
Abstract
Pacific lamprey Entosphenus tridentatus has an anadromous life cycle that begins with larvae that filter-feed in freshwater, followed by transformation into juveniles that migrate to the ocean where they parasitize hosts, and adults that migrate into freshwater to spawn and die. The marine-phase (i.e., juvenile life stage) is important yet poorly understood, and is associated with growth opportunities to achieve maximum body size and recruitment. The four goals of this paper are to: (1) synthesize the literature to identify patterns in the marine biology of Pacific lamprey; (2) develop hypotheses to explain these patterns; (3) identify limiting factors and threats, and (4) identify research needs. We hypothesize that recruitment of adult lamprey to spawning populations is influenced by oceanographic regimes through impacts on host abundance. Three marine factors that may be limiting lamprey abundance include: (1) predation and fisheries bycatch; (2) host availability; and (3) host contaminant loads. Four potential marine-related threats to lamprey include: (1) pollution; (2) climate change; (3) unfavorable oceanographic regimes; and (4) the effects of interactions between climate and regimes. Pacific lamprey is not philopatric and the extent to which host migrations and other factors influence lamprey entry into rivers is unclear. Research is needed to fill information gaps on how, when, why, and where lamprey move, feed, and grow in the ocean. Their widespread distribution, parasitic life history, diverse hosts, and multiple predators suggest that Pacific lamprey is integrated into diverse marine ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Anadromous lampreys (Petromyzontiformes) have complex life cycles that rely on a succession of freshwater and marine habitats (Hardisty 2006). Larval lamprey emerge from nests in rivers and streams after a few weeks of egg incubation and disperse downstream to depositional areas with soft silt and sand, where they burrow, filter feed, and grow (Fig. 1; Dawson et al. 2015; Sutton 2017). After at least 3 years rearing in freshwater, a combination of physiological and environmental signals cue larvae to transform from eyeless, toothless filter feeders into eyed, sharp-toothed juveniles that emigrate downstream into estuaries and the ocean (e.g., Fig. 1; Dawson et al. 2015; Manzon et al. 2015).

Life cycle of the Pacific lamprey. This paper focuses on the juvenile ocean parasite life stage. Image adapted and used with permission from Jeremy Five Crows, Columbia River Inter-Tribal Fish Commission
These fishes rear as juveniles in estuaries and oceans where they parasitize large vertebrates and consume their blood, fluids, and flesh (e.g., Fig. 1; Renaud et al. 2009a; Renaud 2011). At some point during their marine phase, anadromous lampreys stop feeding and migrate into freshwater streams where they spawn and die during spring-early summer (Clemens et al. 2010; Renaud 2011; Johnson et al. 2015).
The anadromous Pacific lamprey Entosphenus tridentatus is a culturally important species that is used by local indigenous peoples for food, medicine, and ceremony (Close et al. 2002, 2004; Petersen-Lewis 2009). This species is also an important ecological connector to many other organisms throughout the food web. For example, as larvae, they filter feed on diatoms and provide a food source for various predators (Beamish 1980b; Downey et al. 1996; Close et al. 2002). Adult lamprey migrating into rivers provide marine-derived nutrients to terrestrial ecosystems (Nislow and Kynard 2009; Wipfli and Baxter 2010).
Pacific lamprey faces numerous limiting factors and threats to persistence (CRITFC 2011; Luzier et al. 2011; Maitland et al. 2015; Clemens et al. 2017a). The abundance of adult Pacific lamprey migrating into the Columbia River Basin (USA) has declined over the last 50+ years (CRITFC 2011). However, abundance trends for adult Pacific lamprey in freshwater is highly variable across its distribution (Murauskas et al. 2016). Due to declines in abundance and perceived threats, Pacific lamprey is a federal species of concern in the U.S. (USFWS 2018). In 2003, a petition to list Pacific lamprey under the Endangered Species Act (ESA 1973) was unsuccessful. The 90-day finding by the U.S. Fish and Wildlife Service concluded that listing was not justified due to a paucity of information on biology, ecology, and threats to Pacific lamprey at any life stage (USFWS 2004). A recent status assessment indicated that Pacific lamprey were at “high conservation risk” in most watersheds in the U.S. (Wang and Schaller 2015). Pacific lamprey is “apparently secure” in Canada (Renaud et al. 2009b), “critically endangered” where status has been determined in Japan (Hokkaido Prefecture 2000; Tochigi Prefecture 2005), and has no conservation status in Russia (Dyldin and Orlov 2016).
Recent efforts have been made to improve understanding of the biology of Pacific lamprey, but most have focused on freshwater life stages. For example, recent research has concentrated on spawning (Stone 2006; Brumo et al. 2009; Gunckel et al. 2009; Johnson et al. 2015; Whitlock et al. 2017), embryonic development (Meeuwig et al. 2005; Lampman et al. 2016), development and ecology of larvae (McGree et al. 2008; Jolley et al. 2012, 2015; Schultz et al. 2014; Harris and Jolley 2017), ecology of emigrating juveniles (Moser et al. 2015b), ecology of adults migrating in freshwater (Moser et al. 2015a; Clemens et al. 2016, 2017b), and limiting factors in freshwater systems (Maitland et al. 2015; Clemens et al. 2017a).
By contrast, information on the marine phase (juvenile life stage) of the Pacific lamprey is sparse (Clemens et al. 2010). Information on some aspects of the marine biology of Pacific lamprey was presented nearly 40 years ago (Beamish 1980b). Since that time, a significant amount of research has occurred, enabling a more thorough and wider-ranging synthesis of the marine biology of Pacific lamprey. The four goals of this paper are to: (1) synthesize the peer-reviewed literature pertaining to the marine biology of Pacific lamprey to identify patterns; (2) develop hypotheses to explain these patterns; (3) identify potential limiting factors and threats in the estuary and ocean; and (4) identify research needs in the estuary and ocean. This review covers all aspects of the marine biology of Pacific lamprey, including their freshwater/marine transitions; distribution and movements; genetic population structure; feeding behavior and hosts; abundance, recruitment, and growth rates; predators; life-history diversity; and marine-related limiting factors and threats. Much of the information we synthesize comes from a few geographic areas, particularly the Columbia River estuary, USA, and estuarine habitat near Vancouver Island, Canada. In the absence of extensive data across the species’ range, we assume that information from the Pacific Northwest of North America applies to Pacific lamprey throughout its range. A comprehensive synthesis of the literature on the marine biology of Pacific lamprey is essential to identifying potential management actions and research that will help this species persist in a changing and uncertain future.
Marine biology
Freshwater and marine transitions and timing
Juveniles generally enter seawater in winter through spring, in association with high river discharge (Beamish 1980b; Beamish and Levings 1991; van de Wetering 1999; Moyle 2002; Weitkamp et al. 2015). Within this general pattern, the transformation timing of filter-feeding larvae into parasitic juveniles and subsequent ocean entry varies by latitude among river systems along the West Coast of North America, and also by the size of the river basin. Pacific lamprey generally transform earlier in California (November–May; Goodman et al. 2015) than they do in Oregon and British Columbia (July–December; Beamish 1980b; McGree et al. 2008). Pacific lamprey also emigrate earlier in California (winter–spring; Moyle 2002) than they do in British Columbia (spring–summer; Beamish 1980b; Beamish and Levings 1991). Transformation timing is intermediate in Oregon and Washington (early winter–spring; Kostow 2002; Hayes et al. 2013; Weitkamp et al. 2015). Notable exceptions to these patterns exist; for example, some juvenile Pacific lamprey may emigrate throughout the year (Hayes et al. 2013; Moser et al. 2015b). Basin size may also influence outmigration timing, with earlier (autumn–winter) outmigration occurring in small rivers than in larger systems (winter–spring; Kan 1975; van de Wetering 1999). Further research is needed to elucidate the influence of latitude, river size, river discharge, temperature, and other factors on outmigration timing.
Adult Pacific lamprey can migrate into freshwater year round (Beamish 1980b; Chase 2001; Moyle 2002; Weitkamp et al. 2015; Parker et al. 2019). The pre-spawning, upstream migration of adult Pacific lamprey purportedly occurs earlier in southern latitudes in comparison with northern latitudes, similar to anadromous sea lamprey Petromyzon marinus in North America (reviewed in Clemens et al. 2010). These latitudinal differences in freshwater entry may be a function of water temperature and the distance to the spawning grounds (reviewed in Clemens et al. 2010). In addition, lampreys appear to be attracted to the amount of river discharge (reviewed in Clemens et al. 2016), and Pacific lamprey can migrate further upstream in years when discharge is greater (Clemens et al. 2017b). We hypothesize that the earlier upstream migration timing in southern latitudes may be related to the timing of peak river discharges or river temperatures along the West Coast of North America.
An additional factor that complicates cross-basin comparisons of river entry time is how far upstream the migration timing was documented. Pacific lamprey in the Columbia River may take 2–5 months to traverse the 235 km from the river mouth to Bonneville Dam (Weitkamp et al. 2015). This relatively slow upstream migration rate (< 4 km/day), if typical of other systems, suggests that estimates of “river entry” made upstream of the river mouth may include substantial time while the fish were already in-river. The freshwater migration distance and collection season can also affect the likelihood of observing a stream-maturing or ocean-maturing Pacific lamprey (Parker et al. 2019). Documentation of adult Pacific lamprey immigration into estuaries would help delineate regional patterns of freshwater entry timing.
Based on laboratory observations, the duration of ocean residence by Pacific lamprey was hypothesized to be ≤ 3.5 years (Beamish 1980b). However, field studies are lacking, individual lamprey may differ in ocean residence times, and variability in adult body size may reflect different feeding behaviors (Beamish 1980b; Weitkamp et al. 2015) or reflect growth differences from feeding on different host species. Field studies on anadromous sea lamprey Petromyzon marinus have indicated that their juvenile feeding stage is relatively short at 10–20 months (Silva et al. 2013, 2016). The body size of adult Pacific lamprey migrating into freshwater to spawn differs significantly across locations (Fig. 2). However, different collection protocols and collection distances from the ocean may influence this pattern. Age estimates using anatomical structures or isotopic compositions from Pacific lamprey may yield insights on the duration of their marine residence (if methods can be developed to effectively apply these techniques; reviewed in Hansen et al. 2016 for anadromous sea lamprey). Additional research is needed to document ocean-residence time for Pacific lamprey and factors that control it.

