
Abstract
Browning, as the most important problem to be solved in callus differentiation, occupies an important position in tissue culture. In this study, plant regeneration was induced using leaves from germinated Isatis indigotica Fortune seedlings and mature plants as explants. Activated carbon (AC) and a solidification agent (Phytagel) were added simultaneously to inhibit browning. We measured the total phenol content and the PPO and POD activities in callus-induced seedling leaves and mature plant leaves of different ploidy at various stages. The results demonstrated that single plant growth regulators could induce calli with higher rooting and browning rates. Despite the addition of AC and phytagel to the medium, half of the mature plant calli turned yellow and eventually died. The total phenol content and POD activity in calli from mature leaves showed significant differences within 0–4 weeks (P < 0.05) but were still higher than the biochemical parameters in calli induced from seedlings, while the PPO activity did not differ significantly (P > 0.05). Mature materials of different ploidy yielded similar results. The addition of 2 mg·L−1 BAP, 0.1 mg·L−1 NAA, and 1.5 mg·L−1 AC and solidification with 0.2% phytagel in MS medium resulted in the highest callus regeneration rate and the lowest browning rate. During dedifferentiation, the browning of the two may be associated with PPO activity and total phenol content.
Key message
The regeneration system of Isatis indigotica Fortune was optimized and the browning problem was solved. The browning of callus is related to PPO activity and the total phenol content.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Isatis indigotica Fortune is a cruciferous herb. Its root is Isatidis Radix (Banlangen in Chinese), its leaf is Isatidis Folium (Daqingye in Chinese) (Chen et al. 2014), and it is also the original plant of indigo naturalis (Qing dai in Chinese) (Wang 2018). The primary active compounds isolated from this species comprise terpenoids, lignans, and indole alkaloids. These compounds have proven antiviral, antibacterial, anti-inflammatory, and antileukaemia properties (Kang et al. 2020). I. indigotica has been used as an important medicinal plant, the therapeutic amount required is relatively large, and demand exceeds supply. Effective prescriptions screened by traditional Chinese medicine to treat COVID-19 include “Three medicines and three prescriptions,” of which Isatidis Radix is the main component of the “Lianhuaqingwen capsule” (Xiao et al. 2020).
The leaves, roots, and hypocotyls of I. indigotica are often used to induce calli (Ke et al. 2009), and the induction rate of different parts is also different (Chen et al. 2015). Although the induced differentiation rate is high, browning is the most common problem. In recent years, many scholars have carried out extensive research on browning. These researchers believe that plant browning is caused by the action of oxidases, namely, polyphenol oxidase and peroxidase, including polyphenol oxidase (PPO) and peroxidase (POD), on natural substrate phenols (Sharma et al. 2019). In normal plant tissues, the simultaneous presence of substrate, oxygen, and PPO does not cause browning because polyphenols are distributed in vacuoles in normal tissue cells, and enzymes are distributed in various plastids and the cytoplasm. This regional distribution prevents the substrate from contacting polyphenols, thus avoiding the occurrence of enzymatic browning in normal tissues (Koohi et al. 2020). Phenols participate in respiratory metabolism as respiratory transmitters in normal plant tissues, and the redox between phenols and quinones is in dynamic balance. When plant tissue cells are damaged, the typical respiratory chain is interrupted, and the invasion of oxygen causes phenolic substances to rapidly oxidize to O-quinones under the action of oxidase and then form brown pigment or melanin through rapid nonenzymatic polymerization (Gong et al. 2015). As the most challenging problem to be solved in the regeneration system, there is an urgent need to solve the browning problem to improve the value-added rate of plants to accumulate secondary metabolites and characterize genes.
In this study, different concentrations of plant growth regulators (PGRs) were used to induce the dedifferentiation of the young leaves of seedlings germinated from I. indigotica seeds and the leaves of mature plants to form a callus and then to differentiate into seedlings. The effects of different antibrowning inhibitors on the induction and regeneration of calli were evaluated, and an effective high-frequency transformation method for indigo was established on this basis. This method can promote the breeding of excellent varieties of I. indigotica and effectively protect wild I. indigotica resources.
