
Abstract
The significance of temperature-dependent regulation of photosynthetic apparatus (PSA) is determined by the fact that plant temperature changes with environmental temperature. In this work, we present a brief overview of temperature-dependent regulation of photosynthetic processes in class B chloroplasts (thylakoids) and analyze these processes using a computer model that takes into account the key stages of electron and proton transport coupled to ATP synthesis. The rate constants of partial reactions were parametrized on the basis of experimental temperature dependences of partial photosynthetic processes: (1) photosystem II (PSII) turnover and plastoquinone (PQ) reduction, (2) the plastoquinol (PQH2) oxidation by the cytochrome (Cyt) b6f complex, (3) the ATP synthase activity, and (4) the proton leak from the thylakoid lumen. We consider that PQH2 oxidation is the rate-limiting step in the intersystem electron transport. The parametrization of the rate constants of these processes is based on earlier experimental data demonstrating strong correlations between the functional and structural properties of thylakoid membranes that were probed with the lipid-soluble spin labels embedded into the membranes. Within the framework of our model, we could adequately describe a number of experimental temperature dependences of photosynthetic reactions in thylakoids. Computer modeling of electron and proton transport coupled to ATP synthesis supports the notion that PQH2 oxidation by the Cyt b6f complex and proton pumping into the lumen are the basic temperature-dependent processes that determine the overall electron flux from PSII to molecular oxygen and the net ATP synthesis upon variations of temperature. The model describes two branches of the temperature dependence of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) characterized by different activation energies (about 60 and ≤ 3.5 kJ mol−1). The model predicts the bell-like temperature dependence of ATP formation, which arises from the balance of several factors: (1) the thermo-induced acceleration of electron transport through the Cyt b6f complex, (2) deactivation of PSII photochemistry at sufficiently high temperatures, and (3) acceleration of the passive proton outflow from the thylakoid lumen bypassing the ATP synthase complex. The model describes the temperature dependence of experimentally measured parameter P/2e, determined as the ratio between the rates of ATP synthesis and pseudocyclic electron transport (H2O → PSII → PSI → O2).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In plant cells, photosynthesis occurs in chloroplasts, the energy-transducing organelles that assimilate carbon dioxide (CO2) and produce molecular oxygen (O2) using the solar energy absorbed by photosynthetic antennas of photosystem I (PSI) and photosystem II (PSII). The protein–pigment complexes of PSI and PSII are embedded into the thylakoid membranes, which form closed vesicles surrounded by the chloroplast envelope. The piles of stacked flattened thylakoids form grana, which are linked to each other by means of the inter-granal thylakoids. The light energy absorbed by the light-harvesting complexes migrate to photoreaction centers of PSI and PSII (Nelson and Yocum 2006; Mamedov et al. 2015). Operating in tandem, PSI and PSII provide electron transfer from the water molecules, oxidized by the water-oxidizing complex (WOC) of PSII, to NADP+, the terminal electron acceptor reduced by PSI. Two photosystems, PSII and PSI, are interconnected by the cytochrome (Cyt) b6f complex and mobile electron carriers (plastoquinone and plastocyanin, PQ and Pc): PSII → PQ → Cyt b6f → Pc → PSI → NADP+. Photosynthetic electron transport is coupled to generation of the trans-thylakoid difference in electrochemical potentials of protons (\( \Delta \tilde{\mu }_{{{\text{H}}^{ + } }} \), termed as the proton-motive force, pmf), which is a source of energy to drive the H+-ATP synthase: ADP + Pi → ATP (Boyer 1997; Walker 2013). NADPH and ATP, the macroergic products of the light-induced processes of photosynthesis, are used in biosynthetic reactions of the Calvin–Benson cycle (CBC) reactions (the fixation of CO2 into carbohydrates) (Edwards and Walker 1983).
Photosynthetic protein complexes are embedded into the thylakoid membrane. Thylakoids are densely packed with proteins that constitute about 70% of membranes. The physico-chemical properties of the membrane bilayer are determined by the composition and characteristics of the individual lipids. There are four major glycerolipids of chloroplasts membranes: monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG), sulfoquinovosyldiacylglycerol (SQDG), and phosphatidylglycerol (PG). Glycerolipids contain two fatty acids linked to glycerol. MGDG and DGDG are the main building blocks of the thylakoid membrane, which provide the matrix for embedding the photosynthetic complexes into the membrane. The membrane lipids allow lateral diffusion of plastoquinone molecules in the thylakoid membrane. MGDG and SQDG have been found in the Cyt b6f complexes from plants and Chlamydomonas; lipids are involved in maintaining dimeric structure of photosynthetic electron transport complexes (Boudiere et al. 2014; Cramer and Hasan 2016).
The electron transport and ATP synthase complexes are distributed non-uniformly over the membranes of granal and stromal thylakoids (Albertsson 2001; Staehelin 2003; Dekker and Boekema 2005). Stacked thylakoids of grana are enriched with PSII; most of PSI and ATP synthase complexes are localized in the unstacked domains of stroma-exposed thylakoids, grana margins, and grana end membranes. The Cyt b6f complexes are spread uniformly along the thylakoid membranes (Anderson 1982). There are two diffusion-controlled stages of the long-range communication between PSII and PSI: (i) electron transport from PSII to the Cyt b6f complex mediated by PQH2 molecules diffusing in the thylakoid membrane, and (ii) electron transfer from the Cyt b6f complex to PSI mediated by Pc diffusing within the thylakoid lumen. The rate of the intersystem electron transport is determined by PQ turnover as a shuttle connecting PSII and Cyt b6f complexes (Witt 1979; Haehnel 1984; Cardona et al. 2012). The rate of PQ turnover is determined by (i) the reduction of the secondary quinone QB to QBH2, (ii) the dissociation of QBH2 from PSII into the bulk phase of the thylakoid membrane (QBH2 → PQH2), (iii) PQH2 diffusion towards the Cyt b6f complex, and (iv) PQH2 oxidation at the Qo site of the Cyt b6f complex. The light-induced reduction of QB and the appearance of PQH2 in PSII (t1/2 ≈ 0.6–0.8 ms) occur more rapidly than the oxidation of PQH2 by the Cyt b6f complex (t1/2 ≥ 5–20 ms, at room temperatures).
