
Abstract
The development of hydropower and other infrastructure that disrupts river connectivity poses a serious threat to highly endemic and genetically distinct freshwater fish species in temperate parts of the Southern Hemisphere. Such locations have been neglected in previous reviews on fish passage. Fishways have long been constructed to mitigate the impacts of riverine barriers on fish, yet they have often failed for all but the largest, strongest swimming taxa. This is a particular problem in the temperate south, which is home to native species that are non-recreational and generally small-bodied with weak swimming abilities (e.g. Galaxiidae) relative to typical target species for fishway design (e.g. Salmonidae). Using the Eco Evidence method for rapid evidence synthesis, we undertook an assessment of evidence for effective fishway design focusing on species representative of the temperate south, including eel and lamprey. Systematic literature searches resulted in 630 publications. Through a rigorous screening process these were reduced to 46 publications containing 76 evidence items across 19 hypotheses relating to design criteria for upstream and downstream passage. We found an overwhelming lack of evidence for effective fishway design in the temperate south. Particular deficiencies were found with regard to the design of effective facilities for downstream passage. The attraction and entrance of upstream migrating fish into fishways is also relatively under-researched. Given the urgent need for effective fishways in the temperate south, these results justify an approach to fishway design based on a combination of empirical data and expert knowledge. In the meantime, significant resources should be assigned to improve the evidence base through high quality research. The particular deficiencies identified here could guide that research agenda.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Given that the majority of freshwater fish species must undertake some form of movement (e.g. for feeding, refuge, reproduction) in order to complete their life-cycle (Schlosser and Angermeier 1995), loss of connectivity caused by hydropower dams poses a serious problem. A range of smaller structures such as low-head hydropower plants, culverts, weirs and tidegates can also represent barriers to fish (Kemp and O’Hanley 2010), and their cumulative impacts can be severe (Larinier 2008; McKay et al. 2013). Pertinent examples include the decline of fall Chinook salmon (Oncorhynchus tshawytscha, Salmonidae) in the Columbia River system, USA (Dauble and Geist 2000), and the disappearance of anadromous species from major rivers in France and northern Spain (e.g. Reyes-Gavilán et al. 1996). Facilities designed to maintain passable conditions for fish have been constructed for centuries but often fail for all but the strongest swimming taxa, such as the salmonids native to northern Europe and North America (Katopodis and Williams 2012), and effective design can be challenging even for these species (Roscoe and Hinch 2010; Noonan et al. 2012; Bunt et al. 2016). The negative environmental and indeed economic consequences of impoundments are such that many large economies in the Northern Hemisphere have begun to remove barriers to migration, including large dams in some cases (Poff and Hart 2002; e.g. East et al. 2015).
The Southern Hemisphere, however, presents a different problem because of intense pressure for rapid economic development and the relative lack of knowledge on the needs of native species (Roscoe and Hinch 2010). Attention has been drawn to the inadequate provision for passage of large, migratory fish in neotropical South America, where hydropower development is especially rapid (Barletta et al. 2010; Zarfl et al. 2015). The conclusion consistently reached by scholars is that designs exported from the Northern Hemisphere are unsuitable for passing diverse neotropical communities (Quirós 1989; Makrakis et al. 2011; Roscoe and Hinch 2010; Duarte and Ramos 2012; Katopodis and Williams 2012). The situation is just as serious in temperate regions of the Southern Hemisphere currently experiencing a hydropower boom (e.g. Chile), which have been neglected in important reviews on fish passage by Quirós (1989), Pringle et al. (2000) and Barletta et al. (2010). The smaller-bodied, non-recreational fish that characterise this region are likely to present even more of a challenge for fishway technology during a global hydropower boom because many hydraulic structures may constitute ‘velocity barriers’ to movement (Link and Habit 2015).
Fish communities of the temperate Southern Hemisphere typically have low species richness but high levels of genetic diversity and endemism (Ormazabal 1993; Gehrke and Harris 2000; Ruzzante et al. 2006; Zemlak et al. 2008; Muñoz-Ramírez et al. 2014). These communities are indicated by the presence of the catadromous common jollytail or ‘inanga’ (Galaxias maculatus, Galaxiidae) and the anadromous pouched lamprey (Geotria australis, Geotridae), whose extant distributions encompass New Zealand and southern parts of Australia, Argentina and Chile (Fig. 1; McDowall 2002). The galaxiids and related taxa are the dominant groups in this zone, comprised of over 50 species from the families Leptogalaxiidae (Australia), Retropinnidae (Australia and New Zealand) and Galaxiidae (whole range). These fish are already experiencing a major decline due to the effects of habitat deterioration, overexploitation and displacement by introduced species (McDowall 2006; Habit et al. 2010). The majority of species native to Chile and 74% of New Zealand’s species are threatened or at risk (Link and Habit 2015; Goodman et al. 2014). The high degree of diadromy in these fish communities (McDowall 2002), and the fact that most upstream migrations occur during juvenile life stages, further exacerbates the problem. Mitigation for fish passage under certain conditions is required by law in all of the aforementioned countries. Article 168 of the Law of Fisheries and Aquaculture in Chile, for example, obligates the owners of barriers that ‘prevent the natural migration of fish’ to ‘carry out a programme of stocking fish’ or ‘build the civil works that allow such migrations’. However, the fishways constructed through implementation of these laws may be more suitable for non-native, recreational species, such as salmonids, rather than native taxa (e.g. Servicio de Evaluación Ambiental 2017).

Joint distribution of Galaxias maculatus and Geotria australis (dashed black lines) delimiting the temperate Southern Hemisphere according to McDowall (2002). Symbols denote the locations of 46 studies included in this review
The overall ‘effectiveness’ of a fishway (sensu Kemp and O’Hanley 2010) is indicated by a suite of metrics describing the ability of individuals of target species to locate and enter the facility and pass the barrier without significant consequences in terms of fitness, i.e. growth, survival and reproduction. We use the term ‘fishway’ in the most general sense to refer to any infrastructure specifically designed to pass fish in an upstream or downstream direction. Effectiveness for upstream passage is a composite of three ‘efficiency’ metrics (Cooke and Hinch 2013). Attraction efficiency describes the proportion of fish motivated to pass the barrier that can locate the entrance to the fishway. Entrance efficiency is the proportion of fish attracted to the entrance that subsequently enter. Passage efficiency is defined as the number of fish exiting the fishway as a proportion of those entering. For downstream passage, guidance efficiency is the proportion of fish passing through the route intended by the design of screens and bypasses, rather than through hydropower turbines. Turbine entrainment may result in injury and mortality due to excessive shear, turbulence and pressure fluctuations, in addition to mechanical injuries such as blade strike (Pracheil et al. 2016a). Thus, ‘fish friendly’ turbine designs are of interest (e.g. Hogan et al. 2014; Dixon and Hogan 2015). The effectiveness of fishways, as well as mortality during turbine entrainment, is related to both engineering parameters (e.g. hydraulics, turbine design) and the biological (e.g. body length, swim bladder morphology) characteristics of target species (Bunt et al. 2012; Pracheil et al. 2016a).
Given the proliferation of hydropower dams and other barriers to fish movement in the temperate south, our aim was to assess the evidence for design criteria that would optimise the effectiveness of fishways for native species in this region. Given the urgency of the situation, we used the Eco Evidence method for rapid evidence synthesis (Norris et al. 2012; Webb et al. 2015) and included evidence on small-bodied and non-recreational species (including eel and lamprey) from anywhere in the world.
