
Abstract
Two species of Bychowskicotylidae (Monogenoidea: Gastrocotylinea) were collected from the gills of the sweetlips Diagramma labiosum Macleay (Haemulidae) obtained from Moreton Bay, Queensland, Australia, during January 2016. Lebedevia n. gen. is proposed for bychowskicotylids having 1) a bilaterally symmetrical haptor with four pairs of gastrocotylid-like clamps and lacking an armed lappet (anchors and hooks absent); 2) longitudinal sclerotized internal ridges lining the vas deferens, 3) an unarmed male copulatory organ, and 4) four interconnected vesicles located posterior to the genital atrium and lying on the body midline dorsal to the uterus and ventral to the vas deferens. Lebedevia lebedevi n. sp., the type and only species of the genus, is described, and Tonkinopsis transfretanus Lebedev, 1972 is recorded for the first time in Moreton Bay. Yamaguticotyla Price, 1959 (Bychowskicotylidae) is considered nomen nudum, and the authorship of the genus is corrected to Yamaguticotyla Yamaguti, 1963.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During a survey of the parasites of marine fishes occurring in Moreton Bay, Queensland, Australia, two bychowskicotylid species (Monogenoidea; Gastrocotylinea) were collected from the gills of the sweetlips Diagramma labiosum Macleay (Haemulidae). The helminth specimens were identified as Tonkinopsis transfretanus Lebedev, 1972 and as an undescribed species herein assigned to a new genus. Species of Benedenia and Encotyllabe (both Capsalidae) were also collected from the sweetlips and will be formally reported in a subsequent paper. The present paper is the seventh on the monogenoids recovered from Moreton Bay during the survey (see Kritsky, 2018a, b, 2019; Kritsky & Nitta, 2019; Kritsky & Chisholm, 2020; Chisholm & Kritsky, 2020) and includes the diagnosis for the new genus, the description of the new species, and the first report of T. transfretanus in Morton Bay.
Materials and methods
Specimens of the sweetlips D. labiosum were collected from Moreton Bay, Queensland, Australia, during January 2016. The fishes were transported alive to the Moreton Bay Research Station located in Dunwich, North Stradbroke Island, Queensland, where they were euthanized, identified using Johnson (2010), and necropsied for parasitic infections. Methods for collection, preparation, illustration, and measurement of the monogenoidean specimens were those of Kritsky (2018a). Measurements, all in micrometers, represented straight-line distances between extreme points and were expressed as the range followed by the mean and number (n) of structures measured in parentheses; body length included that of the haptor. The scientific name of the host was that presented in Johnson (2010) and verified in Fricke et al. (2022). Type and voucher specimens of helminths were deposited in the Queensland Museum, Brisbane, Australia (QM), and the University of Nebraska State Museum, Harold W. Manter Laboratory, Lincoln, Nebraska, U.S.A. (HWML), as indicated in the following species accounts.
Results
The gills of two sweetlips D. labiosum were examined for monogenoids. Both hosts were parasitized by two species of Bychowskicotylidae: Lebedevia lebedevi n. gen., n. sp. (hosts infected with one and five specimens, respectively) and T. transfretanus (hosts infected with one specimen each).
-
Subclass Heteronchoinea Boeger & Kritsky, 2002
-
Infrasubclass Oligonchoinea Bychowsky, 1937
-
Order Mazocraeidea Bychowsky, 1937
-
Suborder Gastrocotylinea Lebedev, 1972
-
Bychowskicotylidae Lebedev, 1969
Lebedevia n. gen.
Type and only species: Lebedevia lebedevi n. sp. from Diagramma labiosum Macleay (Haemulidae).
Etymology: The names of the genus and its type species are proposed in honor of the late Dr. Boris Lebedev, formerly of the Far East Science Center, USSR Academy of Sciences, Vladivostok, in recognition of our friendship and as an acknowledgment of his important contributions regarding the taxonomy and systematics of the Monogenoidea, particularly those dealing with the Suborder Gastrocotylinea. Dr. Lebedev’s (1986) comprehensive revision of the suborder remains the principal reference for anyone working on this group of helminths.
