stability of plant-expressed β-glucosidase in crude extracts was tested as described in methods. Almost no loss of enzymatic activity was observed after 1 month of storage at − 80 °C. In contrast, enzymatic activity gradually decreased to 78%, 64% and 55% after 1 month of storage at − 20, 4 °C or room temperature, respectively (Supplementary Fig. 6a). Hence, plant-expressed β-glucosidase in the storage form of liquid crude extracts may be relatively stable.
Previously, the activity of tobacco-expressed β-glucosidase from T. maritima could be preserved in leaves after lyophilization, but decreased by over 70% with drying at room temperature (Jung et al. 2010). In the present study, the storage stability of plant-expressed β-glucosidase in preserved leaf was investigated according to the description in methods. After 1 week of storage at room temperature, the enzymatic activity still retained approximately 88% activity as compared with fresh harvested leaf, but the activity rapidly decreased to about 42% after 2 weeks, then gradually to 25% of initial activity after 5 weeks (Supplementary Fig. 6b). These results suggest that short-term storage (about 1 week) of plant-expressed β-glucosidase in tissue might be feasible.
Plant-expressed β-glucosidase catalyzes the release of glucose from cellobiose
In previous study, conversion of plant extracts enriched for endogenous polysaccharides into free glucose was demonstrated by using plant extracts of transgenic tobacco lines expressing and accumulating β-glucosidase in cytosol (Montalvo-Rodriguez et al. 2000). In this study, commercial cellobiose was used as substrates for tobacco-expressed β-glucosidase, purified from transgenic pBin-Cp54 or wild-type plants as control, and levels of liberated glucose were determined. Glucose release was significantly increased in plant-expressed β-glucosidase as compared with that of wild type. In contrast, extremely low level of glucose was present in reactions when the protein was heat-denatured (Fig. 4a). In addition, the initial activity of plant-expressed β-glucosidase could reach up to 188 units, and with increasing concentration of cellobiose from 0.5 to 16%, the initial reaction rate was slightly decreased up to 14% (Fig. 4b). Our results suggested that plant-expressed β-glucosidase is able to catalyze the hydrolysis of plant saccharide.

Release of glucose by plant-expressed β-glucosidase action on cellobiose. The cellobiose was used as a substrate, and levels of glucose released by plant-expressed β-glucosidase were determined by glucose oxidase assay. The proteins were isolated from pBin-Cp54 (Cp54) transgenic plants and untransformed wild-type (Wt) plants. a The reaction was incubated at 70 °C for 2 h, and pH 5.0 containing 8 μg protein and 5 mM cellobiose as substrate. The protein was either heat-pretreated at 100 °C for 10 min as control (white) or not heat-pretreated (grey). Data are mean ± SD (n = 3). b The effects of cellobiose concentration (0.5–16%, w/v) on the initial reaction rates of β-glucosidase, the reactions were performed at 70 °C for 10 min, and pH 5.0 containing 15 μg protein purified from Cp54 transgenic plants. Data are mean ± SD (n = 3)
Plant-expressed recombinant enzyme conferred β-galactosidase activity but no β-glucuronidase activity
Previous studies have shown that β-glucosidase of S. solfataricus exhibited broad substrate specificity such as β-galactosidase and xylanase activities (Moracci et al. 1995), but no reports of β-glucuronidase activity are available. In the present study, we tested whether plant-expressed β-glucosidase conferred β-galactosidase and β-glucuronidase activity using the chemiluminecent substrates as described in methods. With Galacton-Star used as a substrate, β-galactosidase activity was detected at as low as 10 ng of TSP in transgenic pBin-Cp54, and with increased amount of protein, higher enzymatic activity was detected (Fig. 5a). In addition, the activity of β-galactosidase was further increased with increasing reaction temperature up to 60 °C but decreased with further increased reaction temperature probably because of the instability of substrate (Fig. 5b). In contrast, only an extremely low level of β-galactosidase activity was detectable in the leaves of wild-type plants. Alternatively, plant TSP was heat-pretreated or not at 65 °C for 30 min to get rid of most endogenous plant proteins. Then the thermostability of heat-pretreated plant extracts was investigated