Materials and methods
Materials and conditions
Diploid seeds of I. indigotica (Baoding, Hebei) and tetraploid seeds of I. indigotica (Qianxian, Shaanxi) were supplied by Resources Sanjiu Medical & Pharmaceutical Co., Ltd. of China. Dr. Qiaosheng Guo of Nanjing Agricultural University identified the seeds. Mature I. indigotica plants (1 year old), including I. indigotica (Baoding, Hebei) and I. indigotica (Qianxian, Shaanxi), were planted at the Institute of Chinese Medicinal Materials, Nanjing Agricultural University, Nanjing, Jiangsu Province (32°2’6.25"N, 118°50’23.47"E, altitude, 50.21 m). The voucher specimen was kept at the Institute of Chinese Medicinal Materials, Nanjing Agricultural University.
Duan’s method was used to disinfect the seeds and mature plant leaves and conduct differentiation in Murashige and skoog (MS) medium with different concentrations of PGRS (Duan et al. 2019). The MS medium contained 3% (w·v−1) sucrose and 0.7% agar. The pH of the medium was 5.8-6.0, and the mixture was autoclaved at 121 °C for 20 min. Sucrose was purchased from China Guanghua Co., Ltd. MS medium and agar were purchased from Beijing Solarbio Technology Co., Ltd.
Callus induction
The leaves were from 14 d seedlings, and mature plants were cut into approximately 1 cm2 pieces with a blade and placed on callus induction medium (CIM). The medium contained 2,4-dichlorophenoxyacetic acid (2,4-D; 2, 1 or 0.1 mg·L−1), 1-naphthylacetic acid (NAA; 2, 1 or 0.1 mg·L−1), kinetin (KT; 2, 1 or 0.1 mg·L−1) and 6-benzylaminopurine (BAP; 2, 1 or 0.1 mg·L−1). After four weeks of culture, the quantity of callus generated through dedifferentiation was recorded.
Antibrowning treatments
The calli browned in the process of redifferentiation. We added activated carbon (AC) and phytagel to the medium antibrowning treatment. The MS medium used contains 0.00001% copper sorbitol, 0.5% sulfate, 0.7% agar or 0.2% phytagel, and AC. At the same time, 2 mg·L−1 BAP and 0.1 mg·L−1 NAA were added for culture. Leaves were placed on different treatment media, and the browning percentage was recorded over two weeks.
Redifferentiation
The experiment involved optimizing the medium using various proportions of auxin and cytokinin. Select the optimal treatment among antibrowning treatments to apply to the redifferentiation. The medium contained 0.00001% copper sorbitol, 0.5% sulfate, and different concentrations of BAP and NAA, i.e., high, middle, and low concentrations of 2, 1, and 0.1 mg·L−1, respectively. After four weeks, the percentage of differentiated calli was determined. The buds differentiated from the callus will continue to grow in the differentiation medium. after they grew to 2 ~ 5 cm, they were transferred to 1/2 MS and allowed to take root. After one month of cultivation, the rooted seedlings were transferred to flower pots and grown in a greenhouse (25 ± 1 °C).
Determination of biochemical parameters
The physiological indices of calli in redifferentiation medium were determined, including the total phenol content and activities of PPO and POD. Calli from 5 periods (0, 1, 2, 3, 4 weeks) were collected and dried at room temperature (20 ± 1 °C) and then ground into a fine powder. The dried samples were passed through a 100-mesh sieve and stored in a refrigerator at 4 °C. One gram of sample was weighed, and 3 mL of methanol was used for ultrasonic extraction for 60 min, followed by determination by spectrophotometry (Alpha-1506, Ltd., Shanghai, China). The total phenol content was determined by Folin–Ciocalteu colorimetry at OD725.
The activity of PPO was determined by spectrophotometry with catechol solution as the substrate. The results are expressed as OD525 min−1 g−1 of protein. The activity of POD was determined by spectrophotometry using guaiacol and H2O2 solution as substrates. The results are expressed as OD470 min−1 g−1 protein (Yang et al. 2009).
Statistical analysis
SPSS 23.0 was used for statistical analysis. Analysis of variance and Duncan’s multiple range test were used for significance analysis of the data.
Results
Induction of calli from various parts of I. indigotica
To identify the most efficient induction method, the leaves of seedlings and mature plants were treated (Fig. 1a, d). The leaves were transferred to CIM with BAP, NAA, 2,4-D, and KT, as listed in Table 1. Different hormones in the medium could induce calli, and the induction rate was 53 ~ 96%. BAP and NAA played an active role in the induction process. When the concentrations of BAP and NAA were 2 mg·L−1, the induction rates were over 90%. Calli appeared around the explants after 10 d of the culture-inducing medium. Figure 1b, e shows the calli of I. indigotica (Baoding, Hebei) growing around the perimeter after two weeks.