ZooBank registration: To comply with the regulations set out in article 8.5 of the 2012 version of the International Code of Zoological Nomenclature (ICZN, 2012), details of the new genus have been submitted to ZooBank. The Life Science Identifier (LSID) for Lebedevia n. gen. is urn:lsid:zoobank.org:act:D3DB2C60-9965-4C79-B79E-6109FF9CDF38.
Diagnosis
Body comprising cephalic region, trunk, haptor; peduncle absent. Haptor symmetrical, with two bilateral rows of four gastrocotylinean clamps each; haptoral lappet, anchors, hooks absent. Paired prohaptoral suckers with septa, small denticles sensu Chisholm et al. (1996) along margins of anterior lips. Eyespots absent. Mouth subterminal, ventral; pharynx muscular; esophagus bifurcating to form two intestinal ceca; ceca terminating blindly in haptor. Common genital pore midventral at level of esophageal bifurcation; genital atrium muscular, armed with elongate delicate spines. Male copulatory organ (MCO) unarmed. Seminal vesicle absent; vas deferens with longitudinal internal sclerotized ridges extending from level of anterior limit of gonadal field into MCO. Four interconnected vesicles lying posterior to genital atrium, ventral to vas deferens, dorsal to uterus. Testes numerous, intercecal, postgermarial. Germarium shaped as inverted U; genitointestinal canal present; seminal receptacle lying to left of body midline anterior to testicular field; uterus extending along body midline to genital atrium and ventral to vas deferens. Vitellarium coextensive with intestinal ceca, extending into haptor; vitelline reservoir ventral to uterus. Vaginal pore, vagina, egg not observed.
Remarks
The Bychowskicotylidae presently contains Yamaguticotyla Yamaguti, 1963 (syn. Yamaguticotyla Price, 1959, see below) with two species and four monotypic genera: Bychowskicotyle Lebedev, 1969, Tonkinopsis Lebedev, 1972, Gaterinia Lebedev, 1972, and Lebedevia n. gen. All bychowskicotylids are parasitic on the gills of haemulid fishes of the Indo-Pacific Ocean. Lebedevia n. gen., represented by its type and only species Lebedevia lebedevi n. sp., differs from other genera assigned to the Bychowskicotylidae by its species having 1) a bilaterally symmetrical haptor armed with four pairs of gastrocotylid-like clamps (10 to 13 pairs in Bychowskicotyle spp., six or seven in Gaterinia spp., 10–12 in Yamaguticotyla spp., and three in Tonkinopsis spp.), 2) sclerotized ridges lining the vas deferens (ridges absent in all other bychowskicotylids), 3) an unarmed MCO (MCO armed with spines in the remaining species of the family), and 4) four interconnected vesicles located posterior to the genital atrium and lying dorsal to the uterus and ventral to the vas deferens (homologs absent in other bychowskicotylid spp.). Lebedevia is most similar to Tonkinopsis by their species having comparatively few pairs of haptoral clamps (see Lebedev, 1986; Chisholm et al., 1996).
Lebedevia lebedevi n. sp.
Type host: Sweetlips, Diagramma labiosum Macleay (Haemulidae).
Type locality: Moreton Bay off Dunwich, North Stradbroke Island, Queensland, Australia (27°29’S, 153°23’E), 10, 19 January 2016.
Infection site: Gills.
Specimens studied: Holotype, QM G240115; 5 paratypes, QM G240116–G240117, HWML 216821.
ZooBank registration: To comply with the regulations set out in Article 8.5 of the 2012 version of the International Code of Zoological Nomenclature (ICZN, 2012), details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) for Lebedevia lebedevi n. sp. is urn:lsid:zoobank.org:act:75B741AD-2D85-4C69-BA8C-810F7CFCA4C9.
Description (Figs. 1–8)

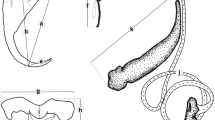
Lebedevia lebedevi n. sp. from the gills of the sweetlips Diagramma labiosum from Moreton Bay, Queensland, Australia. 1, Whole-mount (composite, ventral view); 2, Haptoral clamp (ventral view; the perpendicular distance between the short parallel lines indicates the measurement [width] taken during this study); 3, Hook of the genital atrium; 4, Terminal male genitalia (ventral view); 5, Medial clamp sclerite (ventral view); 6, Left accessory clamp sclerite; 7, Left ventrolateral clamp sclerite (ventral view); 8, Left dorsolateral clamp sclerite. Abbreviations: g, germarium; ga, genetial atrium; gp, genital pore; iv, interconnected vesicles; mco, male copulatory organ; p, pharynx; ps, prohaptoral sucker; sr, seminal receptacle; t, testis; u, uterus; vd, vas deferens; vr, vitelline reservoir.