Plastoquinone diffusion in the lipid moiety of the membrane is a central event for the electronic connection between PSII and the Cyt b6f complex. Over a wide range of pH, ionic strength, and temperature, the light-induced reduction of PQ to PQH2, its dissociation from PSII and PQH2 diffusion towards the Cyt b6f complex occur more rapidly than PQH2 oxidation (see Tikhonov 2013, 2014 and references therein). The electron transfer from PSII to the Cyt b6f complex may be retarded due to slow percolation of PQH2 through the lipid domains of the membrane over-crowded with protein complexes (Kirchhoff 2008, 2014). There are experimental reasons to believe, however, that the lateral diffusion of PQH2 within the membrane, as well as Pc movement in the lumen, should not limit the overall rate of electron transfer between PSII and PSI (Haehnel 1976; Tikhonov et al. 1984). Indeed, although significant amounts of PSI and PSII complexes are laterally segregated, most of them are in close contact with the Cyt b6f complexes, which are evenly distributed over the thylakoid membrane (Albertsson 2001). The distribution of Cyt b6f complexes among PSII supercomplexes localized in granal membranes minimizes the average distance traversed by plastoquinone molecules, providing rapid exchange of PQ and PQH2 between the Cyt b6f and PSII complexes (Kirchhoff et al. 2000; Tremmel et al. 2003). Obstructed diffusion of Pc within the narrow lumen may restrict electron communication between the Cyt b6f and PSI complexes (Kirchhoff et al. 2011). However, electron transfer from the Cyt b6f complex to Pc, and further from Pc− to \( {\text{P}}_{700}^{ + } \), occurs more rapidly (t1/2 ~ 5–350 μs, and t1/2 ~ 20–200 μs) as compared to PQ turnover (t1/2 ≥ 4–20 ms) (Haehnel 1984; Sigfridsson 1998). Thus, Pc− diffusion within the lumen should not limit the rate of the intersystem electron transport.
The intersystem electron transport is governed by the light-induced changes in the lumen pH (pHin). There are two main mechanisms of the feedback control of photosynthetic electron transport: (i) the deceleration of PQH2 oxidation by the Cyt b6f complex caused by the lumen acidification (Tikhonov 2014, 2018 and references therein), and (ii) the attenuation of PSII activity due to ΔpH-dependent enhancement of thermal dissipation of absorbed light energy in LHCII known as non-photochemical quenching (NPQ) of chlorophyll (Chl) a excitation (Li et al. 2009; Demmig-Adams et al. 2012; Horton 2012). Thus, the light-induced acidification of the lumen reduces the rate of the intersystem electron transfer from PSII to PSI. There is also the mechanism of “metabolic” control, which means that the rate of electron flow in chloroplasts correlates with the so-called “phosphate potential”, P = [ATP]/([ADP] × [Pi]), where [ATP], [ADP], and [Pi] are the concentrations of ATP, ADP, and Pi (Foyer et al. 2012). Depending on the ADP/ATP ratio, the ATP synthase functions either in the ATP synthesis mode or in the ATPase mode (ATP hydrolysis). In the metabolic “state 4” (the state of “photosynthetic control”, exhausted pools of ADP and/or Pi), when the overall proton flux through the CF0–CF1 complex and ATP production virtually tend to zero, the intersystem electron flow decelerates due to sufficiently strong acidification of the lumen (pHin < 6). In the metabolic “state 3”, the rate of the intersystem electron flow is high, because ATP synthesis is accompanied by stoichiometric drain of protons from the lumen to stroma, thus precluding too strong acidification of the lumen (pHin ≈ 6–6.2; Tikhonov 2013).
Photosynthetic apparatus is sensitive to changes in plant environment, including variations of temperature. The significance of temperature-dependent regulation of photosynthetic apparatus is determined by the fact that plants are poikilothermic organisms, meaning that their own temperature varies with environmental temperature. Thylakoid lipids play an important role in adaptation of chloroplasts to temperature variations. Galactolipids MGDG and DGDG are involved in the maintenance of membrane fluidity of the thylakoid membranes; they contain high amounts of polyunsaturated fatty acids. Adaptation of photosynthetic apparatus to low (or high) temperatures can proceed due to an increase (or a decrease) in the desaturation degree of fatty acids in galactolipids (Wallis and Browse 2002; Zhou et al. 2016). The response of photosynthetic apparatus to variations of temperature reveals itself as an interplay of a number of different partial photosynthetic processes. In the literature one can find varied information on temperature dependence of partial energy-transducing reactions in chloroplasts such as electron transfer and proton translocation (Kraayenhof et al. 1971; Shneyour et al. 1973; Nolan and Smillie 1976, 1977; Nolan 1980, 1981; Schuurmans and Kraayenhof 1983) and structural transitions in thylakoid membranes detected by the fluorescent and/or paramagnetic probes (Torres-Pereira et al. 1974; Yamamoto and Nishimura 1976; Murata and Fork 1977; Ford et al. 1982; Tikhonov and Subczynski 2005). It is remarkable that most of the Arrhenius plots for partial reactions of photosynthesis show inflexions (or even discontinuities) (Kumamoto et al. 1971; Inout 1978). There are, however, some inconsistent results on the estimation of the apparent activation energies formally determined for partial reactions near the transition temperatures. This diversity may be accounted for by using of various plant species and different experimental conditions.