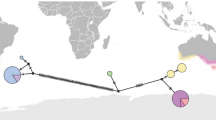
Mean total body lengths (values shown) of adult Pacific lamprey migrating into freshwater (or that have already migrated into freshwater) to spawn. Data are shown across different locations (left-to-right = south to north) from different studies in North America: Klamath estuary, California, USA (Parker 2018); Coquille River, Oregon, USA (Brumo 2006); Smith river, Oregon, USA (Starcevich et al. 2014); North Fork Umpqua River, Oregon, USA (Lampman 2011); Columbia River estuary, Oregon and Washington, USA (Weitkamp et al. 2015); Willamette River, Oregon, USA (Porter et al. 2017); Columbia River, Oregon and Washington, USA (Keefer et al. 2009); Puget Sound, Washington, USA (“PS watersheds”; Hayes et al. 2013); and British Columbia, Canada (“BC watersheds”; Beamish 1980b). Different symbols within the same location indicate different years. With the exception of the Coquille and North Fork Umpqua rivers, all 95% confidence intervals (CI) were within the area indicated by symbols. Weitkamp et al. (2015), Hayes et al. (2013), and Beamish (1980b) did not report standard deviation or 95% CI. Data for British Columbia watersheds is the mean of the means. The lack of significant overlap in 95% CI among locations indicates significant differences in body sizes of adult Pacific lamprey. See text for challenges in making comparisons across studies
Distribution and movements
Considering the vast hordes of Entosphenus tridentatus that at various sizes migrate up many coastal streams of the North Pacific to spawn, there are relatively few records from the sea… (Hubbs 1967).
The latitudinal range of Pacific lamprey in the ocean is the greatest of any lamprey in the world, with observations ranging from southwest of Baja California north to the Bering and Chukchi seas and westward into Russia and Japan (Renaud 2008; Orlov et al. 2009; Renaud 2011). High catches of Pacific lamprey in the Pacific Ocean have been reported along the slope area of the Bering Sea, with some occurrences in the Gulf of Alaska, from southeast Alaska to the eastern Aleutian Islands and westward off the Kamchatka peninsula (Orlov et al. 2008a; AFSC 2019). Pacific lamprey have also been caught in the ocean off the Washington and Oregon coasts by the commercial fishery for Pacific hake Merluccius productus and stock assessment surveys for Pacific hake and groundfishes (Fleischer et al. 2005; Keller et al. 2008).
Movements of Pacific lamprey in the ocean are poorly understood. Although catch records exist for Pacific lamprey in the ocean (e.g., Beamish 1980b; Orlov et al. 2008a; Siwicke and Seitz 2017), where they originated from or the route taken to arrive at their location, or whether observations reflect typical seasonal movements is not known. It is also unclear whether dispersal occurs primarily through independent swimming or when attached to hosts. Statistically-significant associations have been reported between the abundance of adult Pacific lamprey returning to the Columbia River Basin and the relative abundance of their hosts, including Pacific herring Clupea pallasii, Chinook salmon Oncorhynchus tshawytscha, Pacific cod Gadus macrocephalus, walleye pollock Gadus chalcogrammus, and Pacific hake (Murauskas et al. 2013). Pacific lamprey may migrate with their host species in the ocean north of the Columbia River mouth to feed on fish stocks off Vancouver Island, British Columbia (Murauskas et al. 2013). Further evidence for this hypothesis comes from genetic stock structuring of Pacific lamprey, which suggests dispersal in the ocean may be related to host movements (Spice et al. 2012).
Pacific lamprey observed in the Bering Sea off Alaska and Russia may originate from rivers in Canada and the U.S., since there appears to be no large lamprey populations occupying river basins emptying into the Bering Sea (Murauskas et al. 2013). Although evidence of Pacific lamprey originating in North America and rearing in the Bering Sea is lacking, evidence of a reverse relationship (rearing in the Bering Sea and migrating to freshwater in North America) exists. An adult Pacific lamprey tagged with a PIT (passive-integrated transponder) tag in the Bering Sea was subsequently detected at Bonneville Dam and the Deschutes River in Oregon (USA), a distance of thousands of kilometers (Murauskas et al. 2019). Further studies are needed to elucidate movements of Pacific lamprey at multiple spatial and temporal scales in the ocean and to clarify connections between marine feeding grounds and migrations to particular rivers for spawning.
Although their depth distribution is highly variable, Pacific lamprey are generally distributed at mid-water depths. In the open ocean, Pacific lamprey have been caught from the surface down to 1485 m in depth (Orlov et al. 2008a), but they are most often caught between the surface and 500 m (Orlov et al. 2008a; Wade and Beamish 2016). In surveys in the Strait of Georgia, Pacific lamprey were commonly caught between 31 and 100 m, followed by 101–500 m (Wade and Beamish 2016). Similarly, the largest catches made in NOAA Fisheries surveys occurred at open-water survey depths of 100–300 m, where bottom depths were 100–800 m (AFSC 2019).
Regional differences in depth distributions of Pacific lamprey may also occur. For example, very few lamprey have been caught by the extensive bottom trawls of the West Coast groundfish survey (Bradburn et al. 2011), suggesting a pelagic rather than demersal existence. In contrast, Pacific lamprey are frequently caught by bottom trawls on the Bering Sea slope survey (AFSC 2019), suggesting a demersal predisposition in that area. The reasons for these patterns are unclear, but may be related to the depth distributions of hosts of Pacific lamprey. The greater catches of Pacific lamprey by the Bering Sea slope survey are probably not caused by differences in trawl gear because the codend liner used in these surveys (31.8 mm stretched mesh size) is bigger than that used by the West Coast groundfish survey (19.1 mm stretched mesh), and too fine to allow all but the smallest lamprey to escape. This suggests that the regional differences in catch and therefore inferred depth distributions of Pacific lamprey noted above are not caused by gear type.
Pacific lamprey caught at 100–250 m are “slightly larger than the size of the average downstream migrant”, and Pacific hake and walleye pollock caught at these depths occasionally display lamprey wounds (Beamish 1980b). The largest catches made through trawl surveys occurred at open-water survey depths of 100–300 m (bottom depths were 100–800 m), and the largest catches of Pacific lamprey were from stock-assessment surveys for Pacific hake (Fleischer et al. 2005). However, a very large catch of adult Pacific lamprey was recently made in association with a school of walleye pollock at a depth of 45 m (Wade and Beamish 2016). This information suggests that depth distribution of Pacific lamprey is associated with that of their common hosts, including walleye pollock and Pacific hake. Pacific lamprey may undergo daily vertical migrations in the water column (shallower at night and deeper by day), which has been linked to vertical migrations (DVM) in walleye pollock (Orlov et al. 2008a), and may also be linked to the DVM of Pacific hake (Ressler et al. 2008). It is not known whether lamprey undergo these vertical migrations while attached to their hosts, while swimming independently, or both.
Genetic population structure
Use of genetic technology on Pacific lamprey in the Eastern Pacific Ocean has provided considerable insights into genetic diversity that differs by each of two marker types. Neutral genetic markers (i.e., mitochondrial DNA and microsatellite markers) are thought to be unaffected by natural selection (Goodman et al. 2008; Spice et al. 2012), whereas adaptive genetic markers (i.e., single nucleotide polymorphisms) are subjected to natural selection (Hess et al. 2013, 2015). Studies using neutral genetic markers suggest very low genetic population structure, with high gene flow across large geographical areas (Goodman et al. 2008; Spice et al. 2012). Based on neutral variation, Pacific lamprey in North America can be grouped into three large geographic regions: (1) Northern British Columbia; (2) Vancouver Island, British Columbia/Puget Sound, Washington; and (3) West Coast of the U.S. and the Columbia River (Hess et al. 2013). Studies using adaptive markers suggest a greater level of genetic structure than neutral markers, and adaptive markers can differentiate Pacific lamprey from the lower versus the interior Columbia River (Hess et al. 2013). This adaptive variation in adult Pacific lamprey appears to be associated with migration timing, body size, and upstream migration distance. Adaptive genetic markers were found to have the strongest association with body size (Hess et al. 2014). Although the genetic basis of adult body size in Pacific lamprey appears to be important for the evolution of this species, it is unclear what selective mechanism(s) drive variation (Hess et al. 2014). Many adult migration traits can be correlated, which makes it difficult to determine the true selective mechanisms at work.
A better understanding of the selective mechanisms acting on Pacific lamprey may be possible if more information can be gathered on juveniles, as they have the greatest potential for divergence in body size. For example, lamprey may diverge in body size if they distribute in different areas of the ocean, feed on different hosts, spend different periods of time in the ocean, and grow at different rates. The adaptive genetic markers that associate with adult body size may not be underlying a physiological mechanism of body size, but rather could predetermine individual lamprey fate to exhibit behavioral difference(s) in the ocean (Hess et al. 2014).
Feeding behavior and hosts
Pacific lamprey are parasitic during their juvenile life stage in the ocean, feeding on the blood, body fluids (Potter and Hilliard 1987; Renaud et al. 2009a), and flesh (Orlov et al. 2007) of their hosts. Pacific lamprey attach to the anterior and ventral body areas of hosts with their oral disc, which is covered with sharp teeth (Hart 1973; Cochran 1986; Orlov et al. 2009). Thirty-two different species of fishes and mammals have been documented as hosts for Pacific lamprey in the Pacific Ocean (Table 1; Fig. 3); yet many data gaps exist regarding when, where, why, and how Pacific lamprey parasitize them, and the degree of switching hosts (Beamish 1980b; Renaud et al. 2009a; Clemens et al. 2010; Murauskas et al. 2013, 2016; Orlov 2016; Wade and Beamish 2016). The large number of diverse host species upon which Pacific lamprey feed suggests this species is opportunistic. Another widely-distributed, large anadromous lamprey, the sea lamprey, also parasitizes diverse species (Silva et al. 2014). One key parameter necessary to understand lamprey feeding is the duration of attachment to individual hosts (hours, days, or months) and whether serving as host is fatal or impairs the ability of the host to grow.