Body proper dorsoventrally flattened, tapering anteriorly from level of testicular field; haptor poorly differentiated from trunk (Fig. 1). Haptoral clamps subequal in size (Fig. 2), pedunculate; peduncles short. Ventrolateral clamp sclerite with ventral and dorsal arms, each with blunt ends (Fig. 7); posterolateral clamp sclerite J shaped, with acute blade-like posterior end (Fig. 8); median clamp sclerite (spring) with straight ventral arm having rounded tip, vase-shaped dorsal arm having two spikes directed posteriorly (Fig. 5); accessory clamp sclerite fusiform with medial end bent perpendicularly and directed posteriorly in clamp (Fig. 6). Each prohaptoral sucker with thickened posterior lip, anterior lip with numerous small denticles, internal septum delicate, often inconspicuous. Mouth located in cephalic depression, flanked by prohaptoral suckers. Pharynx subspherical, immediately posterior to mouth and prohaptoral suckers; esophagus elongate, bifurcating dorsal to genital atrium to form intestinal ceca; esophageal and intestinal diverticula absent. Common genital pore anterior to genital atrium; genital atrium doughnut shaped, with thin-walled cylinder arising from its inner circular margin, armed with approximately 10 delicate hair-like spines (Figs. 3, 4). Approximately 20–30 subspherical testes. MCO penis like, usually protruding from genital atrium and genital pore; prostates not observed. Germarium an inverted U; oviduct directed anteriorly from distal end of germarium, then recurved posteriorly to unite with other female ducts; genitointestinal canal uniting with right intestinal cecum; vitelline reservoir bifurcating anterior to germarium. Vitellarium coextensive with intestinal ceca, extending into haptor.

Genital corona of Tonkinopsis transfretanus Lebedev, 1972, from the gills of sweetlips Diagramma labiosum from Moreton Bay, Queensland, Australia.
Measurements: Body 1,080–1,280 (1,150; n = 5) long; width of trunk (at level of testicular field) 222–319 (278; n = 5). Haptor 267–301 (281; n = 3) long; 352–395 (372; n = 4) wide. Clamp pair 1 (anterior most) 61–67 (64; n = 4) wide; clamp 2 65–70 (68; n = 4) wide; clamp 3 64–75 (69; n = 5) wide; clamp 4 (posterior most) 65–70 (67; n = 5) wide. Prohaptoral sucker 38–44 (41; n = 5) long, 44–49 (46; n = 5) wide. Pharynx 33–43 (38; n = 5) wide. Testicular field 157–203 (186; n = 4) long, 74–121 (98; n = 4) wide; testis 34–48 (40; n = 4) in diameter. Genital atrium 37–47 (43; n = 5) long, 39–48 (44; n = 5) wide. Germarium 152–171 (161; n = 4) long, 104–143 (122; n = 5) wide.
Remarks
Lebedevia lebedevi n. sp. is the type species of the genus.
Tonkinopsis transfretanus Lebedev, 1972
(Fig. 9)
Type host: Crescent sweetlips, Plectorthynchus cinctus (Temminck & Schlegel) (Haemulidae).
Type locality: Gulf of Tonkin, South China Sea.
Current record: Sweetlips, Diagramma labiosum Macleay (Haemulidae): Moreton Bay off Dunwich, North Stradbroke Island, Queensland, Australia (27°29’S, 153°23’E), 10, 19 January 2016.
Previous records: P. cinctus: Gulf of Tonkin, South China Sea (Lebedev, 1972b). D. labiosum: Heron Island, Great Barrier Reef, Queensland, Australia (23°27’S, 151°55’E) (Chisholm et al., 1996).
Infection site: Gills.
Specimens studied: 2 voucher specimens, QM G240118–240119.