Thermo-induced structural changes in thylakoid membranes belong to basic factors that determine the chloroplast response to fluctuations of temperature (for references, see Hirano et al. 1981; Barber et al. 1984; Los and Murata 2004; Tikhonov and Subczynski 2005; Allakhverdiev et al. 2008; Los et al. 2013; Yamori et al. 2014; Yamamoto 2016; Maksimov et al. 2017; Nievola et al. 2017; Hu et al. 2020). One of the mechanisms for supporting a sufficiently high activity of photosynthetic apparatus upon variations of temperature is associated with the reorganization of membrane structures, including changes in the physical state (fluidity) of the thylakoid membrane (Quinn and Williams 1978; Yamamoto et al. 1981; Mizusawa and Wada 2012; Yamori et al. 2014; Niu and Xiang 2018). Temperature-induced re-modeling of photosynthetic lipid–protein structures can affect the rates of electron and proton transport processes coupled to ATP synthesis, thereby providing optimal fitting of photosynthetic apparatus to environmental temperature. There are good reasons to believe that the acclimation of plants to environmental temperature is realized by changes in the composition of membrane lipids (saturated/desaturated lipids) that determine the local viscosity of the lipid domains (Heise and Harnischfeger 1978; Kern et al. 2009; Tietz et al. 2015; Maksimov et al. 2017). The fluidity of membrane lipids plays an important role in controlling photosynthetic processes. Variations of temperature directly alter the physical state of thylakoid membranes. Changes in the composition of lipids (in particular, the relative content of unsaturated fatty acids) manifest themselves in temperature dependences of physical parameters of biological and model membranes (for references, see Berliner 1976; Griffith and Jost 1976; McConnell 1976; Lee 1977; Margolis et al. 1980; Luzikov et al. 1983, 1984; Aloia and Boggs 1985; Lutova and Tikhonov 1988). Lipids with unsaturated fatty acids have lower “melting” temperature than lipids containing saturated fatty acids; the proportionality between these lipids is one of the key factors that determine the membrane fluidity and lipid diffusion in the thylakoid membranes (Sarcina et al. 2003; Tietz et al. 2015). A decrease in the fluidity of lipid domains of thylakoid membranes caused, for example, by the implementation of cholesterol, is accompanied by an inhibition of the intersystem electron transfer (Yamamoto et al. 1981; Ford and Barber 1983; Barber et al. 1984). The ratio of saturated and unsaturated fatty acids depends on the plant growth conditions (Sawada and Miyachi 1974). Fluidity of membrane lipids is often considered as a peculiar sensor that triggers the retrograde signals controlling the expression of desaturases, adjusting the thylakoid membrane to environmental temperature and, thereby, optimizing the energy transduction in photosynthetic organisms (Los and Murata 2004; Los et al. 2013).
In our previous works (Tikhonov et al. 1980, 1981, 1983, 1984; Timoshin et al. 1984; Tikhonov and Subczynski 2005), we used the electron paramagnetic resonance (EPR) technique for scrutinizing the structure–function relationships in class B chloroplasts (thylakoids) isolated from bean leaves. The advantage of the EPR method is that it allows measuring the functional (electron transport) and structural characteristics of thylakoid membranes under the same experimental conditions. Class B chloroplasts present a suitable model for analyzing the regulation of photosynthetic processes, because they are deprived of the outer shell and the CBC enzymes. This is because the processes beyond the thylakoids will not interfere with the membrane-dependent reactions of electron transport. In the meantime, the integrity of closed thylakoids enables them to generate \( \Delta \tilde{\mu }_{{{\text{H}}^{ + } }} \) and to support the operation of the ATP synthase. Investigating the structure–function relationships in bean thylakoids, we have found strong correlations between the temperature dependences of the intersystem electron transport and ATP synthesis, on the one hand, and structural changes in the lipid domains of thylakoid membranes, on the other hand (the results of these studies are briefly summarized in Tikhonov and Subczynski (2005) and Tikhonov (2020)). The lipid-soluble nitroxide radicals (spin probes) were used for probing the structural transitions in the lipid domains of thylakoid membranes. These changes manifest themselves as the inflexions (or breaks) in the plots of spectral parameters. The EPR spectra of spin probes depend on their local surroundings and ordering of nitroxide radicals in the membrane moiety. It is important to note, however, that spin probes localized at different depths from the membrane surface indicate on the cooperative character of thermo-induced structural transients in the lipid domains of thylakoid membranes (Lee 1977; Tikhonov and Subczynski 2005).
The current work was inspired by the necessity of numerical simulation of temperature-dependent photosynthetic processes in chloroplasts. The importance of computer modeling of temperature-dependent photosynthetic processes is determined by the complexity and variability of electron and proton transport events in chloroplasts (for reviews, see Kukushkin and Tikhonov 1988; Karavaev and Kukushkin 1993; Laisk et al. 2009; Lazár and Schansker 2009; Riznichenko et al. 2009; Arnold and Nikoloski 2011; Igamberdiev 2011; Zaks et al. 2012; Zhu et al. 2013; Rubin and Riznichenko 2014; Tikhonov and Vershubskii 2014; Tikhonov 2016; Stirbet and Govindjee 2016; Cherepanov et al. 2017; Morales et al. 2018; Stirbet et al. 2014, 2019). The responses of photosynthetic apparatus to temperature manifest itself as the interplay of different processes; temperature-dependent regulation of photosynthesis is achieved by cooperation of several feedbacks. Computer modeling of photosynthetic processes would gain a better insight into understanding the temperature-dependent regulation of photosynthesis, which is important from both fundamental and applied viewpoints. The main purpose of this study is the computer analysis of relationships between electron transport, proton translocation, and ATP synthesis processes in thylakoids. Our model mimics the influence of the membrane physical state on the key steps of electron transport in thylakoids. Below, describing the model, we briefly overview the relationships between the photosynthetic processes (electron transport, proton translocation, and ATP synthesis) and structural transitions in the lipid domains of bean thylakoids. Results of our calculations strongly support the notion that the structural changes in the lipid domains and protein complexes, which control the photochemical activity of PSII, the rate of PQH2 oxidation by the Cyt b6f complex and the trans-thylakoid proton transfer through the ATP synthase, are among the crucial factors of the temperature-dependent regulation of electron transport and ATP synthesis in chloroplasts.
Description of the model
General properties of the model
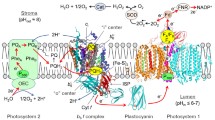
Figure 1 (the bottom panel a) depicts the layout of electron carriers functioning in the chain of electron transport from H2O to O2 through the membrane-bound protein complexes (PSII, b6f, and PSI) and mobile electron carriers, plastoquinone (PQ). The model describes the key stages of electron transfer from the water-oxidizing complex (WOC) of PSII to molecular oxygen, the terminal electron acceptors of PSI in class B chloroplasts (H2O → PSII → PQ → b6f → Pc → PSI → O2). Mobile electron carriers, plastoquinone (PQ) and plastocyanin (Pc), mediate electron transfer between the PSII, Cyt b6f, and PSI complexes. Reduced plastoquinol molecules (PQH2) connect PSII with the Cyt b6f complex. PQH2 oxidation by the Cyt b6f complex is considered as the rate-limiting step in the intersystem chain of electron transport. We take into account that the rate of PQH2 oxidation is controlled by the intra-thylakoid pH (pHin), because the PQH2 oxidation is coupled to dissociation of two protons into the thylakoid lumen (PQH2 → PQ + 2\( {\text{H}}_{\text{in}}^{ + } \) + 2e−). Pc molecules, reduced by the Cyt b6f complexes, rapidly moving within the lumen, provide the reduction of \( {\text{P}}_{ 7 0 0}^{ + } \). As noted above, the PQH2 formation in PSII and its diffusion to the Cyt b6f complex usually occur more rapidly (τ1/2 < 1–5 ms) than electron transfer from PQH2 to \( {\text{P}}_{ 7 0 0}^{ + } \) (τ1/2 > 5–20 ms) via the Cyt b6f complex and Pc (for references, see Siggel 1976; Sanderson et al. 1986; Sigfridsson 1998; Hope 2000; Santabarbara et al. 2009; Tikhonov 2013, 2014, 2018). This means that the rate of \( {\text{P}}_{ 7 0 0}^{ + } \) reduction by electrons injected to the intersystem ETC from PSII should be determined predominantly by the rate of electron transfer from the PQH2 pool to the Cyt b6f complex. On the acceptor side of PSI, reduced ferredoxin molecules bound to PSI (FA and/or FB) donate electrons to O2. Molecular oxygen which serves as the terminal electron acceptor in the chain of pseudocyclic ETC (the “water-water” cycle; Asada 1999; Ort and Baker 2002; Cherepanov et al. 2017), since type B chloroplasts have no envelope, and thus neither FNR nor the CBC enzymes.