Examples of wounds (shown by black arrows) by Pacific lamprey on hosts in the ocean. Pictures from NOAA Fisheries groundfish surveys; courtesy of NOAA Fisheries
Pacific lamprey do not always kill their hosts, as evidenced by the capture of hosts with healed scars (e.g., Siwicke and Seitz 2015). The extent of sub-lethal and lethal impacts by Pacific lamprey to their hosts is unknown and may depend on many factors specific to the biology and size of each host, duration spent attached to individual hosts, and host physiology and health (e.g., see work on landlocked sea lamprey by Swink 2003; Patrick et al. 2009; Sepulveda et al. 2012). Host switching by Pacific lamprey likely occurs as they grow, based on the duration of estimated time at sea and the growth that occurs during the juvenile life stage. Wounds on Pacific herring are smaller than those on walleye pollock, which are smaller than those on Pacific cod (Orlov et al. 2009). This suggests that larger Pacific lamprey, like anadromous sea lamprey (Silva et al. 2014), may selectively parasitize larger individuals or host survival may be size dependent. Wounding rates by Pacific lamprey on Pacific cod in the Bering Sea were most prevalent on relatively large Pacific cod (Siwicke and Seitz 2015). Data on these wounds were used to infer that some mortality of Pacific cod was caused by Pacific lamprey (Siwicke and Seitz 2015).
Host species of Pacific lamprey have been identified by the wounds and scars on them that match the shape and size of the oral disc of Pacific lamprey (Siwicke and Seitz 2015). In addition, hosts are sometimes identified by finding Pacific lamprey in samples with fishes bearing lamprey wounds. However lamprey wounds and scars may not always clearly indicate which species of lamprey caused the wound, and Pacific lamprey typically detach from their host upon capture with fishing nets (Nikiforov et al. 2018; L. Weitkamp, pers. obs.). Wounds and scars made by Pacific lamprey can be difficult to distinguish from those made by Arctic lamprey Lethenteron camtschaticum (Siwicke and Seitz 2015; Orlov and Baitaliuk 2016). In these situations genetic sampling (scraping) the tissues of hosts with fresh wounds for DNA may be an effective method to identify the particular species of lamprey that caused those wounds (Nikiforov et al. 2018). Another method to identify host species is to genetically sample lamprey gut contents, although examination of these revealed blood and crushed tissue that could not be traced back to particular host species (Orlov et al. 2007). Environmental DNA (eDNA) has been used to identify host species from the gut contents of Arctic lamprey (Shink et al. 2019), and it has been proposed that eDNA sampling of the gut contents of Pacific lamprey should be conducted (Nikiforov et al. 2018). If, however, one is more interested in the general trophic position of feeding rather than identifying particular prey species, then assaying fatty acid profiles or stable isotopes from lamprey tissue can be used (e.g., for sea lamprey, see Harvey et al. 2008; Lança et al. 2013; Happel et al. 2017).
Despite the diversity of known hosts, two species of gadids, walleye pollock and Pacific hake, appear to be common hosts for Pacific lamprey. These species exhibit a relatively high frequency of lamprey wounds, and relatively large numbers of lamprey are caught in fisheries or surveys targeting them (Fleischer et al. 2005; Siwicke and Seitz 2015; L. Weitkamp unpubl. data). Walleye pollock have a more northern distribution with the highest abundance in Alaskan waters (Hart 1973; Mecklenburg et al. 2002), and this species appears to be the most common host for Pacific lamprey in the Strait of Georgia and farther north (Orlov et al. 2008b; Wade and Beamish 2016; Orlov 2016). By contrast, Pacific hake are most abundant in the California Current off the coasts of Washington, Oregon, and California (Ressler et al. 2008; Mecklenburg et al. 2002), and this species is likely a common host in this region (Orlov et al. 2008b; Wade and Beamish 2016).
Abundance, recruitment, and growth rates
The abundance of Pacific lamprey is likely associated with the abundance of their hosts. Time-series data for Pacific lamprey suggests decadal cycles of abundance, which has been attributed to their approximate life span (Murauskas et al. 2013). Marine survival of Pacific lamprey may be limited by the availability of hosts, which is influenced by environmental conditions and therefore may be important in determining the abundance of adult Pacific lamprey migrating into freshwater (Murauskas et al. 2013; Wade and Beamish 2016). However, the survival rate of Pacific lamprey from larval to adult life stages is not known, and no relationship between adult and larval counts has been established (Clemens et al. 2017a).
A potential model for how marine conditions might affect Pacific lamprey recruitment to adult spawners stems from studies of Pacific salmon. Research on Pacific salmon has developed a theory of population recruitment pertaining to the Pacific Decadal Oscillation (PDO). Cold phases of the PDO have been associated with increasing bottom-up processes (i.e., upwelling and subsequent nutrient infusion into surface sea water), subsequent proliferation of zooplankton with high lipid content, and the increase of zooplanktivorous fishes, including juvenile Pacific salmon. Warm phases of the PDO, by contrast, lead to top-down processes, which include increased numbers of piscivores that migrate northward in the California Current and feed on small fishes at the same time that upwelling and nutrient infusions into surface seawaters are reduced, along with the zooplankton and fishes that feed on them. Hence cool PDOs are linked to greater marine survival and returns of Pacific salmon, whereas warm PDOs are associated with poor survival and returns (Mantua et al. 1997; Peterson and Schwing 2003; Peterson et al. 2014). Pacific lamprey parasitize fishes that would benefit from the cool-phase PDO, and the abundance of these fishes is associated with numbers of adult Pacific lamprey migrating into the Columbia River (Murauskas et al. 2013). As a result, we hypothesize that Pacific lamprey, like Pacific salmon, may benefit from cool PDOs in the Pacific Northwest. That is, we hypothesize that marine survival of Pacific lamprey is influenced by PDO regimes through its impact on host abundances, which determines the abundance of adult lamprey migrating into freshwater (Fig. 4). We acknowledge that this is a simplistic hypothesis that may not adequately fit the complex biology of Pacific lamprey that have many possible hosts (Table 1).

Conceptual model of how oceanographic regime changes may affect recruitment of juvenile Pacific lamprey in the ocean. Arrows can be read as “leads to”, ‘−’ decreased, and ‘+’ increased. Each of the boxes directly influences those factors indicated in the boxes to the right. For example, the top line of the figure indicates that a warm phase of the Pacific Decadal Oscillation (PDO) has been linked to decreased upwelling and nutrient infusion into surface seawater, which leads to less production of zooplankton and reduced recruitment of zooplanktivorous fishes for juvenile Pacific lamprey to parasitize, leading to reduced numbers of them. In addition, the warm phase of the PDO is associated with increased numbers of piscivores moving northward from the California Current into the waters of the Pacific Northwest, and these may depredate juvenile Pacific lamprey. This is referred to as “top-down” process, because of the influence of predators from atop the food web on production of fishes (in this case lamprey) during the warm phase of the PDO. The cold phase of the PDO yields opposite reactions in all cases, and is referred to as a “bottom-up” process because increased production from the bottom of the food web leads to increased production of fishes
Pacific lamprey attain their maximum body size during their juvenile life stage in the ocean (Beamish 1980b; Weitkamp et al. 2015). Although empirical data on growth rates for Pacific lamprey as juveniles are lacking, some information exists. The mean total length of Pacific lamprey caught in the Columbia River estuary immediately before and after their marine phase was 133 mm and 596 mm, respectively (Weitkamp et al. 2015). If these lamprey spent 3 to 4 years in the ocean (Beamish 1980b), then growth rates are estimated to be 0.42 mm/d for 3 years and 0.32 mm/d for 4 years. Mean weight data from the same individuals (3.6 g juveniles, 460 g adults) indicate growth rates of 0.42 and 0.31 g/day or 0.19% and 0.14% body weight/day if they spend three and 4 years in the ocean, respectively. This is somewhat lower than the 0.65–0.79 g/day estimated by Beamish (1980b) for anadromous sea lamprey in the Western Atlantic Ocean and landlocked sea lamprey the Great Lakes (0.89 g/day, Applegate 1950; 0.5–2.0%/day, Farmer 1980), both of which have shorter parasitic phases at 1–2.5 years (Beamish 1980b). Recent information suggests that anadromous sea lamprey feed as juveniles for only about one to 1.5 years (Silva et al. 2013, 2016). Improvements in the understanding of how long Pacific lamprey rear in the ocean, seasonal patterns of growth, empirical information on feeding behavior and hosts, temperatures experienced, and bioenergetics modelling (e.g., Madenjian et al. 2008; Jorgensen and Kitchell 2005) will be necessary to refine these growth estimates.
Predators
Pacific lamprey exhibit two characteristics that make them a high-quality prey: (1) they have high lipids and high energy content; and (2) they do not have the swimming capabilities necessary to avoid many predators (Close et al. 2002). Caloric values of Pacific lamprey range from 5.92 to 6.34 kcal/g wet mass (Whyte et al. 1993), which is 3.3–6.2 times higher than five species of Pacific salmon (Oncorhynchus spp.; 1.03–1.79 kcal/g; O’Neill et al. 2014). Pacific lamprey are known to serve as prey for at least 17 different species of fishes, birds, and mammals in estuarine and ocean waters (Table 2), with more likely to be identified. The lack of a bony skeleton may enable more rapid digestion of Pacific lamprey, leading to under-estimates of presence in the guts of predators (Cochran 2009).
Some studies reported the frequency with which Pacific lamprey appeared in gut contents in relation to other prey. For example, California sea lions Zalophus californianus, Steller sea lions Eumetopias jubatus, and Pacific harbor seals Phoca vitulina richardii consumed Pacific lamprey more than any other fish species in the Rogue River (Roff and Mate 1984). Pacific lamprey are likely a trophic buffer to some species of migrating salmon, as pinnipeds may preferentially consume Pacific lamprey (Close et al. 2002). In the Columbia River estuary, large colonies of gulls Larus spp., Caspian terns Sterna caspia, and double-crested cormorants Phalacrocorax auritus reside on dredge spoils and consume outmigrating larval and juvenile lampreys, which comprised < 1.9% of the diet samples of these birds (e.g., Collis et al. 2002).