by incubating samples under different temperatures (70, 80, 90 °C) for 120 min before separation on 8% PAGE. Zymography analysis was performed with X-gal as substrate at 70 °C for 45 min, and the intensity of bands was quantified by using ImageJ. Previous study showed that the functional archaebacterial enzymes consist of a tetramer with molecular weight about 240 kD (Moracci et al. 1995; Aguilar et al. 1997). Our zymography analysis and gel filtration chromatography revealed β-galactosidase activity of multi-subunits complex from TSP of transgenic pBin-Cp54, but was almost undetectable in wild-type plants (Fig. 5c and Supplementary Fig. 7). In addition, 79% β-galactosidase activity remained when plant-expressed recombinant enzyme isolated from pBin-Cp54 transgenic plant was heat-pretreated at 70 °C for 120 min versus no heat pretreatment (Fig. 5c). In contrast, the enzymatic activity was significantly decreased to 16% on further increasing the pretreatment temperature to 80 °C, and the activity was almost undetectable after 90 °C heat pretreatment. The thermostability of the plant-expressed recombinant enzyme for β-galactosidase activity is less than that of β-glucosidase activity (Supplementary Fig. 5). In addition, histochemical X-gal staining further revealed that β-galactosidase mainly expressed in leaf tissue of transgenic pBin-Cp54 but not wild-type plants, and the activity was higher with the reaction temperature increasing to 60 °C (Fig. 5d). However, when the Glucuron was used as a substrate, β-glucuronidase activity was not detected, even when using 20 μg TSP from leaf tissues of pBin-Cp54 and wild-type plants (data not shown). In addition, histochemical GUS staining also did not reveal any β-gucuronidase activity in leaf tissue of transgenic pBin-Cp54 or wild-type plants (Fig. 5d). Overall, our results demonstrated that plant-expressed β-glucosidase also conferred high β-galactosidase activity but no β-glucuronidase activity. In addition, on the basis of β-galactosidase activity, the archaebacterial multi-functional β-glucosidase might have potential as a reporter in plants.


Detection of the β-galactosidase and β-glucuronidase activity in transgenic plants. Different amounts of protein extracts (equivalent to 0.01, 0.1, 0.5, 1, 2 and 4 μg TSP) with or without heat-pretreatment, from pBin-Cp54 (Cp54) transgenic or untransformed wild-type (Wt) plants were incubated in triplicate with chemiluminecent substrate Glacton-Star at a 40 °C for 10 min or b 30, 40, 50, 60, 70 and 80 °C for 10 min before measuring light emission. Data are mean ± SD (n = 3). c Plant TSP was with or without heat-pretreated at 65 °C for 30 min to get rid of most endogenous plant proteins. Then the heat-pretreated plant extracts were further incubated under 70, 80 or 90 °C for 120 min before separation on 8% native PAGE. Zymography analysis was performed with X-gal as substrate at 70 °C for 45 min, and the intensity of bands was quantified by using ImageJ as shown in the bottom. d Histochemical X-gal staining of 3-week-old seedlings from transgenic (Cp54) or untransformed (Wt) plants was carried out at room temperature or 60 °C for 4 h (top). Histochemical GUS staining of 3-week-old seedlings from transgenic (Cp54) or untransformed (Wt) plants was carried out at room temperature or 60 °C for 20 h (bottom). The ruler is 5 mm
Discussion
The β-glucosidase has great potential to be used in many biotechnological applications such as in biofuels and the food and beverage and pharmaceutical industries. The β-glucosidases of S. solfataricus show broad substrate specificity, which include β-glucosidase, β-galactosidase, β-fucosidase and β-xylosidase activities (Grogan 1991; Nucci et al. 1993; this study). Besides its broad substrate specificity, with high thermostability and good catalytic kinetics, β-glucosidase of S. solfataricus is useful in the industrial hydrolysis of phytochemicals such as oleuropein, ginsenoside and isoflavone glycosides (Briante et al. 2000; Noh et al. 2009; Kim et al. 2012). Despite the current commercial source of β-glucosidases mainly from microbial fermentation (Ahmed et al. 2017), transgenic plants have been developed as bioreactors for alternative production of β-glucosidases because of the potential low cost (Park et al. 2016). In the present study, the β-glucosidase gene from S. solfataricus was overexpressed in the three distinct subcompartments, chloroplasts, ER and vacuoles of transgenic tobacco.
