
Abstract
Fish behavior is important to consider when developing selective fishing gear. In studies designed to investigate the size selective properties of towed fishing gears such as trawls, fish behavior is mainly documented by underwater video recordings. Because fishing gear can be operated at great depths or in other low light environments, artificial light is often required for underwater recordings. However, artificial light can influence fish behavior, which casts doubt on the validity of behavioral observations obtained in the presence of artificial light. However, removing artificial light disables video recordings and the possibility to study fish behavior in relation to selectivity devices towed fishing gears in low light environments. To date, little is known about the extent to which artificial light used for video observations affects fish behavior with respect to fishing gear. Therefore, we conducted fishing trials in the Barents Sea demersal trawl fishery to assess the effect of light sources on fish behavior by using size selectivity results in towed fishing gears. We found that the behavior of cod (Gadus morhua) was unaffected by the light sources, whereas the behavior of haddock (Melanogrammus aeglefinus), saithe (Pollachius virens) and redfish (Sebastes spp.) significantly changed when red light and white light were employed. Our results also demonstrated significant differences in fish behavior between white and red light.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Size and species selectivity in towed fishing gear have been studied for decades to address the issue of unwanted by-catch (Kennelly and Broadhurst 2021). Size selectivity studies normally provide information about the probability that a given fish species will be retained or escape depending on its length, which is estimated based on length measurements of retained and escaped fish collected during sea trials (Wileman et al. 1996). However, size selectivity estimation does not provide any direct information about the behavior of fish, which can be decisive for the fate of fish in the gear (i.e., retention or escape). Information about fish behavior in relation to selectivity is important because in many cases it can explain patterns in size selectivity curves that quantify the length-dependent retention probability (Wileman et al. 1996). Additionally, the knowledge obtained can be applied to the development of selective fishing gear.
Fish behavior has been observed using a variety of direct methods, including observation by humans (Hemmings 1973), with video cameras and hydro-acoustics (Urquhart and Stewart 1993; Graham et al. 2004). Video cameras are the most common direct method to study animal behavior in relation to fishing gear. However, many towed fishing gears are operated in light environments that are below the sensitivity of optical cameras (e.g., at great depths or at night) (Karlsen et al. 2021). In such circumstances, artificial light is required to illuminate the field of view.
Many studies have demonstrated that artificial light can affect fish behavior (Weinberg and Munro 1999; Gordon et al. 2002; Marchesan et al. 2005). In selectivity studies, researchers have used artificial light to purposely affect fish behavior and reduce unwanted by-catch (Hannah et al. 2015; Grimaldo et al. 2018; Lomeli et al. 2018a, b, 2019; Lomeli and Wakefield 2019; Yochum et al. 2022). Even mechanically stimulated bioluminescence can affect the behavior of fish in front of a trawl (Jamieson et al. 2006). The influence of artificial light is highly dependent on the species (Grimaldo et al. 2018; Geraci et al. 2022), the characteristics of the light (especially wavelength and light intensity) (Marchesan et al. 2005; Mueller et al. 2006; Yochum et al. 2022), and the context in which the light is deployed (Hannah et al. 2015). Thus, when artificial light is used to observe fish behavior, gear selectivity may be affected. However, it would not be possible to observe fish behavior in relation to fishing gear in deep waters using optic cameras without artificial light.
In the Barents Sea demersal trawl fishery that targets gadoids, selectivity studies have been conducted for several decades (Kennelly and Broadhurst 2021), especially since the introduction of the sorting grid in 1997 (Isaksen et al. 1992; Larsen and Isaksen 1993). In this fishery a grid with a 55 mm bar spacing is mandatory. Cod (Gadus morhua), haddock (Melanogrammus aeglefinus), and saithe (Pollachius virens) are the main target species, and redfish (Sebastes spp.) are among the main bycatch species. The size selectivity of these species, especially cod and haddock, has been studied extensively (e.g., Sistiaga et al. 2016; 2022; Grimaldo et al. 2018; Brinkhof et al. 2020; 2022). Fishing depths are ranging between 80 and 400 m, and the main fishery is conducted during winter when there is no sunlight. Red light has been used in some studies based on the assumption that red light does not affect fish behavior (Grimaldo et al. 2018; Underwood et al. 2021) because it is believed to be outside the wavelength sensitivity range of many species of fish (Goldsmith and Fernandez 1968; Meyer-Rochow and Tiang 1984; Douglas, et al. 1995). However, the potential for making biased conclusions about fish behavior when exposed to red light has not been investigated. As the visual sensitivity of many commercial species is poorly known and many sources of red light are not monochromatic (i.e., they contain a range of wavelengths), a response to red light in the fishery cannot be excluded.
