
Abstract
Dendromonocotyle rajidicola n. sp. is described from the dorsal surface of the yellownose skate Zearaja chilensis (Guichenot) (Rajiformes) caught on the Argentine shelf. Dendromonocotyle rajidicola n. sp. can be distinguished from the other 17 species in the genus by the morphology of the distal portion of the male copulatory organ and by the unique morphology of the sclerotised proximal portion of the vagina. This is the first species of Dendromonocotyle to be described from a host in the Rajiformes and also the first record of this genus in the southwestern Atlantic Ocean.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Dendromonocotyle Hargis, 1955 (Monogenea: Monocotylidae) currently comprises 17 species. These species have been reported on the dorsal surface of stingrays (Myliobatiformes) belonging to Dasyatidae, Myliobatidae and Urotrygonidae from the Pacific and North Atlantic Oceans (Chisholm & Whittington, 1995, 2005; Chisholm et al., 2005), as well as from dasyatid and myliobatid hosts kept in aquaria in different regions of the world (Chisholm et al., 2001; Chisholm & Whittington, 2004, 2009; Vaughan et al., 2008; Vaughan & Chisholm, 2009). In the present study, specimens of Dendromonocotyle found on the yellownose skate Zearaja chilensis (Guichenot) (Rajiformes) caught during research cruises on the Argentine shelf, are described and a new species is proposed. The locality and unique host family record for this species are discussed.
Materials and methods
A total of 80 specimens of Z. chilensis was examined. Fishes were caught from October 2011 to August 2012 off the Argentine shelf during research cruises of the Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP). Fish were individually stored in plastic bags and frozen at −20°C. After thawing, fish were necropsied; the dorsal skin was washed in tap water, mucus was sieved and examined for monogeneans using a dissecting microscope with transmitted light. Monogeneans were washed in a saline solution, fixed in a 4% formaldehyde solution before being transferred to 70% ethanol for storage. Fixed monogeneans were stained with alcoholic chlorhydric carmine, dehydrated in a graded ethanol series, cleared in methyl salicylate and mounted permanently in Canada balsam. A single specimen was stained in Gomori’s trichome solution to study the sclerotised structures. Mounted worms were examined using a Leica compound microscope (model: DM 2500) equipped with differential interference contrast (DIC) optics and drawings made using an attached drawing tube. Measurements were made with the aid of an eyepiece micrometer and are given in micrometres (μm), as the range followed by the mean ± standard variation and the number of specimens measured in parentheses. The total length was measured including the haptor, width was measured at the level of the testis. The male copulatory organ (MCO) was measured following a straight line from proximal to distal extremes. The egg was measured following a straight line parallel to a side. Haptoral terminology follows that of Chisholm & Whittington (1995). The taxonomy of hosts is in accordance with Eschmeyer & Fricke (2015). Host identification was made following the key to species of skates of Cousseau et al. (2007).
Type-material was deposited in the Helminthological Collection of the Museo de La Plata, La Plata, Argentina (MLP-He). The following Dendromonocotyle specimens from the Australian Helminthological Collection (AHC) of the South Australian Museum, Adelaide, South Australia, Australia were examined for comparative purposes: D. bradsmithi Chisholm, Glennon & Whittington, 2005 (holotype; AHC 28776); D. californica Olson & Jeffries, 1983 (1 voucher; AHC 45381); D. citrosa Vaughan, Chisholm & Christison, 2008 (2 paratypes; AHC 29170 and 29171; D. colorni Chisholm, Whittington & Kearn, 2001 (2 vouchers; AHC 29172 and 29173); D. lasti Chisholm & Whittington, 2005 (holotype; AHC 28577); D. lotteri Vaughan & Chisholm, 2009 (2 paratypes; AHC 29678 and 29679); D. pipinna Chisholm & Whittington, 2004 (2 paratypes; AHC 28438 and 28439); D. torosa Chisholm & Whittington, 2004 (holotype; AHC 28537), D. ukuthena Vaughan, Chisholm & Christison, 2008 (1 paratype; AHC 29290); D. urogymni Chisholm & Whittington, 2009 (holotype; AHC 35655).
Family Monocotylidae Taschenberg, 1879
Subfamily Monocotylinae Taschenberg, 1879
Genus Dendromonocotyle Hargis, 1955
Dendromonocotyle rajidicola n. sp.
Type-host: Zearaja chilensis (Guichenot) (Rajiformes, Rajidae).
Type-locality: Southern Argentine Sea (49.28°S, 67.02°W, holotype and 3 paratypes).