A scheme of the photosynthetic electron and proton transport processes considered in the model and the arrangement of the four main protein complexes (photosystem I, photosystem II, cytochrome b6f, and ATP synthase) in the thylakoid membrane (panel a). Blue arrows show electron transfer reactions, and red arrows depict proton transport pathways. The top panels b, c, and d illustrate the impact of temperature on the partial reactions in the chain of linear electron transport. Panel b shows the temperature dependences of the model parameters \( \xi (T ) \) and \( \kappa_{\text{PSII}} (T ) = f\left( T \right)/f_{\text{o}} \), which determine the operation of PSII (based on data presented in Tikhonov et al. 1983; see text for explanations). Parameter \( \xi (T ) \) characterizes the photochemical activity of PSII in response to a short light flash (7 μs) inducing single turnover of PSII. Parameter \( \kappa_{\text{PSII}}^{{}} (T ) \) characterizes acceleration of electron transfer from P680 to the PQ pool (for details, see text and Fig. 13 in “Appendix 3”). Panel c shows the temperature dependence of parameter \( \tau_{1/2}^{ - 1} \), where τ1/2 is the half-time of the post-illumination reduction of \( {\text{P}}_{ 7 0 0}^{ + } \) (based on data presented in Tikhonov et al. 1984). Panel d shows the temperature dependence of O2 solubility in water solutions (based on data presented in Melnichenko et al. 2008)
Electron transport processes are accompanied by translocation of protons into the thylakoid lumen (\( {\text{H}}_{\text{out}}^{ + } \) → \( {\text{H}}_{\text{in}}^{ + } \)) and generation of the trans-thylakoid pH difference (ΔpH = pHout − pHin). The acidification of the lumen (pHin < pHout) occurs due to functioning of WOC in PSII and the PQ shuttle (PQ + 2e− + 2\( {\text{H}}_{\text{out}}^{ + } \) → PQH2 → PQ + 2e− + 2\( {\text{H}}_{\text{in}}^{ + } \)). The proton leakage from the lumen occurs by two ways: (a) the proton flux through the ATP synthase (JATP), coupled to ATP formation from ADP and inorganic phosphate Pi, and (b) the passive proton flux through the membrane (Jpass). We assume that the pH value of the suspension is constant (pHout = 8), owing to sufficiently high buffer capacity of the external medium.
The following variables of the model are considered: the relative concentrations of oxidized primary electron donors in PSI and PSII ([\( {\text{P}}_{700}^{ + } \)] and [\( {\text{P}}_{680}^{ + } \)]); the relative concentration of oxidized plastoquinone, [PQ]; the relative concentration of oxidized ferredoxin bound to PSI, [Fd]; the relative concentration of oxidized plastocyanin, [Pc]. The proton transport is described by the variable [\( {\text{H}}_{\text{in}}^{ + } \)], the concentration of hydrogen ions within the lumen. The light-induced changes in ATP concentration, described by the variable [ATP], is determined by the balance between the ATP synthesis and the ATP hydrolysis processes as described earlier (Tikhonov and Vershubskii 2014; Vershubskii et al. 2017). The model parameters L1 and L2 characterize the photosynthetically active fluxes of light quanta exciting PSI and PSII, respectively. The ratio L1/L2 = 10 was accepted for mimicking the far-red light (“Light 1”) exciting preferentially PSI. The ratio L1/L2 = 1 was used for modeling illumination of chloroplasts by light efficiently exciting both PSI and PSII (“Light 2”).
A system of non-linear ordinary differential equations (ODE) was used to describe the dynamics of the model system (for details, see Tikhonov and Vershubskii 2014; Vershubskii et al. 2017, 2018). The set of ODE is presented in “Appendix 1”. The appropriate choice of the apparent rate constants of partial reactions of electron and proton transport was described in our previous works (Vershubskii et al. 2011; see also Table 1 in “Appendix 1”). We have analyzed chloroplast functioning in three metabolic states: state 3 refers to the quasi-steady-state of chloroplasts during active ATP synthesis in the presence of the surplus amounts of ADP and Pi, state 4 corresponds to the state of “photosynthetic control” (no total synthesis of ATP), state 5 pertains to the situation when ΔpH = 0 (uncoupled thylakoids).
Effects of temperature on the partial reactions of electron transport in thylakoids
The top panels in Fig. 1 illustrate how variations of temperature influence the photochemical activity of PSII (panel b), the rate of electron transfer from PQH2 to \( {\text{P}}_{ 7 0 0}^{ + } \) (via the Cyt b6f complex and Pc; panel c), and the solubility of O2 in water solutions (panel d). We analyze electron transport processes in the temperatures range from 0 to 45 °C, because the PSII activity is known to be completely inhibited at temperatures ≥ 45 °C (see, e.g., Lutova and Tikhonov 1983; Benkov et al. 2019). Parametrization of temperature dependences of the partial reactions of electron transport is based on experimental data borrowed from our previous works on class B bean chloroplasts (Tikhonov et al. 1980, 1983, 1984; Timoshin et al. 1984). Below we briefly consider the peculiarities of temperature-dependent partial reactions of electron and proton transport considered in this work. Plant materials and some principal details of experimental methods used in the above-cited works are presented in “Appendix 3”.