Methods
We used the Eco Evidence method, described in full by Norris et al. (2012), because it was specifically designed to rapidly evaluate cause-effect relationships in the environmental sciences. The method is best classified as belonging to the emerging group of ‘rapid review’ methods, which seek to maintain the rigour and objectivity of full ‘systematic review’ (CEBC 2010) methods, inspired by those used in medical sciences, but at a fraction of the cost and time required. Eco Evidence maximises transparency and repeatability, and provides readily interpretable results (Webb et al. 2013). The method centres on the synthesis of evidence items (sensu Webb et al. 2015), which are the summarized findings from a study (hypothesised cause and effect, experimental design classification, presence or absence of cause and effect association). There are eight stages to an Eco Evidence review (Norris et al. 2012). These stages can be consolidated into four broad categories: (1) problem formulation and context; (2) hypothesis generation; (3) literature search and evidence extraction; and (4) evidence assessment and reporting. We describe each of these stages below within a framework that follows the Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) statement (Liberati et al. 2009) as closely as possible for ecological studies (Nakagawa and Poulin 2012).
Problem formulation and context
Our overall research question was: is there sufficient evidence to inform effective fishway design in the temperate Southern Hemisphere? Though our review focused on species native to the temperate south, the scarcity of empirical data relating specifically to these species necessitated a wider scope. We therefore considered evidence relating to any freshwater species globally with a maximum adult body length of <250 mm TL, a broader category than previously proposed for non-recreational (‘non-sport’) fish (<150 mm TL; Link and Habit 2015). This was not intended as a reclassification of non-recreational fish. It was merely a practical definition for the purposes of the review. Our rationale for this was that body length is a fundamental trait influencing swimming speed (Lauder 2015) and mortality due to turbine entrainment (Coutant and Whitney 2000), rendering evidence from larger species of increasingly limited relevance.
Hypothesis generation
We focused on four effects consistent with the literature on fishway effectiveness metrics, namely attraction, entrance, passage and guidance efficiency (Kemp and O’Hanley 2010), plus fish mortality due to turbine entrainment (Table 1). Our initial set of causes was based on our understanding of fishway design criteria from the global literature, which included important contributions from previous reviews that primarily focused on fish native to the Northern Hemisphere (Coutant and Whitney 2000; Larinier and Marmulla 2004; Katopodis 2005; Roscoe and Hinch 2010; Bunt et al. 2012, 2016; Brown et al. 2014; Pracheil et al. 2016a). To further define our hypotheses, we consulted several international fishway design experts. During the literature search we refined our set of hypotheses, excluding, separating or aggregating them as necessary to avoid over-specifying hypotheses in circumstances where there is little evidence available (e.g. Webb et al. 2012). We provide detailed descriptions of the final set of causes in Table 2.
Literature search and evidence extraction
The literature search focused on two databases: ISI Web of Science and the University of Massachusetts Fish Passage Reference Database (EWRI-ACS 2009), which contains theses, unpublished reports, conference proceedings and miscellaneous publications on fish passage. The search strings used when querying the literature databases are provided in Appendix 1 in Supplementary Material. However, Greenhalgh and Peacock (2005) have shown how systematic reviews of complex evidence cannot rely solely on predefined search strategies. Therefore, in addition to the results of the systematic literature search, a number of other sources relevant to the review were included that were sourced through colleagues, our own knowledge of the literature, and from the reference lists of obtained publications. It was impossible to include these in the systematic literature search as they did not appear in either database.
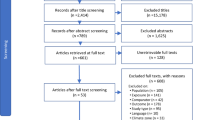
Results of the literature search were filtered by reviewing abstracts or, in the case of several results from the University of Massachusetts database, by scanning the full source. A total of 630 unique articles (Table S1) were filtered down to 72 articles through this initial screening process (Fig. 2). The remaining articles were assessed in full (see below) by at least one assessor but could still be excluded at this stage due to insufficient reporting of results with regards to fishway characteristics or fish response, limitations in the study design (confounding variables) or because results were reported for disqualified (non-anguilliform > 250 mm TL) or ambiguous species (Table S2). After this final screening, we were left with 46 articles containing 76 individual evidence items across the 19 hypotheses (Fig. 2).

Inclusion and exclusion of studies in the systematic review, as suggested by the PRISMA statement (Liberati et al. 2009). n number of studies, e number of evidence items, WoS Web of Science, UMass University of Massachusetts Fish Passage Reference Database
Evidence assessment
An individual article could contain evidence across one or more hypotheses. Each evidence item considered appropriate for inclusion was given a weight based on its inferential strength (a combination of study design and replication, with higher weights attributed to stronger experimental designs) using the standard Eco Evidence weightings (see Norris et al. 2012). For each hypothesis we summed evidence weights supporting the hypothesis and weights refuting the hypothesis. We used the standard Eco Evidence thresholds for assigning a nominal outcome for each hypothesis as ‘support hypothesis’, ‘reject hypothesis’, ‘inconsistent evidence’, ‘insufficient evidence’ or ‘no evidence’ (Fig. 3). The weightings and thresholds used in Eco Evidence were set through an extensive expert consultation process (Norris et al. 2012). However, as the thresholds are somewhat arbitrary, there is a need for careful interpretation when weightings are close to boundaries between outcomes.

Eco Evidence outcome thresholds. Axis units are summed evidence points across evidence items supporting (x) and refuting (y) the hypothesis
For hypotheses with sufficient but somewhat inconsistent evidence (weight > 20 for support and >0 for refute) we decomposed results into sub-hypotheses focusing on three taxonomic groups: (1) angulliform fish; (2) Galaxiidae (including Retropinnidae); and (3) other taxa. These groups were used to reduce potential differences (e.g. rheotactic behaviour) in the response of the taxa considered. If articles reported evidence for multiple taxonomic groups within the same hypothesis, a separate item of evidence was considered for each group.
When evidence for a hypothesis was assessed by more than one assessor we calculated the ‘internal validity’ as the mean and maximum differences in the weightings attributed to individual evidence items across that hypothesis. Where there was a difference in the weighting assigned by different assessors we conservatively used the lower weighting when reporting outcomes. We also recorded ‘external validity’ as the level of agreement between assessors in the inclusion or exclusion of an evidence item.
Results
Internal validity
Out of a total of 121 evidence items remaining after the initial screening, 40 were independently assessed by at least two assessors. There was 100% agreement among assessors about appropriateness of the study design (i.e. whether or not the evidence item was included). There was also complete agreement among assessors about whether the hypothesis was supported or refuted by the evidence. Differences in the weights reported were rare, and in all but one case (hypothesis P4c) equated to only a modest variation (analogous to measurement error in primary data; Webb et al. 2012) in the evidence weight (Table 3, Table S2).
Summary of evidence
The 46 articles retained after completing the screening process came from studies conducted in Europe, North America, New Zealand, Australia, Central America and South America (in order of frequency; Fig. 1). Across the 19 hypotheses tested, plus six sub-hypotheses split by taxonomic group or qualitative cause, the most common outcome was ‘insufficient evidence’ (n = 16, including one instance of ‘no evidence’). Seven hypotheses were supported, one was rejected, and there was one instance of inconsistent evidence (Fig. 4; Table 4).

Attraction efficiency
We did not find sufficient evidence to support either hypothesis relating to attraction efficiency (Fig. 4). Only a small proportion of the evidence gathered across the whole systematic review related to this effect (5%). The majority of evidence items initially screened for hypothesis A1 (↑ Proportion of flow at fishway entrance: ↑ Attraction efficiency) were excluded, largely because they failed to quantify attraction efficiency (Table S2).