Callus-mediated plant regeneration in I. indigotica (Baoding, Hebei). a. Seven-day-old seedlings; b. Callus induction from the leaf tissue of seedlings on CIM for 3 weeks; c. A regenerated plantlet from a seedling-derived callus; d. Field-grown mature plants with young leaves used as explants; e. Callus induction from leaf tissue of field-grown mature plants on CIM for 3 weeks; f. Mature plant-derived calli on regeneration medium for 3 weeks; g. Rooting of a plantlet for 3 weeks; h. A regenerated plant transplanted to soil for 4 weeks Bars = 1 cm
Treatment with antibrowning agents in different media
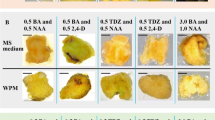
After incubation in MS medium supplemented with 0.7% agar, more than 70% of the mature explants were browned, and the seedlings were not all healthy (Fig. 1h). After adding AC and phytagel to the MS medium, 50% of the mature calli developed well, while 88–93% of the seedling calli grew without browning.
Different antibrowning agents were selected to study the difference in the callus differentiation rate between mature plants and seedlings. In other media, phytagel had a better antibrowning rate than agar, while AC did not differ significantly. Seedling calli were inoculated on regeneration medium with 2.0 g·L−1 phytagel. With 1.5 g·L−1 AC, browning started after 21 d, 12 d later than a mature callus. The callus differentiation in mature plants increased from 33 to 50%, but half were unhealthy. The best antibrowning medium included 1.5 g·L−1 AC solidified with 0.2% phytagel in MS medium, and the browning rate and rooting rate were the lowest under this condition (Table 2).
Callus redifferentiation to form seedlings
Although seedlings and mature explants could dedifferentiate to form calli, the ability to redifferentiate into shoots was quite different. Calli from the leaves of mature plants were over half browned and eventually died after two weeks (Fig. 1e, f). After 14 d, calli from seedlings redifferentiated into form buds (Fig. 1b, c). Combining BAP and NAA, two-factor induction was used to screen the dedifferentiation medium. The results showed that when 2 mg·L−1 BAP and 0.1 mg·L−1 NAA were added to the MS medium, the differentiation rate was the highest, reaching 85.8%. The rooting rate was 0%, and the differentiation rate was the highest (Table 3).
When the seedlings grew to 3 ~ 5 cm on the redifferentiation medium, the seedlings were cut and inoculated into 1/2 MS medium. After approximately 18 d, the seedlings developed roots (Fig. 1g). After opening the sealing film on the medium, cultivating the seedlings for one week, and then transferring them to potting soil (Fig. 1h), a higher survival rate could be obtained.
Comparative analysis of the total phenol content and the PPO and POD activities in different materials
Browning is the most common problem when growing in medium (Tang et al. 2004). When the growth reaches a particular stage, it will turn black, affecting the callus’s growth and development. Studies have shown that these browning phenomena are usually related to the total phenol content and oxidase activity (PPO and POD). Plants of different ploidy in experiments demonstrated the same trend. The total phenol content and the PPO and POD activities in calluses induced from seed-derived seedlings was lower than those in calluses induced from mature plants.
The total phenol content and the activities of PPO and POD all declined in diploids (I. indigotica, Baoding) and tetraploids (I. indigotica, Xianxian) during the differentiation culture period. As time passed, the decreases observed in calli derived from seedlings were higher than those of calli induced from mature plants. After four weeks, the total phenol content and the activities of POD and PPO in the calli derived from diploid seedlings decreased by 53.4%, 42.2%, and 65.9%, respectively. The calli derived from the mature diploid leaves decreased by 42.6%, 49.3%, and 30.7%. The total phenol content and PPO activity of calli derived from diploid seedlings were significantly lower than those before four weeks.
In contrast, the POD activity did not change significantly but was still lower than that of calli derived from mature plant leaf tissue. The tetraploid also showed the same trend (Table 4). The differences in callus browning during culture might be related to the total phenol content and PPO activity.
