
Abstract
Recreational spearfishing is a fishing method that occurs globally, yet receives considerably less attention in the scientific literature relative to other recreational fishing methods, such as angling. Lack of scientific information on spearfishing may negatively affect the development and management of marine recreational fisheries. We conducted a systematic review of 102 peer-reviewed papers published between 1967 and 2022 pertaining to marine recreational spearfishing. Based on this literature review, we provide an overview of key insights across social, economic, and ecological dimensions of marine recreational spearfishing. While spearfishers represent less than 5% of marine recreational fishers, the participants are younger and may differ from recreational anglers in their motivations, with suggestions of increased well-being generated from a close connection with the sea during underwater fishing. Recreational spearfishers mostly target species of moderate to high levels of vulnerability that are mid to high trophic level carnivores. Though spearfishers can deliberately target larger individuals of exploited populations, this is not a generalizable pattern. Despite a growing body of research on the ecological impacts of marine recreational spearfishing, there is limited knowledge of these effects and their mechanisms across biological levels of organization (e.g., individual, population, community and ecosystem) compared with those of other fishing methods. Recreational spearfishers can contribute to advances in marine ecological knowledge, and inclusive participatory management could represent a key step towards transformative sustainable development of marine recreational spearfishing. Throughout the review, we identify gaps in the research and areas where future research is needed to better inform the socio-economic importance, ecosystem impacts and future management of marine recreational spearfishing.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Definition of marine recreational spearfishing
Spearfishing is the use of hand-held underwater gear to capture marine organisms such as fish, cephalopods and crustaceans. Underwater refers exclusively to fishing actions undertaken by means of freediving, SCUBA diving or hookah diving (Fig. 1). The most common underwater harvesting gear are spearguns, pole spears, and Hawaiian slings. However, more rudimentary tools can also be used, such as hand spears or hand hooks (Fig. 1). Recreational spearfishing is defined as “fishing of aquatic organisms that neither constitute the individual’s primary source of nutrition nor are sold or otherwise traded on any market” (FAO 2012). Thus, while a key motivation of recreational spearfishers may relate to catching food, unlike commercial or subsistence fishers, these activities are not integral to the livelihood or nutritional needs of the individual and their family (FAO 2012).

Source of pics: (A, D, E, I: Marco Bardi; B: Sisbiota-Mar; C, F, G, H: from the manufacturers’ websites)
Spearfishing can be practiced by means of three main diving techniques: freediving (A); scuba diving (B); and hookah diving (C). Spearfishers can use a variety of underwater gear such as spearguns with slings (D) or pneumatic (E), pole spears (F), Hawaiian slings (G), hand spears (H), or hand hooks (I).
Introduction
The act of catching aquatic organisms with a spear is an ancient human activity that dates back at least 90,000 years (Yellen et al. 1995), likely preceding the development of more complex gear types, such as hooks, nets, and lines. Presently, spearfishing is practiced for recreational, subsistence, or commercial purposes. While scientific literature pertaining to recreational fishing has primarily focused on rod-and-line angling (e.g., Cooke et al. 2019, 2021; Lewin et al. 2019), there may be social, economic, and ecological differences between recreational angling and spearfishing (Gordoa et al. 2019; Sbragaglia et al. 2020b), such that it is crucial to devise specific management actions for each sector.
Spearfishing may differ from other recreational fishing methods in several ways. First, spearfishers often diverge demographically and attitudinally from anglers. For example, spearfishers have been found to report greater satisfaction related to their catch and associated activity when compared to anglers (e.g., web-surveyed Spanish recreational fishers; Gordoa et al. 2019; see also "Discussion of social and economic aspects" section) and appear to show different engagement dynamics on social media relative to recreational anglers (Sbragaglia et al. 2020b; see also "Discussion of social and economic aspects" section). Second, in contrast to most types of recreational fishing, spearfishing is an active, underwater method of fishing where, almost uniquely, fishers are able to select which individuals they harvest because they can visually identify and selectively target all potential catch (Pavlowich and Kapuscinski 2017). Third, spearfishers use several “hunting” techniques that are more behaviorally comparable to those used by natural predators (e.g., ambush, sit-and-wait, or active hunting strategies) than other fishing methods such as angling or netting. Thus, spearfishing may trigger defensive behaviors in targeted species that are analogous to natural anti-predator responses (Samia et al. 2019). Fourth, the selectivity patterns of spearfishing may differ from other fishing methods because of the depth limitations of spearfishers, especially in freediving (Lindfield et al. 2014; Sbragaglia et al. 2020b). Combined, these attributes of recreational spearfishing could drive differences in socio-economic and ecological outcomes of spearfishing relative to other fishing methods. For example, recreational spearfishing could foster unique psychological and social drivers, such as a stronger connection with the marine environment and different motivation for fishing compared to angling. Recreational spearfishers could also provide a unique form of local ecological knowledge, including a distinct sensitivity in understanding and monitoring changes of marine ecosystems due to their perspective of the underwater environment. Furthermore, the selective and active nature of spearfishing could potentially drive distinct cascading effects on ecosystem processes and functioning different to that expected from other fishing methods (e.g., cascading effects from selective predator removal; Scheffer et al. 2005). However, differences in socio-economic and ecological outcomes of spearfishing relative to other recreational fishing methods are largely hypothetical and, when scientific evidence is available, it is not synthetized and accessible for managers and policy makers. The consequences of this lack of knowledge could result in poor management decisions about spearfishing with respect to other fishing methods. On one hand, excessive limitations of the activity of spearfishers could negatively affect socio-economic outcomes. On the other hand, lack of appropriate and tailored management regulations for spearfishers could have negative ecological consequences. Therefore, it is critical to provide an updated synthesis of socio-economic and ecological aspects of spearfishing to inform managers and policy makers and guide future research activity. This is largely available for recreational fisheries as a whole (e.g., Cooke et al. 2019, 2021; Lewin et al. 2019), but much of this is related to rod-and-line angling, and consequently not useful for addressing specific challenges in management and research of recreational spearfishing.
We first conducted a systematic review of published peer-reviewed research papers including both marine recreational and subsistence/artisanal/commercial spearfishing. This was done to compare between recreational and other forms of spearfishing in terms of geographical and temporal patterns. We then developed an integrative review to characterize and synthesize the main topics of research on social, economic, and ecological dimensions of marine recreational spearfishing. Specifically, we focused on demography and motivation for the social dimension, expenditure for the economic dimension, while for the ecological dimension we synthetize information on catch per unit effort, selectivity, change in behavioral phenotypes, and local ecological knowledge. Though our review focuses on marine recreational spearfishing, many aspects may be applicable to freshwater recreational spearfishing as well as to subsistence, artisanal, or commercial spearfishing. We mainly address spearfishing of finfishes where it is necessary to use spearguns, pole spears, or Hawaiian slings to catch the target species. However, we recognize that cephalopods (e.g., cuttlefish and octopuses), crustaceans (e.g., crabs and lobsters), molluscs (e.g., scallops and bivalves), and echinoderms (e.g., sea urchins and sea cucumbers) may also be harvested by spearfishers with rudimentary gear such as hand spears, hand hooks, or taken by bare hands in distinct regions of the world.
Methodological approach
We used the keywords “spearfishing” or “spear fishing” to search published peer-reviewed papers (written in English) on Web of Science and Scopus databases. We identified and retrieved a total of 462 papers based on queries of the two databases on 21st of April 2022 (see diagram referring to the Preferred Reporting Items for Systematic Reviews and Meta Analyses, PRISMA, in Figure S1). After a first screening of titles and abstracts, we excluded 268 papers mainly because they were not related to spearfishing. The large proportion (49%) of these papers referred to the genus Tetrapturus, which comprises several fish species commonly known as “spearfish” (e.g., Mediterranean spearfish, Tetrapturus belone). Next, we accessed the full-text version of the remaining papers and excluded 47 of them because they were related to freshwater spearfishing, to fishing with a spear from outside of the water or to papers where spearfishing was not empirically evaluated, but only referred to anecdotally as a possible cause of the observed ecological patterns. After these two screenings, we retained 147 papers related to marine spearfishing for further analyses (Fig. S1; Table S1).
With the 147 published papers, we conducted a systematic review of the full-text documents to determine their spatial, temporal, and thematic scope. We extracted the following information ("Results of the systematic review" section): (1) geographic location where the study was conducted; (2) year of publication; (3) whether the study was related to recreational or other forms of spearfishing (i.e., subsistence, artisanal, or commercial); (4) whether the study evaluated spearfishing as a response or predictor (i.e., research on spearfishing), versus whether spearfishing was used as a sampling method or metric for other purposes or inquiry (e.g., for tracking changes in marine ecosystems); and (5) the main research dimension of the study, which included the following: ecological (e.g., catch composition or fish behavioral response to spearfishing), social (e.g., human motivations and fishing activity satisfaction), or economic (e.g., expenditures).
Focusing on the subset of published papers exclusively related to marine recreational spearfishing, we built a co-occurrence network with the keywords that most often appear in the abstract of the studies to have a comprehensive view of the topics of interest of the studies and how such topics were connected among them. We cleaned the text of the abstracts of the papers by removing numbers, stopwords and punctuations. Subsequently, we tokenized the text by isolating all the possible two-words combinations and quantified their frequency of occurrence. Then, we created a co-occurrence matrix to visualize the connections between the different words that occurred at least five times. This allows one to identify the pattern of connections between co-occurring words suggesting their importance in the entire body of literature and also provide insight about the strength and centrality of specific words (e.g., Lozano et al. 2019). The visual display of the network was organized according to the Fruchterman-Reingold layout, which is a force-directed layout algorithm which treats edges (i.e. words) like springs that move vertexes closer or further from each other in an attempt to find an equilibrium that minimizes the energy of the network (Csardi and Nepusz 2006). Furthermore, we analyzed the structure of communities in the network using a fast greedy modularity optimization algorithm (Clauset et al. 2004), which highlights how co-occurring networks of words may cluster around specific topics indicating main research interest. We ran quantitative analysis of comments in R version 3.5.0 (R, Core Team 2018) with the additional packages “quanteda” (Benoit et al. 2018), and “igraph” (Csardi 2013).
Results of the systematic review
The papers analyzed during the systematic review were published from 1967 to 2022. Of the 147 selected papers, 102 (69%) focused on recreational spearfishing and 45 (31%) on subsistence, artisanal, or commercial spearfishing. The latter 45 were grouped as “others” because the main focus of this review was marine recreational spearfishing. Of the 102 marine recreational spearfishing papers (Table S1), 65 (64%) directly investigated spearfishing and 37 (39%) used spearfishing for other purposes. Of the remaining 45 papers focusing on other forms of marine spearfishing (i.e., subsistence, artisanal, or commercial), 34 (76%) directly investigated spearfishing and 11 (24%) used spearfishing for other purposes. In the following sections we describe the geographical distribution, the temporal pattern, and the research dimensions of the 147 papers included in the systematic review. However, considering that the focus of the review is recreational spearfishing, we only synthesized the social, economic, and ecological dimensions of the 102 marine recreational spearfishing papers ("Discussion of social and economic aspects" and "Discussion of ecological aspects" sections; Table S1).