Indigenous peoples harvest Pacific lamprey for food, medicine, and ceremonial purposes in estuaries and rivers (Close et al. 2002; Petersen-Lewis 2009). When other high lipid foods (e.g., Pacific salmon Oncorhynchus spp.) are seasonally unavailable, the capture of Pacific lamprey provides high caloric food (Close et al. 1995) for indigenous people coinciding with the coldest season of the year (Parker 2018). While not targeted directly by ocean fisheries, Pacific lamprey are caught as bycatch by marine fisheries for other species. The geographically-wide distribution of Pacific lamprey in the ocean indicates they are potentially vulnerable to numerous fisheries. However, preliminary data suggest that bycatch is greatest in fisheries targeting walleye pollock (Orlov and Pelenev 2009; Orlov 2016) and Pacific hake (Beamish 1980b). While Pacific lamprey bycatch has not been quantified nor has post-release survival been evaluated, persistent and high bycatch rates could be a conservation concern.
Life-history diversity
It is already evident that this species, [Pacific lamprey] like salmon and some other anadromous fishes, comprises a multitude of races, the members of which differ sharply in body size… (Hubbs 1967).
Life-history diversity in adult Pacific lamprey that have immigrated into freshwater for spawning migrations has been described in at least three dichotomies (which may not be mutually exclusive), including:
- 1.
Stream-maturing and ocean-maturing Some Pacific lamprey enter freshwater with immature gonads (stream-maturing genotype) and overwinter in freshwater before spawning, whereas others enter with more mature gonads (ocean-maturing genotype) and likely spawn within several weeks (Clemens et al. 2013; Porter et al. 2017; Parker et al. 2019). Unlike similar life histories characterized in a different anadromous species (e.g., premature and mature steelhead; Hess et al. 2016), these two Pacific lamprey life histories display overlapping freshwater-entry timing (Parker et al. 2019).
- 2.
Day eels and night eels Native American tribes often refer to Pacific lamprey as “eels” (Miller 2012). Day eels are described as small in body size (short body length) and light in coloration, as opposed to night eels which are large and dark in coloration (Downey et al. 1996; Close et al. 2004; Sheoships 2014). It has been hypothesized that day eels might have been in freshwater for an extended period of time (possibly equivalent to the stream-maturing, life-history strategy), whereas night eels may be recent migrants to freshwater ecosystems (Close et al. 2004).
- 3.
Normal and dwarf Observations on normal and dwarf life-history strategies of Pacific lamprey have been made in the Coquille River, Oregon (Kostow 2002) and Puget Sound, Washington (Hayes et al. 2013). The dwarf life-history is smaller in body size and has been observed to spawn at a different time within the same season than the normal life-history strategy (Kostow 2002). Genetic markers that are associated with the dwarf life history of Pacific lamprey have been identified (see Hess et al. 2013). What we refer to as dwarf Pacific lamprey here is presumably still anadromous (they are not blocked from the ocean). This is different from the dwarf life-history of European river lamprey Lampetra fluviatilis in the UK, which are freshwater resident parasites (Maitland 2003).
More research and careful comparisons are needed to understand whether these paired terms are describing the same genotypic and phenotypic diversity among Pacific lamprey.
Demographic parameters (e.g., feeding, growth, mortality, etc.) have been implicated in life-history diversity of fishes (e.g., McMillan et al. 2012; Tattam et al. 2013; Sloat and Reeves 2014); thus, factors in the ocean (where Pacific lamprey are rearing to maximum body sizes) may select for life-history diversity. Life-history diversity of Pacific lamprey may be most evident in body size and maturation timing, which may be associated with the duration of ocean residency or the type of host (Beamish 1980b; Weitkamp et al. 2015). Adult body size of Pacific lamprey is evolutionarily adaptive (Hess et al. 2014), and is directly related to three key aspects of lamprey behavior and fitness. First, adult body size is directly proportional to the ability of Pacific lamprey to ascend and pass natural and artificial obstacles to upstream migration, such as rapids, waterfalls, and hydroelectric dams (Keefer et al. 2009, 2010). Second, adult body size is directly proportional to the maximum distance upstream that Pacific lamprey migrate (Clemens et al. 2010; Hess et al. 2014). Third, like most fishes, (Kamler 2005), adult body size is directly proportional to the absolute fecundity of female Pacific lamprey (Clemens et al. 2010, 2013). Therefore, environmental and genetic influences on body size may influence life-history diversity in Pacific lamprey. Considerations of adult body size of lamprey are interesting in the context of regional patterns in body size (Fig. 2). However, attempts to identify life history by one metric alone (such as with body size) may be insufficient to identify life-history diversity (Clemens et al. 2013) for at least three reasons. First, the body lengths of Pacific lamprey can decrease 18–30% after entering freshwater (due to cessation of feeding and lack of skeletal components in their bodies; Beamish 1980b; Clemens et al. 2009, 2010). Second, differences in the distance upstream where lamprey were sampled exist across studies. For example, sampling in the estuary may yield recent migrants at their maximum body sizes in comparison with sites further upstream. Third, diversity in maturation timing of Pacific lamprey (stream maturing and ocean maturing life histories) has only been distinguished through a number of characteristics, and not by a single metric alone (Clemens et al. 2013; Parker et al. 2019).
Potential marine-related limiting factors and threats
The widespread distribution of Pacific lamprey, its parasitic life history, use of diverse hosts, and multiple predators suggests that this species is integrated into diverse marine ecosystems in the North Pacific Ocean. However, significant data gaps exist in understanding the limiting factorsFootnote 1 and threatsFootnote 2 to Pacific lamprey in estuaries in the marine environment.
Oceanographic work on various marine and anadromous species (e.g., Keller et al. 2010; Wainwright and Weitkamp 2013; Peterson et al. 2014; Cheung et al. 2015; Beamish 2017) and general knowledge of the biology of Pacific lamprey (this review) enables speculation about potential marine-related limiting factors and threats to them. Three potential marine-related limiting factors are: (1) predation (Table 2), including bycatch; (2) availability of hosts (Table 1); and (3) contaminant loads of hosts––which Pacific lamprey may bioaccumulate as a parasite (e.g., Drevnick et al. 2006). Predation and bycatch can be categorized as top-down limiting factors resulting from top predators, including humans. The last two, availability of hosts and contaminant loads of hosts, can be categorized as bottom-up limiting factors. Pacific lamprey is not exploited commercially or recreationally, which may explain why, historically, this species has not been consistently monitored. Bycatch rates of Pacific lamprey in the ocean and sampling to address questions about the effects of limiting factors and threats on their status is lacking. Bycatch is discussed further in the “Predators” section. Marine-related threats to Pacific lamprey may include: (1) pollution from humans; (2) changes in oceanographic regimes that are unfavorable to recruitment of lamprey (Fig. 4); and (3) climate change.
Pollution occurs from two general categories: (1) human-produced, artificial substances (xenobiotics) that can be ingested by organisms and then bioaccumulate in predators like lampreys (Gray 2002; Drevnick et al. 2006; Boerger et al. 2010; Zenker et al. 2014); and (2) releases of excessive nutrients through human processes (eutrophication) into estuaries and the ocean. Excessive nutrients can induce zones of hypoxia and affect ecosystem functioning, which can displace, change behavior, and cause die-offs (i.e., “dead zones”; Diaz and Rosenberg 2008; Keller et al. 2010; Somero et al. 2016). With a few exceptions, eutrophication is not a problem in most estuaries in the West Coast of North America. Xenobiotics and eutrophication have the potential to negatively impact Pacific lamprey directly by affecting water quality and causing acute stress and death or indirectly, through ingestion of contaminated host tissue. The potential limiting factors and threats to lamprey that we have identified in the marine environment are broad characterizations across the vast North Pacific Ocean, and these may not apply in particular locations.
Estuarine and ocean conditions may influence the abundance of Pacific lamprey in many ways, including via oceanographic regimes (Fig. 4) and interactions of climate change with these regimes. For example, climate change has been implicated in the following recent physical, chemical, and biological changes in the marine environment. These changes include: increased sea surface temperatures (ISAB 2007; Mote et al. 2014; Sharma et al. 2016), hypoxia of bottom waters (Williams et al. 2015), ocean acidification (Haigh et al. 2015; Mathis et al. 2015), increased sea levels and estuarine/marsh inundation (ISAB 2007; Mote et al. 2014), reduced upwelling (Rykaczewski et al. 2015; Sharma et al. 2016), change in distribution of host species (discussed below), decreased host populations (ISAB 2007; Wade et al. 2013; Williams et al. 2015), and decreased productivity of food webs (ISAB 2007; Crozier 2016; Sharma et al. 2016). Climate change has and will continue to cause environmental and ecological changes that will be increasingly evident by the mid and late 21st century (IPCC 2014), and include exacerbation of existing limiting factors to Pacific lamprey in both indirect and direct ways. Environmental alterations will affect organisms inhabiting coastal waters directly (e.g., physiological stress due to increased temperatures or low dissolved oxygen; Somero et al. 2016), and indirectly through changes to ecosystems (e.g., novel predator–prey interactions; Cheung et al. 2015; Rehage and Blanchard 2016). How this multitude of physical and ecosystem changes will affect Pacific lamprey is difficult to predict. Increased ocean temperatures are expected to alter fish and invertebrate distributions to deeper depths and toward the poles (Pinsky et al. 2013; Cheung et al. 2015), and these changes were observed in the recent marine heat wave (Leising et al. 2015; Peterson et al. 2017; Morgan et al. 2019). The poleward movement of animals includes Pacific hake (Cheung et al. 2015; Auth et al. 2017), known hosts of Pacific lamprey (Table 1). Pacific lamprey distributions may also change as they either follow their hosts northwards or switch to new hosts that have not been encountered with frequency in the past. The effects of climate change will have complex, cumulative, and potentially synergistic effects that are difficult to predict, but at a minimum may affect host availability, population recruitment, and growth rates of Pacific lamprey.