Based on this lack of knowledge, the aim of this study was to investigate if, and to what extent, different wavelengths of artificial light used for underwater recordings unintentionally affect fish behavior in towed fishing gear such as bottom trawls. Specifically, the following research questions were addressed.
-
(i)
Does artificial light used for underwater observations in a sorting grid section affect the behavior of cod, haddock, saithe, and redfish?
-
(ii)
Do fish exhibit differences in behavior when subjected to red or white light in a sorting grid section?
-
(iii)
Is the effect of artificial light different for cod, haddock, saithe, and redfish?
Materials and methods
Assessment method
The effect of a treatment is often evaluated by comparing the results of samples to which the treatment is applied with the results of untreated samples (control), keeping the rest of the experimental conditions equal. The aim of this study was to investigate whether artificial light used for underwater recordings unintentionally affected fish behavior in trawls and thus biased potential behavioral analyses based on video observations. Acoustic methods, which are often used to observe fish in low light or turbid environments, could be an alternative, but they do not provide the necessary resolution to carry out such a comparison. Fish behavior and fish morphology are the two fish-specific factors that can affect selectivity in fishing gear. A straightforward experimental design to test the effect of artificial light on fish behavior would be to directly observe and compare fish behavior in the trawl with and without light. However, the effect of light increases its relevance in darkness beyond the sensitivity of the optic camera deployed, and in the dark fish can only be observed in detail using lights.
It is reasonable to assume that light cannot change fish morphology or the physical properties of a gear. Consequently, any potential change in the size selective properties of the gear resulting from the application of light would originate from the potential effect of light on fish behavior. Therefore, we applied an indirect method to assess the impact of artificial light on fish behavior (Fig. 1). We used a sorting grid that is mandatory in the bottom trawl fishery for gadoids in the Barents Sea and assessed its size selective properties with and without artificial light (i.e., the treatment). The data were then to determine if there were significant differences in size selectivity, which would prove that the specific lights used for fish observation in this study affected the behavior of fish and vice versa (Fig. 1).

Schematic overview of the assessment method used to determine if fish behavior related to size selectivity is affected by artificial light
Sea trials
The experimental fishing trials were conducted onboard the R/V “Helmer Hanssen” (63.8 m length and 4,080 HP engine power) in the southern Barents Sea off the coast of Norway from February 26 to March 5, 2022. A single Alfredo No. 3 bottom trawl was used during the trials. The otter boards (Injector Scorpion, 3,100 kg, and 8 m2 each) were followed by 3 m long backstraps and 7 m long connector chains, which were connected to the 60 m long sweeps. To avoid abrasion of the sweeps, a Ø53 cm steel bobbin was placed mid-way between the connector chain and the attachment to the ground gear. The ground gear was 46 m long and consisted of an 18.9 m long rockhopper gear (Ø 53 cm) in the middle and a 14 m long chain (Ø19 mm) with three equally spaced steel bobbins (Ø 53 cm) on each side. The rockhopper gear was attached to the 19.2 m long fishing line of the trawl. The Alfredo No. 3 trawl is a two-panel trawl with 420 meshes in circumference, and it was built entirely out of polyethylene with a mesh size of 155 mm.
After the trawl belly, a Sort-V grid section containing a 1,750 mm × 1,234 mm steel grid with a bar spacing of 55.05 ± 0.84 mm (mean ± SD) was mounted in the trawl. To retain the escapees the grid outlet was fitted with a cover, which had an inner mesh size of 44.32 ± 1.06 mm (Fig. 2). A 10 m long extension piece was inserted between the grid section and the codend. The 12 m long codend was constructed of two panels. It had 60 meshes of single braided Ø8 mm polyethylene twine in the circumference. The codend was blinded with a small-meshed liner with a mesh size of 49.70 ± 4.99 mm to stop fish from escaping from it. The meshes in the codend, cover, small-meshed liner, and grid bar spacing were measured using an OMEGA gauge following the procedure described in Wileman et al. (1996). For the hauls in which artificial light was used, either a white or red dive torch (Brinyte DIV01V, 120° light beam angle) was mounted in front of the grid. The light beam was directed towards the grid, and it illuminated the entire grid surface (Fig. 2).

Trawl rigging showing the experimental configuration with the Sort-V grid, grid cover (GC), blinded codend with a small-meshed liner (C), and the positioning and direction of the video camera and lights (when used)
The scalar irradiance spectrum of the two lights was measured in air for each 3.3 nm (Karlsen et al. 2021). The measurements were conducted in a dark room using a hyperspectral radiometer (Ramses ASC VIS, TriOS GmbH, Rastede, Germany) placed 30 cm away from the light source. The sensitivity of the sensor was 6 × 10–4 mW m–2 nm–1 at 500 nm (corresponding to 2.5 × 10–6 μmol m–2 s–1).