Geographical distribution
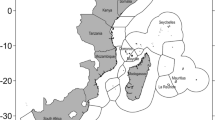
Published papers related to marine recreational spearfishing activities were distributed across the globe, and were particularly concentrated in the northeastern Atlantic Ocean and the Mediterranean Sea (74% in developed countries; Fig. 2A). By contrast, scientific papers related to other forms of spearfishing were also distributed across the globe, but were more heavily concentrated in the western and central Indo-Pacific and the Caribbean Sea (11% in developed countries; Fig. 2A). This could highlight the dominant type of spearfishing that occurs in those areas or may also represent a publishing or research bias. For example, we did not identify any paper dealing with commercial spearfishing from Europe, which is a minor activity compared to the magnitude of recreational spearfishing, but countries such as Italy allow a limited number of commercial spearfishing licenses (V.S. personal observation). Studies related to research on spearfishing were distributed widely around the world (56% in developed countries), while papers using spearfishing for other scientific purposes (e.g., tracking distributional range shifts using local ecological knowledge) were mostly confined to the Mediterranean and the Caribbean seas (51% in developed countries; Fig. 2B).

Geographical distribution of the published papers identified during the systematic review according to; A the typology of spearfishing (i.e. recreational or others, i.e., subsistence, artisanal or commercial) and B whether the papers were related to research on spearfishing or using spearfishing for other scientific purposes
Temporal pattern
In the last decade, there has been an increase in the number of published papers on marine recreational spearfishing relative to those on other forms of spearfishing (Fig. 3A). For example, in 2006–2010 only four studies were published characterizing marine recreational spearfishing, while in 2018–2022, 26 papers were published on this topic (Fig. 3A). In 2006–2014, there was also an increase in the number of studies published on other forms (i.e., subsistence, artisanal, or commercial) of spearfishing, with 10 papers published on this topic in 2010–2014, though the number of new papers published on this topic has declined slightly since 2014 (Fig. 3B), suggesting an increasing interest in empirical study of marine recreational spearfishing relative to other forms of spearfishing. In 2010–2018, we also observed an increase in the number of studies using recreational spearfishing for other research purposes, but this has declined slightly since 2018–2022 (Fig. 3B). By contrast, the number of studies investigating other forms of spearfishing for other purposes has remained consistently low over time (Fig. 3A, B). As an important caveat, results for 2022 represent only 4 months (i.e., papers we accessed in April 2022), which could affect our observed temporal trends.

The total number of new studies published per 4-year time period related to spearfishing, as identified during the systematic review. The papers were grouped according to the spearfishing typology (recreational or others, i.e. subsistence, artisanal or commercial) and whether the papers were related to A research on spearfishing or B using spearfishing for other purposes
Research topics and dimensions
Of the 102 published studies on marine recreational spearfishing, the ecological dimension was most commonly studied (90 studies) relative to social (20) and economic (6) dimensions (Table S1). The majority of studies investigated topics related to only one of the three research dimensions, i.e. ecological, economic, or social (93 out of 102 studies; Table 1). By contrast, only a few papers investigated all the three dimensions simultaneously (4 out of 102; Table 1).
The co-occurrence analysis showed two main word networks composed by three different communities (Fig. 4; Table S2). The first network was composed by one main community centered around the words “recreational” and “fishing”, which was linked to two other communities: one centered around the word “management”, and the other centered around the words “marine” and “reserve” (Fig. 4; Table S2). The second network was composed by one main community centered around the word “fish”, which was linked to two other communities: one centered around the words “total” and “catch”, and the other centered around the word “reef” (Fig. 4; Table S2). Other small communities with terms such as “goliath grouper”, “body size”, “visual census”, “trophic level”, and “social media” were also identified (Fig. 4; Table S2).

Co-occurrence network analysis of the most common keywords used in the abstract of the 102 published studies analyzed here. The width of the edges (i.e., grey lines) between nodes (i.e., words) represents the strength (i.e., the sum of weights attached to ties belonging to a node; see also Table S2). The network is organized according to the Fruchterman-Reingold layout, while colored areas represent community structure resulted by a fast greedy modularity optimization algorithm (see methods for more details)
Discussion of social and economic aspects
Social aspects
We identified 19 published papers that directly investigated the social aspects of marine recreational spearfishing. According to the reviewed studies, spearfishers make up less than 5% of marine recreational fishers in countries such as Spain (Morales-Nin et al. 2005; Pita et al. 2018; Gordoa et al. 2019), Denmark (Skov et al. 2020), and South Africa (Mann et al. 1997). The average age of recreational spearfishers varied among countries, and was 30 years (South Africa; Mann et al. 1997), 33 years (Portugal; Assis et al. 2018; Martínez-Escauriaza et al. 2020), and 36–37 years (Spain and Jamaica; Ennis and Aiken 2014; Pita et al. 2018; Gordoa et al. 2019). Web-surveyed Spanish recreational spearfishers were younger (36 years) than recreational shore (41) and boat (45) anglers (Gordoa et al. 2019). This result is also supported by a recent on-site survey about recreational fishing, conducted in Catalonia (Spain), which reinforced the finding that recreational spearfishers are younger (37) than recreational shore (48) and boat (53) anglers (Vitale et al. 2021). Given the physical nature of spearfishing (especially in freediving), the pattern is not surprising and may extend to other countries and regions. We presume that the physical fitness required to practice recreational spearfishing, especially freediving, is more demanding than that required by other forms such as recreational angling.
Outdoor activities, such as recreational fishing in general, provide a range of social, psychological, and physiological benefits (Manfredo et al. 1996; Parkkila et al. 2010; Fig. 5). Though there is less published information on the motivations and received benefits of recreational spearfishers compared to recreational anglers, spearfishing has both catch and non-catch motivations, similar to the case of angling (Fedler and Ditton 1994). The acquisition of high quality food can also be an important motivation for many recreational fishers, particularly those that use lethal methods, such as spearfishers (Assis et al. 2018; Terlizzi et al. 2022), and where the decision to target and harvest the fish is a deliberate choice (Cooke et al. 2018). For example, among the many identified non-catch related motivations for recreational spearfishers in Portugal, one of the main motivations was “to be in touch with the sea” (Assis et al. 2018). Moreover, recreational spearfishing provides different social benefits relative to angling, as shown by web-surveyed Spanish recreational spearfishers that report higher levels of catch and activity satisfaction than recreational anglers (Gordoa et al. 2019).
The additional social benefits related to spearfishing may relate to the underwater experience (Young et al. 2016), but robust scientific evidence is still lacking (Januchowski-Hartley et al. 2020). In Italy, recreational spearfishing videos triggered strong appreciation for the freediving and fishing actions displayed on social media, something not observed for anglers (Sbragaglia et al. 2020b). Indeed, spearfishers use the term “aquaticity” to refer to their capacity to relax and move in a fluid and smooth way underwater. A compelling and systematic review on the term “aquaticity” highlighted how human contact with the water could promote physical as well as psychological and emotional well-being (Varveri et al. 2016). Images and sounds of marine ecosystems have also demonstrated beneficial psychological and physiological effects that include reducing stress, and helping reduce sleep disorders and depression (Bratman et al. 2019). Moreover, the human body undergoes physiological changes when diving, especially freediving (i.e., the human diving response; Foster and Sheel 2005), some of which could result in evolutionary adaptations. For example, the indigenous Bajau people (in southeast Asia; Abrahamsson and Schagatay 2014) evolved an increased spleen size (an adaptation to providing them with a larger reservoir of oxygenated red blood cells during spearfishing diving) and other adaptations related to genes controlling the diving response (Ilardo et al. 2018). Although recreational spearfishing and angling provide health benefits associated with experiencing blue spaces (Pretty et al. 2006), it is plausible that social, psychological, and physiological benefits associated to the underwater aspects of recreational spearfishing differ from those of recreational angling, but no rigorous comparisons exist.
In summary, spearfishers represent a younger numerical minority within the population of marine recreational fishers, but other demographic information (e.g., gender, race, socioeconomic status) is currently limited. They have a strong motivation to be underwater, may generally be in good physical condition, and may experience higher well-being rewards from spearfishing activity relative to anglers. Future research is especially needed on the demographics of recreational spearfishers, to better inform the social implications of this activity and its cultural importance relative to recreational angling. Additional research should also evaluate whether the athletic performance required for practicing recreational spearfishing could promote a healthier lifestyle and state of well-being—as well as exposing spearfishers to risks such those related to hypoxia and blackout (e.g., Lindholm and Lundgren 2009)—relative to other recreational consumptive (e.g., recreational angling) and non-consumptive (e.g., scuba diving) recreational activities.
Economic impact
We identified six published papers that investigated certain economic aspects of recreational spearfishing. Several of these studies contained expenditures related to spearfishing, which often differ by region within and across countries. For example, in Spain, recreational spearfishers in Catalonia spent roughly €800 per person per year in acquiring goods and services directly related to spearfishing activities (Lloret et al. 2008), versus €1700 per year in Galicia (Pita et al. 2018). For comparison, in the Canary Islands, Jiménez-Alvarado et al. (2020) estimated that recreational spearfishers spent on average €484 as an initial investment in equipment, and then an average annual expenditure of €245 (including travel, maintenance, equipment replacement, insurance and a fishing license). The estimates for Catalonia and Canary Islands are similar to the estimates for Spanish marine recreational fishers (€672 per year; Hyder et al. 2018), while estimates for Galicia are twice as much. This may be related to the fact that GDP is higher in the Atlantic than in the Mediterranean areas with obvious repercussion on recreational fishing expenditure (Hyder et al. 2018). Economic estimates for mainland Portugal also report that 65% of recreational spearfishers spent less than €500 per year in direct expenditures related to the activity (namely on equipment and trips), while the remaining 35% spent over €500 per year for the same expenditures (Assis et al. 2018). This estimate is lower than that obtained by Hyder et al. (2018) for the average annual spending by Portuguese marine recreational fishers considering all fishing methods (€796; Hyder et al. 2018). This may be expected since the great majority of marine recreational fishers in Portugal are shore anglers with lower income and with less expenditures than spearfishers and boat anglers (Diogo et al. 2020). In Cyprus, recreational spearfishers spent on average €748, which is less than what was estimated for marine recreational anglers in the same study (Michailidis et al. 2020), but it is more than double that of previous estimates of expenditure for marine recreational fishers in Cyprus (€300; Hyder et al. 2018).