Recommendations and conclusions
We reviewed the literature on the marine biology of juvenile Pacific lamprey and updated and expanded on information provided nearly 40 years ago by Beamish (1980b). We synthesized information on the freshwater/marine transitions of Pacific lamprey; their distribution and marine movements; genetic population structure; feeding behavior and hosts; abundance, recruitment, and growth rates; predators; life-history diversity; and marine-related limiting factors and threats. The juvenile life stage of Pacific lamprey in the ocean is important for the attainment of maximum body size, which directly correlates with three key aspects to the behavior and fitness of Pacific lamprey: (1) passage (Keefer et al. 2009, 2010); (2) migration distance in freshwater (Hess et al. 2014); and fecundity (Clemens et al. 2013). The abundance of hosts for Pacific lamprey in the ocean correlates with abundance of adult migrations into freshwater (Murauskas et al. 2013).
New, key insights into the marine biology of Pacific lamprey include apparent latitudinal differences in transformation and ocean entrance timing; updated host species and predator lists; a hypothesis of population recruitment related to the PDO (Fig. 4) and climate change; a synthesis of descriptions of life history diversity; a meta-analysis on regional differences in body size (Fig. 2); and the first identification of potential, marine-related limiting factors and threats. Our review has identified 26 key research needs pertaining to the marine biology and marine-related limiting factors and threats to Pacific lamprey (Table 3). Based on patterns that have emerged from our review of the literature, we have identified and advanced eight hypotheses (highlighted in Table 4). Many data gaps remain regarding the biology, limiting factors, and threats to Pacific lamprey in the marine environment (Clemens et al. 2010; this review). Therefore research may be the most important goal towards furthering science and management of Pacific lamprey. The widespread distribution of Pacific lamprey, its parasitic life history, and diverse hosts suggest that this species is integrated into diverse marine ecosystems.
Notes
We define limiting factors to this species as the physical, chemical, or biological conditions of the environment that constrain the behavior, abundance, productivity, diversity, or distribution. Therefore, direct challenges to Pacific lamprey in estuaries and the Pacific Ocean include physiological stress, delayed mortality, misalignment of environmental cues with species adaptations, and instantaneous mortality via acute trauma or predation.
We define threats as human-induced or natural processes or actions that may create or exacerbate limiting factors.
References
Abakumov VA (1964) Concerning the sea period of life of the Pacific lamprey Enthosphenus tridentatus (Richardson). Tr VNIRO 49:253–256
AFSC (Alaska Fisheries Science Center) (2019). https://www.fisheries.noaa.gov/resource/map/alaska-groundfish-fishery-observer-data-map. Accessed Apr 2019
Antonelis GA Jr, Fiscus CH (1980) The pinnipeds of the California sea current. CalCOFI Rep 21:68–73
Auth TD, Daly EA, Brodeur RD, Fisher JL (2017) Phenological and distributional shifts in ichthyoplankton associated with recent warming in the northeast Pacific Ocean. Glob Change Biol 24:259–272
Beamish FWH (1980a) Biology of the North American anadromous sea lamprey, Petromyzon marinus. Can J Fish Aquat Sci 37:1924–1943
Beamish RJ (1980b) Adult biology of the river lamprey (Lampetra ayresi) and the Pacific lamprey (Lampetra tridentata) from the Pacific coast of Canada. Can J Fish Aquat Sci 37:1906–1923
Beamish RJ (2017) What the past tells us about the future of Pacific salmon research. Fish Fish 18:1161–1175
Beamish RJ, Levings CD (1991) Abundance and freshwater migrations of the anadromous parasitic lamprey, Lampetra tridentata, in a tributary of the Fraser River, British Columbia. Can J Fish Aquat Sci 48:1250–1263
Boerger CM, Lattin GL, Moore SL, Moore CJ (2010) Plastic ingestion by planktivorous fishes in the North Pacific central gyre. Mar Pollut Bull 60:2275–2278
Bradburn MJ, Keller AA, Horness BH (2011) The 2003–2008 US West Coast bottom trawl surveys of groundfish resources off Washington, Oregon, and California: estimates of distribution, abundance, length, and age composition. US Department of Commerce, NOAA Technical Memorandum NMFS-NWFSC-114
Brumo A (2006) Spawning, larval recruitment, and early life survival of Pacific lampreys in the South Fork Coquille River, Oregon. Master’s thesis, Oregon State University
Brumo AF, Grandmontagne L, Namitz SN, Markle DF (2009) Approaches for monitoring Pacific lamprey spawning populations in a coastal Oregon stream. In: Brown LR, Chase SD, Mesa MG, Beamish RJ, Moyle PB (eds) Biology, management, and conservation of lampreys in North America. Symposium 72. American Fisheries Society, Bethesda, pp 203–222
Chase SD (2001) Contributions to the life history of adult Pacific lamprey (Lampetra tridentata) in the Santa Clara River of Southern California. South Calif Acad Sci Bull 100:74–85
Cheung WWL, Brodeur RD, Okey TA, Pauly D (2015) Projecting future changes in distribution of pelagic fish species of Northeast Pacific shelf seas. Prog Oceanogr 130:19–31
Clemens WA, Wilby GV (1949) Fishes of the Pacific coast of Canada. Bull Fish Res Board Can 68:1–368
Clemens BJ, van de Wetering S, Kaufman J, Holt R, Schreck CB (2009) Do summertime temperatures trigger springtime maturation in adult Pacific lamprey, Entosphenus tridentatus? Ecol Freshw Fish 18:418–426
Clemens BJ, Binder TR, Docker MF, Moser ML, Sower SA (2010) Similarities, differences, and unknowns in biology and management of three parasitic lampreys of North America. J Fish 35:580–594
Clemens BJ, van de Wetering SJ, Sower SA, Schreck CB (2013) Maturation characteristics and life history strategies of the Pacific lamprey, Entosphenus tridentatus. Can J Zool 91:775–788
Clemens B, Schreck C, van de Wetering S, Sower S (2016) The potential roles of river environments in selecting for stream- and ocean-maturing Pacific lamprey, Entosphenus tridentatus (Gairdner, 1836). In: Orlov AM, Beamish RJ (eds) Jawless fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 299–322
Clemens BJ et al (2017a) Conservation challenges and research needs for Pacific lamprey in the Columbia River Basin. J Fish 42:268–280
Clemens BJ, Wyss L, McCoun R, Courter I, Schwabe L, Peery C, Schreck CB, Spice EK, Docker MF (2017b) Temporal genetic population structure and interannual variation in migration behavior of Pacific lamprey Entosphenus tridentatus. Hydrobiologia 794:223–240
Close DA, Fitzpatrick M, Li H, Parker B, Hatch D, James G (1995) Status report of the Pacific Lamprey (Lampetra tridentata) in the Columbia River basin annual report of the Oregon cooperative Fish and Wildlife research unit to the Bonneville power administration. Portland, Oregon
Close DA, Fitzpatrick MS, Li HW (2002) The ecological and cultural importance of a species at risk of extinction, Pacific lamprey. J Fish 27:19–25
Close DA, Jackson AD, Conner BP, Li HW (2004) Traditional ecological knowledge of Pacific lamprey (Entosphenus tridentatus) in northeastern Oregon and southeastern Washington from indigenous peoples of the Confederated Tribes of the Umatilla Indian Reservation. J Northwest Anthropol 38:141–162
Cochran PA (1986) Attachment sites of parasitic lampreys: comparisons among species. Environ Biol Fish 17:71–79
Cochran PA (2009) Predation on lampreys. In: Brown LR, Chase SD, Mesa MG, Beamish RJ, Moyle PB (eds) Biology, management, and conservation of lampreys in North America. Symposium 72. American Fisheries Society, Bethesda, pp 139–151
Collis K, Roby DD, Craig DP, Adamany S, Adkins J, Lyons DE (2002) Colony size and diet composition of piscivorous waterbirds on the Lower Columbia River: implications for losses of juvenile salmonids to avian predation. Trans Am Fish Soc 131:537–550
Collis K, Roby DD, Couch C, Dorsey G, Fischer K, Lyons DE, Myers AM, Nelson SK, Adkins JY, Evans AE (2006) Piscivorous waterbird research on the Columbia River Final 2004 Season Summary Report, 91 electronic pp
Condit R, Le Boeuf BJ (1984) Feeding habits and feeding grounds of the northern elephant seal. J Mammal 65:281–290
CRITFC (Columbia River Inter-Tribal Fish Commission) (2011) Tribal Pacific lamprey restoration plan for the Columbia River basin CRITFC, Portland, Oregon. https://www.critfc.org/wp-content/uploads/2012/12/lamprey_plan.pdf. Accessed Aug 2016
Crozier L (2016) Impacts of climate change on salmon of the Pacific Northwest: a review of the scientific literature published in 2015. Northwest Fisheries Science Center, National Marine Fisheries Service, 42 electronic pp
Dawson HA, Quintella BR, Almeida PR, Treble AJ, Jolley JC (2015) The ecology of larval and metamorphosing lampreys. In: Docker MF (ed) Lampreys: biology, conservation and control. Fish and Fisheries monograph series, vol 1. Springer, New York, pp 75–137
Diaz RJ, Rosenberg R (2008) Spreading dead zones and consequences for marine ecosystems. Science 321:926–929