During towing, the trawl performance and geometry were continuously monitored by a set of Scanmar sensors. Parameters measured were the otter board spread, trawl height, and catch volume. When the trawl was hauled on board the vessel, the catch from the codend and the grid cover were kept in separate holding bins. The length of all cod, haddock, saithe, and redfish above 20 cm were measured to the nearest centimeter below.
Statistical analysis
Analysis of grid size selection data
The analysis of the data collected for each species was performed separately and independently for the control (no light) and the two different treatments (red light or white light). In the experimental design applied, fish were either retained by the grid cover or by the blinded codend, so the collected catch data were binominal.
The retention probability for fish of length l in the blinded codend in haul j is expressed by a function rj(l). The purpose of the analysis was to estimate the values of this function for all relevant sizes of the species investigated individually. Between hauls with the same treatment, the value of rj(l) is expected to vary (Fryer 1991). We were interested in the length-dependent values of r(l) averaged over hauls to provide information about the average consequences for the size selection process of applying the specific treatment (or the control). The estimation of the average size selection \({r}_{av}\left(l,{\varvec{v}}\right)\) involved pooling data from the hauls conducted with a specific treatment (Herrmann et al. 2012). The \({\varvec{v}}\) is a vector consisting of the parameters of the model. Four different models (Logistic, Probit, Gompertz, and Richards) were chosen as basic candidates to describe \({r}_{av}\left(l,{\varvec{v}}\right)\) for the baseline/control, the two treatments, and the species individually. The first three models were fully described by the selection parameters length at 50% retention probability (L50) and selection range (SR), whereas the Richards model required an additional parameter (δ). The formulas for the four selection models, together with additional information, can be found in Wileman et al. (1996). These four classical size-selection models assume that all individual fish entering the grid section can contact the grid in a way that provides a size-dependent probability of escape into the cover. In addition, four corresponding models, which accounted for the potential fraction of fish that could not contact the grid to provide a size-dependent chance of escape, were also considered. In total, eight models were tested for each of the data series \({r}_{av}\left(l,{\varvec{v}}\right)\):
For the four last models in (1), C represents the assumed length-independent contact probability with the grid that provides fish with a length-dependent chance of escape. Thus, C is a number between 0.0 and 1.0. If all fish were able to contact the grid sufficiently to obtain a length-dependent chance of escape, C would be 1.0 and the last four models in (1) would simplify to the first four. In contrast, if only 75% of fish were able to make sufficient contact with the grid, C would be 0.75 (see Herrmann et al. (2013) for further details about this type of size selection model). The subscript g on the selection parameters (L50, SR, 1/δ) in the last four models in (1) indicates that these are values of the parameters for the fraction of the fish that contact the grid. In those models, the overall L50 and SR also consider the fraction of fish not making sufficient contact with the grid, and they are estimated based on the value of the subscribed parameters and the estimated value of C obtained by the approach described in Sistiaga et al. (2010). The reason we included the last four models in (1) as candidates to describe the size selection in a grid section of a bottom fishery is that several studies have shown that often a significant fraction of the fish entering such a section do not make contact with the grid in a way that provides them with a length-dependent chance to escape (Sistiaga et al. 2010; Herrmann et al. 2019; Brinkhof et al. 2022).
The selection curves described by the formulas in (1) and the associated selectivity parameters were estimated using the maximum likelihood method and by minimizing the negative of the log-likelihood function derived from the binomial probability mass function (Wileman et al. 1996):
The log-likelihood function (2) includes a summation over hauls {j = 1,…,m} for the specific treatment or the control, with ncjl and ngjl being the number of fish of the specific species caught in the codend and grid, respectively, in haul j belonging to length class l.
Our evaluation of the ability of a model to describe the data sufficiently was based on a calculation of the corresponding p-value, which expresses the likelihood of obtaining at least as big a discrepancy between the fitted model and the observed experimental data by coincidence. Therefore, for the fitted model to be a candidate to model the size selection data, this p-value should not be < 0.05 (Wileman et al. 1996). The best model among the eight considered in (1) was selected based on Akaike’s Information Criterion (AIC); the model with the lowest AIC value was selected (Akaike 1974).