Discussion
Optimization of plant growth regulators in the I. indigotica regeneration system
Establishing an efficient, rapid, and stable genetic transformation system is essential to validate candidate genes (Abdallat et al. 2011). With the gradual modernization of traditional Chinese medicine, plant tissue culture technology can rapidly reproduce plant tissue in vitro in a short period (Sundararajan et al. 2022). This method is fast, efficient, controllable and convenient for large-scale industrial production. Callus-mediated transformation is the most common method of obtaining transgenic plants (Xu et al. 2017). Reportedly, the stems, hypocotyls, true leaves, cotyledons, and petioles of watercress can induce a callus, and its leaves and stems have higher induction rates (Li et al. 2023). Callus induction experiments in Erysimum scoparium (Cruciferous) explants cultured in MS medium showed that the leaves and hypocotyls have a high induction rate, while the roots have a low induction rate. It has also been reported that the leaves of 10- to 14-d-old sterile cauliflower seedlings are better than 15- to 20-d-old leaves. Moreover, the differentiation and induction rates of protoplasts are higher. The vacuoles are small and dispersed, and the chloroplasts are uniformly distributed (Gerszberg 2018). Consequently, leaves are the best material for inducing calli, and their cell totipotency determines the differentiated shoots. In Narcissus confusus (Amaryllidaceae), calli induced by sterile seedlings cultured from seed had a high regeneration capacity. Moreover, all regenerated plants showed typical morphological characteristics (Sellés et al. 1999; Santos et al. 2020). Using sterile seedling explants to induce differentiation for plant regeneration is ideal for cultivating directly inserted petiolate cotyledons (Yao et al. 2011).
The commonly used exogenous plant growth substances can be divided into two categories, auxin and cytokinin. The combination of auxin and cytokinin in proportion can promote plant induction (stem induction, proliferation induction, callus induction, and tissue culture seedling rooting) (Meng et al. 2014). When the two are present in low amounts, the induction effect is not significant. Excessively high concentrations not only inhibit the formation of buds but also increase the differentiation of roots. He found that different plant growth substance combinations of 2,4-D, 6-BA, and NAA played a crucial role in Brassica oleracea var. albiflora Kuntze. The optimal induction medium was MS + 0.1 mg·L−1, 2,4-D + 0.02 mg·L−1 6-BA, and the induction rate of hypocotyls was as high as 80%. The optimal medium for the regeneration culture process was MS + 0.75 mg·L−1 6-BA + 0.03 mg·L−1 NAA, with the adventitious bud differentiation rate and rooting rate reaching 94% and 67%, respectively (He et al. 2019). In general, 6-BA is used in the proliferation culture of the primary induction medium; NAA is used in the induction of callus, monocotyledonous plants require more hormone concentration than dicotyledonous plants, and induction rooting culture generally uses indole-3-butyric acid (IBA).
In this experiment, the PGR types and combinations of concentrations were intricate, different concentrations of a single PGR were used to induce calli, and the ratios of dual PGR combinations was optimized to induce calli to differentiate into shoots. In MS medium, different concentrations of PGRs, including BAP, NAA, 2,4-D, and KT, could cause calli at rates higher than 80% (Table 1). Both mature plants and 14-d-old seedlings can induce calli and redifferentiate into seedlings, and the differentiation rate of seedlings is higher than that of mature plants. Table 1 shows that NAA and BAP can induce higher levels of callus formation. Based on previous reports (He et al. 2019), the concentration range was determined to be 0.1 ~ 2 mg·L−1. A combination of BAP and NAA was used to induce adventitious buds. The results showed that the callus induction rate was the highest under treatment with 2 mg·L−1 BAP and 0.1 mg·L−1 NAA, especially since its rooting rate was 0%. The differentiation rate of different explants exhibited significant disparity upon additional processing, as shown in Table 3.
Causes of browning in I. indigotica
Although the regeneration medium included browning inhibitors, calli derived from mature leaves still easily browned and were challenging to differentiate. We wanted to take measures to reduce browning to confirm the reasons for rooting and browning in mature explants. There are many ways to alleviate or delay the browning of explants in tissue culture, such as antioxidants (Aghayeh et al. 2021) and altering culture conditions such as the temperature, pH, moisture, and salinity stress (Ndakidemi et al. 2014). Duan added AC, ascorbic acid (ASA) and solidification agent to the medium and found that the nonbrowning rate of calli from seedlings in the regeneration medium containing phytagel and ASA was as high as 83.3%, which was consistent with previous results (Duan et al. 2019). The browning rate was reduced by changing the curing agent of the medium in this experiment. Compared with agar, phytagel significantly affected the callus, delaying the browning rate by 2 d and increasing the browning rate by 16.7% and 8%, respectively (Table 2). The results showed that adding phytagel to the medium could effectively alleviate the browning rate of I. indigotica calli.