Downey T, Rilatos D, Sondenaa A, Zybach B (1996) Skwakol: the decline of the Siletz lamprey EEL population during the 20th century Oregon State University Chapter. American Indians in Science and Engineering Society (AISES), Oregon State University, Corvallis, p 90
Drevnick PE, Horgan MJ, Oris JT, Kynard BE (2006) Ontogenetic dynamics of mercury accumulation in northwest Atlantic sea lamprey (Petromyzon marinus). Can J Fish Aquat Sci 63:1058–1066
Dyldin YV, Orlov AM (2016) Ichthyofauna of fresh and brackish waters of Sakhalin Island: an annotated list with taxonomic comments: 1. Petromyzontidae–Clupeidae families. J Ichthol 56:534–555
Ebert DA (1986) Biological aspects of sixgill shark, Hexanchus griseus. Copeia 1986:131–135
ESA (Endangered Species Act) (1973) US Code, vol 16, sections 1531-1544
Farmer GJ (1980) Biology and physiology of feeding in adult Lampreys. Can J Fish Aquat Sci 37:1751–1761
Fleischer GW, Cooke KD, Ressler PH, Thomas RE, de Blois SK, Hufnagle LC, Kronlund AR, Holmes JA, Wilson CD (2005) The 2003 integrated acoustic and trawl survey of Pacific hake, Merluccius productus. In: US and Canadian waters off the Pacific coast US Department of Commerce, NOAA Technical Memorandum NMFS-NWFSC-65
Flinn RD, Trites AW, Gregr EJ (2002) Diets of fin, sei, and sperm whales in British Columbia: an analysis of commercial whaling records, 1963–1967. Mar Mamm Sci 18:663–679
Goodman DH, Reid SB, Docker MF, Haas GR, Kinziger AP (2008) Mitochondrial DNA evidence for high levels of gene flow among populations of a widely distributed anadromous lamprey Entosphenus tridentatus (Petromyzontidae). J Fish Biol 72:400–417
Goodman DH, Reid SB, Som NA, Poytress WR (2015) The punctuated seaward migration of Pacific lamprey (Entosphenus tridentatus): environmental cues and implications for streamflow management. Can J Fish Aquat Sci 72:1–12
Gray JS (2002) Biomagnification in marine systems: the perspective of an ecologist. Mar Pollut Bull 45:46–52
Gunckel SL, Jones KK, Jacobs SE (2009) Spawning distribution and habitat use of adult Pacific and Western brook lampreys in Smith River, Oregon pp. 173–190. In: Brown LR, Chase SD, Mesa MG, Beamish RJ, Moyle PB (eds) Biology, management, and conservation of lampreys in North America. Symposium 72. American Fisheries Society, Bethesda, pp 73–190
Haigh R, Ianson D, Holt CA, Neate HE, Edwards AM (2015) Effects of ocean acidification on temperate coastal marine ecosystems and fisheries in the Northeast Pacific. PLoS ONE 10(2):e0117533. https://doi.org/10.1371/journalpone0117533
Hansen MJ, Madenjian CP, Slade JW, Steeves TB, Almeida PR, Quintella BR (2016) Population ecology of the sea lamprey (Petromyzon marinus) as an invasive species in the Laurentian Great Lakes and an imperiled species in Europe. Rev Fish Biol Fish 26:509–535
Happel A, Rinchard J, Czesny S (2017) Variability in sea lamprey fatty acid profiles indicates a range of host species utilization in Lake Michigan. J Gt Lakes Res 43:182–188
Hardisty MW (2006) Lampreys: life without jaws. Forest Text, Cardigan
Harris JE, Jolley J (2017) Estimation of occupancy, density, and abundance of larval lampreys in tributary river mouths upstream of dams on the Columbia River, Washington and Oregon. Can J Fish Aquat Sci 74:843–852
Hart JL (1973) Pacific fishes of Canada; Bulletin 180. Canadian Bulletin of Fisheries and Aquatic Sciences, Ottawa
Harvey JT (1989) Food habits, seasonal abundance, size, and sex of the blue shark, Prionace gluaca, in Monterey Bay, California. Calif Fish Game 75:33–44
Harvey CJ, Ebener MP, White CK (2008) Spatial and ontogenetic variability of sea lamprey diets in Lake Superior. J Great Lakes Res 34:434–449
Hayes MC, Hays R, Rubin SP, Chase DM, Hallock M, Cook-Tabor C, Luzier CW, Moser ML (2013) Distribution of Pacific lamprey Entosphenus tridentatus in watersheds of Puget Sound based on smolt monitoring data. Northwest Sci 87:95–105
Hess JE, Campbell NR, Close DA, Docker MF, Narum SR (2013) Population genomics of Pacific lamprey: adaptive variation in a highly dispersive species. Mol Ecol 22:2898–2916
Hess JE, Caudill CC, Keefer ML, McIlraith BJ, Moser ML, Narum SR (2014) Genes predict long distance migration and large body size in a migratory fish, Pacific lamprey. Evol Appl 7:1192–1208
Hess JE, Campbell NR, Docker MF, Baker C, Jackson A, Lampman R, McIlraith B, Moser ML, Statler DP, Young WP, Wildbill AJ, Narum SR (2015) Use of genotyping by sequencing data to develop a high-throughput and multifunctional SNP panel for conservation applications in Pacific lamprey. Mol Ecol Res 15:187–202
Hess JE, Zendt JS, Matala AR, Narum SR (2016) Genetic basis of adult migration timing in anadromous steelhead discovered through multivariate association testing. Proc R Soc B: Biol Sci 283:20153064. https://doi.org/10.1098/rspb20153064
Hokkaido Prefecture (2000) Hokkaido Red Data Book. http://rdb.hokkaido-ies.go.jp/page/detail1.asp?spc=000000000000000000620. Accessed Jan 2019 (in Japanese)
Hubbs CL (1967) Occurrence of the Pacific lamprey, Entosphenus tridentatus, off Baja California and in streams of southern California; with remarks on its nomenclature. Trans San Diego Soc Nat Hist 14:301–312
IPCC (International Panel on Climate Change) (2014) Climate Change 2014: synthesis report. Contribution of Working Groups I, II and III to the fifth assessment report of the intergovernmental panel on Climate Change [Core Writing Team, RK Pachauri and LA Meyer (eds)] IPCC, Geneva
ISAB (Independent Scientific Advisory Board) (2007) Climate change impacts on Columbia River Basin fish and wildlife, 146 electronic pp
Johnson NS, Buchinger TJ, Li W (2015) Reproductive ecology of lampreys. In: Docker MF (ed) Lampreys: biology, conservation and control. Fish and Fisheries monograph series, vol 1. Springer, New York, pp 265–303
Jolley JC, Silver GS, Whitesel TA (2012) Occupancy and detection of larval Pacific Lampreys and Lampetra spp. in a large river: the lower Willamette River. Trans Am Fish Soc 141:305–312
Jolley J, Uh CT, Silver GS, Whitesel TA (2015) Feeding and growth of larval Pacific lamprey reared in captivity. N Am J Aquac 77:449–459
Jorgensen JC, Kitchell JF (2005) Growth potential and host mortality of the parasitic phase of the sea lamprey (Petromyzon marinus) in Lake Superior. Can J Fish Aquat Sci 62:2343–2353
Kamler E (2005) Parent-egg-progeny relationships in teleost fishes: an energetics perspective. Rev Fish Biol Fish 15:399–421
Kan TT (1975) Systematics, variation, distribution, and biology of lampreys of the genus Lampetra in Oregon. Ph.D. dissertation, Oregon State University
Keefer ML, Moser ML, Boggs CY, Daigle WR, Peery CA (2009) Effects of body size and river environment on the upstream migration of adult Pacific lampreys (Lampetra tridentata). N Am J Fish Manag 29:1214–1224
Keefer ML, Daigle WR, Peery CA, Pennington HT, Lee SR, Moser ML (2010) Testing adult Pacific lamprey performance at structural challenges in fishways. N Am J Fish Manag 30:376–385
Keller AA, Horness BH, Fruh EL, Simon VH, Tuttle VJ, Bosley KL, Buchanan JC, Kamikawa DJ, Wallace JR (2008) The 2005 U.S. West Coast bottom trawl survey of groundfish resources off Washington, Oregon, and California: estimates of distribution, abundance, and length composition. U.S. Department of Commerce, NOAA Technical Memorandum, NMFS-NWFSC-93
Keller AA, Simon V, Chan F, Wakefield WW, Clarke ME, Barth JA, Kamikawa D, Fruh EL (2010) Demersal fish and invertebrate biomass in relation to an offshore hypoxic zone along the US West Coast. Fish Oceanogr 19:67–87
Kostow K (2002) Oregon lampreys: natural history status and problem analysis. Oregon Department of Fish and Wildlife, Portland
Lampman RT (2011) Passage, migration behavior, and autoecology of adult Pacific Lamprey at Winchester Dam and within the North Umpqua River Basin, Oregon, USA. Master’s thesis, Oregon State University
Lampman R, Moser M, Jackson A, Rose R, Gannam A, Barron J (2016) Developing techniques for artificial propagation and early rearing of Pacific lamprey Entosphenus tridentatus for species recovery and restoration. In: Orlov AM, Beamish RJ (eds) Jawless fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 160–195
Lança MJ, Machado M, Ferreira R, Alves-Pereira I, Quintella BR, Almeida PRD (2013) Feeding strategy assessment through fatty acid profiles in muscles of adult sea lampreys from the Western Iberian Coast. Sci Mar 77:281–291
Leising AW et al (2015) State of the California Current 2014–2015: impacts of the warm-water “blob”. CalCOFI Rep 56:31–68
Luzier CW, Schaller HA, Brostrom JK, Cook-Tabor C, Goodman DH, Nelle RD, Ostrand K, Streif B (2011) Pacific lamprey (Entosphenus tridentatus) assessment and template for conservation measures United States Fish and Wildlife service. Portland, Oregon, p 282
Madenjian CP, Chipman BC, Marsden JE (2008) New estimates of lethality of sea lamprey (Petromyzon marinus) attacks on lake trout (Salvelinus namaycush): implications for fisheries management. Can J Fish Aquat Sci 65:535–542
Maitland P (2003) Ecology of the river, brook and sea lamprey Lampetra fluviatilis, Lampetra planeri, Petromyzon marinus. Conserving Natura 2000 Rivers Ecology Series No. 5. English Nature, Peterborough