Most recently, two studies characterized the economic impact of spearfishing tournaments in Australia and Italy respectively, both of which demonstrate the local economic value of spearfishing tournaments and suggest trading-off the economic dimension with social benefits and potential ecological impacts (Schilling et al. 2022; Terlizzi et al. 2022). Although two recent papers have estimated the economic importance of recreational fishing at global (Cisneros-Montemayor and Sumaila 2010) and regional levels (Hyder et al. 2018), specific estimates for spearfishing are missing. The comparison with economic expenditure of European marine recreational fishers (Hyder et al. 2018), suggests that spearfishers may spend more or less than recreational anglers with contrasting patterns across countries and regions. To understand the full spectrum of the economic contribution of recreational spearfishing, future research should move beyond the current characterization of expenditure and investigate the knock-on effects of the activity on other economic sectors (Dyck and Sumaila 2010), as well as the economic and socio-cultural values of ecosystem services provided by spearfishing activities relative to other activities (De Groot et al. 2012). This is especially important because many countries separately manage recreational spearfishing and recreational angling, such that economic evaluations specific to recreational spearfishing could be important to ensure balanced and fair management decisions.
Discussion of ecological aspects
Catch per unit effort and catch characteristics
Among the 53 reviewed papers that investigated ecological aspects of recreational spearfishing, 23 estimated catch per unit of effort (CPUE; see also Table 1). “Fishing effort” also appeared in the central network of the co-occurrence analysis (Fig. 4). All except 2 studies out of the 23 (which reported CPUE as number of fish per day, Table 1) reported biomass of fish extracted by spearfishers per year, day, or hour. Therefore, we were able to standardize harvest rate estimates using a general assumption based on the literature review (i.e., 30 fishing days per year; 4 h per day; considering only most recent values where trends in CPUE were reported) or information presented in the paper (Table 1). Values of CPUE indicated that estimated fishing efficiency varied from 0.08 to 2.6 kg/spearfisher/hour (Table 1). These values are similar to those found for artisanal spearfishing in the Indo-Pacific, which average between 0.5 and 2 kg/spearfisher/hour (Hamilton et al. 2012; Cohen and Alexander 2013; Januchowski-Hartley et al. 2014; Humphries et al. 2019) during normal fishing activities. However, these values fall significantly below the 3.6–9 kg/spearfisher/hour that has been estimated for night spearfishing, or when targeting spawning aggregations (Hamilton et al. 2012; Rhodes et al. 2018).
Comparison of CPUE estimates may not always be appropriate due to the different spatial and temporal scales and methods used. CPUE was estimated during different types of activities, including both recreational activity (15 out of 23) and tournaments (8 out of 23), different time periods (spanning 1996–2022), across several regions of the world, sites with different histories of exploitation, and with considerable differences in catch composition both relative to species targeted and number of species harvested. Tournament data are particularly useful for CPUE comparisons as they allow for longitudinal data analysis and comparisons across controlled conditions in space and time (e.g., sites with similar histories of exploitation, similar catches with regards to species targeted and harvested, under the same environmental conditions, and with the same gear requirements and fishing restrictions), while also representing individual heterogeneity among participants. For example, large differences in CPUE have been documented among participants within the same tournament (Lincoln Smith et al. 1989; Coll et al. 2004), as well as among different types of tournaments (Cinelli and Fresi 1980; Pita and Freire 2014). The drivers governing such differences are still unknown, but Terlizzi et al. (2022) showed that CPUE of spearfishers swimming out from the shore is half that of spearfishers using a boat during tournaments, potentially due to access to fish or to physical effort, which may be of relevance to assessment and management.
The average intrinsic vulnerability index of fishes (i.e., an index ranging from 0 to 100, which integrates life history and ecological characteristics of marine fishes to estimate their intrinsic vulnerability to fishing; Cheung et al. 2005) harvested within recreational spearfishing catches was reported in eight papers from the Mediterranean Sea, North-East Atlantic Ocean, and Australia (Fig. 4; see also Table 1). These values spanned from 36.7 (moderate) to 55.5 (high) intrinsic vulnerability. The trophic levels of species targeted by recreational spearfishers were reported in six papers and ranged between 3.24 and 3.89 (Table 2). This suggests that recreational spearfishers mostly target species of moderate to high levels of vulnerability that are mid- to high-trophic level carnivores.
Selectivity of recreational spearfishing and potential effects on marine ecosystems
Among the 53 reviewed papers that directly investigated ecological aspects of recreational spearfishing, 19 were directly or indirectly related to the size of the fish targeted by recreational spearfishers, and “body size” and “highly selective” appeared as relevant nodes in the co-occurrence analysis (Fig. 4). Of these 19 papers, only three provided evidence that recreational spearfishing targets larger individuals compared with other fishing methods (e.g., recreational angling) that target the same species (Dalzell and Smith 1998; Harper et al. 2000; Frisch et al. 2008). Recent evidence suggests that in some instances recreational angling may select for larger common dentex, Dentex dentex than recreational spearfishers (Marengo et al. 2015; Sbragaglia et al. 2020b), which could be an emerging pattern linked to technological innovations, such as underwater cameras, boat-based electronics and use of live prey as bait (Cooke et al. 2021). Therefore, management actions aiming to mitigate the impact of selective harvesting of larger individuals should carefully consider updated information for the target species in light of recent technological innovations.
We did not find papers that directly addressed the unique effects of recreational spearfishing (relative to other fishing methods) on biomass and food-web dynamics. Spearfishing has been shown to reduce the abundance of exploited predatory species (Godoy et al. 2010; Giglio et al. 2017; Hall et al. 2022). This in turn can reduce density-mediated and risk-mediated effects of these species on prey and competitor populations, with top-down cascading effects on prey and competitive release on community structure and potential changes in ecosystem functions (Baum and Worm 2009; Estes et al. 2011). A classic example—not related to spearfishing—is that overexploitation of otters (i.e., top predators) may lead to explosion of their preys (i.e., mesopredators or grazers) and consequently kelp forest overgrazing (Estes et al. 1978). However, whether similar cascading effects can or has occurred for marine recreational spearfishing in a mixed coastal fisheries context is unknown, and represents an important aspect to address in future research. Furthermore, an additional aspect to consider in this context is that both top predators (i.e., sharks) and humans could present similar lethal threat to mesopredatory fishes in coastal marine systems (Asunsolo-Rivera et al. 2023), with potentially similar implications in controlling top down cascading effects.
One study identified additional effects of recreational spearfishing on marine ecosystems related to inadvertently damaging important habitats such as corals in areas that experience high levels of fishing effort (Giglio et al. 2018). However, it is possible that such damage is less than that associated with other fishing methods, because there may be fewer anchor deployments per trip compared to angling (Frisch et al. 2008). Therefore, recreational spearfishing may exert lower levels of habitat and environmental damage compared with other fishing methods because there is essentially no discarded gear, little if any bycatch, no requirement for bait, and less littering (Frisch et al. 2008; Diogo et al. 2020).
Changes in behavioral phenotypes of fishes
We identified six papers that estimated the effects of spearfishing on fish behavior (Table 1). The most common behavioral traits measured in these studies reportedly influenced by spearfishing were related to anti-predator behavior (a focus on such behavior is supported by “antipredator behavior” appearing as relevant nodes in the co-occurrence analysis; Fig. 4). Antipredator behavior was quantified using a number of metrics, the most common of which was flight-initiation distance, which is defined as the distance at which a prey flees from an approaching predator (Ydenberg and Dill 1986; Stankowich and Blumstein 2005). There is strong agreement among studies that recreational spearfishing pressure increases flight-initiation distance of targeted species, which aligns with studies showing that passive fishing gear (e.g., hook-and-line, traps and stationary nets) can trigger a “timidity syndrome” in exploited fish populations (i.e., the emergence of fish populations that are more timid when exploited compared to unexploited populations of the same species; Arlinghaus et al. 2017b). For example, Sbragaglia et al. (2018) showed that Mediterranean fishes can differentiate spearfishers from snorkelers and adjust their flight initiation distances to be greater than the range of spearguns. Spearfishing may also influence other aspects of defensive behavior, such as pre- and post-flight behavior (Januchowski-Hartley et al. 2011; Nunes et al. 2015; Bergseth et al. 2016), as well as diurnal hiding behavior (Côté et al. 2014). Moreover, flight initiation distance of fishes can decrease with depth (Stamoulis et al. 2019; Pereira et al. 2020), which suggests that spearfishing impact may decrease with increasing depth due to physical limitation of divers. Furthermore, a potential depth refuge could exist in deeper water, likened to a marine protected area with vertical boundaries, where larger fish persist in deeper waters out of the reach of spearfishing can help repopulate the shallower habitats over time (Lindfield et al. 2014).
These various changes in behavioral phenotypes of fishes may result in time and energy costs for individuals (Dill et al. 2003), and may also affect group behavior, with possible repercussions for fisheries and conservation strategies (Sbragaglia et al. 2021b). For example, there can be strong links between anti-predator behavior and other functionally important behaviors (e.g., movement, feeding, mating) and the potential for community-wide dynamics and ecosystem-level consequences of recreational spearfishing. Changes in behavior due to protection may be transmitted outside protected areas, and although likely limited in both spatial and temporal reach, may increase susceptibility to recreational spearfishers (Januchowski-Hartley et al. 2013, 2014). The reduced boldness of fishes may be linked to reduced feeding rates on standard prey in fished areas (with spearfishing) relative to protected areas (Rhoades et al. 2019; Skinner et al. 2019). In particular, spatio-temporal activity patterns can be modified as expected from the “landscape of fear” hypothesis (Brown et al. 1999; Gaynor et al. 2019), but such effects specific to spearfishing are empirically difficult to test especially in mixed coastal fisheries. Ascertaining effects on other functionally important behaviors of targeted species will be an essential area of research to develop effective ecosystem-based management of both commercial and recreational fisheries. Behavioral changes may be used as indicators of spearfishing disturbance of fish population (Bergseth et al. 2016; Tran et al. 2016; Sbragaglia et al. 2018) and to inform associated conservation strategies (e.g., Berger-Tal et al. 2011), but the link to population dynamics is not always straightforward (i.e., behavioral changes do not always affect survival or reproduction; Gill et al. 2001). For example, species that show strong avoidance of spearfishers presence may be in less need of protection relative to those that do not (Gill et al. 2001). Additional study of the effects of recreational spearfishing on other functionally important behaviors and the functional role of targeted species will be important for understanding the individual, population, and community-level consequences of marine recreational spearfishing.