Remarks
Only two specimens of T. transfretanus were collected during the Moreton Bay survey; one was unstained and mounted in Gray & Wess medium and the other stained with hematoxylin and mounted in Canada balsam. Although the two specimens were insufficient for determination of soft anatomy, they clearly are conspecific with those identified as T. transfretanus from the Great Barrier Reef by Chisholm et al. (1996). Present specimens differed from the previous descriptions of the species only by 1) septa being present in the prohaptoral suckers and 2) presence of eight elongate narrow spines in the genital corona (Fig. 9). Lebedev (1972b) and Chisholm et al. (1996) reported a varying number of hooked and narrow spines in the genital corona (see figure 9 in Chisholm et al. [1996] and figure B in Lebedev [1972b]). In addition, Chisholm et al. (1996) considered the hooked spines to be associated with the genital atrium and the narrow spines with the MCO, but these associations were not confirmed in the present specimens.
The finding of T. transfretanus on the gills of the sweetlips in Moreton Bay represents a new faunal record for the bay.
Discussion
Lebedev (1969) proposed the Bychowskicotylinae Lebedev, 1969 as a subordinate taxon of the Gastrocotylidae Price, 1943. The subfamily initially included three genera: Bychowskicotyle Lebedev, 1969, Gaterinia Lebedev, 1969, and Yamaguticotyla Price, 1959 (now Yamaguticotyla Yamaguti, 1963, see below). Shortly thereafter, Lebedev (1972a, b) proposed the Calyxinellidae Lebedev, 1972, for Calyxinella Lebedev, 1972, and Tonkinopsis Lebedev, 1972. Upon determination that Calyxinella was a junior synonym of Yamaguticotyla, Lebedev (1984) elevated the Bychowskicotylinae to family status, to which two subfamilies, the Bychowskicotylinae (containing Bychowskicotyle and Gaterinia) and the Calyxinellinae (containing Yamaguticotyla and Tonkinopsis) were assigned. Prior to the present study, the Bychowskicotylidae contained five described species all parasitic on the gills of haemulid fishes: Bychowskicotyle plectorhynchi Lebedev, 1969 on Plectorhinchus cinctus (Temminck & Schlegel); Gaterinia talaensis Lebedev, 1969 on Gaterin punctatissimus (Playfair) [now Plectorhinchus picus (Cuvier)]; Tonkinopsis transfretanus Lebedev, 1972, on P. cinctus and Diagramma labiosum Macleay; Yamaguticotyla truncata (Goto, 1894) Price, 1959 on Pristipoma japonicum Cuvier [now Parapristipoma trilineatum (Thunberg)], and Yamaguticotyla jucunda (Lebedev, 1972) Lebedev, 1984 (syn. Calyxinella jucunda Lebedev, 1972) on an unidentified fish of the Pomadasidae (now Haemulidae) (see Lebedev, 1986; Chisholm et al., 1996).
In his most recent revision of the Suborder Gastrocotylinea, Lebedev (1986) characterized the Bychowskicotylidae to include monogenoids having postgermarial testes, a (bilaterally) symmetrical haptor armed with two rows of a small number of (gastrocotylid-like) clamps each, and copulatory organs (genital atrium and MCO) with a crown of curved hooks (parentheses ex nobis), while the Bychowskicotylinae and Calyxinellinae were differentiated primarily on morphological differences in the crown of hooks associated with the genital atrium and MCO. Although a feature of all bychowskicotylid species, absence of an armed haptoral lappet was considered a diagnostic feature of only the Bychowskicotylinae; the character was not mentioned in the diagnoses of either the family or that of the Calyxinellinae by Lebedev (1969, 1984, 1986).
Lebedevia n. gen. is clearly a member of the Suborder Gastrocotylinea and of the Bychowskicotylidae by its species having a bilaterally symmetrical haptor armed with comparatively few gastrocotylid-like clamps and lacking an armed haptoral lappet. Assignment of the genus at the subfamily level, however, is problematical. The genus is probably most closely aligned with the Bychowskicotylinae by its species having an unarmed MCO and non-diverging copulatory spines enclosed within a doughnut-shaped genital atrium. In species of the Calyxinellinae, the weakly developed genital atrium and the MCO are armed with diverging spines, a feature that excludes Lebedevia from the subfamily (see Chisholm et al., 1996). The genus differs from Bychowskicotyle and Gaterinia (both Bychowskicotylinae) by its members having four interconnected vesicles lying on the body midline posterior to the genital atrium, by possessing longitudinal sclerotized ridges lining the vas deferens, and by having four pairs of haptoral clamps (vesicles and ridges of the vas deferens absent and six or seven pairs of haptoral clamps in species of Gaterinia; vesicles and ridges absent and 10 to 13 pairs of clamps in species of Bychowskicotyle).