The conversion of cellobiose by β-glucosidases is a critical step to reduce cellobiose inhibition and enhance the efficiency of cellulolytic enzymes in lignocellulosic ethanol production (Ahmed et al. 2017). However, the cost of hydrolytic enzymes is a major limitation in the lignocellulosic biofuel industry (Park et al. 2016). For application in the bioconversion of lignocellulosic biomass and others, many β-glucosidase genes from diverse sources such as fungi, mesophilic bacteria or thermophilic archae have been expressed in plants for cost-effective production of recombinant enzymes. In addition, the accumulation of β-glucosidase in different subcellular compartments of plants could significantly affect their expression. For example, the expression of the β-glucosidase gene from A. niger in five different subcellular compartments such as cytoplasm, cell wall, ER, vacuole and chloroplast of transgenic tobacco, with the vacuole-targeted transgenic plant having highest activity, reached up to 2.3% of TSP (Wei et al. 2004, 2007). In addition, when the β-glucosidase (BglB) gene from Thermotoga maritima was expressed in tobacco and the recombinant enzyme was targeted to the cytosol and chloroplasts, it accumulated up to 4.5% and 5.8% of TSP, respectively (Jung et al. 2010), or even up to 9.3% of TSP when the same gene was driven by the alfalfa rbcSK-1A promoter and the recombinant protein accumulated in chloroplasts (Jung et al. 2013). In another study, BglB of T. maritima driven by the 35S promoter was overexpressed in tobacco and effectively targeted to cytosol or vacuole, with vacuole-targeted transgenic plants having the highest activity (Nguyen et al. 2015). When the β-glucosidase (lacS) gene of S. solfataricus strain MT-4 driven by a CaMV 35S promoter was overexpressed in cytosol of transgenic tobacco, the accumulation of recombinant enzyme could reach up to 0.15% of TSP (Montalvo-Rodriguez et al. 2000). Taken together, many factors could potentially affect the steady state protein level such as transcription, translation, protein targeting, folding, assembly and degradation. Previously, the regulatory 5′ and 3′ non-translated regions of rbcS1 were engineered to drive heterologous expression of various genes in tobacco, and the foreign protein expression could reach up to 10% of TSP (Outchkourov et al. 2003a). In the present study, the same rbcS1 regulatory elements were used to drive the expression of β-glucosidase gene (sso3019) of S. solfataricus strain P2 in tobacco, and subsequently the recombinant proteins were accumulated in chloroplasts, endoplasmic reticulum and vacuoles, respectively. Therefore, we assumed that these three transgenic lines have the same transcription rate. Plant-expressed β-glucosidase with accumulation in chloroplasts could reach up to 1% of TSP, significantly higher than that in the ER and vacuole (Fig. 2 and Supplementary Fig. 2), but far less than previously reported to express GusA in tobacco (Outchkourov et al. 2003a). On the basis of the assumption of no difference in protein translation, folding and assembly, under different subcellular compartments, a wide variety of proteases might be involved in the processing and degradation of proteins (Outchkourov et al. 2003b). Therefore, differentially proteolytic degradation of heterologously expressed β-glucosidase might be one of the major limiting factors for the steady-state protein expression level in specific compartments in plants.
By stacking the transgenes via crossing among different transgenic events in the pBin-Cp and pBin-Er lines, we could further increase the level of β-glucosidase in plants (Supplementary Fig. 3). Therefore, the β-glucosidase level could be further improved by stacking the transgenes via crossing among different transgenic lines or events in the future, although gene-silencing might be encountered when transgenes are driven by the same promoter. Alternatively, transgene integration into the plastid genome for expression by chloroplast transformation is particularly attractive, because of the high-level accumulation of foreign protein in chloroplasts, absence of gene silencing and improved transgene containment. Previously, the expression of the β-glucosidase gene from the thermophilic bacterium T. fusca or fungus A. niger expressed in transplantomic tobacco was 2% to 5% of TSP without causing any deleterious phenotypic effect (Petersen and Bock 2011; Yu et al. 2007; Espinoza-Sánchez et al. 2016). To further increase the transgene level in transplastomic tobacco, the β-glucosidase from T. fusca was fused with different downstream box regions, and the recombinant enzyme accumulated up to 9.6% of TSP (Gray et al. 2011). In a recent study, the expression of β-glucosidase gene from a hyperthermophilic species of Archaea, Pyrococcus furiosus, in transplastomic tobacco could reach up to 75% of TSP (Castiglia et al. 2016). These examples demonstrate that using transplastomic plants is useful to produce recombinant β-glucosidase proteins. Therefore, the level of β-glucosidase gene from S. solfataricus might be further increased in transplastomic plants in near future.