Additional localities: Central and northern Argentine Sea (47.13°S, 61.13°W, 1 specimen; 40.08°S, 59.37°W, 1 specimen).
Site on host: Dorsal skin surface.
Prevalence and intensity: Three out of 80 skates infected (prevalence 3.75%) with 1, 1, 4 (mean intensity = 2) specimens, respectively.
Etymology: The specific name refers to the family Rajidae, which is here reported as host for the genus Dendromonocotyle for the first time.
Type-material: Holotype MLP-He 7102, 3 paratypes MLP-He 7103.
Description (Figs. 1–5)
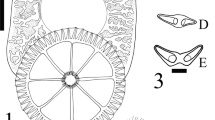
[Based on 5 sexually mature specimens, one additional juvenile specimen examined to investigate male copulatory organ (MCO) development.] Body 3,320–6,880 (4,720 ± 1,394; n = 5) long, 1,500–3,760 (2,476 ± 881; n = 5) wide (Fig. 1A). Haptor diameter 1,570–3,020 (2,036 ± 583; n = 5), divided into 1 central and 8 peripheral loculi (Fig. 1A). Hamuli absent. Hooklets 14, distributed regularly in thicker portion of marginal valve between every 4 papillae (Fig. 1A, B). Haptoral rim with 56 marginal papillae: 6 papillae associated with each anterior loculus; 7 papillae associated with each posterolateral and anterolateral loculus; and 8 papillae associated with each posterior loculus (Fig. 1A). Marginal papillae with 9–13 sclerites (mode = 10) (Figs. 1A, 5A), terminal papillary sclerite distinct (Figs. 2A, 5B) from other papillary sclerites (Fig. 2B). Radial septa with 26–32 sclerites (mode = 29), inner ring septa with 76–85 sclerites, outer ring septa with 26–41 sclerites per loculus; sclerites more numerous on posterior loculi decreasing in number anteriorly. Sclerites on radial, inner and outer ring septa similar in shape (Fig. 2C, E–G). Tripartite sclerites sometimes present at junction of radial septum with inner or outer ring septum (Fig. 2D). Mouth ventral, subterminal. Five anterolateral gland duct openings on either side of head (Fig. 1A). Subterminal groove absent at anterior end. Eyespots 2 pairs, anterodorsal to pharynx; anterior pair very close to posterior pair. Pharynx 535–1,450 (912 ± 311; n = 5) in diameter. Intestinal diverticula extend from just posterior to level of mouth to posterior margins of body proper (Fig. 1A). Dark pigment associated with lining of intestinal diverticula present in all specimens collected, including juvenile.

Dendromonocotyle rajidicola n. sp. A, Ventral view, composite; B, Marginal hooklet; C, Egg. Abbreviations: alg, anterolateral gland duct openings; e, eyespot; i, intestinal caecum; m, mouth; mh, marginal hooklet; mp, marginal papilla; p, pharynx; tps, terminal papillary sclerite. Scale-bars: A, 500 μm; B, 5 μm; C, 100 μm

Haptoral sclerites of Dendromonocotyle rajidicola n. sp. A, Terminal papillary sclerite; B, Papillary sclerite; C, D, Radial septa sclerites; E, F, Inner ring septa sclerites; G, outer ring septa sclerite. Scale-bars: A, 10 μm; B–G, 5 μm
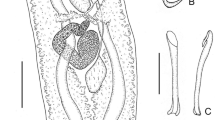
Testis single 490–710 (628 ± 105; n = 5) wide. Vas deferens arises from left side of testis, runs anteriorly dorsal to vagina, then curves to right side of body, ventral to ejaculatory bulb inflating to form a looping seminal vesicle, then narrows, looping around male copulatory organ before entering ejaculatory bulb posteriorly (Fig. 3). Ejaculatory bulb 228–320 (267 ± 41; n = 5) long, 175–260 (225 ± 34; n = 5) wide. Male accessory glands entering base of ejaculatory bulb not seen. Male copulatory organ sclerotised tube 1,037–1,135 (1,096 ± 36; n = 5) long, 18–20 (19 ± 1; n = 5) wide; surrounded by muscular sheath (Figs. 1A, 3). Male copulatory organ with a single sperm duct opening terminally. Distal end of male copulatory organ armed with 3 sclerotised accessory flanges and a coiled accessory filament as illustrated (Figs. 4B, C, 5C). In single juvenile specimen studied, proximal end of male copulatory organ looped (Fig. 5D) and accessory filament and flanges not seen at distal end.