Entrance efficiency
Of the four hypotheses relating to entrance efficiency, an outcome of insufficient evidence was returned for hypotheses E1 (↓ Mean water velocity at entrance: ↑ Entrance efficiency) and E4 (↓ Turbulence intensity at entrance: ↑ Entrance efficiency) (Fig. 4). Hypothesis E5 (↓ Drop height: ↑ Entrance efficiency) was strongly supported, containing 9% of all evidence items considered across the review. We were unable to find any evidence for hypothesis E2 (↓ Velocity gradient: ↑ Entrance efficiency).
Passage efficiency
We found insufficient evidence to support hypothesis P1 (Δ Fishway type: ↑ Passage efficiency; Fig. 4). An outcome of inconsistent evidence was returned for hypothesis P2 (↓ Mean water velocity in fishway: ↑ Passage efficiency) but decomposition into taxonomic groups demonstrated sufficient evidence to support the hypothesis for anguilliform and galaxiid species, whilst the hypothesis was rejected by a small margin for other taxa. Evidence relating to hypothesis P2 constituted the largest proportion (27%) of items found across the whole review. Hypothesis P4b (Δ Baffle presence and configuration: ↑ Passage efficiency) was supported across pooled taxa. However, a more detailed analysis showed that there was only sufficient evidence to support this hypothesis for galaxiids. We also found support for P4c (Δ Flow regime: ↑ Passage efficiency) but this related to only two species across three separate studies. Outcomes of insufficient evidence were returned for hypotheses P4a (↓ Turbulence intensity in fishway: ↑ Passage efficiency) and P4d (↑ Climbing substrate: ↑ Passage efficiency).
Guidance efficiency
We found sufficient evidence to support hypothesis G1 (Δ Screen design: ↑ Guidance efficiency) by a narrow margin (Fig. 4). An outcome of insufficient evidence was returned for hypothesis G2 (Δ Bypass design: ↑ Guidance efficiency), with only four evidence items all relating to a single species, Anguilla anguilla Anguillidae (Table S2).
Turbine entrainment (mortality)
We found insufficient evidence to support any of the four hypotheses relating to mortality due to turbine entrainment (Fig. 4), with just four studies contributing evidence.
Discussion
Overall, we found insufficient evidence to inform effective fishway design in the temperate Southern Hemisphere. The only clear exceptions to this (weighting ≫ 20 for support, ≪20 for refute) related to hypotheses P4b (Δ Baffle presence and configuration: ↑ Passage efficiency) and P2 (↓ Mean water velocity in fishway: ↑ Passage efficiency) for Galaxiidae and Retropinnidae, as well as hypothesis E5 (↓ Drop height: ↑ Entrance efficiency) for all taxonomic groups.
Workers in the field of fish passage have consistently bemoaned the disproportionate focus of fishway research and design on large, relatively strong swimming species native to the Northern Hemisphere (Quirós 1989; Roscoe and Hinch 2010; Makrakis et al. 2011; Duarte and Ramos 2012; Katopodis and Williams 2012). Despite this, there have been no previous attempts to synthesise the evidence for fishway design criteria specifically relating to non-recreational species. The large proportion of ‘insufficient evidence’ results returned here supports the contention that non-recreational fish passage is an under-researched area not only in the temperate Southern Hemisphere but in other regions dominated by relatively small-bodied, weak swimming species, such as Turkey (e.g. Kucukali and Hassinger 2016) and the Great Plains of the USA (e.g. Pennock et al. 2017).
Evidence for downstream fishway design
We found a particular deficiency in evidence relating to downstream passage, echoing several previous commentaries highlighting the disproportionate focus on upstream migration in the wider fish passage literature (Kemp and O’Hanley 2010; Pompeu et al. 2012, 2015; Baumgartner et al. 2014). This disproportionate effort risks the creation of ecological traps upstream of barriers and eventually local extinction (Pelicice and Agostinho 2008; Agostinho et al. 2011). The major issues with downstream passage have been cited as mortality due to pressure fluctuations (barotrauma), fluid shear and blade strike during turbine entrainment, and the challenges of guiding fish away from potentially harmful routes (Coutant and Whitney 2000; Katopodis 2005; Brown et al. 2014; Pracheil et al. 2016a).
Though we were able to support hypothesis G1, that a qualitative change in screen design would affect guidance efficiency, the four evidence items included evaluated the performance of three different screen types. Baker and Aldridge (2010) evaluated the effect of modification to a physical screen on three species native to New Zealand (in both anguilliform and Galaxiidae groups). Johnson and Miehls (2013) tested the response of Petromyzon marinus Petromyzontidae to two different electrical screens. Finally, Piper et al. (2015) focused on hydrodynamic screening of migrant A. anguilla. The effectiveness of screens is related to many factors that are highly species- and site- specific (Katopodis 2005), making the definition of general design criteria challenging.
The only evidence for the effectiveness of bypass types (hypothesis G2) and mortality due to blade strike (T2, T3), shear and turbulence (T4) during turbine entrainment was limited to a single species, A. anguilla. Evidence for the effect of pressure fluctuations on mortality due to barotrauma came from just two studies investigating a total of four species; Entosphenus tridentatus Petromyzontidae and Lampetra richardonii Petromyzontidae in Colotelo et al. (2012) and Astyanax bimaculatus Characidae and Leporinus reinhardtii Anostomidae in Pompeu et al. (2009). However, it could be misleading to include lamprey (e.g. E. tridentatus, L. richardonii) together with other species as evidence for this hypothesis. This is because the lack of a swim bladder in lamprey appears to render them insusceptible to barotrauma (Colotelo et al. 2012).
Evidence for upstream fishway design
We found that factors affecting attraction and entrance efficiency have been poorly researched for all taxonomic groups representative of the temperate south. The exception to this was in the case of hypothesis E5, demonstrating that the occurrence of drops between the downstream water surface and the upstream bed level would constitute poor fishway design. The general lack of evidence for attraction efficiency hypotheses is of major concern as poor attraction is one of the primary reasons for fishway failure worldwide (Larinier and Marmulla 2004). Bunt et al. (2012) describe attraction and entrance failure mechanisms as: (1) poor entrance location; (2) insufficient discharge relative to competing flow; and (3) excessive turbulence and velocities. We did not find sufficient evidence to evaluate hypotheses for any of these causes. Furthermore, the majority of evidence we did find was limited to eel and lamprey. We found no evidence at all for hypothesis E2, which is surprising given the wider literature suggesting that fish avoid areas with abrupt velocity accelerations and decelerations, albeit when migrating in a downstream direction (Haro et al. 1998; Kemp et al. 2005; Enders et al. 2009, 2012; Vowles and Kemp 2012; Vowles et al. 2014). It is likely that such extreme conditions are not found at entrances to fishways designed for upstream passage.
Causes relating to passage efficiency had comparatively more evidence. However, several design criteria relating to upstream passage are still not sufficiently researched to reach any general conclusions. For example, evidence for hypothesis P1 (Δ Fishway type: ↑ Passage efficiency) included studies on a range of fishway types. Foulds and Lucas (2013) and Matondo et al. (2015) compared the passage efficiency of Denil and vertical slot fishways respectively with pool-and-weir designs. The former found extremely low passage efficiencies of 0% (Denil fishway) and 5% (pool-and-weir) for Lampetra fluviatilis Petromyzontidae. Stuart et al. (2008a) evaluated passage efficiency improvements resulting from the modification of a vertical slot fishway with lock gates, finding inconsistent effects of the modification among three non-recreational species. Noonan et al’s (2012) meta-analysis reported passage efficiencies for a variety of fishway types, whereas Newbold et al. (2014) focused on alternative designs for culverts. Clearly, these studies do not form coherent evidence that is able to inform fishway design in any detail, and this may partly explain the inconsistency of findings.