The ecological knowledge of recreational spearfishers
We retrieved 37 papers that used spearfishing for other scientific purposes (26 in a systematic way, and 11 in a haphazard way; Table 2). Interests spanned from understanding trends of fish populations (27%), tracking occurrence of non-indigenous species (19%), quantifying distributional range-shifts of species (16%), characterizing fish assemblages (16%), and analyzing general fish biology (11%; see other categories in Table 2). Ecological information has also been extracted from spearfisher-held underwater cameras in papers published after 2011 (30%; Table 2). Underwater videos actively recorded by recreational spearfishers may be useful for monitoring fish assemblages, analyzing fish behavior, and testing hypotheses at large spatial and temporal scales (Bulleri and Benedetti-Cecchi 2014). Additionally, the use of social media as a tool for passive data collection or interaction with spearfishers appeared in papers published after 2016 (13%; Table 2), with “social media” appearing as relevant nodes in the co-occurrence analysis (Fig. 4). Emerging digital approaches represent a promising tool for characterizing spatio-temporal distribution and dynamics of marine life, and the use of digital data can benefit from the high spatial dispersion of marine recreational spearfishers, and by accessing the information directly from digital platforms (Sbragaglia et al. 2021a; Lennox et al. 2022). However, operationalization of data-mining approaches from digital data is still ongoing, and future integration of machine learning and harmonized datasets could boost applications of these methods inside and outside academia (Lennox et al. 2022). Data from spearfishing tournaments have been used for gathering ecological knowledge (9 out of 37; Table 2), such as catch data providing information on marine fish communities over time (Mann et al. 1997; Pita and Freire 2014; Gledhill et al. 2015; Boada et al. 2017).
Spearfishing tournaments may also be used to mitigate the invasion of non-native species such as lionfish, Pterois volitans, in the Atlantic Ocean (Barbour et al. 2011; Harris et al. 2019, 2020). Culling of lionfish by spearfishing appears to be one of the most effective community-led methods that might help mitigate the spread and ecological impacts of invasive lionfish (Malpica-Cruz et al. 2016). Indeed, most of the papers we identified using spearfishing for other scientific purposes were from the Caribbean Sea, where lionfish invasions are causing severe damage (Barbour et al. 2011). The interaction between spearfishing tournaments and biological invasions represents an important example to understand how synergies between spearfishing groups and conservation authorities can react and respond to complex ecological challenges.
In summary, the use of recreational spearfishers in scientific studies, combined with their time underwater, suggests that over time, experienced recreational spearfishers may develop a deep knowledge of habitat preferences, behavior and occurrence of targeted species which leads them to develop stewardship ethics as recently conceptualized for recreational anglers (Shephard et al., in press). Recreational spearfishing can directly (e.g., by spearfisher local ecological knowledge) or indirectly (e.g., by passive mining of digital data or tournaments) contribute to ecological understanding and conservation of marine ecosystems (e.g., Sbragaglia et al. 2020a; Foster et al. 2023). Such use may be encouraged, especially as voluntary bottom-up resource-conserving actions (Sbragaglia and Arlinghaus 2020), because they provide complementary and alternative information to costly institutional actions (Cooke et al. 2013; Fujitani et al. 2017; Arlinghaus et al. 2019). An overarching question is whether the ecological knowledge of recreational spearfishers differs from that of recreational anglers and other types of fishers, and how it can be integrated with other knowledge systems to better inform fisheries management (e.g., Lee et al. 2019).
Concluding remarks and future directions
We provide a multidisciplinary and global characterization of marine recreational spearfishing spanning the range of social, economic, and ecological dimensions as reported in the current literature (Fig. 5). Research interest in marine recreational spearfishing is increasing, but there are several key points to keep in mind for the future study and management of marine recreational spearfishing.

Conceptual representation of the different processes involved in multidisciplinary research applied to marine recreational spearfishing. Socio-economic dynamics are presented at the top: contribution to scientific research ("The ecological knowledge of recreational spearfishers" section); social ("Social aspects" section) and economic ("Economic impact" section) aspects. Ecological processes are presented at the bottom; downsizing of individuals ("Changes in behavioral phenotypes of fishes" section); timidity syndrome ("Selectivity of recreational spearfishing and potential effects on marine ecosystems" and "Changes in behavioral phenotypes of fishes" sections); possible cascading effects at ecosystem level ("Changes in behavioral phenotypes of fishes" section). Symbols are courtesy of the Integration and Application Network (http://ian.umces.edu/symbols/)
Recreational spearfishers may consistently differ from recreational anglers in their motivations and the human well-being generated by the fishing activity, though the demography of spearfishers (gender, race, and socioeconomic status) remains largely unstudied. Considering that recreational fisheries are complex adaptive social-ecological systems (Hunt et al. 2013; Arlinghaus et al. 2017a), differences within recreational fishing should be accounted for specifically in those situations where recreational angling and spearfishing are managed with different regulations and paradigms. We showed an increasing number of papers focused on understanding marine recreational spearfishing in the last decade, which could help to fill such knowledge gaps in the future.
Recreational spearfishers selectively target mid- to high-trophic level and mid- to high-vulnerability species (e.g., slow life history traits), which may exacerbate their ecological impact in comparison to other fishing methods (for example, triggering density-mediated or risk-mediated effects with consequent cascading effects; Baum and Worm 2009; Estes et al. 2011; Boaden and Kingsford 2015). This should be carefully evaluated with respect to other fishing methods considering recent technological development. For example, in terms of size selectivity, there are two aspects to consider when looking at ecological impacts on exploited fish population dynamics. Firstly, depending on the species, other fishing methods could potentially exert the same or even a stronger pressure on larger individuals and vulnerable species (e.g., such as the case of Dentex dentex in the Mediterranean Sea; Marengo et al. 2015; Sbragaglia et al. 2020b). Management in these contexts requires appropriately assessing the relative impacts of fishing methods in light of recent fishing technological innovations (Cooke et al. 2021). Secondly, other fishing methods could potentially exert stronger pressures on immature individuals compared to recreational spearfishing (Frisch et al. 2008). Therefore, predictions of ecological impacts in terms of species-specific population productivity should consider fishing mortality of larger individuals with greater reproductive potential, as well as fishing mortality of immature individuals (Barneche et al. 2018; Ahrens et al. 2020; Marshall et al. 2021). From an ecosystem perspective, there is little empirical evidence on whether the potential impact of recreational spearfishing differs from that of other fishing methods, and this deserves specific research attention in the future.
There is strong evidence that spearfishing increases wariness of exploited fish populations. Human-induced changes of behavior have been shown in response to several fishing techniques spanning from active to passive and from recreational to commercial fishing gear (Arlinghaus et al. 2017b; Diaz Pauli and Sih 2017; Sbragaglia et al. 2021b). However, what is unique in behavioral interactions between spearfishing and fish is that fish wariness is directly linked to underwater human presence (Samia et al. 2019), and that antipredator responses to spearfishers may mimic responses to natural predators as recently demonstrated for snorkelers (Asunsolo-Rivera et al. 2023). The possible consequences of human-induced fish behavioral changes should be carefully quantified from an ecosystem perspective (Gaynor et al. 2019; Wilson et al. 2020; Rahman and Candolin 2022), particularly linking changes in anti-predator behavior to changes in other functionally important behaviors (e.g., movement, feeding, mating). In particular, cumulative impacts on consumptive (Arlinghaus et al. 2016) and non-consumptive (Geffroy et al. 2015; Asunsolo-Rivera et al. 2023) activities should be accounted for in population, community and ecosystem dynamics. This would enable a comprehensive understanding of spearfishing impacts relative to human-induced effects and inform effective management regulations for sustainable fishing, tourism, and conservation.
Alongside possible ecological impacts of spearfishing, there is also evidence suggesting that recreational spearfishing can contribute to advances in marine ecological knowledge. Growing research relationships and networks between recreational spearfishers and researchers can also help to build diverse knowledge systems to navigate future management scenarios of increasing protection of coastal areas and management of recreational spearfishing, which is an important topic (supported by “marine reserve” appearing as important nodes in the co-occurrence analysis; Fig. 4). In this context, the inclusion of recreational spearfishers in participatory processes (e.g., within the framework of co-management systems) may be advisable for two main reasons. Firstly, spearfishers can provide early warning and knowledge about conservation problems where existing monitoring data are insufficient (Gledhill et al. 2015; Zapelini et al. 2017; Sbragaglia et al. 2020a; Middleton et al. 2021; Foster et al. 2023). Secondly, fostering an inclusive process could reduce conflicts and stimulate cooperation, trust, proactive behaviors, and even change existing misperceptions such as those related to systematic harvesting of large, long-lived, and slow-growing species (Gledhill et al. 2015; Horta e Costa et al., 2022; see also "Changes in behavioral phenotypes of fishes" section). In this context, considering that spearfishers are usually younger than other groups of recreational fishers, it is advisable to engage them using social media and digital methods (Allison et al. 2022; Lennox et al. 2022), which could be more effective than traditional engagement strategies. A significant proportion of Australian and Mediterranean recreational spearfishers support self-regulatory actions (Young et al. 2014; Sbragaglia et al. 2016) and perceive that no-take marine protected areas are important tools for the conservation of marine ecosystems (Mann et al. 1997; Assis et al. 2018). Inclusive, participatory and collaborative management is important in the current scenario of increasing protection of marine coastal areas for achieving biodiversity conservation, developing an inclusive blue economy, and increasing acceptance of new regulations (Di Franco et al. 2020; Grorud-Colvert et al. 2021; Horta e Costa et al., 2022). Beyond daily bag limits and size limits, possible alternative management strategies could consist of periodic spatial and/or temporal closures (Goetze et al. 2018; Carvalho et al. 2019). In particular, the wariness of fish could be a good indicator of ecological disturbance of marine recreational spearfishing and could be used to apply periodic spatial and/or temporal closures (Goetze et al. 2017). Thus far, such approaches have mainly been implemented in subsistence fisheries, but they could similarly be applied to recreational spearfishing.
The present and future of recreational spearfishing are facing a change in perceptions, beliefs, and behavior of its practitioners, all of which could support the future sustainability of the sector. For example, the attitudes of some recreational spearfishers are already changing from being a group exclusively interested in “adventure-seeking hunting” to a group that is interested in “sea-appreciating hunting” (i.e., towards sharks and rays on the Great Barrier Reef, Whatmough et al. 2011). Such changes in attitudes are also suggested by an increase in the conservation awareness of recreational spearfishers in the Mediterranean Sea (Sbragaglia and Arlinghaus 2020). Scientists and managers would benefit from encouraging this ongoing transition, towards the spreading of best practices driven by recreational spearfishers that exemplify these new attitudes of marine stewardship. This could be a triggering factor to successfully navigate toward the sustainable development of recreational spearfishing in the future, as has been demonstrated for recreational angling (Fujitani et al. 2017). We hope this review will help to stimulate such transitions and encourage future research on marine recreational fisheries.