Reproductive system of Dendromonocotyle rajidicola n. sp. in ventral view. Abbreviations: dmco, distal portion of male copulatory organ; eb, ejaculatory bulb; eg, egg; mco, male copulatory organ; mg, Mehlis’ gland; ms, muscular sheath surrounding male copulatory organ; o, ovary; ob, ovarian branch; oo, oötype; spv, sclerotised proximal portion of vagina; sr, seminal receptacle; sv, seminal vesicle; t, testis; tvd, transverse vitelline duct; v, vagina; vd, vas deferens; vp, vaginal pore. Scale-bar: 250 μm

Dendromonocotyle rajidicola n. sp. A, Sclerotised proximal portion of vagina; B, Distal portion of male copulatory organ; C, Details of male copulatory organ, showing sclerotised accessory flanges, accessory filament omitted. Abbreviations: af, accessory filament; ed, ejaculatory duct; fl, flanges. Scale-bars: A, B, 100 μm; C, 50 μm

Photomicrographs of Dendromonocotyle rajidicola n. sp. A, Papillary sclerites; B, Terminal papillary sclerite; C, Distal portion of male copulatory organ; D, Proximal portion of male copulatory organ in juvenile specimen; E, Sclerotised proximal portion of vagina. Scale-bars: A, C, D, E, 50 μm; B, 10 μm
Ovary bi-lobed, with single branch on either side of testis. Anteromedial extension loops right intestinal caecum dorsoventrally and forms oviduct (Fig. 3). Oviduct receives common vitelline duct and duct from seminal receptacle and forms ovovitelline duct which enters oötype posteriorly. Oötype 250–888 (468 ± 249; n = 5) long, 63–163 (125 ± 37; n = 5) wide. Mehlis’ gland present. Vaginal pore ventral, on left side of body at level of posterior portion of pharynx. Proximal part of vagina sclerotised 313–338 (327 ± 10; n = 5) (Figs. 3, 4A, 5E); vagina then narrows and enters seminal receptacle 125–213 (148 ± 43; n = 4) long, 125–185 (144 ± 27; n = 4) wide (Fig. 3).
Vitellarium inconspicuous, largely obscured by extensive pigmented intestinal diverticula; extends from level of pharynx to posterior portion of body proper. Transverse vitelline duct at level of anterior portion of seminal receptacle (Fig. 3); common vitelline duct joins oviduct. Egg tetrahedral, side length 125–138 (131 ± 9; n = 2), with short appendage on 1 pole (Fig. 1C).
Remarks
The majority of Dendromonocotyle spp. including D. rajidicola n. sp. have 56 marginal papillae. The exceptions are D. ardea Chisholm & Whittington 1995 and D. taeniurae Euzet & Maillard 1967, which both have 42, and D. lasti which has 38 (Vaughan et al., 2008). Among the members of Dendromonocotyle with 56 marginal papillae, species can be separated into two distinct groups based on the papillae-to-loculus associations (Vaughan et al., 2008). One group includes D. colorni and D. lotteri with papillae-to-loculus associations as follows: anterior and anterolateral loculus pairs with six associated papillae, posterolateral and posterior loculus pairs associated with eight papillae each, represented numerically as 6-6-8-8. The other group has the anteriormost loculi each with six associated papillae, the anterolateral and posterolateral loculi with seven associated papillae each, and the two posterior loculi with eight associated papillae, represented numerically as 6-7-7-8. This group includes our new species D. rajidicola along with D. akajeii Ho & Perkins, 1980, D. bradsmithi, D. californica, D. centrourae Cheung & Whitaker, 1993, D. citrosa, D. cortesi Bravo-Hollis, 1969, D. kuhlii Young, 1967, D. octodiscus Hargis, 1955, D. pipinna, D. torosa, D. ukuthena and D. urogymni. Dendromonocotyle rajidicola n. sp. can be distinguished from the other 12 members of this group by the morphology of the distal portion of the male copulatory organ and the number of radial and papillary sclerites. Additionally, the morphology of sclerotised proximal portion of the vagina in D. rajidicola is unique.
Discussion
With the description of D. rajidicola n. sp. there are now 18 species in the genus. The first species of Dendromonocotyle were reported from stingrays caught in natural habitats, including D. akajeii, D. ardea, D. californica, D. centrourae, D. cortesi, D. kuhlii, D. octodiscus, D. taeniurae (see Chisholm & Whittington, 1995; Pulido-Flores & Monks, 2005). The majority of species that have been described more recently are from hosts kept in public aquaria. These include D. citrosa, D. colorni, D. lotteri, D. pipinna, D. torosa, D. ukuthena and D. urogymni (see Chisholm & Whittington, 1995, 2009; Chisholm et al., 2001, 2004; Vaughan et al., 2008).