Mean velocity or fishway slope (hypothesis P2) was by far the most well-researched design criterion, with a total of 20 individual evidence items. We found support for the hypothesis that the passage of anguilliform and galaxiid species is improved as mean velocity or longitudinal slope is decreased. For other species there was a greater weight of evidence for the opposite effect. The majority of evidence refuting hypothesis P2 for non-anguilliform and non-galaxiid species comes from two studies that tested the passage of fish through culverts (Bouska and Paukert 2010; David and Hamer 2012), a somewhat different context to fishway design at hydropower barriers. Further evidence of a negative correlation between passage efficiency and mean velocity came from the meta-analysis of Noonan et al. (2012), reported for all non-salmonid taxa together. We therefore interpret this finding with caution.
We found that there was insufficient evidence to support hypothesis P3, that passage efficiency would increase with decreasing fishway length. Minimisation of fishway length has often been included in best practice guidelines for the hydraulic design of fishways, for reasons relating to swimming performance and energetics (e.g. Laborde et al. 2016). However, we found a comparable weight of evidence supporting and refuting this hypothesis. The evidence contributing to this outcome comes from two very different contexts: Baker (2014) evaluated passage efficiency over ramps up to 6 m long, whereas Noonan et al. (2012) included full-scale fishways. The latter found a positive relationship between fishway length and passage efficiency but this correlation was likely at least partly driven by the negative relationship between fishway length and slope, with slope being the decisive factor. There is clearly a need to disentangle the effects of these key design criteria.
Turbulence (hypothesis P4a) and the installation of baffles (P4b) have also been included in design criteria for fishways for reasons relating to swimming energetics (Feurich et al. 2012; Bretón et al. 2013; Baki et al. 2014a, b). However, we found insufficient evidence for the benefits of these design parameters. The exception to this was for the response of galaxiids to baffle design, which was among our most strongly supported hypotheses. Several studies support the installation of complex baffle arrangements and rough substrates to improve the passage of galaxiid and related species (e.g. Baker and Boubée 2006; MacDonald and Davies 2007; Mallen-Cooper et al. 2008). Turbulence is a complex phenomenon that can be described in a variety of ways, including intensity, periodicity, orientation and scale (Lacey et al. 2012). The elucidation of relationships between fish swimming performance and turbulence, especially in the context of fishway design, remains a major challenge (Wilkes et al. 2013).
We were able to support hypothesis P4c, that a qualitative change in flow regime affects passage efficiency. However, the three studies that contributed to this evidence suggested opposite effects for two species. Piper et al. (2012) found that A. anguilla passage over an intertidal weir was significantly higher under plunging flow, whereas Branco et al. (2013a, b) reported that Squalius pyrenaicus Cyprinidae passage through an experimental pool-and-weir fishway was significantly higher under streaming flow. This conflicting evidence points to fundamental differences in the behaviour of eels and other species around barriers. Eel passage may be stimulated under plunging flow because the flow regime has a greater influence on velocities and turbulence near the water surface, where juveniles are more likely to be migrating (Clay 1995; Tesch 2003), whereas other non-recreational species lack the strong rheotactic behaviour to deal with these conditions and may become disorientated by the flow patterns found under plunging flow regimes (Branco et al. 2013a, b).
A key characteristic of several species of the temperate south is their ability to climb vertical surfaces (e.g. Galaxias fasciatus Galaxiidae, Gobiomorphus huttoni Eleotridae, Anguilla dieffenbachii Anguillidae). We therefore tested hypothesis P4d, that the presence and type of climbing substrate would affect passage efficiency. However, we found only one study evaluating this hypothesis (David and Hamer 2012), which reported an increase in passage when mussel spat ropes were installed at a perched culvert entrance, and for only one (juvenile G. fasciatus) of three climbing species native to New Zealand. This may be due to the different climbing styles exhibited by the three species. For example, G. fasciatus uses its whole body to climb in a continuous movement, whereas G. huttoni attaches intermittently with specialised pectoral fins. This apparent difference justifies further research into the efficacy of a variety of substrates to improve the passage of climbing species.
Fish passage research needs
We found that most studies excluded at the screening stage were highly site-specific, lacking a sufficiently robust study design to generate transferable knowledge. Another shortcoming apparent in the extant literature is the lack of standardised reporting of efficiency estimates and variances, a problem also highlighted by Bunt et al. (2012, 2016) and Pracheil et al. (2016a) for the global literature on upstream passage and turbine entrainment respectively. The omission of standardised estimates and variances precludes the use of formal meta-analyses (Williams and Katopodis 2016).
Roscoe and Hinch (2010) found that less than 5% of published fishway evaluations had been performed in the temperate south. The lack of studies investigating passage failure mechanisms is a particular deficiency of work in the Southern Hemisphere (Roscoe and Hinch 2010). Not a single study from the Southern Hemisphere met the criteria for inclusion in the meta-analyses on the performance of fishways by Bunt et al. (2012, 2016). These criteria were: (1) fish were individually monitored using biotelemetry; (2) data were from migratory fish actively migrating within a single spawning season; and (3) observations were made under natural conditions without any intervention to coerce fish into ascending structures. In part, the exclusion of work from the temperate south based on such criteria is due to the difficulty or impossibility of adopting currently available biotelemetry technologies for use with small-bodied fish. For example, even the smallest passive integrated transponder (PIT) tags are approximately 25% of the length and 10% of the weight of a juvenile galaxiid (e.g. G. maculatus; Chapman et al. 2006).
Direct fishway evaluations in the temperate south often rely on trapping campaigns at the upstream and downstream ends of fishways on alternate days, with a comparison of species composition and body length distributions indicating the degree to which the fishway is effective (e.g. Stuart et al. 2008b). Whilst this is a pragmatic study design, it fails to produce a metric of passage efficiency, ignores attraction and entrance efficiency, and is not able to indicate mechanisms for passage success or failure. An alternative approach has been to stain and release fish downstream of a fishway (e.g. Amtstaetter et al. 2015). Recapture rates in a netted section upstream can then be used to infer passage efficiency. However, this method is susceptible to losses of stained fish and is not suitable in large rivers where isolating a section with nets would be impractical.
There is an urgent need to develop a set of methods for robust design and evaluation of fishways in the temperate south. This may include PIT and acoustic tagging, although experimental work is required to check what, if any, negative impacts on mortality and swimming performance result from using miniaturised PIT and acoustic tags in non-recreational fish. Further laboratory work on swimming performance and behaviour in prototype fishways will also be useful to define biological design criteria for non-recreational species (e.g. Laborde et al. 2016). This experimental approach should be extended to include the assessment of screen and bypass designs and the definition of safe thresholds for shear and pressure fluctuations (Brown et al. 2014). Such dose–response studies are expensive and time-intensive. Thus, attempts should be made to synthesise work already completed on a range of recreational species worldwide in order to derive transferrable knowledge for native species of the temperate Southern Hemisphere. There is also a need to model blade strike injury and mortality for a range of turbine designs and discharges (e.g. Deng et al. 2011). All such modelling and laboratory results must be validated in situ in order to incorporate the full set of processes influencing fishway efficiency in complex, real-world situations (e.g. Baumgartner et al. 2012).