Data availability
The full list of papers used for the systematic review is available as a supplementary table.
References
Abrahamsson E, Schagatay E (2014) A living based on breath-hold diving in the bajau laut. Hum Evol 29:171–183
Adel M, Nour OM, Al Mabruk SA, Zava B, Deidun A, Corsini-Foka M (2022) The yellowfin surgeonfish acanthurus xanthopterus valenciennes, 1835 (actinopterygii: Perciformes: Acanthuridae) from mediterranean egyptian waters. Mediterr Mar Sci 23:134–139
Aguilar-Perera A, Carrillo-Barragán J (2019) A poorly known snapper in the gulf of mexico: Cubera, lutjanus cyanopterus,(teleostei: Lutjanidae) off the northern coast of the yucatan peninsula, mexico. Thalassas Int J Mar Sci 35:587–590
Ahrens RNM, Allen MS, Walters C, Arlinghaus R (2020) Saving large fish through harvest slots outperforms the classical minimum-length limit when the aim is to achieve multiple harvest and catch-related fisheries objectives. Fish Fish 21:483–510
Allison C, Winkler AC, Childs AR, Muller C, Potts WM (2022) Can social media platforms be used to foster improved environmental behaviour in recreational fisheries? Fish Res 258:106544
Arechavala-Lopez P, Izquierdo-Gomez D, Forcada A, Fernandez-Jover D, Toledo-Guedes K, Valle C, Sanchez-Jerez P (2018) Recapturing fish escapes from coastal farms in the western mediterranean sea: insights for potential contingency plans. Ocean Coast Manage 151:69–76
Arlinghaus R, Alós J, Klefoth T, Laskowski K, Monk CT, Nakayama S, Schröder A (2016) Consumptive tourism causes timidity, rather than boldness, syndromes: a response to geffroy et al. Trends Ecol Evol 31:92–94
Arlinghaus R, Alós J, Beardmore B, Daedlow K, Dorow M, Fujitani M, Hühn D et al (2017a) Understanding and managing freshwater recreational fisheries as complex adaptive social-ecological systems. Rev Fish Sci Aquac 25:1–41
Arlinghaus R, Laskowski KL, Alós J, Klefoth T, Monk CT, Nakayama S, Schröder A (2017b) Passive gear-induced timidity syndrome in wild fish populations and its potential ecological and managerial implications. Fish Fish 18:360–373
Arlinghaus R, Abbott JK, Fenichel EP, Carpenter SR, Hunt LM, Alós J, Klefoth T et al (2019) Governing the recreational dimension of global fisheries. Proc Natl Acad Sci USA 116:5209–5213
Assis J, Gonçalves JMS, Veiga P, Pita C (2018) Spearfishing in portugal: a baseline study on spearfishers’ profiles, habits and perceptions towards management measures. Fish Manage Ecol 25:417–428
Asunsolo-Rivera A, Lester E, Langlois T, Vaughan B, McCormick MI, Simpson SD, Meekan MG (2023) Behaviour of mesopredatory coral reef fishes in response to threats from sharks and humans. Sci Rep 13:6714
Barbosa MC, Luiz OJ, Cordeiro CA, Giglio VJ, Ferreira CE (2021) Fish and spearfisher traits contributing to catch composition. Fish Res 241:105988
Barbour AB, Allen MS, Frazer TK, Sherman KD (2011) Evaluating the potential efficacy of invasive lionfish (pterois volitans) removals. PLoS ONE 6:e19666
Barcelos LMD, Azevedo J, Pollerspöck J, Barreiros JP (2018) Review of the records of the smalltooth sand tiger shark, odontaspis ferox (elasmobranchii: Lamniformes: Odontaspididae), in the azores. Acta Ichthyol Pisc 48:189–194
Barneche DR, Robertson DR, White CR, Marshall DJ (2018) Fish reproductive-energy output increases disproportionately with body size. Science 360:642–645
Baum JK, Worm B (2009) Cascading top-down effects of changing oceanic predator abundances. J Anim Ecol 78:699–714
Bender MG, Machado GR, Silva PJdA, Floeter SR, Monteiro-Netto C, Luiz OJ, Ferreira CE (2014) Local ecological knowledge and scientific data reveal overexploitation by multigear artisanal fisheries in the southwestern atlantic. PLoS ONE 9:e110332
Benevides LJ, Nunes JdACC, Costa TLA, Sampaio CLS (2016) Flight response of the barber surgeonfish, acanthurus bahianus castelnau, 1855 (teleostei: Acanthuridae), to spearfisher presence. Neotrop Ichthyol 14:e150010
Benevides LJ, Pinto TK, José de Anchieta C, Sampaio CL (2018) Fish escape behavior as a monitoring tool in the largest brazilian multiple-use marine protected area. Ocean Coast Manag 152:154–162
Benoit K, Watanabe K, Wang H, Nulty P, Obeng A, Müller S, Matsuo A (2018) Quanteda: an r package for the quantitative analysis of textual data. J Open Sour Softw 3:774
Berger-Tal O, Polak T, Oron A, Lubin Y, Kotler BP, Saltz D (2011) Integrating animal behavior and conservation biology: a conceptual framework. Behav Ecol 22:236–239
Bergseth BJ, Williamson DH, Frisch AJ, Russ GR (2016) Protected areas preserve natural behaviour of a targeted fish species on coral reefs. Biol Conserv 198:202–209
Boada J, Sagué O, Gordoa A (2017) Spearfishing data reveals the littoral fish communities’ association to coastal configuration. Estuar Coast Shelf Sci 199:152–160
Boaden AE, Kingsford MJ (2015) Predators drive community structure in coral reef fish assemblages. Ecosphere 6:art46
Bradford T, Wolfe K, Mumby PJ (2019) Preferences and perceptions of the recreational spearfishery of the great barrier reef. PLoS ONE 14:e0221855
Bratman GN, Anderson CB, Berman MG, Cochran B, de Vries S, Flanders J, Folke C et al (2019) Nature and mental health: an ecosystem service perspective. Sci Adv 5:eaax0903
Brown JS, Laundré JW, Gurung M (1999) The ecology of fear: optimal foraging, game theory, and trophic interactions. J Mammal 80:385–399
Bulleri F, Benedetti-Cecchi L (2014) Chasing fish and catching data: recreational spearfishing videos as a tool for assessing the structure of fish assemblages on shallow rocky reefs. Mar Ecol Prog Ser 506:255–265
Carvalho PG, Jupiter SD, Januchowski-Hartley FA, Goetze J, Claudet J, Weeks R, Humphries A et al (2019) Optimized fishing through periodically harvested closures. J Appl Ecol 56:1927–1936
Castellanos-Galindo GA, Chong-Montenegro C, Baos ERA, Zapata LA, Tompkins P, Graham RT, Craig M (2018) Using landing statistics and fishers’ traditional ecological knowledge to assess conservation threats to pacific goliath grouper in colombia. Aquat Conserv Mar Freshwat Ecosyst 28:305–314
Cavallaro M, Marino G, and Ammendolia G (2016) First record of a lessepsian migrant, the dusky spinefoot, siganus luridus (rüppell, 1829) in the strait of messina (central mediterranean sea)/prima segnalazione di pesce coniglio (specie migrante lessepsiana), siganus luridus (rüppell, 1829), nello stretto di messina (mediterraneo centrale). In Annales: Series Historia Naturalis, (p 19). Scientific and Research Center of the Republic of Slovenia
Chapman MR, Kramer DL (1999) Gradients in coral reef fish density and size across the barbados marine reserve boundary: effects of reserve protection and habitat characteristics. Mar Ecol Prog Ser 181:81–96
Cheung WWL, Pitcher TJ, Pauly D (2005) A fuzzy logic expert system to estimate intrinsic extinction vulnerabilities of marine fishes to fishing. Biol Conserv 124:97–111
Cinelli F, and Fresi E (1980). Contributo alla valutazione dell'effecttiva incidenza della pesca subacquea sul patrimonio biologico delle acque consiere italiane. Atti; Serie B: Memorie-Societa toscana di scienze naturali pp 330–339
Cisneros-Montemayor AM, Sumaila UR (2010) A global estimate of benefits from ecosystem-based marine recreation: potential impacts and implications for management. J Bioecon 12:245–268
Clauset A, Newman ME, Moore C (2004) Finding community structure in very large networks. Phys Rev E 70:066111
Cohen PJ, Alexander TJ (2013) Catch rates, composition and fish size from reefs managed with periodically-harvested closures. PLoS ONE 8:e73383
Coll J, Linde M, García-Rubies A, Riera F, Grau AM (2004) Spear fishing in the balearic islands (west central mediterranean): species affected and catch evolution during the period 1975–2001. Fish Res 70:97–111
Cooke SJ, Suski CD, Arlinghaus R, Danylchuk AJ (2013) Voluntary institutions and behaviours as alternatives to formal regulations in recreational fisheries management. Fish Fish 14:439–457
Cooke SJ, Twardek WM, Lennox RJ, Zolderdo AJ, Bower SD, Gutowsky LFG, Danylchuk AJ et al (2018) The nexus of fun and nutrition: recreational fishing is also about food. Fish Fish 19:201–224
Cooke SJ, Twardek WM, Reid AJ, Lennox RJ, Danylchuk SC, Brownscombe JW, Bower SD et al (2019) Searching for responsible and sustainable recreational fisheries in the anthropocene. J Fish Biol 94:845–856
Cooke SJ, Venturelli P, Twardek WM, Lennox RJ, Brownscombe JW, Skov C, Hyder K et al (2021) Technological innovations in the recreational fishing sector: implications for fisheries management and policy. Rev Fish Biol Fish 31:253–288
Côté IM, Darling ES, Malpica-Cruz L, Smith NS, Green SJ, Curtis-Quick J, Layman C (2014) What doesn’t kill you makes you wary? Effect of repeated culling on the behaviour of an invasive predator. PLoS ONE 9:e94248
Csardi MG (2013) Package ‘igraph.’ Last Accessed 3:2013