A note on the nomenclatural status of Yamaguticotyla Price, 1959
In a published abstract of a paper presented at the Thirty-Fourth Annual Meeting of the American Society of Parasitologists, Price (1959) introduced the genus-group Yamaguticotyla as a member of the Gastrocotylidae (Gastrocotylinae) with Yamaguticotyla truncata (Goto, 1894) Price, 1959 as its type and only species. Yamaguticotyla Price, 1959 was accepted as an available and valid taxon sensu the International Code of Zoological Nomenclature (ICZN) by Yamaguti (1963) and subsequently by Lebedev (1972a, b, 1984, 1986). However, Article 13.1.1 of the ICZN (International Commission on Zoological Nomenclature, 1999) requires that every new name published after 1930 must “be accompanied by a description or definition that states in words characters that … differentiate the taxon.” Other than introducing Yamaguticotyla as a new genus and identifying its type species, Price (1959) failed to provide any statement that could be considered to have fulfilled Article 13.1.1. Thus, Yamaguticotyla Price, 1959 did not meet the requirements for availability sensu the ICZN and is herein considered nomen nudum.
Yamaguti (1963) first made the epithet Yamaguticotyla available, when in addition to identifying the type species as Y. truncata, he provided a generic diagnosis for the taxon and fulfilled all other nomenclatural requirements (Articles 10 to 20) for publication. Thus, the correct authorship for the taxon is Yamaguticotyla Yamaguti, 1963, not Price, 1959.
Data availability
All data used in during this study is presented within the paper.
References
Chisholm, L. A., Agrawal, N., Whittington, I. D. (1996). A redescription and new host record of Tonkinopsis transfretanus Lebedev, 1972 (Monogenea: Polyopisthocotylea: Bychowskicotylidae), with a description of the oncomiracidium from Diagramma labiosum (Haemulidae) from Heron Island, Great Barrier Reef, Australia. Systematic Parasitology, 34, 79−87.
Chisholm, L. A., & Kritsky, D. C. (2020). Heterocotyle whittingtoni n. sp. (Monogenea: Monocotylidae) from the gills of the black-spotted whipray, Maculabatis toshi (Whitley) (Myliobatiformes: Dasyatidae), collected in coastal waters of Queensland, Australia. Systematic Parasitology, 97, 561–567.
Fricke, R., Eschmeyer, W. N., & Van der Laan, R. (eds). (2022). Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available at http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Accessed 8 March 2022.
International Commission on Zoological Nomenclature. (1999). International Code of Zoological Nomenclature, Fourth Edition. London: International Trust for Zoological Nomenclature, 306 pp.
International Commission on Zoological Nomenclature. (2012). Amendment of Articles 8, 9, 10, 22 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Bulletin of Zoological Nomenclature, 69, 161–169.
Johnson, J. W. (2010). Fishes of the Moreton Bay Marine Park and adjacent continental shelf waters, Queensland, Australia. Memoirs of the Queensland Museum - Nature, 54, 299−353.
Kritsky, D. C. (2018a). Dactylogyrids (Monogenoidea) infecting the gill lamellae of some beloniform fishes from Moreton Bay, Queensland, Australia, with a redescription of Hareocephalus thaisae Young, 1969 and descriptions of six new species of Hemirhamphiculus Bychowsky & Nagibina, 1969. Systematic Parasitology, 95, 33–54.
Kritsky, D. C. (2018b). Species of Monogenoidea infecting the gill lamellae of the common silver-biddy Gerres oyena (Forsskål) and the common silver belly Gerres subfasciatus Cuvier (Perciformes: Gerreidae) in Moreton Bay, Queensland, Australia. Systematic Parasitology, 95, 499–525.