The human β-glucocerebrosidase (acid β-glucosidase) has been overexpressed in plant cells for the enzyme replacement therapy of Gaucher disease, and N-linked glycosylation was required for the enzymatic activity (Tekoah et al. 2013, 2015). We predicted the presence of a potential N-linked glycosylation site (Asn-Ser-Ser) in theβ-glucosidase of S. olfataricus by using NetNGlyc 1.0 Server (https://www.cbs.dtu.dk/services/NetNGlyc/). The N-linked glycosylation of proteins first processed in ER, and then further modified in Goigi apparatus through the secretory pathway in plants (Strasser 2014). Therefore, in the present study, the β-glucosidase in ER- or vacuole-expressed lines might be glycosylated, but not that of chloroplast-expressed line. However, we did not observe any change of molecular weight in β-glucosidase via the SDS-PAGE among chloroplast-, ER-, or vacuole-expressed lines (Fig. 2a). In addition, the enzymatic activity of plant-expressed β-glucosidase was positively co-related with their expression level among transgenic lines (Fig. 2a and b). Our analysis suggested that plant-expressed β-glucosidase conferred no glycosylation.
The β-glucosidase has been shown to play an important role in regulating homeostatic levels of plant hormones by converting the glucose-conjugated inactive form to an active form. Therefore, overexpression and accumulation of β-glucosidase in different subcellular compartments not only has a profound effect on their expression level, but also might influence plant growth and development as well as respond to the environment. For example, transgenic tobacco plants overexpressing β-glucosidase from maize showed a substantial increase in cytokinin level by releasing active cytokinin from the conjugated form, although the plants showed no apparent alteration in phenotype (Kiran et al. 2006). However, overexpression of β-glucosidase from Arabidopsis in creeping bentgrass resulted in higher abscisic acid levels and enhanced drought tolerance but a dwarf phenotype as compared with the wild type (Han et al. 2012). In addition, overexpression of β-glucosidase and accumulation in the vacuole of Artemisia annua could increase trichome density to 20% and 66% in leaves and flowers, respectively, which also enhanced artemisinin yield in transgenic plants (Singh et al. 2016). Furthermore, expression of the β-glucosidase gene from A. niger in transgenic tobacco had a significant effect on volatile emissions from leaves (Wei et al. 2004). Expression of the β-glucosidase gene from A. niger in apoplasts rather than other compartments significantly delayed seed germination and flowering and increased the insecticidal effect and density of secretory glandular trichomes in leaf (Wei et al. 2007). Additionally, expression of a β-glucosidase gene from the ruminal bacterium Butyrivibrio fibrisolvens in transgenic tobacco increased salicylic acid levels and conferred more resistance to tobacco mosaic virus (Yao et al. 2007). Furthermore, the β-glucosidase gene of T. maritima driven by 35S promoter and overexpressed in tobacco effectively targeted the vacuole. The recombinant enzyme could liberate the conjugated form of abscisic acid, indole-3-acetic acid, and cytokinin stored in the vacuole to enhance growth, development, and stress tolerance in transgenic plants (Nguyen et al. 2015). Moreover, transplastomic tobacco plants expressing the β-glucosidase gene from Trichoderma reesei showed increased biomass, height, trichome density, and leaf area; the elevated levels of gibberellin, cytokinin and auxin might have contributed to the phenotypic effect in plants (Jin et al. 2011). In the present study, hyperthermophilic β-glucosidase rather than mesophilic or thermophilic enzymes selected for overexpression in tobacco could minimize the enzymatic effect resulting from residual activity at plant growth temperatures. Indeed, transgenic seedlings expressing the β-glucosidase gene from S. solfataricus and accumulated in subcompartments of chloroplast, ER or vacuole showed no obvious developmental defects in shoot growth (Supplementary Fig. 8a), but slightly suffered the growth retardation in root (Supplementary Fig. 8b and c).