Often, once the specific selection model is identified for a particular species and treatment (or control), bootstrapping is applied to estimate the confidence limits for the average size selection keeping the model fixed to the one selected based on the data collected. However, this procedure does not account for uncertainty in model selection due to data fluctuations (collected data being a random sample) (Burnham and Anderson 2002; Efron 2014; Preacher and Merkle 2012). Therefore, to account for this additional uncertainty for the size selection curves we adapted the bootstrap model selection (BMS) approach described by Jacques et al. (2024). Specially, this implied conducting model selection among the eight size selection models in (1) in each of the 1000 bootstrap iterations carried out. Specially, in each bootstrap iteration independently the model having the lowest AIC value for the specific resampled data was selected to model that data. The resampling procedure in this bootstrapping approach was identical to the one described in Millar (1993) and took both within-haul and between-haul variation into consideration. The hauls for the control and two treatments were used to define a group of hauls. To account for between-haul variation, an outer bootstrap resample with replacement from the group of hauls was included in the procedure. Within each resampled haul, the data for each length class were bootstrapped in an inner bootstrap to account for within-haul variation. Each bootstrap iteration resulted in a “pooled” set of data, which then was analysed performing the model selection among the eight candidates in (1). Thus, each bootstrap run resulted in an average selection curve. For each case analysed (specific species for specific treatment or control), 1,000 bootstrap repetitions were conducted to estimate the Efron percentile 95% confidence intervals (CIs) (Herrmann et al. 2012). We used the software tool SELNET (Herrmann et al. 2012) for the size selection analysis (version date 31 May 2024).
Inference of the effect of treatment on fish behavior
To investigate the extent to which artificial light affects behavior and thereby the size selection in the grid section, changes in the length-dependent retention probability were quantified (delta). Specifically, the difference in \({r}_{av}\left(l,{\varvec{v}}\right)\) between treatment (t) or control (q) was obtained species-wise by estimating the difference in the length-dependent retention probability (\(\Delta {r}_{{av}_{qt}}\left(l\right)={r}_{{av}_{t}}\left(l,{{\varvec{v}}}_{{\varvec{t}}}\right)-{r}_{{av}_{q}}\left(l,{{\varvec{v}}}_{{\varvec{q}}}\right)\)), where \({r}_{{av}_{q}}\left(l,{\varvec{v}}\right)\) and \({r}_{{av}_{t}}\left(l,{\varvec{v}}\right)\) represent \({r}_{av}\left(l,{\varvec{v}}\right)\) obtained using (1) in (2) for control and treatment, respectively. The 95% CIs for (\(\Delta {r}_{{av}_{qt}}\left(l\right)\) were obtained based on the two bootstrap populations for both \({r}_{{av}_{t}}\left(l,{{\varvec{v}}}_{{\varvec{t}}}\right)\) and \({r}_{{av}_{q}}\left(l,{{\varvec{v}}}_{{\varvec{q}}}\right)\) using the method described in Larsen et al. (2018). In cases of length classes for which the 95% CIs did not contain the value 0.0, we concluded that there was a difference in fish behavior between a treatment t and the control q or between two treatments. In contrast, if the 95% CIs for all fish sizes included the value 0.0, we concluded that the treatment had no effect on fish behavior.
Results
Light source measurements
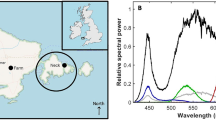
The energy of the emitted white light peaked at 443 nm (full-with-half-max, FWHM, range: 430–453 nm) and 547 nm (FWHM range: 503–627 nm), while that of the emitted red light peaked at 630 nm (FWHM range: 620–640 nm) (Fig. 3).

Light spectra of the white (left) and red (right) light sources measured as scalar irradiance at 30 cm with the center of the light beam pointing directly at the light sensor (0°, dotted line), tilted 45° relative to the sensor (45°, broken line), or perpendicular to the sensor (90°, solid line)
Results from sea trials
During the cruise, we conducted 39 valid hauls consisting of 14 hauls without artificial light, 14 hauls with white light, and 11 hauls with red light (Table 1). The towing depth ranged between 236 and 315 m, and towing time ranged from 35 min to 1 h 45 min. The total length of 13,153 cod, 7497 haddock, 2830 saithe, and 1218 redfish were measured during the cruise. If fewer than 15 individuals of one species were caught during a haul, it was eliminated from the statistical analysis for that species (Table 1).
Each of the models in Eq. (1) where fitted to the experimental data model for each species and light case (without artificial light, white light, red light) separately to conduct model selection for each case according to lowest AIC value (Table 2). The models chosen to quantify the retention probability for cod, haddock, saithe, and redfish without the use of artificial light or when exposed to white or red light fitted the experimental data points well (p-value > 0.05) (Fig. 4, Table 2.) The distribution curves in Fig. 4 showed that few cod were retained in the cover, probably because there were few individuals in the area that were small enough to pass through the grid bars. For haddock, saithe, and redfish, however, a considerable proportion managed to escape and was retained in the grid cover.