Kritsky, D. C. (2019). Euryhaliotrema spp. (Monogenoidea: Dactylogyridae) parasitic on the gill lamellae of perciform fishes in Moreton Bay, Queensland, Australia, with the description of Euryhaliotrema solenophallus sp. nov. from the silver moony Monodactylus argenteus (Linnaeus) (Monodactylidae). Acta Parasitologica, 74, 223–227.
Kritsky, D. C., & Chisholm, L. A. (2020). Monocotylids (Monogenoidea) infecting elasmobranchs in Moreton Bay, Queensland, Australia, with descriptions of Calicotyle cutmorei n. sp. (Calicotylinae) and Dendromonocotyle raiae n.sp. (Monocotylinae). Systematic Parasitology, 97, 569–589.
Kritsky, D. C., & Nitta, M. (2019). Dactylogyrids (Platyhelminthes: Monogenoidea) infecting the gill lamellae of flatheads (Scorpaeniformes: Platycephalidae), with proposal of Platycephalotrema n. gen. and descriptions of new species from Australia and Japan. Diversity, 11, 132.
Lebedev, B. I. (1969). [Bychowskicotylinae subfam. n. and some notes on the system of monogeneans of the family Gastrocotylidae Price, 1943.] Parazitologicheskii Sbornik, 24, 156–165. (In Russian)
Lebedev, B. I. (1972a). [The taxonomy of monogeneans of Suborder Gastrocotylinea.] Trudy Biologo-Pochvennogo Instituta, Vladivostok, 11, 121–145. (In Russian)
Lebedev, B. I. (1972b). [The new family of higher monogeneans—Calyxinellidae fam. nov. (Monogenoidea).] Trudy Biologo-Pochvennogo Instituta, Vladivostok, 11, 146–153. (In Russian)
Lebedev, B. I. (1984). [System of the monogenean Suborder Gastrocotylinea.] In: Mamaev, Yu. L. (ed.), Parasites of Animals and Plants. Vladivostok: Far Eastern Scientific Center of the Academy of Sciences of the USSR, p. 3–16. (In Russian)
Lebedev, B. I. (1986). [Monogenes of the suborder Gastrocotylinea.] Leningrad: Akademia Nauk Izdatel’stvo “Nauka”, 200 pp. (In Russian)
Price, E. W. (1959). A proposed reclassification of the gastrocotyloid Monogenea. Journal of Parasitology, 45 (Suppl., Section 2), 22–23.
Yamaguti, S. (1963). Systema Helminthum. Volume IV. Monogenea and Aspidocotylea. John Wiley and Sons, New York, 699 pp.
Acknowledgments
Specimens of Diagramma labiosum were collected as part of surveys of the parasites of the fishes of Queensland, supported by the Australian Biological Resources Study (National Taxonomy Research Grants RF215-40 and RG19-37, awarded to Drs Tom Cribb and Scott Cutmore, University of Queensland). The author is especially grateful to Tom and Scott for the invitation to participate in these funded surveys, and special thanks are extended to Dr. Jean-Lou Justine (Museum National d’Histoire Naturelle, Paris) and Dr. Gabor Racz (University of Nebraska, Harold W. Manter Laboratory) for locating old versions of the ICZN for the present study.
Funding
No external funding was used in development of this paper.
Author information
Authors and Affiliations
Contributions
The author was responsible for all aspects of the submission.
Corresponding author
Ethics declarations
Competing interests
The author declare no competing interests.
Conflict of interest
The author declares that they have no conflict of interest.
Ethical approval
The author asserts that all applicable institutional, national, and international guidelines for the care and use of animals were followed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article was registered in the Official Register of Zoological Nomenclature (ZooBank) as urn:lsid:zoobank.org:pub:974FF2E1-B9D9-4FB4-B835-A38D296AEA29. This article was published as an Online First article on the online publication date shown on this page. The article should be cited by using the doi number. This is the Version of Record.
Rights and permissions
About this article

Cite this article
Kritsky, D.C. Bychowskicotylids (Monogenoidea: Gastrocotylinea) infecting the gills of the sweetlips Diagramma labiosum Macleay (Haemulidae) in Moreton Bay, Queensland, Australia, with the proposal of Lebedevia n. gen. and description of Lebedevia lebedevi n. sp.. Syst Parasitol 99, 563–569 (2022). https://doi.org/10.1007/s11230-022-10046-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11230-022-10046-4