Csardi G, Nepusz T (2006) The igraph software package for complex network research. InterJ Comp Syst 1695:1–9
Curley BG, Glasby TM, Curley AJ, Creese RG, Kingsford MJ (2013) Enhanced numbers of two temperate reef fishes in a small, partial-take marine protected area related to spearfisher exclusion. Biol Conserv 167:435–445
Dahl KA, Patterson WF III (2014) Habitat-specific density and diet of rapidly expanding invasive red lionfish, pterois volitans, populations in the northern gulf of mexico. PLoS ONE 9:e105852
Dahl KA, Patterson WF III, Snyder RA (2016) Experimental assessment of lionfish removals to mitigate reef fish community shifts on northern gulf of mexico artificial reefs. Mar Ecol Prog Ser 558:207–221
Dalzell P, Smith A (1998) Rapid data collection and assessment of the biology of acanthurus nigrofuscus at woleai atoll, micronesia. J Fish Biol 52:386–397
De Groot R, Brander L, Van Der Ploeg S, Costanza R, Bernard F, Braat L, Christie M et al (2012) Global estimates of the value of ecosystems and their services in monetary units. Ecosyst Serv 1:50–61
Dedeu AL, Boada J, Gordoa A (2019) The first estimates of species compositions of spanish marine recreational fishing reveal the activity’s inner and geographical variability. Fish Res 216:65–73
Di Franco A, Hogg KE, Calò A, Bennett NJ, Sévin-Allouet M-A, Esparza Alaminos O, Lang M et al (2020) Improving marine protected area governance through collaboration and co-production. J Environ Manag 269:110757
Diaz Pauli B, Sih A (2017) Behavioural responses to human-induced change: Why fishing should not be ignored. Evol Appl 10:231–240
Dill LM, Heithaus MR, Walters CJ (2003) Behaviorally mediated indirect interactions in marine communities and their conservation implications. Ecology 84:1151–1157
Diogo HMC, Pereira JG (2013) Impact evaluation of spear fishing on fish communities in an urban area of são miguel island (azores archipelago). Fish Manage Ecol 20:473–483
Diogo H, Pereira JG (2014) Assessing the potential biological implications of recreational inshore fisheries on sub-tidal fish communities of azores (north-east atlantic ocean) using catch and effort data. J Fish Biol 84:952–970
Diogo H, Pereira JG, Schmiing M (2017) Experience counts: Integrating spearfishers’ skills and knowledge in the evaluation of biological and ecological impacts. Fish Manage Ecol 24:95–102
Diogo H, Veiga P, Pita C, Sousa A, Lima D, Pereira JG, Gonçalves JMS et al (2020) Marine recreational fishing in portugal: current knowledge, challenges, and future perspectives. Rev Fish Sci Aquac 28:536–560
Dyck AJ, Sumaila UR (2010) Economic impact of ocean fish populations in the global fishery. J Bioecon 12:227–243
Ennis Z, Aiken K (2014) Spearfishing as a potential threat to fishery sustainability in jamaica: a survey of 23 fishing beaches. Rev Biol Trop 62:392–400
Espedido J, Parducho V, Yap M, and Palomares M (2014) The recreational marine sport fisheries catch of the philippines, 1950–2010. Philippine marine fisheries catches: a bottom-up reconstruction: 92–104
Esposito G, Prearo M, Menconi V, Mugetti D, Meloni D, Tomasoni M, Pizzul E et al (2021) Northward spread of the parrotfish sparisoma cretense (teleostei: Scaridae) in the mediterranean sea: an update on current distribution with two new records from sardinia. J Mar Sci Eng 9:536
Estes JE, Smith NS, Palmisano JF (1978) Sea otter predation and community organization in the western aleutian islands, alaska. Ecology 59:822–833
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, Carpenter SR et al (2011) Trophic downgrading of planet earth. Science 333:301–306
Fairclough D, Clarke K, Valesini F, Potter I (2008) Habitat partitioning by five congeneric and abundant choerodon species (labridae) in a large subtropical marine embayment. Estuar Coast Shelf Sci 77:446–456
FAO (2012). Recreational fisheries. Fao technical guidelines for responsible fisheries: Recreational fisheries (food and agricultural organization, rome) Rome
Fedler AJ, Ditton RB (1994) Understanding angler motivations in fisheries management. Fisheries 19:6–13
Foo SA, Walsh WJ, Lecky J, Marcoux S, Asner GP (2021) Impacts of pollution, fishing pressure, and reef rugosity on resource fish biomass in west hawaii. Ecol Appl 31:e2213
Foster GE, Sheel AW (2005) The human diving response, its function, and its control. Scand J Med Sci Sports 15:3–12
Foster RM, Childs AR, Mann BQ, Potts WM (2023) Specialist spearfishers’ ecological knowledge provides accurate information that improves the description of the reproduction of a data-deficient species, Oplegnathus conwayi in South Africa. Fish Res 257:106513
Frisch AJ, Baker R, Hobbs JPA, Nankervis L (2008) A quantitative comparison of recreational spearfishing and linefishing on the great barrier reef: implications for management of multi-sector coral reef fisheries. Coral Reefs 27:85–95
Frisch AJ, Cole AJ, Hobbs J-PA, Rizzari JR, Munkres KP (2012) Effects of spearfishing on reef fish populations in a multi-use conservation area. PLoS ONE 7:e51938
Fujitani M, McFall A, Randler C, Arlinghaus R (2017) Participatory adaptive management leads to environmental learning outcomes extending beyond the sphere of science. Sci Adv 3:e1602516
Gaynor KM, Brown JS, Middleton AD, Power ME, Brashares JS (2019) Landscapes of fear: spatial patterns of risk perception and response. Trends Ecol Evol 34:355–368
Geffroy B, Samia DS, Bessa E, Blumstein DT (2015) How nature-based tourism might increase prey vulnerability to predators. Trends Ecol Evol 30:755–765
Giddens J, Friedlander AM, Conklin E, Wiggins C, Stamoulis K, Donovan MK (2014) Experimental removal of the invasive peacock hind (roi) cephalopholis argus, in puakō, hawai ‘i: Methods for assessing and managing marine invasive species. Mar Ecol Prog Ser 511:209–221
Giglio VJ, Bender MG, Zapelini C, Ferreira CEL (2017) The end of the line? Rapid depletion of a large-sized grouper through spearfishing in a subtropical marginal reef. Perspect Ecol Conserv 15:115–118
Giglio VJ, Luiz OJ, Barbosa MC, Ferreira CEL (2018) Behaviour of recreational spearfishers and its impacts on corals. Aquat Conserv Mar Freshwat Ecosyst 28:167–174
Giglio VJ, Suhett AC, Zapelini CS, Ramiro AS, Quimbayo JP (2020) Assessing captures of recreational spearfishing in abrolhos reefs, brazil, through social media. Regional Studies in Marine Science 34:100995
Gill JA, Norris K, Sutherland WJ (2001) Why behavioural responses may not reflect the population consequences of human disturbance. Biol Conserv 97:265–268
Gledhill DC, Hobday AJ, Welch DJ, Sutton SG, Lansdell MJ, Koopman M, Jeloudev A et al (2015) Collaborative approaches to accessing and utilising historical citizen science data: a case-study with spearfishers from eastern australia. Mar Freshw Res 66:195–201
Godoy N, Gelcich S, Vásquez JA, Castilla JC (2010) Spearfishing to depletion: evidence from temperate reef fishes in chile. Ecol Appl 20:1504–1511
Goetze JS, Januchowski-Hartley FA, Claudet J, Langlois TJ, Wilson SK, Jupiter SD (2017) Fish wariness is a more sensitive indicator to changes in fishing pressure than abundance, length or biomass. Ecol Appl 27:1178–1189
Goetze JS, Claudet J, Januchowski-Hartley F, Langlois TJ, Wilson SK, White C, Weeks R et al (2018) Demonstrating multiple benefits from periodically harvested fisheries closures. J Appl Ecol 55:1102–1113
Gordoa A, Dedeu AL, Boada J (2019) Recreational fishing in spain: first national estimates of fisher population size, fishing activity and fisher social profile. Fish Res 211:1–12
Grorud-Colvert K, Sullivan-Stack J, Roberts C, Constant V, Horta e Costa B, Pike EP, Kingston N et al (2021) The mpa guide: a framework to achieve global goals for the ocean. Science 373:eabf0861
Guabiroba HC, Santos MEA, Pinheiro HT, Simon T, Pimentel CR, Vilar CC, Joyeux J-C (2020) Trends in recreational fisheries and reef fish community structure indicate decline in target species population in an isolated tropical oceanic island. Ocean Coast Manag 191:105194
Hall AE, Cameron DS, Kingsford MJ (2021) Partially protected areas as a management tool on inshore reefs. Rev Fish Biol Fish 31:631–651
Hall A, Cameron D, Kingsford M (2022) Prohibiting spearfishing boosts conservation outcomes for partially protected areas. Biol Conserv 272:109662
Hamilton R, Giningele M, Aswani S, Ecochard J (2012) Fishing in the dark-local knowledge, night spearfishing and spawning aggregations in the western solomon islands. Biol Conserv 145:246–257
Harmelin-Vivien M, Cottalorda J-M, Dominici J-M, Harmelin J-G, Le Diréach L, Ruitton S (2015) Effects of reserve protection level on the vulnerable fish species sciaena umbra and implications for fishing management and policy. Global Ecol Conserv 3:279–287
Harper DE, Bohnsack JA, and Lockwood BR (2000) Recreational fisheries in biscayne national park, florida, 1976–19.