Our analyses confirm that vastly less is known about fishway design for non-recreational fish than for salmonids. The evidence is not well documented even for anguilliform species, which have been the focus of much legislation and conservation effort globally (Haro et al. 2000; Dekker 2003; Masters et al. 2006; Bark et al. 2007; Lucas et al. 2009; Russon and Kemp 2011). The mean overall upstream fishway effectiveness (21%) and the average delay to migration (5.5 days) reported by Noonan et al. (2012) for non-salmonids suggest serious consequences of current fishway designs for the vast majority of species. The current global hydropower boom is affecting areas of the world with diverse non-salmonid species (Zarfl et al. 2015), including the mega-diverse Amazon and Mekong basins (Winemiller et al. 2016). A major problem in tropical basins is the trapping of fish eggs and larvae in quiescent impounded sections, rendering fishway effectiveness a concept of limited relevance (Pompeu et al. 2012). Although the Eco Evidence approach would be suitable for application to this problem, our review has not considered this aspect. This is, in part, because this life-history trait is less prevalent in temperate systems. However, research on ecological traps (Pelicice and Agostinho 2008) and sources and sinks (Godinho and Kynard 2009) caused by impoundments in the temperate south would help to fill this knowledge gap. Such research would ensure that effective fishway design is contributing positively to the viability of populations, rather than facilitating passage into ecological traps, and thus condemning populations to eventual local extinction.
Our inclusion of any species with an adult body length of <250 mm TL plus anguilliform species of any length encompasses fish of a huge diversity of life-histories, behaviours and ecologies. This introduces uncertainties into our review and may be at least partially responsible for our evaluation of inconsistent evidence in the case of hypothesis P2 (↓ Mean water velocity in fishway: ↑ Passage efficiency) for non-galaxiid and non-anguilliform species, which included cypriniform, perciform, characiform and siluriform taxa (Table 4; Table S2). One major challenge for fish passage research globally is to identify and catalogue the traits that are decisive in species’ variable responses to fishway design and hydropower development in general. As a minimum, such a trait catalogue should include information on body length, migration timings and cues, swimming performance, lateral line development, swim bladder morphology and migration depth. These latter two traits are critical to the susceptibility of fish to barotrauma, a previously neglected impact of hydropower turbines and other river infrastructure (Brown et al. 2014; Pracheil et al. 2016a). Limited fish trait information is already available in databases with worldwide (Froese and Pauly 2016) and national (e.g. USA; Frimpong and Angermeier 2009) coverage, and has already been applied in the context of hydropower monitoring in the USA (Pracheil et al. 2016b), but more work is required to develop the full set of relevant traits, particularly for species native to regions outside of the temperate Northern Hemisphere. By allowing the identification of functional groups, compilation of these traits would help to direct efforts to transfer knowledge between biogeographical regions and taxonomic groups, as we have attempted to do in this review.
Conclusions
There is currently very little evidence to support the design of effective fishways for non-recreational fish native to the temperate south, a geographical context that includes areas presently experiencing rapid hydropower development. More research is urgently required in areas relating to attraction, entrance and guidance efficiency and turbine entrainment, but with more robust experimental designs that allow findings to be transferred beyond the system being studied. The most urgent needs are for research into effective design of downstream passage facilities, otherwise there is a risk that resources used to construct effective upstream fishways are wasted when downstream migrants suffer high levels of mortality. This lack of empirical evidence justifies the combination of available data, modelling outputs and expert judgement for informing fishway design decisions until sufficient (and sufficiently robust) data can be collected.
References
Agostinho CS, Pelicice FM, Marques EE, Soares AB, de Almeida DAA (2011) All that goes up must come down? Absence of downstream passage through a fish ladder in a large Amazonian river. Hydrobiol 675(1):1–12
Amtstaetter F, O’Connor J, Borg D, Stuart I, Moloney P, Campbell-Beshorner R (2015) Improving the passage of migrating Galaxias through a culvert. Arthur Rylah Institute for Environmental Research Unpublished Client Report for Melbourne Water, Department of Environment, Land, Water and Planning, Heidelberg, Victoria
Baker CF (2003) Effect of fall height and notch shape on the passage of inanga (Galaxias maculatus) and common bullies (Gobiomorphus cotidianus) over an experimental weir. N Z J Mar Freshw Res 37(2):283–290
Baker CF (2014) Effect of ramp length and slope on the efficacy of a baffled fish pass. J Fish Biol 84(2):491–502
Baker C, Aldridge B (2010) Huntly Power Station—2010 Fisheries Monitoring Report. NIWA Client Report HAM2010-109 September 2010
Baker CF, Boubée JAT (2006) Upstream passage of inanga Galaxias maculatus and redfin bullies Gobiomorphus huttoni over artificial ramps. J Fish Biol 69(3):668–681
Baki ABM, Zhu DZ, Rajaratnam N (2014a) Mean flow characteristics in a rock-ramp-type fish pass. J Hydraul 140(2):156–168
Baki ABM, Zhu DZ, Rajaratnam N (2014b) Turbulence characteristics in a rock-ramp-type fish pass. J Hydraul 141(2):04014075
Bark A, Williams B, Knights B (2007) Current status and temporal trends in stocks of European eel in England and Wales. ICES J Mar Sci 64:1368–1378
Barletta M, Jaureguizar AJ, Baigun C, Fontoura NF, Agostinho AA, Almeida-Val VMF et al (2010) Fish and aquatic habitat conservation in South America: a continental overview with emphasis on neotropical systems. J Fish Biol 76:2118–2176
Baumgartner LJ, Marsden T, Singhanouvong D, Phonekhampheng O, Stuart IG, Thorncraft G (2012) Using an experimental in situ fishway to provide key design criteria for lateral fish passage in tropical rivers: a case study from the Mekong River, central Lao PDR. River Res Appl 28(8):1217–1229
Baumgartner L, Zampatti B, Jones M, Stuart I, Mallen-Cooper M (2014) Fish passage in the Murray-Darling Basin, Australia: not just an upstream battle. Ecol Restor Manag 15(S1):28–39
Bouska WW, Paukert CP (2010) Road crossing designs and their impact on fish assemblages of Great Plains streams. Trans Am Fish Soc 139(1):214–222
Branco P, Santos JM, Katopodis C, Pinheiro A, Ferreira MT (2013a) Effect of flow regime hydraulics on passage performance of Iberian chub (Squalius pyrenaicus) (Günther, 1868) in an experimental pool-and-weir fishway. Hydrobiol 714(1):145–154
Branco P, Santos JM, Katopodis C, Pinheiro A, Ferreira MT (2013b) Pool-type fishways: two different morpho-ecological cyprinid species facing plunging and streaming flows. PLoS ONE 8(5):e65089