Although phytagel was used in the regeneration medium, the calli induced from the leaves of mature plants were mostly browned and not differentiated. Likewise, the leaves from young shoots germinated from Isodon amethystoides seeds induced a higher rate of callus differentiation compared with leaves from mature plants (Duan et al. 2019). This finding shows that the growth vigour of the leaves of young shoots germinated from seeds is higher than that of mature plants. At the same time, the oxidation reaction may also be another reason for its browning (Schuch et al. 2010).
However, polyphenols, as substrates for PPO and POD reactions, have been implicated as one of the factors affecting the browning of explants in tissue culture (Leng et al. 2009). The oxidation of phenolic compounds into their corresponding O-quinones catalysed by PPO is now well documented (Lim et al. 2019). PPO is also the major protease in plant tissue browning (Xu et al. 2015). Additionally, POD can oxidize various phenolic compounds, including monohydroxy and catholic phenolics (Soltani et al. 2023). PPO is responsible for the enzymatic browning of apples; apples lacking PPO activity might be helpful not only in the food industry but also in studies of the metabolism of polyphenols and the function of PPO (Wu et al. 2021). PPO and POD can convert phenolic compounds into O-quinones (Garcia et al. 2007). We determined the total phenol content and activities of PPO and POD in differentiating calli from seedling and mature plants at different ploidy levels. While POD activity exhibited an insignificant difference, the polyphenol content and PPO activity differed significantly. Both of them decreased in activities of PPO and total polyphenols. Materials with different ploidy levels demonstrated the same trend. These results are consistent with those for other plant species in previous reports. The activities of PPO and total polyphenols may be related to the browning of I. indigotica calli.
Conclusion
We optimized the I. indigotica plant regeneration system and selected the medium with the highest differentiation rate, i.e., 2 mg·L−1 BAP and 0.1 mg·L−1 NAA in MS medium. The solidification agent was optimized, and activated carbon was added to solve the browning problem. The total phenol content and the PPO and POD activities of materials of different ploidy were compared. The results showed that the browning rate was related to the PPO activity and the total phenolic substances. Using seedlings as starting materials, genetic transformation can lead to rapid differentiation with the lowest browning rate, which lays a foundation for subsequent gene mining.
Data Availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- Isatis indigotica Fortune:
-
I. indigotica
- BAP:
-
6-benzylaminopurine
- NAA:
-
1-naphthylacetic acid
- 2,4-D:
-
2,4-dichlorophenoxyacetic acid
- KT:
-
Kinetin
- MS:
-
Murashige and skoog
- CIM:
-
Callus induction medium
- PPO:
-
Polyphenol oxidase
- POD:
-
Peroxidase
- AC:
-
Activated carbon
- ASA:
-
Ascorbic acid
- D:
-
Day
- H:
-
Hour
- Min:
-
Minute
References
Abdallat AM, Suwwan J, Zoubi B (2011) Agrobacteriumtumefaciens-mediated transformation of callus cells of Crataegusaronia. Plant Cell Tiss Org 104(1):31–39. https://doi.org/10.1007/s11240-010-9798-1
Aghayeh R, Bahram A, Ahmad B, Leila S, Ali T (2021) The first successful report: control of Browning Problem inin vitroCulture of iranian Seedless Barberry, aMedicinally important species. Erwerbs-Obstbau 63(3):319–329. https://doi.org/10.1007/S10341-021-00574-6
Chen R, Li Q, Tan H, Chen J, Xiao Y, Ma R, Gao S, Zerbe P, Chen W, Zhang L (2015) Gene-to-metabolite network for biosynthesis of lignans in MeJA-elicited Isatis indigotica hairy root cultures. Front Plant Sci 6:952. https://doi.org/10.3389/fpls.2015.00952