Retention probability curves for cod, haddock, saithe, and redfish (black solid curves) with 95% CIs (blue, yellow and red shaded areas). The black dots denote the experimental data points, and the grey and black distribution curves denote the number of fish caught in each length class in the codend and grid cover, respectively
When comparing the effect of no light, white light, and red light for the four species, no significant effect of light was detected for cod (Fig. 5). However, red and white light caused a significantly higher retention probability of haddock in the codend compared to the control without light for a large span of length groups (Fig. 5). The same patterns were detected for saithe, but only for a few length groups of the largest individuals. For redfish, significantly more of the largest length groups (> 48 cm) were retained in the white light treatment compared to the hauls conducted without light or with red light (Fig. 5).

Comparison of the retention probability curves for cod, haddock, saithe, and redfish when caught with no light (solid black curves), white light (dashed black curves), and red light (dotted black curves), with 95% CIs (colored areas)
The delta plots comparing the hauls conducted without light to the hauls conducted with white light demonstrated no significant effect for cod, whereas white light resulted in a significantly higher retention of haddock, saithe, and redfish (Fig. 6). Comparing the hauls conducted without light to the hauls conducted with red light revealed similar results. For redfish, a significant difference was only detected for a few length groups, with the hauls conducted with red light having a slightly higher retention efficiency compared to the hauls conducted without light (Fig. 6). The delta plots comparing the hauls conducted with white light to those conducted with red light demonstrated no significant difference for cod and haddock. For saithe and redfish, however, the hauls with white light retained significantly more of the largest fish compared to the hauls with red light (Fig. 6).

Delta curves (black curves) represent the differences in selection properties for cod, haddock, saithe, and redfish when caught with no light, white light, and red light, with 95% CIs (grey shaded areas). Horizontal dashed lines denote the line of no difference in selectivity
For the selectivity parameters L50 and SR, no significant difference was detected for cod, saithe, and redfish for no light vs. white light, no light vs. red light, or white light vs. red light (Table 2). For haddock, however, a significant reduction in the L50 and increase in the SR was estimated between the hauls conducted with no light compared to those conducted with either white or red light (Table 2). No significant difference was detected between the hauls conducted with white or red light for haddock.
Discussion
In selectivity studies, fish behavior is often an important explanatory factor. Therefore, besides the regular catch measurements necessary to evaluate the selective performance of fishing gear, video recordings are often conducted to investigate the relationship between fish behavior and gear performance (e.g., Sistiaga et al. 2016; Grimaldo et al. 2018; Brinkhof et al. 2020; Santos et al. 2020). In many cases, such video recordings are conducted in environments where the light conditions are below the sensitivity levels of optical cameras (e.g., at great depths or at night) (Karlsen et al. 2021) and require the use of artificial light to illuminate the field of view. Whether and to what extent different types of artificial light affect the behavior of fish has been a long and ongoing debate, because this potential effect could affect the size selectivity performance of fishing gear. It is also commonly claimed in the literature that the effect of red light on fish behavior is limited (Sistiaga et al. 2017; Grimaldo et al. 2018). Therefore, the purpose of this study was to assess whether artificial light used for underwater observations in towed fishing gears affects fish behavior and therefore risks providing biased information about the behavior of fish. Specifically, we tested whether there is a potential effect on fish behavior when exposed to different sources of light, and whether the effects differ among cod, haddock, saithe, and redfish.
The results of this study demonstrated that artificial light used for underwater observations affected fish behavior and consequently the selective performance of fishing gear. They also demonstrated that the impact of light depended on both the light color used and the traits of the species exposed to light. Melli et al. (2018) and O’Neill et al. (2022) previously documented the effect of artificial light on the behavior of cod. Other studies (Anthony and Hawkins 1983; Larsen et al. 2017; Grimaldo et al. 2018), including the present one, found that cod was not affected by artificial light. We also discovered that haddock, saithe, and redfish were each affected by both white and red light. For saithe and redfish, significant differences between the hauls conducted with white or red light were detected, with hauls with red light retaining more fish. Cod, contrary to the other species, seem less reactive to the light treatments, which fits with well with the general knowledge regarding differences in fish behaviour between these species (Larsen et al. 2017; Grimaldo et al. 2018).