Bretón F, Baki ABM, Link O, Zhu DZ, Rajaratnam N (2013) Flow in nature-like fishway and its relation to fish behaviour. Can J Civ Eng 40(6):567–573
Brown RS, Colotelo AH, Pflugrath BD, Boys CA, Baumgartner LJ, Deng ZD, Silva LGM et al (2014) Understanding barotrauma in fish passing hydro structures: a global strategy for sustainable development of water resources. Fisheries 39(3):108–122
Bunt CM, Castro-Santos T, Haro A (2012) Performance of fish passage structures at upstream barriers to migration. River Res Appl 28(4):457–478
Bunt CM, Castro-Santos T, Haro A (2016) Reinforcement and validation of the analyses and conclusions related to fishway evaluation data from Bunt et al.: ‘performance of fish passage structures at upstream barriers to migration’. River Res Appl. doi:10.1002/rra.3095
Buysse D, Mouton AM, Stevens M, den Neucker T, Coeck J (2013) Mortality of European eel after downstream migration through two types of pumping stations. Fish Manag Ecol 21(1):13–21
Buysse D, Mouton AM, Baeyens R, Stevens M, den Neucker T, Coeck J (2015) Evaluation of downstream migration mitigation actions for eel at an Archimedes screw pump pumping station. Fish Manag Ecol 22(4):286–294
Calles O, Karlsson S, Hebrand M, Comoglio C (2012) Evaluating technical improvements for downstream migrating diadromous fish at a hydroelectric plant. Ecol Eng 48:30–37
CEBC (2010) Guidelines for systematic review in environmental management. Centre for Evidence-Based Conservation and Collaboration for Environmental Evidence, Bangor
Chapman A, Morgan DL, Beatty SJ, Gill HS (2006) Variation in life history of land-locked lacustrine and riverine populations of Galaxias maculatus (Jenyns 1842) in Western Australia. Environ Biol Fishes 77(1):21–37
Clay CH (1995) Design of Fishways and other Fish Facilities. Lewis, Boca Raton
Colotelo AH, Pflugrath BD, Brown RS, Brauner CJ, Mueller RP, Carlson TJ, Trumbo BA et al (2012) The effect of rapid and sustained decompression on barotrauma in juvenile brook lamprey and Pacific lamprey: implications for passage at hydroelectric facilities. Fish Res 129:17–20
Cooke SJ, Hinch SG (2013) Improving the reliability of fishway attraction and passage efficiency estimates to inform fishway engineering, science and practice. Ecol Eng 58:123–132
Cooney PB, Kwak TJ (2013) Spatial extent and dynamics of dam impacts on tropical island freshwater fish assemblages. Bioscience 63(3):176–190
Coutant CC, Whitney RR (2000) Fish behavior in relation to passage through hydropower turbines: a review. Trans Am Fish Soc 129(2):351–380
da Silva LGMD, Nogueira LB, Maia BP, Resende LBD (2012) Fish passage post-construction issues: analysis of distribution, attraction and passage efficiency metrics at the Baguari Dam fish ladder to approach the problem. Neotrop Ichthyol 10(4):751–762
Dauble DD, Geist DR (2000) Comparison of mainstem spawning habitats for two populations of fall Chinook salmon in the Columbia River Basin. Regul Rivers 16(4):345–361
David BO, Hamer MP (2012) Remediation of a perched stream culvert with ropes improves fish passage. Mar Freshw Res 63(5):440–449
David BO, Tonkin JD, Taipeti KW, Hokianga HT (2014) Learning the ropes: mussel spat ropes improve fish and shrimp passage through culverts. J Appl Ecol 51(1):214–223
Dekker W (2003) Did lack of spawners cause the collapse of the European eel, Anguilla anguilla? Fish Manag Ecol 10:365–376
Deng Z, Carlson TJ, Dauble DD, Ploskey GR (2011) Fish passage assessment of an advanced hydropower turbine and conventional turbine using blade-strike modeling. Energies 4(1):57–67
Dixon D, Hogan T (2015) Alden fish-friendly hydropower turbine: history and development status. In: International conference on engineering and ecohydrology for fish passage, Groningen, June 2015
Doehring K, Young RG, McIntosh AR (2011) Factors affecting juvenile galaxiid fish passage at culverts. Mar Freshw Res 62(1):38–45
Doehring K, Young RG, McIntosh AR (2012) Facilitation of upstream passage for juveniles of a weakly swimming migratory galaxiid. N Z J Mar Freshw Res 46(3):303–313
Duarte BADF, Ramos ICR (2012) Reynolds shear-stress and velocity: positive biological response of neotropical fishes to hydraulic parameters in a vertical slot fishway. Neotrop Ichthyol 10(4):813–819
Durif C, Elie P, Gosset C, Rives J, Travade F (2002) Behavioral study of downstream migrating eels by radio-telemetry at a small hydroelectric power plant. In: Dixon DA (ed) Biology, management and protection of catadromous eels. American Fisheries Society, Bethesda, pp 343–356
East AE, Pess GR, Bountry JA, Magirl CS, Ritchie AC, Logan JB, Randle TJ et al (2015) Large-scale dam removal on the Elwha River, Washington, USA: river channel and floodplain geomorphic change. Geomorphology 228:765–786
Enders EC, Gessel MH, Williams JG (2009) Development of successful fish passage structures for downstream migrants requires knowledge of their behavioural response to accelerating flow. Can J Fish Aquat Sci 66(12):2109–2117
Enders EC, Gessel MH, Anderson JJ, Williams JG (2012) Effects of decelerating and accelerating flows on juvenile salmonid behavior. Trans Am Fish Soc 141(2):357–364
EWRI-ACS (2009) Joint EWRI-AFS Fish Passage Reference Database (online). http://scholarworks.umass.edu/fishpassage_database/. Accessed 4 Oct 2016
Feurich R, Boubée J, Olsen NRB (2012) Improvement of fish passage in culverts using CFD. Ecol Eng 47:1–8
Ficke AD, Myrick CA, Jud N (2011) The swimming and jumping ability of three small Great Plains fishes: implications for fishway design. Trans Am Fish Soc 140(6):1521–1531
Foulds WL, Lucas MC (2013) Extreme inefficiency of two conventional, technical fishways used by European river lamprey (Lampetra fluviatilis). Ecol Eng 58:423–433
Frimpong EA, Angermeier PL (2009) Fish traits: a database of ecological and life-history traits of freshwater fishes of the United States. Fisheries 34(10):487–495
Froese R, Pauly D (eds) (2016) FishBase. www.fishbase.org
Gehrke PC, Harris JH (2000) Large-scale patterns in species richness and composition of temperate riverine fish communities, south-eastern Australia. Mar Freshw Res 51(2):165–182
Godinho AL, Kynard B (2009) Migratory fishes of Brazil: life history and fish passage needs. River Res Appl 25(6):702–712
Goodman JM, Dunn NR, Ravenscroft PJ, Allibone RM, Boubee JA, David BO, Rolfe JR et al (2014) Conservation status of New Zealand freshwater fish, 2013. N Z Threat Classif Ser 7:12
Gosset C, Travade F, Durif C, Rives J, Elie P (2005) Tests of two types of bypass for downstream migration of eels at a small hydroelectric power plant. River Res Appl 21(10):1095–1105
Greenhalgh T, Peacock R (2005) Effectiveness and efficiency of search methods in systematic reviews of complex evidence: audit of primary sources. BMJ 331:1064–1065
Habit E, Piedra P, Ruzzante DE, Walde SJ, Belk MC, Cussac VE, Colin N et al (2010) Changes in the distribution of native fishes in response to introduced species and other anthropogenic effects. Glob Ecol Biogeogr 19(5):697–710
Haro A, Odeh M, Noreika J, Castro-Santos T (1998) Effect of water acceleration on downstream migratory behavior and passage of Atlantic salmon smolts and juvenile American shad at surface bypasses. Trans Am Fish Soc 127(1):118–127