Photosystem II
For parametrization of PSII activity, we used the temperature dependences of the relative numbers of electrons injected into the intersystem ETC in response to light flashes of various duration (Tikhonov et al. 1980; Tikhonov and Vershubskii 2017; for details, see “Appendix 3”). The model parameters \( \xi (T ) \) and \( \kappa_{\text{PSII}}^{{}} (T ) \) characterize the temperature dependence of PSII activity. Parametrization of \( \xi (T ) \) was performed on the basis of the kinetics of P700 redox transients induced by a short pulse (t1/2 = 7 μs) of white light of saturating intensity, which provided a single turnover of PSII (Stiehl and Witt 1969; Witt 1979). Parameter \( \xi (T ) \) decreases at temperatures ≥ 25 °C (Fig. 1b), tending to zero at 45 °C (complete inhibition of PSII activity).
In response to a long flash (t1/2 = 750 μs), each PSII donated several electrons to the PQ pool. We assume that the apparent rate constant of electron transfer from PSII to PQ (the rate constant \( {k}_{{{\text{P}}_{ 6 8 0} }} \), Eq. A4 in “Appendix 1”) to be proportional to the ratio f = W2(T)/W1(T), where W2(T) and W1(T) denote the relative numbers of electrons donated into the intersystem ETC in response to the long (τ1/2 = 750 μs) and short (τ1/2 = 7 μs) flashes, respectively (for definition of W2 and W1, see Fig. 13 in “Appendix 3”). Figure 1b shows the temperature dependence of the ratio f(T) = W2/W1 normalized to f0 = f(25 °C). Parameter \( \kappa_{\text{PSII}} (T) = f\left( T \right)/f \) (25 °C) increases with the rise of temperature. This means that the apparent rate constant of electron transfer from PSII to the PQ pool increases with temperature. A clear inflexion of \( \kappa_{\text{PSII}}^{{}} (T ) \) at 25 °C is likely to reflect structural changes in the membrane that have impact on PSII activity. In order to simulate the influence of the membrane physical state on the rate of electron transfer from PSII to the PQ pool, we performed calculations for different patterns of \( \kappa_{\text{PSII}}^{{}} (T ) \), characterized by different values of the model parameter to (Fig. 2a).

Temperature dependences of the partial reactions of photosynthesis related to different values of the model parameters t0. The value of parameters t0 reflects the temperature of the membrane structural transition (for more explanations, see the main text): t0 = 20 °C (labeled by one asterisk), t0 = 25 °C (the basic model), and t0 = 30 °C (labeled by two asterisks). Panel a: Normalized temperature dependence of the correction factor \( \kappa_{\text{PSII}}^{{}} (T )= f\left( T \right)/f\left( {t_{0} } \right) \), where the ratio f(T) = W2(T)/W1(T) characterizes the acceleration of PSII turnover with temperature (for definition of W2(T) and W1(T), see Fig. 13 in “Appendix 3”). Panel b: Normalized temperature dependence of the correction factor \( \kappa_{\text{Q}}^{{}} (T ) \) characterizing the rate of electron transfer from PQH2 to Pc via the Cyt b6f complex (see text for explanations). Panel c: Normalized temperature dependence of the correction factor \( \kappa_{\text{ATP}}^{{}} (T ) \) characterizing the effect of temperature on the activity of the ATP synthase complex. Panel d: Normalized temperature dependence of the correction factor \( \kappa_{\text{pass}}^{{}} (T ) \) characterizing the passive H+ ion transfer through the thylakoid membrane
PQH 2 oxidation
Modeling the chain of linear electron transfer (H2O → PSII → PQ → b6f → Pc → PSI → Fd → O2), we assume that the reaction of PQH2 oxidation is the rate-limiting step in the intersystem ETC (Stiehl and Witt 1969). In the whole range of temperatures considered in our work (0–45 °C), the rate of PQH2 oxidation is significantly slower than the Pc turnover between the Cyt b6f complex and PSI (see, e.g., Tikhonov et al. 1984, 2018 and references therein). Rapid shuttling of electrons between the PSII and Cyt b6f complexes is determined by a high mobility of PQH2 and PQ molecules of the photo-reducible plastoquinone pool in the thylakoid membrane. Close location of PSII and granal Cyt b6f complexes and sufficiently high fluidity of membrane lipids promote rapid formation of the “substrate–enzyme” complex (PQH2-b6f). The overall rate of the intersystem electron transfer should be determined mainly by the rate of PQH2 oxidation at the quinol-binding portal Qo of the Cyt b6f complex (Tikhonov 2014, 2016, 2018). The rate of this process is controlled by the intra-thylakoid pH (pHin). Oxidation of PQH2 is accompanied by the release of two protons into the lumen. The back-pressure of the protons pumped into the lumen decelerates the oxidation of PQH2, thereby slowing down the intersystem electron transport with the lumen acidification (pHin ↓).
Within the framework of our model, we consider the influence of pHin on the rate of PQH2 oxidation, using the function \( k_{\text{Q}}^{0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) = 1/\tau_{\text{Q}} \), which was first suggested by Dubinskii and Tikhonov (1997). We used this function in our works (Vershubskii et al. 2011, 2018; Tikhonov and Vershubskii 2014, 2017), assuming that the rate of PQH2 oxidation can be found as the reciprocal value of the overall time \( \tau_{\text{Q}} \) of PQH2 turnover related to electron flow from PQH2 to Pc via the Cyt b6f complex:
The \( \tau_{\text{Q}} \) value is the sum of characteristic times related to the following steps of PQH2 oxidation by the Cyt b6f complex and further electron transfer to Pc: (1) the PQH2 binding to quinol-binding center “o”, (2) the two-electron oxidation of PQH2, and (3) the electron transfer from the Cyt b6f complex to plastocyanin (Pc). Here, \( [ {\text{PQH}}_{ 2} ]= [ {\text{PQ]}}_{ 0} - [ {\text{PQ]}} \) is the concentration of PQH2, ko is the binding constant of PQH2 to the Qo center; \( \tau_{1}^{\text{o}} \),\( \tau_{2}^{\text{o}} \), h1, and h2 are the model parameters characterizing two steps of PQH2 oxidation inside the Cyt b6f complex; kf is the rate constant of electron transfer from the Cyt b6f complex to the oxidized Pc molecule. Parameters h1 and h2 are the normalizing coefficients, the magnitudes of which are determined by the pKa values of the first and second stages of PQH2 deprotonation. The deprotonation of PQH2, associated with the proton release into the lumen, to be considered as the prerequisite for PQH2 oxidation (Brandt 1996; Link 1997; Crofts et al. 2000, 2013). Therefore, the rate of PQH2 oxidation appears to be dependent on the concentration of hydrogen ions (\( [ {\text{H}}_{\text{in}}^{ + } ] \)) inside the thylakoids (for references, see Tikhonov 2013, 2014, 2018). In this work, the choice of the coefficients related to parametrization of \( k_{\text{Q}}^{0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) \) was performed as described earlier (Vershubskii et al. 2011, 2018; Tikhonov and Vershubskii 2014; see also “Appendix 1”). A good agreement between the experimental and model pHin dependences of the kinetic parameter τ1/2, which characterizes the half-time of the post-illumination reduction of \( {\text{P}}_{ 7 0 0}^{ + } \), proves an adequacy of \( k_{\text{Q}}^{0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) \) parametrization (see Fig. 13 in “Appendix 3”).