In some studies, hauls for which artificial light was applied for underwater observations have been omitted from the selectivity analysis as a precautionary approach (Brinkhof et al. 2017; Grimaldo et al. 2018). Other studies included hauls that used artificial light for underwater observations (Sistiaga et al. 2017; Underwood et al. 2021) based on the assumption that red light is outside the visible wavelengths of the species of interest and therefore would not affect fish behavior and potentially the selectivity results (Goldsmith and Fernandez 1968; Meyer-Rochow and Tiang 1984; Douglas et al. 1995). However, our results demonstrated that even the use of red light for underwater observations affected fish behavior and therefore support omitting such hauls from size selectivity analysis.
On the other hand, the number of selectivity studies that used artificial light to deliberately affect the behavior of fish and crustaceans, and subsequently the selectivity results, has increased substantially (Hannah et al. 2015; Grimaldo et al. 2018; Cuende et al. 2020; Ingólfsson et al. 2021). In some of these studies, artificial light was used to reduce the retention of unwanted by-catch species (Hannah et al., 2015; Lomeli et al. 2018a,b) or sizes (Grimaldo et al. 2018; Ingólfsson et al. 2021). Other studies have documented increased retention of target species (Hannah et al. 2015) or unwanted by-catch species (Larsen et al. 2017; 2018; Lomeli et al. 2018a; Cuende et al. 2020). The different results obtained in studies looking at catch separation (Melli et al. 2018; Karlsen et al. 2021; O’Neill et al. 2022) and effects of light characteristics in the laboratory (Yochum et al. 2022) demonstrate that artificial light impacts behavior and therefore can influence selectivity. The studies also demonstrate that the impact is both species and size dependent, and also that the type of artificial light (wavelength and intensity) and the location of the light in the gear is important. Our study confirmed that even the use of light for underwater observations affected fish behavior and therefore could be used to manipulate size selectivity in fishing gears.
An alternate method that can be used to observe fish behavior is high resolution hydroacoustic sonar, such as the DIDSON (Belcher et al. 2002). The drawback of hydroacoustic devices for behavioral observations is that it can be difficult to interpret the picture and differentiate between similar species. Another possibility is to use indirect methods in which the researcher conducts the evaluation by linking the outcome of a process in order to say something about the internal processes using so-called structural models (Santos et al. 2016) and therefore do not depend on direct observing the internal processes. However, the challenge of how to study fish behavior in fishing gear under low light level conditions remains open, as we demonstrated the risk of biased fish behavior data caused by the light sources required to make observations.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article, and can also be shared upon request.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723. https://doi.org/10.1109/TAC.1974.1100705
Anthony PD, Hawkins AD (1983) Spectral sensitivity of the cod, Gadus morhua L. Mar Behav Phys 10:145–166
Belcher E, Hanot W, Burch J (2002) Dual-frequency identification sonar (DIDSON). In Proceedings of the 2002 International Symposium on Underwater Technology (Cat. No. 02EX556):187–192, IEEE
Brinkhof J, Larsen RB, Herrmann B, Grimaldo E (2017) Improving catch efficiency by changing ground gear design: case study of Northeast Atlantic cod (Gadus morhua) in the Barents Sea bottom trawl fishery. Fish Res 186:269–282
Brinkhof J, Larsen RB, Herrmann B, Sistiaga M (2020) Size selectivity and catch efficiency of bottom trawl with a double sorting grid and diamond mesh codend in the North-east Atlantic gadoid fishery. Fish Res 231:105647
Brinkhof J, Sistiaga M, Herrmann B, Grimaldo E, Larsen RB (2022) Managing size selectivity: the relevance of compulsory and alternative selection devices in the Northeast Atlantic bottom trawl fishery. ICES J Mar Sci 79(9):2399–2412
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. ISBN:978-0-387-22456-5
Cuende E, Arregi L, Herrmann B, Sistiaga M, Onandia I (2020) Stimulating release of undersized fish through a square mesh panel in the Basque otter trawl fishery. Fish Res 224:105431