The development of the male copulatory organ in species of Dendromoncotyle has been investigated previously. Chisholm & Whittington (1995) showed that in D. ardea, the sclerotised filaments associated with distal portion of the male copulatory organ were absent in juveniles and formed as the parasite matured. They also found that the proximal portion of the long male copulatory organ is coiled in juveniles presumably so it can be accommodated in the shorter body but then straightens as the body of the parasite increases in size. We made similar observations in D. rajidicola where the coiled filament seen around the distal portion of the male copulatory organ in the adults is absent in the juvenile and the proximal part of the male copulatory organ is looped in the juvenile specimen but is straight in the adults. The male copulatory organ of D. colorni (see Chisholm et al., 2001) and D. bradsmithi (see Chisholm et al., 2005) also straightens as the parasite develops and this possibly the case for all Dendromonocotyle species that have a long male copulatory organ.
We only found D. rajidicola n. sp. on three of the 80 specimens of Z. chilensis caught off the Argentine shelf. The low prevalence (3.75%) and intensity we found is consistent with the observations that the intensities of monogeneans on wild elasmobranchs are generally lower than in those kept in captivity. In captive environments monogenean intensities can increase significantly due to their direct life-cycle and the hosts reduced ability to fight infection due to stress (see Chisholm et al., 2004). Lesions which may form where the parasites attach can result in the death of the elasmobranch either indirectly (via secondary infections) or directly (Chisholm et al., 2004; Poynton et al., 1997; Bullard et al., 2001).
Dendromonocotyle spp. that have been described from wild hosts, have been reported from stingrays caught in the south Pacific Ocean (D. bradsmithi, D. lasti and D. kuhlii), north Pacific Ocean (D. akajeii and D. californica) and north Atlantic Ocean (D. cortesi, D. octodiscus and D. taeniurae). This is the first report of a species of Dendromonocotyle from the southwestern Atlantic Ocean.
All previously described species of Dendromonocotyle, are parasites of Myliobatiformes, with most of them (D. akajeii, D. ardea, D. centrourae, D. citrosa, D. colorni, D. kuhlii, D. lasti, D. lotteri, D. octodiscus, D. pipinna, D. taeniurae, D. ukuthena and D. urogymni) found on hosts in the Dasyatidae. A few others have been found on hosts in the Myliobatidae (D. bradsmithi, D. californica and D. torosa) and on the Urotrygonidae (D. octodiscus). The present finding constitutes the first record of Dendromonocotyle species on a member of the Rajiformes, namely, Zearaja chilensis (Rajidae). This skate species lives in deep waters of austral South-America and has been little studied for parasitological purposes. At present, only one monogenean species, Merizocotyle euzetii Irigoitia, Cantatore, Delpiani, Incorvaia, Lanfranchi & Timi, 2014 has been recorded on nasal tissue of this skate species (Irigoitia et al., 2014). A high diversity of skates, approximately 30 species, inhabit the Argentine Sea (Menni & Lucifora, 2007). Little is known about their monogenean fauna and therefore it is expected that many new species, including species of Dendomonocotyle remain to be described in this region.
References
Bullard, S. A., Frasca, S, Jr, & Benz, G. W. (2001). Gill lesions associated with Erpocotyle tiburonis (Monogenea: Hexabothriidae) on wild and aquarium-held bonnethead sharks (Sphyrna tiburo). Journal of Parasitology, 87, 972–977.
Chisholm, L. A., & Whittington, I. D. (1995). A revision of Dendromonocotyle Hargis, 1955 (Monogenea: Monocotylidae) with a description of a new species from Pastinachus sephen (Forsskål) (Myliobatiformes: Dasyatidae) from the Great Barrier Reef, Australia. Journal of Natural History, 29, 1093–1120.
Chisholm, L. A., & Whittington, I. D. (2004). Two new species of Dendromonocotyle Hargis, 1955 (Monogenea: Monocotylidae) from the skin of Taeniura meyeni (Dasyatidae) and Aetobatus narinari (Myliobatidae) from aquaria in Queensland, Australia. Systematic Parasitology, 57, 221–228.
Chisholm, L. A., & Whittington, I. D. (2005). Dendromonocotyle lasti n. sp. from the skin and Monocotyle caseyae n. sp. (Monogenea: Monocotylidae) from the gills of Himantura sp. (Dasyatidae) in Moreton Bay, Queensland, Australia. Systematic Parasitology, 60, 81–89.