The oxidation/reduction reactions of plastoquinone are the temperature-dependent processes. In the current work, we used the modified function \( k_{\text{Q}}^{0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) \) to describe the temperature dependence of the intersystem electron transfer. According to experimental data for bean chloroplasts (for review, see Tikhonov 2018, 2020 and reference therein), the temperature dependence of the rate of electron transfer from PQH2 to \( {\text{P}}_{ 7 0 0}^{ + } \) can be approximated by two exponents (Fig. 1c). Taking that at the characteristic temperature t0 the rate of PQH2 oxidation can be described by the function \( k_{\text{Q}}^{ 0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) \), we determined the function \( k_{\text{Q}}^{{}} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ] ,T} \right) \) using the following formula:
where \( \kappa_{\text{Q}}^{{}} (T )= { \exp }\left[ { - E_{\text{a}} /\left( {k_{\text{B}} T} \right) + E_{\text{a}} /\left( {k_{\text{B}} T_{0} } \right)} \right] \) is the temperature-dependent correction factor. Here, Ea is the activation energy, kB is the Boltzmann constant, T denotes the temperature in the Kelvin scale, T0 = 273.16 + t0, where t0 is the peculiar temperature in the Celsius scale. The function \( k_{\text{Q}}^{ 0} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ]} \right) \) describes the overall rate of PQ turnover at the characteristic temperature t0, which corresponds to the break-point in the temperature dependence of the PQH2 oxidation rate determined from the post-illumination reduction of \( {\text{P}}_{ 7 0 0}^{ + } \). According to experimental data on bean chloroplasts presented in Tikhonov et al. (1984), the Arrhenius plot of the overall rate of electron transfer from PQH2 to \( {\text{P}}_{ 7 0 0}^{ + } \) can be approximated by two exponents with activation energies \( E_{\text{a}}^{ ( 1 )} \) and \( E_{\text{a}}^{ ( 2 )} \). Experimental Arrhenius plot of the rate of the post-illumination reduction of \( {\text{P}}_{ 7 0 0}^{ + } \) (Fig. 1c) shows an explicit break at t0 ≈ 25 °C and characterized by \( E_{\text{a}}^{ ( 1 )} \) ≈ 60 kJ/mol and \( E_{\text{a}}^{ ( 2 )} \) ≤ 3.5 kJ/mol. There are good reasons to believe that the two branches in the temperature-dependence plot reflect structural transitions in the lipid domains of the thylakoid membrane. Thus, bearing in mind the structure–function relationships in thylakoid membranes, we can say that the two-branch pattern of the temperature-dependent correction factor \( \kappa_{\text{Q}}^{{}} (T ) \), and, respectively, the function \( k_{\text{Q}}^{{}} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ] ,T} \right) \), mimic the temperature dependence of PQH2 oxidation at different fluidities of the membrane (see Tikhonov 2020 and references therein). In this work, we compared the behavior of the model system for three different patterns of the temperature-dependent correction factor \( \kappa_{\text{Q}}^{{}} (T ) \) (Fig. 2b).
Photosystem I
Electron transfer on the acceptor side of PSI occurs rapidly by the mechanism of quantum mechanical tunneling (Moser et al. 1992; Brettel 1997; Page et al. 1999; Möbius and Savitsky 2009; Shelaev et al. 2010). The outflow of electrons from PSI to O2 (the Mehler reaction; see Asada 1999; Badger et al. 2000; Ort and Baker 2002; Cherepanov et al. 2017) was calculated as JFd–O2 = kMeh·(1 – [Fd])·[O2]. Here, kMeh stands for the apparent rate constant of electron transfer from the reduced terminal electron carriers on the acceptor side of PSI (FA and FB), collectively denoted as Fd, and [O2] is the concentration of molecular oxygen in the chloroplast suspension. As a result of electron transfer from PSI to O2, superoxide radicals \( {\text{O}}_{2}^{ \bullet - } \) are formed. Two \( {\text{O}}_{2}^{ \bullet - } \) molecules dismutate to form hydrogen peroxide (H2O2) and molecular oxygen O2 (2\( {\text{O}}_{2}^{ \bullet - } \)+ 2H+ → H2O2 + O2), and two H2O2 molecules decompose to O2 and water. Thus, electrons from the water molecules oxidized in PSII are transferred to O2 (O2 + e− → \( {\text{O}}_{2}^{ \bullet - } \)); the water molecule formed in the result of further transformations of \( {\text{O}}_{2}^{ \bullet - } \)is the final product of PSI (H2O → PSII → PSI → H2O, the “water–water” cycle; Asada 1999).
It is common knowledge that PSI is less sensitive to injuries at higher temperatures compared to PSII. Variations of temperature within the range 0–45 °C may influence PSI activity, but much less significantly than the intersystem electron transport controlled by PQ reduction by PSII and PQH2 oxidation by the Cyt b6f (see, e.g., Yan et al. 2013). Nevertheless, the terminal stage of electron transfer (the reduction of O2 by Fd−) depends on temperature, because the solubility of O2 in water changes with variations of temperature (Benson and Krause 1984; Melnichenko et al. 2008; Clever et al. 2014). Therefore, to take into account the influence of temperature on the outflow of electrons from PSI to O2, we consider here the temperature dependence of O2 solubility in water (Fig. 1d).