Douglas RH, Partridge JC, Hope AJ (1995) Visual and lenticular pigments in the eyes of demersal deep-sea fishes. J Comp Physiol A 177:111–122
Efron B (2014) Estimation and accuracy after model selection. J Am Stat Assoc 109(507):991–1007. https://doi.org/10.1080/01621459.2013.823775
Fryer RJ (1991) A model of between-haul variation in selectivity. ICES J Mar Sci 48:281–290. https://doi.org/10.1093/icesjms/48.3.281
Geraci ML, Colloca F, Di Maio F, Falsone F, Fiorentino F, Sardo G, Scannela D, Gancitano V, Vitale S (2022) How is artificial lighting affecting the catches in deep water rose shrimp trawl fishery of the central mediterranean sea? Ocean Coast Manag 217:106005
Goldsmith TH, Fernandez HR (1968) Comparative studies of crustacean spectral sensitivity. Z Vgl Physiol 60(2):156–175
Gordon JD, Bergstad OA, Pascoe PL (2002) The influence of artificial light on the capture of deep-water demersal fish by bottom trawling. J Mar Biol Assoc UK 82(2):339–344
Graham N, Jones EG, Reid DG (2004) Review of technological advances for the study of fish behaviour in relation to demersal fishing trawls. ICES J Mar Sci 61(7):1036–1043. https://doi.org/10.1016/j.icesjms.2004.06.006
Grimaldo E, Sistiaga M, Herrmann B, Larsen RB, Brinkhof J, Tatone I (2018) Improving release efficiency of cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) in the Barents Sea demersal trawl fishery by stimulating escape behaviour. Can J Fish Aquat Sci 75(3):402–416
Hannah RW, LomeliMJM JSA (2015) Tests of artificial light for bycatch reduction in an ocean shrimp (Pandalus jordani) trawl: strong but opposite effects at the footrope and near the bycatch reduction device. Fish Res 170:60–67
Hemmings CC (1973) Direct observation of the behaviour of fish in relation to fishing gear. Helgolander Wiss Meeresunters 24:348–360. https://doi.org/10.1007/BF01609525
Herrmann B, Sistiaga M, Nielsen KN, Larsen RB (2012) Understanding the size selectivity of redfish (Sebastes spp.) in North Atlantic trawl codends. J Northwest Atl Fish Sci 44:1–13
Herrmann B, Sistiaga M, Larsen RB, Nielsen KN (2013) Size selectivity of redfish (Sebastes spp.) in the Northeast Atlantic using grid-based selection systems for trawls. Aquat Living Resour 26:109–120
Herrmann B, Sistiaga M, Grimaldo E, Larsen RB, Olsen L, Brinkhof J, Tatone I (2019) Size selectivity and length-dependent escape behaviour of haddock in a sorting device combining a grid and a square mesh panel. Can J Fish Aquat Sci 76(8):1350–1361
Ingólfsson ÓA, Jørgensen T, Sistiaga M, Kvalvik L (2021) Artificial light improves size selection for northern shrimp (Pandalus borealis) in trawls. Can J Fish Aquat Sci 78(12):1910–1917
Isaksen B, Valdemarsen JW, Larsen RB, Karlsen L (1992) Reduction of fish bycatch in shrimp trawl using a rigid separator grid in the aft belly. Fish Res 13:335–352
Jacques N, Herrmann B, Brinkhof J, Sistiaga M (2024) Understanding size selectivity of trawls using structural-based models: methodology and a case study on fish sorting grids. Aqua Fish. https://doi.org/10.1016/j.aaf.2024.03.003
Jamieson AJ, Godø OR, Bagley PM, Partridge JC, Priede IG (2006) Illumination of trawl gear by mechanically stimulated bioluminescence. Fish Res 81(2–3):276–282
Karlsen JD, Melli V, Krag LA (2021) Exploring new netting material for fishing: the low light level of a luminous netting negatively influences species separation in trawls. ICES J Mar Sci 78(8):2818–2829
Kennelly SJ, Broadhurst MK (2021) A review of bycatch reduction in demersal fish trawls. Rev Fish Biol Fisheries 31(2):289–318
Larsen RB, Isaksen B (1993) Size selectivity of rigid sorting grids in bottom trawls for Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus). ICES Mar Sci Symp 196:178–182
Larsen RB, Herrmann B, Sistiaga M, Brinkhof J, Tatone I, Langård L (2017) Performance of the Nordmøre grid in shrimp trawling and potential effects of guiding funnel length and light stimulation. Mar Coast Fish 9(1):479–492
Larsen RB, Herrmann B, Sistiaga M, Brčić J, Brinkhof J, Tatone I (2018) Could green artificial light reduce bycatch during Barents Sea Deep-water shrimp trawling? Fish Res 204:441–447
Lomeli MJM, Wakefield WW (2019) The effect of artificial illumination on Chinook salmon behavior and their escapement out of a midwater trawl bycatch reduction device. Fish Res 218:112–119
Lomeli MJ, Wakefield WW, Herrmann B (2018a) Illuminating the headrope of a selective flatfish trawl: effect on catches of groundfishes, including Pacific halibut. Mar Coast Fish 10(2):118–131