Chen S, Zhu Z, Ma H, Yang J, Guo Q (2014) DNA barcodes for discriminating the medicinal plant Isatis indigotica Fort. (Cruciferae) and its adulterants. Biochem Syst Ecol 57:287–292. https://doi.org/10.1016/j.bse.2014.08.007
Duan Y, Su Y, Chao E, Zhang GR, Zhao FL, Xue T, Sheng W, Teng JT, Xue JP (2019) Callus-mediated plant regeneration in Isodon amethystoides using young seedling leaves as starting materials. Plant Cell Tiss Org 136(2):247–253. https://doi.org/10.1007/s11240-018-1510-x
Garcia A, Wichers H, Wichers HJ (2007) Decrease of the IgE-binding by Mal d 1, the major apple allergen, by means of polyphenol oxidase and peroxidase treatments. Food Chem 103(1):94–100. https://doi.org/10.1016/j.foodchem.2006.07.029
Gerszberg A (2018) Tissue culture and genetic transformation of cabbage (Brassica oleracea var. capitata): an overview. Planta 248:1037–1048. https://doi.org/10.1007/s00425-018-2961-3
Gong ZQ, Li DJ, Liu CQ, Cheng AW, Wang WL (2015) Partial purification and characterization of polyphenol oxidase and peroxidase from chestnut kernel. Lwt-Food Sci Technol 60(2):1095–1099. https://doi.org/10.1016/j.lwt.2014.10.012
He Y, Liang S, Zheng H, Zhang F, Sun B (2019) Callus Induction and Plant Regeneration in Chinese Kale callus induction and plant regeneration in chinese kale. AIP Conference Proceedings 2058(1):1–4. https://doi.org/10.3724/sp.j.1259.2011.00569
Kang M, Wu H, Yang Q, Huang H, Liu J (2020) A chromosome-scale genome assembly of Isatis indigotica, an important medicinal plant used in traditional chinese medicine. Hortic Res-England 7(1):10. https://doi.org/10.1038/s41438-020-0240-5
Ke SY, Zhang SZ, Liu YJ (2009) Changes of antioxidant enzyme activity during vitrification of Isatis indica in vitro. Dissertation North China Agricultural Journal 24(3):5. https://doi.org/10.7668/hbnxb.2009.03.033
Koohi S, Nasernejad B, Zare MH, Elahifard M, Shirazian S, Ghadiri M (2020) Extraction of oxidative enzymes from green tea leaves and optimization of extraction conditions. Chem Eng Technol 43(12):2548–2556. https://doi.org/10.1002/ceat.202000344
Leng P, Su S, Wei F, Yu F, Duan Y (2009) Correlation between browning, total phenolic content, polyphenol oxidase and several antioxidation enzymes during pistachio tissue culture. Acta Hort 829:127–132. https://doi.org/10.17660/ActaHortic.2009.829.17
Lim WY, Cheun CF, Wong CW (2019) Inhibition of enzymatic browning in sweet potato (Ipomoea batatas (L.)) with chemical and natural anti-browning agents. J Food Process Pres 43:11. https://doi.org/10.1111/jfpp.14195
Li Y, Zhao J, Chen H, Yu X, Li H, Zhang Y, Feng L, Wu Z, Xie W, Hou D, Yu M (2023) Plant regeneration via callus-mediated organogenesis in commercial variety of Chuanbeichai No. 1 in Bupleurum chinense DC. Plant Biotechnol Rep 17(2):159–169. https://doi.org/10.1007/s11816-022-00772-y
Meng Q, Liu ZY, Zhang Y, Liu C, Ren F, Feng H (2014) Effects of antibiotics on in vitro-cultured cotyledons. In Vitro Cell Dev-Pl 50(4):436–441. https://doi.org/10.1007/s11627-014-9595-3
Ndakidemi CF, Mneney E, Ndakidemi PA (2014) Effects of ascorbic acid in controlling lethal browning in culture of using nodal segments. Am J Plant Sci 05(1):187–191. https://doi.org/10.4236/ajps.2014.51024
Santos GS, Sinoti SBP, De Almeida FTC, Silveira D, Simeoni LA, Gomes-Copeland KKP (2020) Use of galantamine in the treatment of Alzheimer’s disease and strategies to optimize its biosynthesis using the in vitro culture technique. Plant Cell Tiss Org: An International Journal on in Vitro Culture of Higher Plants 143(1):13–29. https://doi.org/10.1007/s11240-020-01911-5
Schuch MW, Cellini A, Masia A, Marino G (2010) Aluminium-induced effects on growth, morphogenesis and oxidative stress reactions in in vitro cultures of quince. Sci Hortic-Amsterdam 125(2):151–158. https://doi.org/10.1016/j.scienta.2010.03.007