The uidA encoding β-glucuronidase (GUS) from E. coli has been commonly used as a reporter gene in plants, because the intrinsic GUS activity of plant tissues is often rarely present and can be easily distinguished from the introduced activity (Jefferson et al. 1987). Despite the broad substrate of β-glucosidase (sso3019) from the archaebacterium S. solfataricus (Grogan 1991; Nucci et al. 1993), in the present study, we found that the plant-expressed archaebacterial β-glucosidase did not confer GUS activity (Fig. 5d and unpublished data). However, plant-expressed β-glucosidase also conferred high β-galactosidase activity (Fig. 5), which might provide additional advantages for the hydrolysis of complex sugars in biofuel conversion. In addition, β-galactosidase is commonly used as a reporter system in mammalian cells, insects and yeast (Naylor 1999) but not plants because of high endogenous activity. In the present study, the plant-expressed archaebacterial enzyme could hydrolyze the substrate under a broad range of temperature and pH (Supplementary Figs. 4 and 5). By heat-pretreatment or increasing the reaction temperature, the background noise of intrinsic β-galactosidase activity in plants could be greatly reduced or completely eliminated (Fig. 5). Therefore, the multi-functional archaebacterial β-glucosidase (sso3019) might be an alternative source as a reporter gene in plants. Besides, the cost of assays might be significantly decreased because the substrate X-gal for β-galactosidase is much cheaper than the substrate X-gluc for β-glucuronidase.
In conclusion, the β-glucosidase gene from S. solfataricus was driven by the rbcS promoter and overexpressed in transgenic tobacco. Recombinant β-glucosidase was accumulated in chloroplasts, ER and vacuoles. The plant-expressed β-glucosidase has optimal activity at 80 °C and pH 5–6. Plant-expressed β-glucosidase with accumulation in chloroplasts could reach up to 1% of TSP and showed higher activity than that in the ER and vacuole. Transgenic seedlings showed no obvious developmental defects in shoot growth, but slightly conferred the growth retardation in root during early development. To further increase the expression of β-glucosidase in plants, stacking the transgenes by crossing the plants among different transgenic events in the pBin-Cp line or different transgenic lines such as pBin-Cp, pBin-Er and pBin-Va might be feasible. In addition, on the basis of β-galactosidase activity, the archaebacterial β-glucosidase (sso3019) might provide an alternative source as a reporter gene in plants.
References
Aguilar CF, Sanderson I, Moracci M, Ciaramella M, Nucci R, Rossi M, Pearl LH (1997) Crystal structure of the β-glycosidase from the hyperthermophilic archeon Sulfolobus solfataricus: resilience as a key factor in thermostability. J Mol Biol 271:789–802
Ahmed A, Nasim F-H, Batool K, Bibi A (2017) Microbial β-glucosidase: sources, production and applications. Appl Environ Microbiol 5:31–46
Briante R et al (2000) Hydrolysis of oleuropein by recombinant β-glycosidase from hyperthermophilic archaeon Sulfolobus solfataricus immobilised on chitosan matrix. J Biotechnol 77:275–286
Castiglia D et al (2016) High-level expression of thermostable cellulolytic enzymes in tobacco transplastomic plants and their use in hydrolysis of an industrially pretreated Arundo donax L. biomass. Biotechnol Biofuels 9:154
Chang CC, Huang PS, Lin HR, Lu CH (2007) Transactivation of protein expression by rice HSP101 in planta and using Hsp101 as a selection marker for transformation. Plant Cell Physiol 48:1098–1107
D'Auria S, Moracci M, Febbraio F, Tanfani F, Nucci R, Rossi M (1998) Structure-function studies on β-glycosidase from Sulfolobus solfataricus. Molecular bases of thermostability. Biochimie 80:949–957
D'Auria S et al (1999a) β-glycosidase from the hyperthermophilic archaeon Sulfolobus solfataricus: structure and activity in the presence of alcohols. J Biochem 126:545–552
D'Auria S, Nucci R, Rossi M, Gryczynski I, Gryczynski Z, Lakowicz JR (1999b) The β-glycosidase from the hyperthermophilic archaeon Sulfolobus solfataricus: enzyme activity and conformational dynamics at temperatures above 100 °C. Biophys Chem 81:23–31