In this work, we do not consider the cyclic flow of electrons around PSI, in which electrons return from Fd− via the water-soluble ferredoxin and ferredoxin-quinone reductase (FQR) to the ETC segment between PSII and PSI (at the PQ level). This is because Class B chloroplasts lose the water-soluble ferredoxin, a mediator of the cyclic electron transport around PSI (Bendall and Manasse 1995; Strand et al. 2016).
Proton transport and ATP synthesis
We consider that protons accumulate inside the thylakoid lumen due to water oxidation by WOC and PQH2 oxidation by the Cyt b6f complex. The overall balance of the electron and proton transport in PSII is the following: two protons evolve in the lumen per one H2O molecule decomposed in PSII; two electrons extracted from one H2O molecule are used to reduce PQ to PQH2. Oxidation of one PQH2 molecule by the Cyt b6f complex is accompanied by the release of two protons into the lumen. Note that the stoichiometric ratio PQH2/2H+ = 1 is true if the Q-cycle in the Cyt b6f complex is neglected. We do not consider the operation of the Q-cycle, because of the absence of soluble ferredoxin in class B chloroplasts. Otherwise, two protons are released into the lumen per one electron transferred from PQH2 to PSI (Mitchell 1976; Baniulis et al. 2008; Cramer and Hasan 2016; Tikhonov 2018). The overall electron and proton balance is the following: H2O + PQ + 2\( {\text{H}}_{\text{out}}^{ + } \) → 1/2O2 + PQH2 + 2\( {\text{H}}_{\text{in}}^{ + } \). We also take into account that hydrogen ions translocated into the lumen can bind to the proton-accepting (buffer) groups, the concentrations of which significantly (by two orders of magnitude) exceed the concentration of electron carriers (for details, see Tikhonov and Blumenfeld 1985; Tikhonov and Vershubskii 2014, 2017).
The efflux of protons from the lumen is considered to occur through the ATP synthase (the proton flux JATP) and due to the passive leak of protons through the thylakoid membrane (the proton flux Jpass).
The proton flux coupled to ATP synthesis (J ATP )
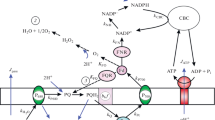
The ATP synthase is a reversible molecular machine operating in both directions, either to form ATP from ADP and Pi (the endergonic process) or to hydrolyze ATP (the exergonic reaction). In Fig. 3, panel a schematically depicts the architecture of the ATP synthase ensemble: the membrane fragment of the ATP synthase complex (CF0) is surrounded by the membrane lipids, the CF1 fragment is exposed to stroma. Similarly to the model described earlier (Tikhonov and Vershubskii 2014; Vershubskii et al. 2017), we consider the proton flow as a two-step process: (1) the proton binding to the membrane-buried carboxy group (–COO− + \( {\text{H}}_{\text{in}}^{ + } \) → –COOH) and (2) the proton dissociation from the protonated group –COOH (–COOH → –COO− + \( {\text{H}}_{\text{out}}^{ + } \)) (Fig. 3b). The proton transfer through the ATP synthase proceeds via the carboxy groups located in the center moiety of the subunits c, which are assembled as the cn-ring buried into the membrane (Fig. 3a). The transmembrane pH difference (ΔpH = pHout − pHin) provides the protonation/deprotonation reactions shown in Fig. 3b, thereby supporting directed transfer of protons \( \left( {{\text{H}}_{\text{in}}^{ + } \to {\text{H}}_{\text{out}}^{ + } } \right) \).

Schemes illustrating the architecture of the ATP synthase ensemble (panel a) and the two-step mechanism modeling the transmembrane proton transfer (panel b). The transmembrane proton transfer, either through the ATP synthase (JATP) or passive flux (Jpass), occurs by means of the protonation/deprotonation exchange with the membrane-buried carboxy group. Panel c presents the force–flux relationship (JATP vs. ΔpH) calculated for the model parameter pKc = 7.3 (see “Appendix 2” for details of JATP calculations)
The ΔpH-driven proton transfer through the ATP synthase is coupled to directed rotation of the cn-ring within the membrane (Junge et al. 1997; Fillingame et al. 2000; Diez et al. 2004; Ariga et al. 2007; Romanovsky and Tikhonov 2010; Junge and Nelson 2015). Rotating due to the energy of the trans-thylakoid pH difference, the cn-ring actuates the operation of the coupling factor CF1, catalyzing the ATP formation (ADP + Pi → ATP). Figure 3c presents the typical pattern of the “flux–force” relationship (JATP vs ΔpH) related to the proton translocation through CF0 at different values of the proton-motive force ΔpH. JATP is calculated for pHout = 8 and pKa = 7.3, where pKa characterizes proton-accepting properties of Glu71 in subunits c of the cn-ring (Vollmar et al. 2009). The equation for the flux JATP is presented in “Appendix 2” (Eq. A8). The sigmoid-type dependence of JATP vs ΔpH demonstrates that efficient flux JATP occurs after the generation of a sufficiently high ΔpH difference (> 1.5–2.0). We do not consider here the partitioning of pmf into ΔpH and Δψ, because a steady-state difference in electric potentials Δψ =ψin − ψout in chloroplasts is negligible under the (quasi)steady-state conditions (Johnson and Ruban 2014; Davis et al. 2017).
One complete rotation of the cn-ring results in the formation of three ATP molecules (Junge et al. 1997; Seelert et al. 2000; Junge and Nelson 2015). Bearing in mind that the rotation of the membrane-buried cn-ring may depend on the fluidity of the membrane lipids surrounding cn, we assume that the rate of ATP synthesis is controlled by the membrane fluidity. There are good reasons to believe that the acceleration of the cn-ring rotation with temperature will stimulate the stoichiometric formation of ATP coupled to directed revolutions of the rotor. Therefore, we consider that the apparent rate constant \( k_{\text{ATP}}^{{}} (T ) \), which stands in the equation describing ATP synthesis (“Appendix 1”, Eq. A6), increases with temperature. We assume that the temperature dependence of \( k_{\text{ATP}}^{{}} (T )= \kappa_{\text{ATP}} (T )\cdot k_{\text{ATP}}^{\text{o}} \), where \( k_{\text{ATP}}^{\text{o}} \) is the model parameter related to t0 = 25 °C, and \( \kappa_{\text{ATP}}^{{}} (T ) \) is the temperature-dependent correction factor described by two exponents (Fig. 2c). The change-over of the apparent activation energy, corresponding to the inflexion in the Arrhenius plot of \( k_{\text{ATP}}^{{}} (T ) \) at a characteristic temperature T0, may be tentatively attributed to the structural changes in the membrane caused by the fluidization of the lipid domains surrounding the ATP synthase. Since the ATP synthase is the reversible enzyme capable of hydrolyzing ATP, we consider that the apparent rate constant of ATP hydrolysis is also the temperature-dependent process (see the temperature dependence of ATP hydrolysis in bean chloroplasts, “Appendix 3”, Fig. 15). Within the framework of our model, we assume that the ratio of the rates of the forward (ATP synthesis) and reverse (ATP hydrolysis) reactions may be taken as kATP/kADP = 0.1. This ratio was derived from the comparison of temperature dependences of the light-induced ATP synthesis and ATP hydrolysis in the dark in isolated bean chloroplasts (Timoshin et al. 1984; see also “Appendix 3” for the description of the measurements of ATP synthesis and ATP hydrolysis).