Haro A, Castro-Santos T, Boubée J (2000) Behaviour and passage of silver-phase eels, Anguilla rostrata (LeSueur), at a small hydroelectric facility. Dana 12:33–42
Hicks B, Roper M, Ito T, Campbell D (2008) Passage of inanga (Galaxias maculatus) over artificial ramps as a means to restoring upstream access to stream habitat. Centre for Biological and Ecological Research Contract Report 89
Hogan TW, Cada GF, Amaral SV (2014) The status of environmentally enhance hydropower turbines. Fisheries 39(4):164–172
Holthe E, Lund E, Finstad B, Thorstad EB, McKinley RS (2005) A fish selective obstacle to prevent dispersion of an unwanted fish species, based on leaping capabilities. Fish Manag Ecol 12(2):143–147
Johnson NS, Miehls S (2013) Guiding out-migrating juvenile sea lamprey (Petromyzon marinus) with pulsed direct current. River Res Appl 30(9):1146–1156
Johnson EL, Caudill CC, Keefer ML, Clabough TS, Peery CA, Jepson MA, Moser ML (2012) Movement of radio-tagged adult Pacific lampreys during a large-scale fishway velocity experiment. Trans Am Fish Soc 141(3):571–579
Katopodis C (2005) Developing a toolkit for fish passage, ecological flow management and fish habitat works. J Hydraul Res 43(5):451–467
Katopodis C, Williams JG (2012) The development of fish passage research in a historical context. Ecol Eng 48:8–18
Keefer ML, Peery CA, Lee SR, Daigle WR, Johnson EL, Moser ML (2011) Behaviour of adult Pacific lamprey in near-field flow and fishway design experiments. Fish Manag Ecol 18(3):177–189
Kemp PS, O’Hanley JR (2010) Procedures for evaluating and prioritising the removal of fish passage barriers: a synthesis. Fish Manag Ecol 17(4):297–322
Kemp PS, Gessel MH, Williams JG (2005) Fine-scale behavioral responses of Pacific salmonid smolts as they encounter divergence and acceleration of flow. Trans Am Fish Soc 134:390–398
Kemp PS, Russon IJ, Vowles AS, Lucas MC (2011) The influence of discharge and temperature on the ability of upstream migrant adult river lamprey (Lampetra fluviatilis) to pass experimental overshot and undershot weirs. River Res Appl 27(4):488–498
Kucukali S, Hassinger R (2016) Flow and turbulence structure in a baffle–brush fish pass. In: Proceedings of the Institution of Civil Engineers-Water Management. Thomas Telford Ltd, London, pp 1–12
Laborde A, González A, Sanhueza C, Arriagada P, Wilkes M, Habit E, Link O (2016) Hydropower development, riverine connectivity, and non-sport fish species: criteria for hydraulic design of fishways. River Res Appl. doi:10.1002/rra.3040
Lacey RWJ, Neary VS, Liao JC, Enders EC, Tritico HM (2012) The IPOS framework: linking fish swimming performance in altered flows from laboratory experiments to rivers. River Res Appl 28(4):429–443
Larinier M (2008) Fish passage experience at small-scale hydro-electric power plants in France. Hydrobiol 609(1):97–108
Larinier M, Marmulla G (2004) Fish passes: types, principles and geographical distribution—an overview. In: Second international symposium on the management of large rivers for fisheries, Phnom Penh, February, 2004
Lauder GV (2015) Fish locomotion: recent advances and new directions. Ann Rev Mar Sci 7:521–545
LeMoine MT, Bodensteiner LR (2014) Barriers to upstream passage by two migratory sculpins, prickly sculpin (Cottus asper) and coastrange sculpin (Cottus aleuticus), in northern Puget Sound lowland streams. Can J Fish Aquat Sci 71(11):1758–1765
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JP, Clarke M et al (2009) The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. Ann Intern Med 151(4):W-65
Link O, Habit E (2015) Requirements and boundary conditions for fish passes of non-recreational fish species based on Chilean experiences. Rev Environ Sci Biol 2015:1–13
Lucas MC, Bubb DH, Jang MH, Ha K, Masters JEG (2009) Availability of and access to critical habitats in regulated rivers: effects of low-head barriers on threatened lampreys. Freshw Biol 54:621–634
MacDonald JI, Davies PE (2007) Improving the upstream passage of two galaxiid fish species through a pipe culvert. Fish Manag Ecol 14(3):221–230
Makrakis S, Miranda LE, Gomes LC, Makrakis MC, Junior HMF (2011) Ascent of neotropical migratory fish in the Itaipu reservoir fish pass. River Res Appl 27(4):511–519
Mallen-Cooper M, Zampatti BP, Stuart IG, Baumgartner LJ (2008) Innovative fishways-manipulating turbulence in the vertical-slot design to improve performance and reduce cost. A report to the Murray Darling Basin Commission, Fishway Consulting Services, Sydney
Marohn L, Prigge E, Hanel R (2014) Escapement success of silver eels from a German river system is low compared to management-based estimates. Freshw Biol 59(1):64–72
Masters JEG, Jang MH, Ha K, Bird PD, Frear PA, Lucas MC (2006) The commercial exploitation of a protected anadromous species, the river lamprey (Lampetra fluviatilis L.), in the tidal River Ouse, north-east England. Aquat Conserv 16:77–92
Matondo BN, Dierckx A, Benitez JP, Ovidio M (2015) Does yellow eel prefer old pool and weir or new vertical slot fish pass during their upstream migration? In: International conference on engineering and ecohydrology for fish passage, Groningen, June 2015
McDowall RM (2002) Accumulating evidence for a dispersal biogeography of southern cool temperate freshwater fishes. J Biogeogr 29(2):207–219
McDowall RM (2006) Crying wolf, crying foul, or crying shame: alien salmonids and a biodiversity crisis in the southern cool-temperate galaxioid fishes? Rev Fish Biol Fish 16(3–4):233–422
McKay SK, Schramski JR, Conyngham JN, Fischenich JC (2013) Assessing upstream fish passage connectivity with network analysis. Ecol Appl 23(6):1396–1409
Moser ML, Matter AL, Stuehrenberg LC, Bjornn TC (2002) Use of an extensive radio receiver network to document Pacific lamprey (Lampetra tridentata) entrance efficiency at fishways in the Lower Columbia River, USA. Aquat Telem 165:5–53
Muñoz-Ramírez CP, Unmack PJ, Habit E, Johnson JB, Cussac VE, Victoriano P (2014) Phylogeography of the ancient catfish family Diplomystidae: biogeographic, systematic, and conservation implications. Mol Phylogenet Evol 73:146–160
Nakagawa S, Poulin R (2012) Meta-analytic insights into Evol Ecol: an introduction and synthesis. Evol Ecol 26(5):1085–1099
Newbold LR, Karageorgopoulos P, Kemp PS (2014) Corner and sloped culvert baffles improve the upstream passage of adult European eels (Anguilla anguilla). Ecol Eng 73:752–759
Noonan MJ, Grant JW, Jackson CD (2012) A quantitative assessment of fish passage efficiency. Fish Fish (Oxf) 13(4):450–464
Norris RH, Webb JA, Nichols SJ, Stewardson MJ, Harrison ET (2012) Analyzing cause and effect in environmental assessments: using weighted evidence from the literature. Freshw Sci 31:5–21
Ormazabal C (1993) The conservation of biodiversity in Chile. Rev Chil Hist Nat 66(4):383–402
Pelicice FM, Agostinho AA (2008) Fish passage facilities as ecological traps in large Neotropical rivers. Conserv Biol 22:180–188
Pennock CA, Bender D, Hofmeier J, Mounts JA, Waters R, Weaver VD, Gido KB (2017) Can fishways mitigate fragmentation effects on great plains fish communities? Can J Fish Aquat Sci. doi:10.1139/cjfas-2016-0466
Piper AT, Wright RM, Kemp PS (2012) The influence of attraction flow on upstream passage of European eel (Anguilla anguilla) at intertidal barriers. Ecol Eng 44:329–336