Maitland PS, Renaud CB, Quintella BR, Close DA, Docker MF (2015) Conservation of native lampreys. In: Docker MF (ed) Lampreys: biology, conservation and control. Fish and Fisheries monograph series, vol 1. Springer, New York, pp 375–428
Mantua NJ, Hare SR, Zhang Y, Wallace JM, Francis RC (1997) A Pacific interdecadal climate oscillation with impacts on salmon production. Bull Am Meteorol Soc 78:1069–1079
Manzon RG, Youson JH, Holmes JA (2015) Lamprey metamorphosis. In: Docker MF (ed) Lampreys: biology, conservation and control. Fish and Fisheries monograph series, vol 1. Springer, New York, pp 139–214
Mathis JT, Cooley SR, Lucey N, Colt S, Ekstrom J, Hurst T, Hauri C, Evans W, Cross JN, Feely RA (2015) Ocean acidification risk assessment for Alaska’s fishery sector. Prog Oceanogr 136:71–91
McGree M, Stone J, Whitesel TA (2008) Metamorphosis, growth, and survival of larval Pacific lamprey reared in captivity. Trans Am Fish Soc 137:1866–1878
McMillan JR, Dunham JB, Reeves GH, Mills JS, Jordan CE (2012) Individual conditions and stream temperature influence early maturation of rainbow steelhead trout, Oncorhynchus mykiss. Environ Biol Fish 93:343–355
Mecklenburg CW, Mecklenburg TA, Thorsteinson LK (2002) Fishes of Alaska. American Fisheries Society Press, Bethesda
Meeuwig MH, Bayer JM, Seelye JG (2005) Effects of temperature on survival and development of early life stage Pacific and Western brook lampreys. Trans Am Fish Soc 134:19–27
Miller J (2012) Lamprey “eels” in the greater Northwest: a survey of tribal sources, experiences, and sciences. J Northwest Anthropol 46:65–84
Morgan CA, Beckman BR, Weitkamp LA, Fresh KL (2019) Recent ecosystem disturbance in the Northern California Current. Fisheries. https://doi.org/10.1002/fsh.10273
Moser ML, Almeida PR, Kemp PS, Sorensen PW (2015a) Lamprey spawning migration. In: Docker MF (ed) Lampreys: biology, conservation and control. Fish and Fisheries monograph series, vol 1. Springer, New York, pp 215–263
Moser ML, Jackson AD, Lucas MC, Mueller RP (2015b) Behavior and potential threats to survival of migrating lamprey ammocoetes and macrophthalmia. Rev Fish Biol Fish 25:103–116
Mote P, Snover AK, Capalbo S, Eigenbrode SD, Glick P, Littell J, Raymondi R, Reeder S (2014) Chapter 21: Northwest. In: Melillo JM, Richmond TC, Yohe GW (eds) Climate change impacts in the United States: the third national climate assessment, US Global Research Program, pp 487–513
Moyle PB (2002) Inland fishes of California. University of California Press, Berkeley
Murauskas JG, Orlov AM, Siwicke KA (2013) Relationships between the abundance of Pacific lamprey in the Columbia River and their common hosts in the marine environment. Trans Am Fish Soc 142:143–155
Murauskas J, Schultz L, Orlov A (2016) Trends of Pacific lamprey populations across a broad geographic range in the North Pacific Ocean, 1939–2014. In: Orlov AM, Beamish RJ (eds) Jawless Fishes of the world. Cambridge Scholars Publishing, New Castle upon Tyne, pp 73–96
Murauskas JG, Orlov AM, Keller L, Maznikova OA, Glebov II (2019) Transoceanic migration of Pacific lamprey, Entosphenus tridentatus. J Ichthyol 59:280–282
Nemoto T (1955) White scars on whales (I) lamprey marks. Sci Rep Whales Res Inst (Tokyo) 10:69–77
Nikiforov AI, Gavrilov BA, Kruglova DK, Posokhova ES, Rabazanov NI, Orlov AM (2018) A research using DNA extracted from aquatic environment: state and prospects. Uspekhi Sovremennoi Biol 138:18–30
Nislow KH, Kynard BE (2009) The role of anadromous sea lamprey in nutrient and material transport between marine and freshwater environments. Am Fish Soc Symp 69:485–494
Novikov NP (1963) Events of attacks of Pacific lamprey Enthosphenus tridentatus (Gairdner) on halibuts and other fish of the Bering Sea. Vopr Ikhtiol 3:567–569
O’Neill SM, Ylitalo GM, West JE (2014) Energy content of Pacific salmon as prey of northern and southern resident killer whales. Endanger Species Res 25:265–281
Orlov AM (2016) Relationships between Pacific lamprey and their prey. In: Orlov A, Beamish RJ (eds) Jawless Fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 234–285
Orlov AM, Baitaliuk AA (2016) Distribution of Arctic and Pacific lampreys in the North Pacific. In: Orlov A, Beamish RJ (eds) Jawless Fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 32–56
Orlov AM, Pelenev DV (2009) Interspecies relationships between the Pacific lamprey Lampetra tridentata and walleye pollock Theragra chalcogramma. Vestnik SVNC DVO RAN 3:46–53
Orlov AM, Vinnikov AV, Pelenev DV (2007) Principles of studies of the sea period of anadromous parasitic lampreys (example of Pacific lamprey Lampetra tridentata (Gairdner, 1836) Petromyzontidae family). Vopr Rybolov 8:287–312
Orlov AM, Pelenev DV, Vinnikov AV (2008a) Pacific lamprey and commercial fish stocks in the Far Eastern waters of Russia. Rybn Khozyaistvo 2:60–65
Orlov AM, Savinyh VF, Pelenev DV (2008b) Features of the spatial distribution and size structure of the Pacific lamprey, Lampetra tridentata in the North Pacific. Russ J Mar Biol 34:276–287
Orlov AM, Beamish RJ, Vinnikov AV, Pelenev D (2009) Feeding and prey of Pacific lamprey on coastal waters of the western North Pacific. In: Haro A, Smith KL, Rulifson RA, Moffitt CM, Klauda RJ, Dadswell MJ, Cunjak RA, Cooper JE, Beal KL, Avery TS (eds) Challenges for diadromous fishes in a dynamic global environment. Symposium 69. American Fisheries Society, Bethesda, pp 875–877
Orr AJ, Banks AS, Mellman S, Huber HR, Delong RL (2004) Examination of the foraging habits of Pacific harbor seal (Phoca vitulina richardsi) to describe their use of the Umpqua River, Oregon, and their predation on salmonids. Fish Bull 102:108–117
Parker K (2018) Evidence for the genetic basis and inheritance of ocean and river-maturing ecotypes of Pacific lamprey (Entosphenus tridentatus) in the Klamath River, California. Master’s thesis, Humboldt State University
Parker K, Hess JE, Narum SR, Kinziger AP (2019) Evidence for the genetic basis and epistatic interactions underlying ocean‐ and river‐maturing ecotypes of Pacific Lamprey (Entosphenus tridentatus) returning to the Klamath River, California. Mol Ecol 28:3171–3185
Patrick HK, Sutton TM, Swink WD (2009) Lethality of sea lamprey parasitism on lake sturgeon. Trans Am Fish Soc 138:1065–1075
Petersen-Lewis RS (2009) Yurok and Karuk traditional ecological knowledge: insights into Pacific lamprey populations of the lower Klamath Basin. In: Brown LR, Chase SD, Mesa MG, Beamish RJ, Moyle PB (eds) Biology, management, and conservation of lampreys in North America. Symposium 72. American Fisheries Society, Bethesda, pp 1–40
Peterson WT, Schwing FB (2003) A new climate regime in Northeast Pacific ecosystems. Geophys Res Lett 30:1–4
Peterson WT, Fisher JL, Peterson JO, Morgan CA, Burke BJ, Fresh KL (2014) Applied fisheries oceanography: ecosystem indicators of ocean conditions inform fisheries management in the California Current. Oceanography 27:80–89
Peterson WT, Fisher JL, Strub PT, Du X, Risien C, Peterson J, Shaw CT (2017) The pelagic ecosystem in the Northern California Current off Oregon during the 2014–2016 warm anomalies within the context of the past 20 years. J Geophys Res: Oceans 122:7267–7290
Pike GC (1951) Lamprey marks on whales. J Fish Res Board Can 8:275–280
Pinsky ML, Worm B, Fogarty MJ, Sarmiento JL, Levin SA (2013) Marine taxa track local climate velocities. Science 341:1239–1242
Porter LL, Galbreath PF, McIlraith BJ, Hess JE (2017) Sex ratio and maturation characteristics of adult Pacific lamprey at Willamette Falls. Oregon, Columbia River Inter-Tribal Fish Commission, technical report 17-01
Potter IC, Hilliard RW (1987) A proposal for the functional and phylogenetic significance of differences in the dentition of lampreys (Agnatha: Petromyzontiformes). J Zool 212:713–737
Prokhorov VG, Grachev LE (1965) On the record of Pacific lamprey Enthosphenus tridentatus (Gairdner) in the western Bering Sea. Vopr Ikhtiol 4:723–726
Rehage JS, Blanchard JR (2016) What can we expect from climate change for species invasions? J Fish 41:405–407
Renaud CB (2008) Petromyzontidae, Entosphenus tridentatus: southern distribution record, Isla Clarión, Revillagigedo Archipelago, Mexico. Check List 4:82–85
Renaud CB (2011) Lampreys of the world: an annotated and illustrated catalogue of lamprey species known to date. Food and Agriculture Organization of the United Nations, Rome Species Catalogue for Fishery Purposes No 5
Renaud CB, Docker MF, Mandrak NE (2009a) Taxonomy, distribution, and conservation of lampreys in Canada. In: Brown LR, Chase SD, Mesa MG, Beamish RJ, Moyle PB (eds) Biology, management, and conservation of lampreys in North America. Symposium 72. American Fisheries Society, Bethesda, pp 293–310
Renaud CB, Gill HS, Potter IC (2009b) Relationships between the diets and characteristics of the dentition, buccal glands and velar tentacles of the adults of the parasitic species of lamprey. J Zool 278:231–242
Ressler PH, Holmes JA, Fleischer GW, Thomas RE, Cooke KC (2008) Pacific hake, Merluccius productus, autoecology: a timely review. Mar Fish Rev 69:1–24