Chisholm, L. A., & Whittington, I. D. (2009). Dendromonocotyle urogymni sp. nov. (Monogenea, Monocotylidae) from Urogymnus asperrimus (Elasmobranchii, Dasyatidae) off eastern Australia. Acta Parasitologica, 54, 113–118.
Chisholm, L. A., Whittington, I. D., & Kearn, G. C. (2001). Dendromonocotyle colorni sp. n. (Monogenea: Monocotylidae) from the skin of Himantura uarnak (Dasyatidae) from Israel and a new host record for D. octodiscus Hargis, 1955 from the Bahamas. Folia Parasitologica, 48, 15–20.
Chisholm, L. A., Whittington, I. D., & Fischer, A. D. P. (2004). A review of Dendromonocotyle (Monogenea: Monocotylidae) from the skin of stingrays and their control in public aquaria. Folia Parasitologica, 51, 123–130.
Chisholm, L. A., Glennon, V., & Whittington, I. D. (2005). Dendromonocotyle bradsmithi n. sp. (Monogenea: Monocotylidae) from the skin of Myliobatis australis (Elasmobranchii: Myliobatidae) off Adelaide and Perth, Australia: description of adult and larva. Zootaxa, 951, 1–15.
Cousseau, M. B., Figueroa, D. E., Díaz de Astarloa, J. M., Mabragaña, E., & Lucifora, L. O. (2007). Rayas, chuchos y otros batoideos del Atlántico Sudoccidental (34°S–55°S). Publicaciones Especiales Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP), Mar del Plata. 104 pp.
Eschmeyer, W. N., & Fricke, R. (2015). Catalog of fishes: Genera, species, references. Retrieved October 14, 2015, from (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp).
Irigoitia, M. M., Cantatore, D. M., Delpiani, G. E., Incorvaia, I. S., Lanfranchi, A. L., & Timi, J. T. (2014). Merizocotyle euzeti sp. n. (Monogenea: Monocotylidae) from the nasal tissue of three deep sea skates (Rajidae) in the southwestern Atlantic Ocean. Folia Parasitologica, 61, 206–212.
Menni, R. C., & Lucifora, L. O. (2007). Condrictios de la Argentina y Uruguay. Lista de Trabajo ProBiota, FCNyM, UNLP, Serie Técnica-Didáctica, La Plata, Argentina, 11, 1–15.
Poynton, S. L., Campbell, T. W., & Palm, H. W. (1997). Skin lesions in captive lemon sharks Negaprion brevirostris (Carcharhinidae) associated with the monogenean Neodermophthirius harkemai Price, 1963 (Microbothriidae). Diseases of Aquatic Organisms, 31, 29–33.
Pulido-Flores, G., & Monks, S. (2005). Monogenean parasites of some elasmobranchs (Chondrichthyes) from the Yucatán Peninsula, Mexico. Comparative Parasitology, 72, 69–74.
Vaughan, D. B., & Chisholm, L. A. (2009). Three Dendromonocotyle species (Monogenea: Monocotylidae) reported from captive rays, including D. lotteri sp. n. from Himantura gerrardi (Elasmobranchiii: Dasyatidae) in the public aquarium at the Atlantis resort, Dubai. Folia Parasitologica, 56, 99–106.
Vaughan, D. B., Chisholm, L. A., & Christison, K. (2008). Overview of South African Dendromonocotyle (Monogenea: Monocotylidae), with descriptions of 2 new species from stingrays (Dasyatidae) kept in public aquaria. Zootaxa, 1826, 26–44.
Acknowledgements
Thanks are extended to the team of the Programa de Condrictios (INIDEP) for assisting in the collection of fish samples; to Dr Santiago Barbini (IIMyC), Dr Lorena Scenna (IIMyC) and Dr Daniel Figueroa (UNMdP) for providing literature about chondrichthyans.
Funding
Financial support was provided by grants from CONICET (PIP # 112-201101-00036) and ANPCYT (PICT 2012 # 02094).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable institutional, national and international guidelines for the care and use of animals were followed.
Rights and permissions
About this article

Cite this article
Irigoitia, M.M., Chisholm, L.A. & Timi, J.T. A new species of Dendromonocotyle Hargis, 1955 (Monogenea: Monocotylidae) from the skin of Zearaja chilensis (Guichenot) (Rajiformes: Rajidae) from the Argentine Sea. Syst Parasitol 93, 367–374 (2016). https://doi.org/10.1007/s11230-016-9624-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11230-016-9624-1