Diez-Municio M, Herrero M, Olano A, Moreno FJ (2014) Synthesis of novel bioactive lactose-derived oligosaccharides by microbial glycoside hydrolases. Microb Biotechnol 7:315–331
Espinoza-Sánchez EA, Torres-Castillo JA, Rascón-Cruz Q, Zavala-García F, Sinagawa-García SRJPBR (2016) Production and characterization of fungal β-glucosidase and bacterial cellulases by tobacco chloroplast transformation. Plant Biotech Reports 10:61–73
Frigerio L, Foresti O, Felipe DH, Neuhaus J-M, Vitale A (2001) The C-terminal tetrapeptide of phaseolin is sufficient to target green fluorescent protein to the vacuole. J Plant Physiol 158:499–503
Gray BN, Yang H, Ahner BA, Hanson MR (2011) An efficient downstream box fusion allows high-level accumulation of active bacterial β-glucosidase in tobacco chloroplasts. Plant Mol Biol 76:345–355
Grogan DW (1991) Evidence that β-galactosidase of Sulfolobus solfataricus is only one of several activities of a thermostable β-D-glycosidase. Appl Environ Microbiol 57:1644–1649
Gueguen Y, Chemardin P, Janbon G, Arnaud A, Galzy P (1996) A very efficient β-glucosidase catalyst for the hydrolysis of flavor precursors of wines and fruit juices. J Agric Food Chem 44:2336–2340
Han YJ, Cho KC, Hwang OJ, Choi YS, Shin AY, Hwang I, Kim JI (2012) Overexpression of an Arabidopsis β-glucosidase gene enhances drought resistance with dwarf phenotype in creeping bentgrass. Plant Cell Rep 31:1677–1686
Hoffman CS, Winston F (1987) A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 57:267–272
Huang LK, Liao SC, Chang CC, Liu HJ (2006) Expression of avian reovirus σC protein in transgenic plants. J Virol Methods 134:217–222
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901–3907
Jiang X, Zhou X, Liu Q, Zheng L, Yu N, Li W (2017) Expression of Acidothermus cellulolyticus thermostable cellulases in tobacco and rice plants. Biotechnol Biotechnol Equ 31:23–28
Jiang XR, Zhou XY, Jiang WY, Gao XR, Li WL (2011) Expressions of thermostable bacterial cellulases in tobacco plant. Biotechnol Lett 33:1797–1803
Jin S, Kanagaraj A, Verma D, Lange T, Daniell H (2011) Release of hormones from conjugates: chloroplast expression of β-glucosidase results in elevated phytohormone levels associated with significant increase in biomass and protection from aphids or whiteflies conferred by sucrose esters. Plant Physiol 155:222–235
Jung S, Kim S, Bae H, Lim HS, Bae HJ (2010) Expression of thermostable bacterial β-glucosidase (BglB) in transgenic tobacco plants. Bioresour Technol 101:7155–7161
Jung S, Lee DS, Kim YO, Joshi CP, Bae HJ (2013) Improved recombinant cellulase expression in chloroplast of tobacco through promoter engineering and 5' amplification promoting sequence. Plant Mol Biol 83:317–328
Kim BN, Yeom SJ, Kim YS, Oh DK (2012) Characterization of a β-glucosidase from Sulfolobus solfataricus for isoflavone glycosides. Biotechnol Lett 34:125–129
Kim S, Lee DS, Choi IS, Ahn SJ, Kim YH, Bae HJ (2010) Arabidopsis thaliana Rubisco small subunit transit peptide increases the accumulation of Thermotoga maritima endoglucanase Cel5A in chloroplasts of transgenic tobacco plants. Transgenic Res 19:489–497
Kiran NS et al (2006) Ectopic over-expression of the maize β-glucosidase Zm-p60.1 perturbs cytokinin homeostasis in transgenic tobacco. J Exp Bot 57:985–996
Lu SW, Wang KC, Liu HJ, Chang CD, Huang HJ, Chang CC (2011) Expression of avian reovirus minor capsid protein in plants. J Virol Methods 173:287–293
Montalvo-Rodriguez R, Haseltine C, Huess-LaRossa K, Clemente T, Soto J, Staswick P, Blum P (2000) Autohydrolysis of plant polysaccharides using transgenic hyperthermophilic enzymes. Biotechnol Bioeng 70:151–159
Moracci M, Nucci R, Febbraio F, Vaccaro C, Vespa N, La Cara F, Rossi M (1995) Expression and extensive characterization of a β-glycosidase from the extreme thermoacidophilic archaeon Sulfolobus solfataricus in Escherichia coli: authenticity of the recombinant enzyme. Enzyme Microb Technol 17:992–997
Naylor LH (1999) Reporter gene technology: the future looks bright. Biochem Pharmacol 58:749–757