Harris HE, Patterson WF, Ahrens RNM, Allen MS (2019) Detection and removal efficiency of invasive lionfish in the northern gulf of mexico. Fish Res 213:22–32
Harris HE, Fogg AQ, Allen MS, Ahrens RNM, Patterson WF (2020) Precipitous declines in northern gulf of mexico invasive lionfish populations following the emergence of an ulcerative skin disease. Sci Rep 10:1934
Herfaut J, Levrel H, Thébaud O, Véron G (2013) The nationwide assessment of marine recreational fishing: a french example. Ocean Coast Manag 78:121–131
Horta e Costa B, Guimarães MH, Rangel M, Ressurreição A, Monteiro P, Oliveira F, Bentes L et al (2022) Co-design of a marine protected area zoning and the lessons learned from it. Front Mar Sci 9:969234
Humphries AT, Gorospe KD, Carvalho PG, Yulianto I, Kartawijaya T, Campbell SJ (2019) Catch composition and selectivity of fishing gears in a multi-species indonesian coral reef fishery. Front Mar Sci 6:378
Hunt L, Sutton S, Arlinghaus R (2013) Illustrating the critical role of human dimensions research for understanding and managing recreational fisheries within a social-ecological system framework. Fish Manage Ecol 20:111–124
Hutchings K, Griffiths M (2010a) Assessment of south african umbrina robinsoni based on per-recruit models. Afr J Mar Sci 32:511–523
Hutchings K, Griffiths M (2010b) Life-history strategies of umbrina robinsoni (sciaenidae) in warm-temperate and subtropical south african marine reserves. Afr J Mar Sci 32:37–53
Hyder K, Weltersbach MS, Armstrong M, Ferter K, Townhill B, Ahvonen A, Arlinghaus R et al (2018) Recreational sea fishing in Europe in a global context-participation rates, fishing effort, expenditure, and implications for monitoring and assessment. Fish Fish 19:225–243
Ilardo MA, Moltke I, Korneliussen TS, Cheng J, Stern AJ, Racimo F, de Barros DP et al (2018) Physiological and genetic adaptations to diving in sea nomads. Cell 173:569-580.e515
Irigoyen-Arredondo MS, Moreno-Sánchez XG, Escobar-Sánchez O, Soto-Jiménez MF, Marín-Enríquez E, Abitia-Cárdenas LA (2022) Essential (cu, zn) and nonessential (pb, cd) metals in the muscle of leopard groupers (mycteroperca rosacea) from a mining port in the gulf of california, mexico: human health risk assessment. Environ Sci Pollut Res 29(23):35001–35011
Januchowski-Hartley FA, Graham NAJ, Feary DA, Morove T, Cinner JE (2011) Fear of fishers: human predation explains behavioral changes in coral reef fishes. PLoS ONE 6:e22761
Januchowski-Hartley FA, Graham NAJ, Cinner JE, Russ GR (2013) Spillover of fish naïveté from marine reserves. Ecol Lett 16:191–197
Januchowski-Hartley FA, Cinner JE, Graham NAJ (2014) Fishery benefits from behavioural modification of fishes in periodically harvested fisheries closures. Aquat Conserv Mar Freshwat Ecosyst 24:777–790
Januchowski-Hartley SR, Bear C, O'Gorman E, and Januchowski-Hartley FA (2020) Underwater. A-Z of shadow places concepts pp 1–5
Jimenez C, Aguilar-Perera A, Dimitriou AC, Papatheodoulou M, Patsalou P (2022) Saddleback syndrome in the common lionfish pterois miles (scorpaeniformes: Scorpaenidae), off cyprus, eastern mediterranean sea. Thalassas Int J Mar Sci 38(1):487–492
Jiménez-Alvarado D, Guerra-Marrero A, Sarmiento-Lezcano A, Meyers EKM, Castro JJ (2020) First assessment of the spearfishing impact in the canary islands. Reg Stud Mar Sci 38:101385
Johnson DW, Stirling BS, Paz J, Satterfield DR (2019) Geographic variation in demography of black perch (embiotoca jacksoni): effects of density, food availability, predation, and fishing. J Exp Mar Biol Ecol 516:16–24
Jouvenel JY, Pollard D (2001) Some effects of marine reserve protection on the population structure of two spearfishing target-fish species, dicentrarchus labrax (moronidae) and sparus aurata (sparidae), in shallow inshore waters, along a rocky coast in the northwestern mediterranean sea. Aquat Conserv Mar Freshwat Ecosyst 11:1–9
Last PR, White WT, Gledhill DC, Hobday AJ, Brown R, Edgar GJ, Pecl G (2011) Long-term shifts in abundance and distribution of a temperate fish fauna: a response to climate change and fishing practices. Global Ecol Biogeogr 20:58–72
Lee LC, Thorley J, Watson J, Reid M, Salomon AK (2019) Diverse knowledge systems reveal social–ecological dynamics that inform species conservation status. Conserv Lett 12:e12613
Lennox R, Sbragaglia V, Vollset K, Sortland L, McClenachan L, Jarić I, Guckian M et al (2022) Digital fisheries data in the internet age: emerging tools for research and monitoring using online data in recreational fisheries. Fish Fish 23:926–940
Lewin W-C, Weltersbach MS, Ferter K, Hyder K, Mugerza E, Prellezo R, Radford Z et al (2019) Potential environmental impacts of recreational fishing on marine fish stocks and ecosystems. Rev Fish Sci Aquac 27:287–330
Lincoln Smith M, Bell J, Pollard D, Russell B (1989) Catch and effort of competition spearfishermen in South eastern Australia. Fish Res 8:45–61
Lindfield SJ, McIlwain JL, Harvey ES (2014) Depth refuge and the impacts of scuba spearfishing on coral reef fishes. PLoS ONE 9:e92628
Lindholm P, Lundgren CE (2009) The physiology and pathophysiology of human breath-hold diving. J Appl Physiol 106:284–292
Lloret J, Font T (2013) A comparative analysis between recreational and artisanal fisheries in a Mediterranean coastal area. Fish Manage Ecol 20:148–160
Lloret J, Zaragoza N, Caballero D, Font T, Casadevall M, Riera V (2008) Spearfishing pressure on fish communities in rocky coastal habitats in a Mediterranean marine protected area. Fish Res 94:84–91
Lloyd P, Plaganyi EE, Weeks SJ, Magno-canto M, and Plaganyi G, (2012) Ocean warming alters species abundance patterns and increases species diversity in an African sub-tropical reef-fish community. Fish Oceanogr 21:78–94
Lowry M, Suthers I (2004) Population structure of aggregations, and response to spear fishing, of a large temperate reef fish Cheilodactylus fuscus. Mar Ecol Prog Ser 273:199–210
Lozano S, Calzada-Infante L, Adenso-Díaz B, García S (2019) Complex network analysis of keywords co-occurrence in the recent efficiency analysis literature. Scientometrics 120:609–629
Malpica-Cruz L, Chaves LCT, Côté IM (2016) Managing marine invasive species through public participation: lionfish derbies as a case study. Mar Policy 74:158–164
Manfredo MJ, Driver BL, Tarrant MA (1996) Measuring leisure motivation: a meta-analysis of the recreation experience preference scales. J Leis Res 28:188–213
Mann B, Scott G, Mann-Lang J, Brouwer S, Lamberth S, Sauer W, Erasmus C (1997) An evaluation of participation in and management of the south african spearfishery. S Afr J Mar Sci 18:179–193
Marengo M, Culioli J-M, Santoni M-C, Marchand B, Durieux EDH (2015) Comparative analysis of artisanal and recreational fisheries for dentex dentex in a marine protected area. Fish Manage Ecol 22:249–260
Marshall DJ, Bode M, Mangel M, Arlinghaus R, Dick EJ (2021) Reproductive hyperallometry and managing the world’s fisheries. Proc Natl Acad Sci 118:e2100695118
Martínez-Escauriaza R, Vieira C, Gouveia L, Gouveia N, Hermida M (2020) Characterization and evolution of spearfishing in madeira archipelago, eastern atlantic. Aquat Living Resour 33:15
Martín-Sosa P (2019) Spearfishing in the canary islands: Is the devil as black as it seems to be? Sci Insul 2:9–36
Maya-Jariego I, Martínez-Alba I, Alieva D (2022) Responsible practice of recreational spearfishing is needed. A response to sbragaglia et al.(2021). Mar Policy 139:105022
Meyer CG (2007) The impacts of spear and other recreational fishers on a small permanent marine protected area and adjacent pulse fished area. Fish Res 84:301–307
Michailidis N, Katsanevakis S, Chartosia N (2020) Recreational fisheries can be of the same magnitude as commercial fisheries: the case of cyprus. Fish Res 231:105711
Middleton I, Aguirre JD, Trnski T, Francis M, Duffy C, Liggins L (2021) Introduced alien, range extension or just visiting? Combining citizen science observations and expert knowledge to classify range dynamics of marine fishes. Divers Distrib 27:1278–1293
Morales-Nin B, Moranta J, García C, Tugores MP, Grau AM, Riera F, Cerdà M (2005) The recreational fishery off majorca island (western mediterranean): some implications for coastal resource management. ICES J Mar Sci 62:727–739
Nunes JACC, Medeiros DV, Reis-Filho JA, Sampaio CLS, Barros F (2012) Reef fishes captured by recreational spearfishing on reefs of bahia state, northeast brazil. Biota Neotrop 12:179–185
Nunes JACC, Sampaio CLS, Barros F (2015) The influence of structural complexity and reef habitat types on flight initiation distance and escape behaviors in labrid fishes. Mar Biol 162:493–499
Nunes JACC, Loiola M, Miranda RJ, Sampaio CL, Barros F (2016) Are abrolhos no-take area sites of naïve fish? An evaluation using flight initiation distance of labrids. Neotrop Ichthyol.