Lomeli MJM, Groth SD, Blume MTO, Herrmann B, Wakefield WW (2018b) Effects on the bycatch of eulachon and juvenile groundfish by altering the level of artificial illumination along an ocean shrimp trawl fishing line. ICES J Mar Sci. https://doi.org/10.1093/icesjms/fsy105
Marchesan M, Spoto M, Verginella L, Ferrero EA (2005) Behavioural effects of artificial light on fish species of commercial interest. Fish Res 73(1–2):171–185. https://doi.org/10.1016/j.fishres.2004.12.009
Melli V, Krag LA, Herrmann B, Karlsen JD (2018) Investigating fish behavioural responses to LED lights in trawls and potential applications for bycatch reduction in the Nephrops-directed fishery. ICES J Mar Sci 75(5):1682–1692
Meyer-Rochow VB, Tiang KM (1984) The eye of Jasus edwardsii (Crustacea, Decapoda, Palinuridae): Electrophysiology, histology, and behaviour. Zoologica 45:1–685
Millar RB (1993) Incorporation of between-haul variation using bootstrapping and nonparametric estimation of selection curves. Fish Bull 91:564–572
Mueller RP, Brown RS, Hop H, Moulton L (2006) Video and acoustic camera techniques for studying fish under ice: a review and comparison. Rev Fish Biol Fish 16(2):213–226
O’Neill FG, Summerbell K, Edridge A, Fryer RJ (2022) Illumination and diel variation modify fish passage through an inclined grid. Fish Res 250:106297
Preacher K, Merkle E (2012) The problem of model selection uncertainty in structural equation modeling. Psychol Methods 17:1–14. https://doi.org/10.1037/a0026804
Santos J, Herrmann B, Mieske B, Stepputtis D, Krumme U, Nilsson H (2016) Reducing flatfish bycatch in roundfish fisheries. Fish Res 184:64–73
Santos J, Herrmann B, Stepputtis D, Kraak SB, Gökçe G, Mieske B (2020) Quantifying the performance of selective devices by combining analysis of catch data and fish behaviour observations: Methodology and case study on a flatfish excluder. ICES J Mar Sci 77(7–8):2840–2856
Sistiaga M, Herrmann B, Grimaldo E, Larsen RB (2010) Assessment of dual selection in grid based selectivity systems. Fish Res 105:187–199
Sistiaga M, Brinkhof J, Herrmann B, Grimaldo E, Langård L, Lilleng D (2016) Size selective performance of two flexible sorting grid designs in the Northeast Arctic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) fishery. Fish Res 183:340–351
Sistiaga M, Brinkhof J, Herrmann B, Larsen RB, Grimaldo E, Cerbule K, Brinkhof I, Jørgensen T (2022) Potential for codends with shortened lastridge ropes to replace mandated selection devices in demersal trawl fisheries. Can J Fish Aquat Sci 79(5):834–849
Sistiaga M, Herrmann B, Grimaldo E, Larsen RB, Olsen L, Brinkhof J, Tatone I (2017) Combination of a sorting grid and a square mesh panel to optimize size selection in the North-East Arctic cod (Gadus morhua) and redfish (Sebastes spp.) trawl fisheries. ICES J Mar Sci 75:1105–1116
Underwood MJ, Palm ACU, Øvredal JT, Bjordal Å (2021) The response of mesopelagic organisms to artificial lights. Aqua Fish 6(5):519–529
Urquhart GG, Stewart PAM (1993) A review of techniques for the observation of fish behaviour in the sea. ICES Mar Sei Symp 196:135–139
Weinberg KL, Munro PT (1999) The effect of artificial light on escapement beneath a survey trawl. ICES J Mar Sci 56(3):266–274
Wileman DA, Ferro RST, Fonteyne, Millar RB (1996) Manual of methods of measuring the selectivity of towed fishing gears. ICES cooperative research report no. 215: 126
Yochum N, Bryan DR, Britt LL, Berejikian BA, Haehn R, McEntire S, Towler R, Atkins J, Gadberry B, Irvin P (2022) Evaluating Chinook salmon (Oncorhynchus tshawytscha) response to artificial light in support of bycatch mitigation. Can J Fish Aquat Sci 79(6):912–924
Acknowledgements
We thank the Norwegian Fisheries and Aquaculture Research Fund (project number 901633) for funding the project, the Arctic University of Norway for logistical support for our sea trials, and the Norwegian Directorate of Fisheries for the necessary permits. We also thank the crew of R/V “Helmer Hanssen”, Ilmar Brinkhof, Hanne Hatlebrekke, Kristine Cerbule, and Enis Kostak for their help during the cruise.
Funding
Open access funding provided by UiT The Arctic University of Norway (incl University Hospital of North Norway).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brinkhof, J., Sistiaga, M., Herrmann, B. et al. Observing fish behavior in towed fishing gear—is there an influence of artificial light?. Rev Fish Biol Fisheries (2024). https://doi.org/10.1007/s11160-024-09889-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11160-024-09889-5