Sellés M, Viladomat F, Bastida J, Codina C (1999) Callus induction, somatic embryogenesis and organogenesis in Narcissus confusus: correlation between the state of differentiation and the content of galanthamine and related alkaloids. Plant Cell Rep 18(7–8):646–651. https://doi.org/10.1007/s002990050636
Sharma H, Chawla N, Dhatt AS (2019) Nutraceutical content and free radical scavenging capacity of brinjal (Solanum melongena L.) genotypes. Sci Hortic-Amsterdam 244:294–303. https://doi.org/10.1016/j.scienta.2018.09.060
Soltani M, Samari E, Vazirifar S, Ahmadian CN, Sharifi M, Fotovat R (2023) Putrescine induces lignans biosynthesis through changing the oxidative status and reprogramming amino acids and carbohydrates levels in Linum album hairy roots. Plant Cell Tiss Org 153(2):387–402. https://doi.org/10.1007/s11240-023-02479-6
Sundararajan S, Rajendran V, Sivakumar HP, Kumariah M, Ramalingam S (2022) Growth modulation by nitric oxide donor sodium nitroprusside in in vitro plant tissue cultures–A review. Biologia 77(7):1699–1711. https://doi.org/10.1007/s11756-022-01027-5
Tang W, Newton, RJ (2004) Increase of polyphenol oxidase and decrease of polyamines correlate with tissue browning in Virginia pine (Pinus virginiana Mill.). Plant Sci 167(3):621–628. https://doi.org/10.1016/j.plantsci.2004.05.024
Wang L, Xue YN, Li ZW, Zhang W, Ji XP, Fan Z, Li YJ (2018) Efficacy and Safety of Indigo Naturalis in Combination with Narrow-Band Ultraviolet B for Treatment of Pityriasis Rosea: A Meta-Analysis. Evid Based Complement Alternat Med 2018:6816981. https://doi.org/10.1155/2018/6816981
Wu G, Wei X, Wang X, Wei Y (2021) Changes in biochemistry and histochemical characteristics during somatic embryogenesis in Ormosia henryi Prain. Plant Cell Tiss Org 144:505–517. https://doi.org/10.1007/s11240-020-01973-5
Xiao M, Tian J, Zhou Y, Xu X, Tong X (2020) Efficacy of Huoxiang Zhengqi dropping pills and Lianhua Qingwen granules in treatment of COVID-19: a randomized controlled trial. Pharmacol Res 161:105–126. https://doi.org/10.1016/j.phrs.2020.105126
Xu CJ, Ru ZW, Li L, Zeng BY, Huang JM, Huang W, Hu O (2015) The Effects of Polyphenol Oxidase and Cycloheximide on the Early Stage of Browning in Phalaenopsis Explants. Hortic Plant J 1(3):172–180. https://doi.org/10.16420/j.issn.2095-9885.2015-0030
Xu K, Zhang X, Fan C, Chen F, Zhu J, Zhang S, Chen Q, Fu Y (2017) A callus transformation system for gene functional studies in soybean. J Integr Agr 16(9):1913–1922. https://doi.org/CNKI:SUN:ZGNX.0.2017-09-004
Yang H, Guo X, Lu R, Niu B, Pasapula V, Pei H, Cai F, Ying X, Chen F (2009) Changes in morphology and biochemical indices in browning callus derived from Jatropha curcas hypocotyls. Plant Cell Tiss Org 98(1):11–17. https://doi.org/10.1007/s11240-009-9533-y
Yao SC, Ling ZZ, Cui LI, Hui-Zhen LU, Zhang ZJ (2011) Research on differentiation and plant regeneration from in vitro leaf tissues of chirita medica d.fang ex w.t.wang. Seed 30(12):30–29. https://doi.org/10.1016/j.jnca.2010.09.003
Funding
This work was supported by the Jiangsu Modern Agricultural Industrial Technology System Construction Project (JATS|[2022]461, JATS[2022]291).
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: YS, MW, QG. Performed the experiments: YS, MW, QG. Analysed the data: YS, MW, QG, JH, KZ, JH. Wrote the paper: YS, MW, QG, JH, KZ, JH.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
We thank all authors for their contributions to this work.
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Communicated by Christell van der Vyver.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Yong Su and Min Wei share co-first authorship.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Su, Y., Wei, M., Guo, Q. et al. Investigating the relationships between callus browning in Isatis indigotica Fortune, total phenol content, and PPO and POD activities. Plant Cell Tiss Organ Cult 155, 175–182 (2023). https://doi.org/10.1007/s11240-023-02567-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-023-02567-7