Nunes JACC, Blumstein DT, Giglio VJ, Barros F, Quimbayo JP (2019) Reef fish antipredator behavior in remote islands does not reflect patterns seen in coastal areas. Ethol Ecol Evol 31:557–567
Ordines F, Deudero S, Sintes-Vila J, Sbragaglia V, Fricke R, Azzurro E (2018) A new record of diodon hystrix (actinopterygii: Tetraodontiformes: Diodontidae) in the mediterranean sea. Acta Ichthyol Pisc 48:403–407
Palacios-Salgado DS, Ramírez-Valdez A, Rojas-Herrera AA, Amores JG, Melo-García MA (2014) Marine fishes of acapulco, mexico (eastern pacific ocean). Mar Biodivers 44:471–490
Papadopoulos A, Touloumis K, Tziolas E, Boulamatsis D, Koutrakis E (2022) Evaluation of marine recreational fisheries and their relation to sustainability of fisheries resources in greece. Sustainability 14:3824
Parkkila K, Arlinghaus R, Artell J, Gentner B, Haider W, Aas O, Barton D, et al (2010) Methodologies for assessing socio-economic benefits of european inland recreational fisheries: European inland fisheries advisory commission. EIFAC Occasional Paper Nº 46. Ankara, FAO, p 112
Pavlowich T, Kapuscinski AR (2017) Understanding spearfishing in a coral reef fishery: fishers’ opportunities, constraints, and decision-making. PLoS ONE 12:e0181617
Pereira PHC, Macedo CHR, de Lima GV, de Jesus BL (2020) Effects of depth on reef fish flight initiation distance: Implications of deeper reefs conservation. Environ Biol Fishes 103:1247–1256
Pita P, Freire J (2014) The use of spearfishing competition data in fisheries management: evidence for a hidden near collapse of a coastal fish community of galicia (ne atlantic ocean). Fish Manage Ecol 21:454–469
Pita P, Freire J (2016) Assessing the impact of spear fishing by using competitions records and underwater visual censuses. Sci Mar 80:27–38
Pita P, Hyder K, Gomes P, Pita C, Rangel M, Veiga P, Vingada J et al (2018) Economic, social and ecological attributes of marine recreational fisheries in galicia, spain. Fish Res 208:58–69
Pretty J, Hine R, Peacock J (2006) Green exercise: the benefits of activities in green places-little has been said about the potential emotional or health benefits of the natural environment in arguments about conservation. Yet Biologist-London 53:143–148
R (Core Team 2018). R: A language and environment for statistical computing. Ed. by RFfS Computing. Vienna
Rahman T, Candolin U (2022) Linking animal behavior to ecosystem change in disturbed environments. Front Ecol Evol 101:893453
Rhoades OK, Lonhart SI, Stachowicz JJ (2019) Human-induced reductions in fish predator boldness decrease their predation rates in kelp forests. Proc Royal Soc b Biol Sci 286:20182745
Rhodes KL, Hernandez-Ortiz DX, Cuetos-Bueno J, Ioanis M, Washington W, Ladore R (2018) A 10-year comparison of the pohnpei, micronesia, commercial inshore fishery reveals an increasingly unsustainable fishery. Fish Res 204:156–164
Rizgalla J, Shinn AP, Ferguson HW, Paladini G, Jayasuriya NS, Bron JE (2017) A novel use of social media to evaluate the occurrence of skin lesions affecting wild dusky grouper, epinephelus marginatus (lowe, 1834), in libyan coastal waters. J Fish Dis 40:609–620
Rocklin D, Tomasini J-A, Culioli J-M, Pelletier D, Mouillot D (2011) Spearfishing regulation benefits artisanal fisheries: the regs indicator and its application to a multiple-use mediterranean marine protected area. PLoS ONE 6:e23820
Roos NC, Longo GO (2021) Critical information for fisheries monitoring may be available in social media. Aquat Conserv Mar Freshwat Ecosyst 31:2420–2428
Russell B (1983) The food and feeding habits of rocky reef fish of north-eastern new zealand. N z J Mar Freshwat Res 17:121–145
Samia DSM, Bessa E, Blumstein DT, Nunes JACC, Azzurro E, Morroni L, Sbragaglia V et al (2019) A meta-analysis of fish behavioural reaction to underwater human presence. Fish Fish 20:817–829
Sangil C, Martín-García L, Clemente S (2013) Assessing the impact of fishing in shallow rocky reefs: a multivariate approach to ecosystem management. Mar Pollut Bull 76:203–213
Sbragaglia V, Arlinghaus R (2020) Conservation bottom-up initiatives in marine recreational spearfishing suggest the emergence of positive attitudes towards conservation. Sci Mar 84:441–444
Sbragaglia V, Pla OS, Gordoa A, Hernández SP, Elias LC, Pulido M, Ruiz RT et al (2016) The barcelona agreement: a manifesto towards the spearfishing of the future. Sci Mar 80:423–426
Sbragaglia V, Morroni L, Bramanti L, Weitzmann B, Arlinghaus R, Azzurro E (2018) Spearfishing modulates flight initiation distance of fishes: the effects of protection, individual size, and bearing a speargun. ICES J Mar Sci 75:1779–1789
Sbragaglia V, Cerri J, Bolognini L, Dragićević B, Dulćić J, Grati F, Azzurro E (2020a) Local ecological knowledge of recreational fishers reveals different meridionalization dynamics of two mediterranean subregions. Mar Ecol Prog Ser 634:147–157
Sbragaglia V, Correia RA, Coco S, Arlinghaus R (2020b) Data mining on youtube reveals fisher group-specific harvesting patterns and social engagement in recreational anglers and spearfishers. ICES J Mar Sci 77:2234–2244
Sbragaglia V, Coco S, Correia RA, Coll M, Arlinghaus R (2021a) Analyzing publicly available videos about recreational fishing reveals key ecological and social insights: a case study about groupers in the mediterranean sea. Sci Total Environ 765:142672
Sbragaglia V, Jolles JW, Coll M, Arlinghaus R (2021b) Fisheries-induced changes of shoaling behaviour: mechanisms and potential consequences. Trend Ecol Evol 36:885–888
Sbragaglia V, Espasandín L, Coco S, Felici A, Correia RA, Coll M, Arlinghaus R (2022) Recreational angling and spearfishing on social media: insights on harvesting patterns, social engagement and sentiments related to the distributional range shift of a marine invasive species. Rev Fish Biol Fish 32:687–700
Scheffer M, Carpenter S, Young Bd (2005) Cascading effects of overfishing marine systems. Trends Ecol Evol 20:579–581
Schilling HT, Riley IJ, Puckeridge AC, Milne-Muller A, Callaghan CT (2022) Economic value of regional spearfishing competitions. Fish Res 250:106289
Schroeder RE, Parrish JD (2005) Resilience of predators to fishing pressure on coral patch reefs. J Exp Mar Biol Ecol 321:93–107
Shephard S, List CJ, and Arlinghaus R (in press). Reviving the unique potential of recreational fishers as environmental stewards of aquatic ecosystems. Fish Fish, n/a.
Skinner C, Newman SP, Box S, Narozanski A, Polunin NV (2019) Chronic spearfishing may indirectly affect reef health through reductions in parrotfish bite rates. J Fish Biol 94:585–594
Skov C, Berg S, Eigaard OR, Jessen TK, Skov PV (2020) Danish fisheries and aquaculture: past, present, and future. Fisheries 45:33–41
Sluka RD, Sullivan KM (1998) The influence of spear fishing on species composition arid size of groupers on patch reefs in the upper florida keys. Fish Bull 96:388–392
Stamoulis KA, Harvey ES, Friedlander AM, Williams ID, Weng KC, Wiggins C, Wagner GW et al (2019) Flight behavior of targeted fishes depends on variables other than fishing. Ecol Indic 96:579–590
Stankowich T, Blumstein DT (2005) Fear in animals: a meta-analysis and review of risk assessment. Proc Royal Soc b Biol Sci 272:2627–2634
Ten Brink TS, Dalton T (2018) Perceptions of commercial and recreational fishers on the potential ecological impacts of the block island wind farm (US). Front Mar Sci 5:439
Terlizzi A, Tarantino G, Sbragaglia V (2022) Ecological, social and economic aspects of italian marine spearfishing tournaments (2009–2020). Front Mar Sci 9:891246
Tiralongo F, Lipari R, Mancini E (2018) A new exotic fish for the mediterranean sea: Chaetodon auriga forsskål, 1775 (perciformes: Chaetodontidae). Mediterr Mar Sci 19:491–493
Tran DSC, Langel KA, Thomas MJ, Blumstein DT (2016) Spearfishing-induced behavioral changes of an unharvested species inside and outside a marine protected area. Curr Zool 62:39–44
Varveri D, Karatzaferi C, Pollatou E, Sakkas GK (2016) Aquaticity: a discussion of the term and of how it applies to humans. J Bodyw Mov Ther 20:219–223
Vitale G, Dedeu AL, Pujol M, Sbragaglia V (2021) Characterizing the profile of recreational fishers who share their catches on social media. Front Mar Sci 8:768047
Welch DJ, Mapstone BD, Davies CR, Russ GR (2010) Spatial and fishing effects on sampling gear biases in a tropical reef line fishery. Mar Freshw Res 61:1134–1146
Whatmough S, Van Putten I, Chin A (2011) From hunters to nature observers: a record of 53 years of diver attitudes towards sharks and rays and marine protected areas. Mar Freshw Res 62:755–763
Wilson MW, Ridlon AD, Gaynor KM, Gaines SD, Stier AC, Halpern BS (2020) Ecological impacts of human-induced animal behaviour change. Ecol Lett 23:1522–1536
Ydenberg RC, Dill LM (1986) The economics of fleeing from predators. Adv Study Behav 16:229–249
Yellen J, Brooks A, Cornelissen E, Mehlman M, Stewart K (1995) A middle stone age worked bone industry from katanda, upper semliki valley, zaire. Science 268:553–556
Young MAL, Foale S, Bellwood DR (2014) Impacts of recreational fishing in australia: historical declines, self-regulation and evidence of an early warning system. Environ Conserv 41:350–356
Young MA, Foale S, Bellwood DR (2015a) Dynamic catch trends in the history of recreational spearfishing in australia. Conserv Biol 29:784–794
Young MAL, Foale S, Bellwood DR (2015b) The last marine wilderness: spearfishing for trophy fishes in the coral sea. Environ Conserv 43:90–95
Young MAL, Foale S, Bellwood DR (2016) Why do fishers fish? A cross-cultural examination of the motivations for fishing. Mar Policy 66:114–123
Zapelini C, Giglio VJ, Carvalho RC, Bender MG, Gerhardinger LC (2017) Assessing fishing experts’ knowledge to improve conservation strategies for an endangered grouper in the southwestern atlantic. J Ethnobiol 37:478–493
Zarauz L, Ruiz J, Urtizberea A, Andonegi E, Mugerza E, Artetxe I (2015) Comparing different survey methods to estimate european sea bass recreational catches in the basque country. ICES J Mar Sci 72:1181–1191
Zeller D, Stoute S, Russ G (2003) Movements of reef fishes across marine reserve boundaries: effects of manipulating a density gradient. Mar Ecol Prog Ser 254:269–280
Acknowledgements
We are grateful to Marta Coll, Arnau L. Dedeu, Nikolas Michailidis for their help. We acknowledge Marco Bardi for providing some of the pictures used in Figure 1. VS was supported by a “Juan de la Cierva Incorporación” (IJC2018-035389-I), and he is now supported by a “Ramón y Cajal” research fellowships (RYC2021-033065-I) granted by the Spanish Ministry of Science and Innovation. SV and PP are supported by the Xunta de Galicia (RECREGES I and II projects under Grants ED481B2014/034-0 and ED481B2018/017), Grupo de Referencia Competitiva GI-2060 AEMI, under Grant ED431C2019/11), and Fundación Biodiversidad, Ministerio para la Transición Ecológica y el Reto Demográfico, Gobierno de España (SICORE and GT PMR projects). VJG received a postdoctoral grant (#2017/22273-0) from São Paulo Research Foundation (FAPESP). MR would like to acknowledge Portuguese national funds from FCT—Foundation for Science and Technology through projects UIDB/04326/2020, UIDP/04326/2020 and LA/P/0101/2020. MR would also like to acknowledge FCT funding through a postdoctoral grant (SFRH/BPD/116307/2016). JACCN thanks to Meros do Brasil Project sponsored by Petrobras. FJH is supported by a Sȇr Cymru European Regional Development Fund Fellowship (80761-SU-135). RA was supported by the German Federal Ministry of Education and Research (Grants 01LC1826E and 033W046A). ML is supported by the Agencia Nacional de Investigación e Innovación (ANII, POS_EXT_2020_1_165362). This work acknowledges the ‘Severo Ochoa Centre of Excellence’ accreditation (CEX2019-000928-S).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sbragaglia, V., Arlinghaus, R., Blumstein, D.T. et al. A global review of marine recreational spearfishing. Rev Fish Biol Fisheries 33, 1199–1222 (2023). https://doi.org/10.1007/s11160-023-09790-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-023-09790-7