Nguyen QA, Lee DS, Jung J, Bae HJ (2015) Phenotypic changes in transgenic tobacco plants overexpressing vacuole-targeted Thermotoga maritima BglB related to elevated levels of liberated hormones. Front Bioeng Biotechnol 3:181
Noh KH, Son JW, Kim HJ, Oh DK (2009) Ginsenoside compound K production from ginseng root extract by a thermostable β-glycosidase from Sulfolobus solfataricus. Biosci Biotechnol Biochem 73:316–321
Nucci R, Moracci M, Vaccaro C, Vespa N, Rossi M (1993) Exo-glucosidase activity and substrate specificity of the β-glycosidase isolated from the extreme thermophile Sulfolobus solfataricus. Biotechnol Appl Biochem 17:239–250
Otto RT, Bornscheuer UT, Syldatk C, Schmid RDJBL (1998) Synthesis of aromatic n-alkyl-glucoside esters in a coupled β-glucosidase and lipase reaction. Biotechnol lett 20:437–440
Outchkourov NS, Peters J, de Jong J, Rademakers W, Jongsma MA (2003a) The promoter-terminator of chrysanthemum rbcS1 directs very high expression levels in plants. Planta 216:1003–1012
Outchkourov NS, Rogelj B, Strukelj B, Jongsma MA (2003b) Expression of sea anemone equistatin in potato. Effects of plant proteases on heterologous protein production. Plant Physiol 133:379–390
Park SH, Ong RG, Sticklen M (2016) Strategies for the production of cell wall-deconstructing enzymes in lignocellulosic biomass and their utilization for biofuel production. Plant Biotechnol J 14:1329–1344
Perugino G, Trincone A, Rossi M, Moracci M (2004) Oligosaccharide synthesis by glycosynthases. Trends Biotechnol 22:31–37
Petersen K, Bock R (2011) High-level expression of a suite of thermostable cell wall-degrading enzymes from the chloroplast genome. Plant Mol Biol 76:311–321
She Q et al (2001) The complete genome of the crenarchaeon Sulfolobus solfataricus P2. Proc Natl Acad Sci USA 98:7835–7840
Singh ND, Kumar S, Daniell H (2016) Expression of β-glucosidase increases trichome density and artemisinin content in transgenic Artemisia annua plants. Plant Biotechnol J 14:1034–1045
Strasser R (2014) Biological significance of complex N-glycans in plants and their impact on plant physiology. Front Plant Sci 5:363
Tekoah Y et al (2013) Glycosylation and functionality of recombinant β-glucocerebrosidase from various production systems. Biosci Rep 33:e00071
Tekoah Y et al (2015) Large-scale production of pharmaceutical proteins in plant cell culture-the Protalix experience. Plant Biotechnol J 13:1199–1208
Tu JM, Chang MC, Huang LL, Chang CD, Huang HJ, Lee RH, Chang CC (2014) The blue fluorescent protein from Vibrio vulnificus CKM-1 is a useful reporter for plant research. Bot Stud 55:79
Wei S, Marton I, Dekel M, Shalitin D, Lewinsohn E, Bravdo BA, Shoseyov O (2004) Manipulating volatile emission in tobacco leaves by expressing Aspergillus niger β-glucosidase in different subcellular compartments. Plant Biotechnol J 2:341–350
Wei S, Semel Y, Bravdo B-A, Czosnek H, Shoseyov O (2007) Expression and subcellular compartmentation of Aspergillus niger β-glucosidase in transgenic tobacco result in an increased insecticidal activity on whiteflies (Bemisia tabaci). Plant Sci 172:1175–1181
Yao J, Huot B, Foune C, Doddapaneni H, Enyedi A (2007) Expression of a β-glucosidase gene results in increased accumulation of salicylic acid in transgenic Nicotiana tabacum cv. Xanthi-nc NN genotype. Plant Cell Rep 26:291–301
Yu LX, Gray BN, Rutzke CJ, Walker LP, Wilson DB, Hanson MR (2007) Expression of thermostable microbial cellulases in the chloroplasts of nicotine-free tobacco. J Biotechnol 131:362–369
Acknowledgements
This work was financially supported in part by the Grant (MOST 106-2313-B-006-003-MY3) to C.-C. Chang from the Ministry of Science and Technology, Taiwan.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Huang, CH., Huang, TL., Liu, YC. et al. Overexpression of a multifunctional β-glucosidase gene from thermophilic archaeon Sulfolobus solfataricus in transgenic tobacco could facilitate glucose release and its use as a reporter. Transgenic Res 29, 511–527 (2020). https://doi.org/10.1007/s11248-020-00212-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-020-00212-z