Riemer SD, Wright BE, Brown RF (2011) Food habits of Steller sea lions (Eumetopias jubatus) off Oregon and northern California, 1986–2007. Fish Bull 109:369–381
Roff TJ, Mate BR (1984) Abundances and feeding habits of pinnipeds in the Rogue River, Oregon. J Wildl Manag 48:1262–1274
Rykaczewski RR, Dunne JP, Sydeman WJ, Garcia-Reyes M, Black BA, Bograd SJ (2015) Poleward displacement of coastal upwelling-favorable winds in the ocean’s eastern boundary currents through the 21st century. Geophys Res Lett 42:6424–6431
Schultz LD, Mayfield MP, Sheoships GT, Wyss LA, Clemens BJ, Whitlock S, Schreck CB (2014) Role of large- and fine-scale variables in predicting catch rates of larval Pacific lamprey in the Willamette basin, Oregon. Ecol Freshw Fish 25:261–271
Semakula SN, Larkin PA (1968) Age, growth, food, and yield of the white sturgeon (Acipenser transmontanus) of the Fraser River, British Columbia. J Fish Res Board Can 25:2589–2602
Sepulveda MS, Sutton TM, Patrick HK (2012) A single sea lamprey attack causes acute anemia and mortality of lake sturgeon. J Aquat Anim Health 24:91–99
Sharma R, Graves D, Farrell A, Mantua N (2016) Investigating freshwater and ocean effects on Pacific lamprey and Pacific Eulachon of the Columbia River Basin: projections within the context of climate change. Columbia River Inter-Tribal Fish Commission, technical report 16-05, 140 electronic pp
Sheoships G (2014) Pacific lamprey Entosphenus tridentatus: integrating ecological knowledge and contemporary values into conservation planning, and stream substrate associations with larval abundance in the Willamette River Basin, Oregon, USA. Master’s thesis, Oregon State University
Shink KG, Sutton TM, Murphy JM, Lopez JA (2019) Utilizing DNA metabarcoding to characterize the diet of marine-phase Arctic lamprey (Lethenteron camtschaticum) in the eastern Bering Sea. Can J Fish Aquat Sci. https://doi.org/10.1139/cjfas-2018-0299
Silva S, Servia MJ, Vieira-Lanero R, Barca S, Cobo F (2013) Life cycle of the sea lamprey Petromyzon marinus: duration of and growth in the marine life stage. Aquat Biol 18:59–62
Silva S, Araújo MJ, Bao M, Mucientes G, Cobo F (2014) The haematophagous feeding stage of anadromous populations of sea lamprey Petromyzon marinus: low host selectivity and wide range of habitats. Hydrobiologia 734:187–199
Silva S, Barca S, Cob F (2016) Advances in the study of sea lamprey Petromyzon marinus Linnaeus, 1758, in the NW of the Iberian Peninsula. In: Orlov AM, Beamish RJ (eds) Jawless fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 346–385
Siwicke KA, Seitz AC (2015) Interpreting lamprey attacks on Pacific Cod in the Eastern Bering Sea. Trans Am Fish Soc 144:1249–1262
Siwicke KA, Seitz AC (2017) Spatial differences in the distributions of Arctic and Pacific lampreys in the Eastern Bering Sea. Trans Am Fish Soc 147:111–127
Sloat MR, Reeves GH (2014) Individual condition, standard metabolic rate, and rearing temperature influence steelhead and rainbow trout (Oncorhynchus mykiss) life histories. Can J Fish Aquat Sci 71:491–501
Somero GN, Beers JM, Chan F, Hill TM, Klinger T, Litvin SY (2016) What changes in the carbonate system, oxygen, and temperature portend for the Northeastern Pacific Ocean: a physiological perspective. Bioscience 66:14–26
Spice EK, Goodman DH, Reid SB, Docker MF (2012) Neither philopatric nor panmictic: microsatellite and mtDNA evidence suggest lack of natal homing but limits to dispersal in Pacific lamprey. Mol Ecol 21:2916–2930
Starcevich SJ, Gunckel SL, Jacobs SE (2014) Movements, habitat use, and population characteristics of adult Pacific lamprey in a coastal river. Environ Biol Fish 97:939–953
Stillwater Sciences (2010) Pacific lamprey in the Eel River basin: a summary of current information and identification of research needs. Stillwater Sciences, Arcata, California for Wiyot Tribe, Loleta, California
Stone J (2006) Observations on nest characteristics, spawning habitat, and spawning behavior of Pacific and Western brook lamprey in a Washington stream. Northwest Nat 87:225–232
Sutton TM (2017) Distribution and ecology of Lethenteron spp. lampreys in interior Alaskan rivers. J Fish Biol 90:1196–1213
Swink WD (2003) Host selection and lethality of attacks by sea lampreys (Petromyzon marinus) in laboratory studies. J Gt Lakes Res 29(Suppl 1):307–319
Tattam IA, Ruzycki JR, Li HW, Giannico GR (2013) Body size and growth rate influence emigration timing of Oncorhynchus mykiss. Trans Am Fish Soc 142:1406–1414
Tinus CA (2012) Prey preference of lingcod (Ophiodon elongatus), a top marine predator: implications for ecosystem-based fisheries management. Fish Bull 110:193–204
Tochigi Prefecture (2005) Tochigi red data book. http://www.pref.tochigi.lg.jp/shizen/sonota/rdb/detail/15/0001.html. Accessed Jan 2019 (in Japanese)
Trites AW, Calkins DG, Winship AJ (2007) Diets of Steller sea lions (Eumetopias jubatus) in southeast Alaska, 1993–1999. Fish Bull 105:234–248
USFWS (US Fish and Wildlife Service) (2004) Endangered and threatened wildlife and plants; 90-day finding on a petition to list three species of lampreys as threatened or endangered. Federal Register 69:257, 27 Dec 2004, pp 77158–77167
USFWS (US Fish and Wildlife Service) (2018) Pacific lamprey Entosphenus tridentatus assessment. 27 Aug 2018 Draft for external review
Van de Wetering SJ (1999) Aspects of life history characteristics and physiological processes in smolting Pacific lamprey, Lampetra tridentata, in a Central Oregon Coast stream. Master’s thesis, Oregon State University
Wade J, Beamish R (2016) Trends in the catches of River and Pacific lamprey in the Strait of Georgia. In: Orlov AM, Beamish RJ (eds) Jawless fishes of the world. Cambridge Scholars Publishing, Newcastle upon Tyne, pp 57–72
Wade AA, Beechie TJ, Fleishman E, Mantua NJ, Wu H, Kimball JS, Stoms DM, Stanford JA (2013) Steelhead vulnerability to climate change in the Pacific Northwest. J Appl Ecol 50:1093–1104
Wainwright TC, Weitkamp LA (2013) Effects of climate change on Oregon Coast coho salmon: habitat and life-cycle interactions. Northwest Sci 87:219–242
Wang C, Schaller H (2015) Conserving pacific lamprey through collaborative efforts. Fisheries 40:72–79
Weitkamp LA, Hinton SA, Bentley PJ (2015) Seasonal abundance, size, and host selection of western river lamprey (Lampetra ayresii) and Pacific (Entosphenus tridentatus) lampreys in the Columbia River estuary. Fish Bull 113:213–226
Whitlock SL, Schultz LD, Schreck CB, Hess JE (2017) Using genetic pedigree reconstruction to estimate effective spawner abundance from redd surveys: an example involving Pacific lamprey Entosphenus tridentatus. Can J Fish Aquat Sci 74:1646–1653
Whyte JNC, Beamish RJ, Ginther NG, Neville C (1993) Nutritional condition of the Pacific lamprey (Lampetra tridentata) deprived of food for periods of up to 2 years. Can J Fish Aquat Sci 50:591–599
Williams JE, Isaak DJ, Imhof J, Hendrickson DA, McMillan JR (2015) Cold-water fishes and climate change in North America. In: Reference module in earth systems and environmental sciences, pp 1–10. https://doi.org/10.1016/b978-0-12-409548-909505-1
Wipfli MS, Baxter CV (2010) Linking ecosystems, food webs, and fish production: subsidies in salmonid watersheds. J Fish 35:373–387
Wolf BO, Jones SL (1989) Great blue heron deaths caused by predation on Pacific lamprey. Condor 91:482–4848
Wright B, Murtaugh T, Brown R (2016) Willamette Falls pinniped monitoring project, 2016. Report by the Oregon Department of Fish and Wildlife, South Willamette Watershed District Office
Zamon JE, Mannas JM, Sandford BP, Evans A, Cramer B (2014) Measuring estuary avian predation on juvenile salmon by electronic recovery of passive integrated transponder tags from nesting colonies, 2013. Report of research for the US Arm Corps of Engineers, Portland District, Nothwestern Division, 69 electronic pp
Zenker A, Cicero MR, Prestinaci F, Bottoni P, Carere M (2014) Bioaccumulation and biomagnification potential of pharmaceuticals with a focus to the aquatic environment. J Environ Manag 133:378–387
Acknowledgements
The idea for this paper was generated by Josh Murauskas. The late Dr. Bill Peterson provided insights on oceanographic regime changes and salmon recruitment at a lamprey workshop that one of us (BJC), along with Dr. David Noakes, convened at the Oregon Hatchery Research Center in Alsea, Oregon, in 2013. Bill’s insights led to our creation of Fig. 4. This paper benefitted from reviews by Tom Stahl, Maggie Sommer, Mary Moser, and Rick Gustafson. Jeremy Five Crows created the image used in Fig. 1. Hiro Arakawa and Ralph Lampman provided insights to the status of Pacific lamprey in Japan. This paper benefitted from the reviews of Sergio Silva and an anonymous reviewer. References to trade names do not imply endorsement by the U.S. Government. The findings and conclusions in this manuscript are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Clemens, B.J., Weitkamp, L., Siwicke, K. et al. Marine biology of the pacific lamprey Entosphenus tridentatus. Rev Fish Biol Fisheries 29, 767–788 (2019). https://doi.org/10.1007/s11160-019-09578-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-019-09578-8