Piper AT, Manes C, Siniscalchi F, Marion A, Wright RM, Kemp PS (2015) Response of seaward-migrating European eel (Anguilla anguilla) to manipulated flow fields. Proc Biol Sci 282(1811):20151098
Poff NL, Hart DD (2002) How Dams Vary and Why it Matters for the Emerging Science of Dam Removal: An ecological classification of dams is needed to characterize how the tremendous variation in the size, operational mode, age, and number of dams in a river basin influences the potential for restoring regulated rivers via dam removal. Bioscience 52(8):659–668
Pompeu PDS, Horta LFM, Martinez CB (2009) Evaluation of the effects of pressure gradients on four Brazilian freshwater fish species. Braz Arch Biol Technol 52(1):111–118
Pompeu PS, Agostinho AA, Pelicice FM (2012) Existing and future challenges: the concept of successful fish passage in South America. River Res Appl 28(4):504–512
Pompeu PS, Suzuki FM, Prado IG, Souza RCR (2015) Downstream fish passage: the new challenge of the hydropower sector for the conservation of the brazilian fish fauna. Am J Hydropower Water Environ Syst. doi:10.14268/ajhwes.2015.00028
Pracheil BM, DeRolph CR, Schramm MP, Bevelhimer MS (2016a) A fish-eye view of riverine hydropower systems: the current understanding of the biological response to turbine passage. Rev Fish Biol Fish. doi:10.1007/s11160-015-9416-8
Pracheil BM, McManamay RA, Bevelhimer MS, DeRolph CR, Čada GF (2016b) A traits-based approach for prioritizing species for monitoring and surrogacy selection. Endanger Species Res 31:243–258
Pringle CM, Freeman MC, Freeman BJ (2000) Regional effects of hydrlogic alterations on riverine macrobiota in the new world: tropical-temperate comparisons. Bioscience 50(9):807–823
Quirós R (1989) Structures assisting the migrations of non-salmonid fish: Latin America. COPESCAL Technical Paper, no. 5, United Nations, Rome
Reyes-Gavilán FG, Garrido R, Nicieza AG, Toledo MM, Brana F (1996) Fish community variation along physical gradients in short streams of northern Spain and the disruptive effect of dams. Hydrobiol 321(2):155–163
Roscoe DW, Hinch SG (2010) Effectiveness monitoring of fish passage facilities: historical trends, geographic patterns and future directions. Fish Fish (Oxf) 11(1):12–33
Russon IJ, Kemp PS (2011) Experimental quantification of the swimming performance and behaviour of spawning run river lamprey Lampetra fluviatilis and European eel Anguilla anguilla. J Fish Bio 78(7):1965–1975
Ruzzante DE, Walde SJ, Cussac VE, Dalebout ML, Seibert J, Ortubay S, Habit E (2006) Phylogeography of the Percichthyidae (Pisces) in Patagonia: roles of orogeny, glaciation, and volcanism. Mol Ecol 15(10):2949–2968
Schlosser IJ, Angermeier PL (1995) Spatial variation in demographic processes in lotic fishes: conceptual models, empirical evidence, and implications for conservation. Am Fish Soc Symp 17:360–370
Servicio de Evaluación Ambiental (2017) Estudio de Impacto Ambiental ‘Central Hidroeléctrica del Río Chaica’ (online). http://seia.sea.gob.cl/documentos/documento.php?idDocumento=2131795393. Accessed 6 April 2017
Stuart IG, Berghuis AP, Long PE, Mallen-Cooper M (2007) Do fish locks have potential in tropical rivers? River Res Appl 23(3):269–286
Stuart IG, Baumgartner LJ, Zampatti BP (2008a) Lock gates improve passage of small-bodied fish and crustaceans in a low gradient vertical-slot fishway. Fish Manag Ecol 15(3):241–248
Stuart IG, Zampatti BP, Baumgartner LJ (2008b) Can a low-gradient vertical-slot fishway provide passage for a lowland river fish community? Mar Freshw Res 59:332–346
Tesch FW (2003) The Eel. Blackwell, Oxford
Tummers JS, Winter E, Silva S, O’Brien P, Jang MH, Lucas MC (2015) Tricky Little Lampreys! Efficacy of an unmodified and modified super-active baffle fish pass for European river lamprey (Lampetra Fluviatilis). In: International conference on engineering and ecohydrology for fish passage, Groningen, June 2015
Van Esch BPM, Spierts ILY (2014) Validation of a model to predict fish passage mortality in pumping stations. Can J Fish Aquat Sci 71(12):1910–1923
Vowles AS, Kemp PS (2012) Effects of light on the behaviour of brown trout (Salmo trutta) encountering accelerating flow: application to downstream fish passage. Ecol Eng 47:247–253
Vowles AS, Anderson JJ, Gessel MH, Williams JG, Kemp PS (2014) Effects of avoidance behaviour on downstream fish passage through areas of accelerating flow when light and dark. Anim Behav 92:101–109
Vowles AS, Don AM, Karageorgopoulos P, Worthington TA, Kemp PS (2015) Efficiency of a dual density studded fish pass designed to mitigate for impeded upstream passage of juvenile European eels (Anguilla anguilla) at a model Crump weir. Fish Manag Ecol 22(4):307–316
Webb JA, Wallis EM, Stewardson MJ (2012) A systematic review of published evidence linking wetland plants to water regime components. Aquat Bot 103:1–14
Webb JA, Miller KA, King EL, de Little S, Stewardson MJ, Zimmerman JKH, Poff NL (2013) Squeezing the most out of existing literature: a systematic re-analysis of published evidence on ecological responses to altered flows. Freshw Biol 58:2439–2451
Webb JA, Miller KA, de Little SC, Stewardson MJ, Nichols SJ, Wealands SR (2015) An online database and desktop assessment software to simplify systematic reviews in environmental science. Environ Model Softw 64:72–79
Wilkes MA, Maddock I, Visser F, Acreman MC (2013) Incorporating hydrodynamics into ecohydraulics: the role of turbulence in the swimming performance and habitat selection of stream-dwelling fish. In: Maddock I, Harby A, Kemp P, Wood P (eds) Ecohydraulics: an integrated approach. Wiley, Chichester, pp 7–30
Williams JG, Katopodis C (2016) Incorrect application of data negates some meta-analysis results in Bunt et al. (2012). River Res Appl. doi:10.1002/rra.3076
Winemiller KO, McIntyre PB, Castello L, Fluet-Chouinard E, Girarizzo T, Nam S, Baird IG et al (2016) Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 351(6269):128–129
Zarfl C, Lumsdon AE, Berlekamp J, Tydecks L, Tockner K (2015) A global boom in hydropower dam construction. Aquat Sci 77(1):161–170
Zemlak TS, Habit EM, Walde SJ, Battini MA, Adams ED, Ruzzante DE (2008) Across the southern Andes on fin: glacial refugia, drainage reversals and a secondary contact zone revealed by the phylogeographical signal of Galaxias platei in Patagonia. Mol Ecol 17(23):5049–5061
Acknowledgements
This work was funded by the European Commission through the Marie Sklodowska-Curie action, ‘Knowledge Exchange for Efficient Passage of Fish in the Southern Hemisphere’ (RISE-2015-690857-KEEPFISH). We would like to thank Justin O’Connor and Frank Amstaetter of the Arthur Rylah Institute, Lee Baumgartner of Charles Sturt University, Martin Mallen-Cooper of Fishway Consulting Services, Paul Franklin and Cindy Baker of the National Institute for Water and Atmospheric Research (NIWA) and Oscar Link and Evelyn Habit of the University of Concepción for their valuable inputs in terms of evidence to be included in the review.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wilkes, M.A., Mckenzie, M. & Webb, J.A. Fish passage design for sustainable hydropower in the temperate Southern Hemisphere: an evidence review. Rev Fish Biol Fisheries 28, 117–135 (2018). https://doi.org/10.1007/s11160-017-9496-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-017-9496-8