The passive flux of protons through the membrane (Jpass)
Along with the active proton transport through the ATP synthase (JATP), the passive outflow of protons from the lumen (Jpass) will contribute to the establishment of ΔpH. For a proper choice of the function Jpass(ΔpH), we turned to experimental data on the H+ ion uptake by thylakoids. Figure 4a shows the kinetics of light-induced acidification (pHout ↑) of weakly buffered suspension of chloroplasts in metabolic state 4 (Tikhonov et al. 1983). Figure 4b depicts the temperature dependence of the proton uptake (ΔH+), demonstrating that ΔH+ increases with the rise of temperature in the range 0–25 °C, but monotonously decreases at higher temperatures. The bell-like temperature dependence of ΔH+ can be explained by the interplay of two effects: (i) the enhancement of proton pumping into the lumen due to the speeding-up of electron transport with temperature, and (ii) the acceleration of proton leak from the lumen caused by the membrane fluidization at sufficiently high temperatures.

Passive proton transfer through the thylakoid membrane. Panel a depicts the kinetics of light-induced pH changes in the weakly buffered suspension of chloroplasts in metabolic state 4. Panel b shows the temperature dependence of the light-induced uptake of proton (parameter ΔH+). In panel c, we compare the normalized temperature dependences of semilogarithmic plots of the proton flux Jpass obtained in experiment (stars) and theory (circles), on the one hand, the structural parameter 2T ′II derived from the EPR spectrum of the lipid-soluble spin-probe 5-SASL (see text for more explanations). Experimental data are from Tikhonov et al. (1983)
The post-illumination decay of pHout reflects the proton leak from the lumen. In analogy to the model of the proton flow through the ATP synthase (Fig. 3b), we consider that the passive flux of protons through the membrane (Jpass) occurs by means of the proton exchange with the membrane-bound acidic groups (Vershubskii et al. 2011). For correct choice of the model parameters that determine Jpass, we compared the calculated values of Jpass with the appropriate experimental data obtained for isolated bean chloroplasts. As a touchstone for fitting the model parameters, we used the experimental curve measured in state 4 (without the addition of ADP). In this case, we could exclude the overestimation of Jpass that might occur due to the ATP synthase activity. Figure 4c shows the normalized semilogarithmic plots of the proton flux Jpass vs. temperature borrowed from experimental (stars) and calculated (circles) data. Experimental values were determined as Jpass ~ 1/τ1/2, where τ1/2 is the half-time of the post-illumination decay of pHout in state 4 (for definition, see Fig. 4a). Note that the experimental dependence demonstrates the characteristic inflexion of Jpass at t0 ≈ 25 °C. In accordance with experimental data, the calculated rate of the proton leakage (the passive flux Jpass) notably increased with the rise of temperature up to 25 °C. At temperatures higher than t0, the rate of the proton leakage also increased with temperature, but less significantly. Theoretical values of Jpass were determined as \( J_{\text{pass}} = k_{{{\text{H}}^{ + } }} (T )\cdot ( [ {\text{H}}_{\text{in}}^{ + } ]- [ {\text{H}}_{\text{out}}^{ + } ] ) \), where the temperature-dependent coefficient \( k_{{{\text{H}}^{ + } }} (T ) \) determines the transmembrane proton flux driven by the “proton force”, \( [ {\text{H}}_{\text{in}}^{ + } ]- [ {\text{H}}_{\text{out}}^{ + } ] \). The function \( k_{{{\text{H}}^{ + } }} (T ) \) was parametrized by fitting calculated values of Jpass to experimentally measured proton fluxes determined for chloroplasts in the metabolic state 4 (compare experimental and theoretical data in Fig. 4c). In this work, we have considered three models of the temperature-dependent correction factor for the passive leak of protons, \( k_{{{\text{H}}^{ + } }} (T )= \kappa_{\text{pass}} (T )\cdot k_{\text{pass}}^{\text{o}} \), where \( k_{\text{pass}}^{\text{o}} \) is the normalizing coefficient related to temperature t0 = 25 °C (Fig. 2d).
The calculated temperature dependence adequately reproduces the experimental data on the proton leak in the temperature range 10–35 °C, demonstrating the inflexion at 25 °C. Thus, following experimental data presented in Nolan (1981) and Tikhonov et al. (1983), we could suggest that the temperature dependence of the function Jpass has two branches described by different apparent activation energies.
Simulation of the membrane fluidity effects on structure–function relationships in thylakoids
In the context of the problem of structure–function relationships, observed upon the comparison of the functional (electron transport, proton translocation, and ATP synthesis) and structural characteristics of thylakoid membranes, it was interesting to consider the models, which differ with respect to the temperature-dependent patterns of partial photosynthetic reactions. We have considered the models characterized by different values of parameter t0 related to the inflexion (break) points of temperature dependence of partial photosynthetic reactions, which are assumed to reflect the temperature of structural transitions in the lipid domains of thylakoid membranes. Figure 2 visualizes our choice of the temperature-dependence plots of four temperature-dependent correction functions related to the break points at t0 = 20, 25, and 30 °C. As a touchstone for the choice of these dependences we use experimental data (Tikhonov et al. 1980, 1981, 1983, 1984; Timoshin et al. 1984) on the study of partial temperature dependences of photosynthetic processes in class B chloroplasts isolated from bean leaves.
Results
Redox transients of P700
P700 redox transients are indicative of the intersystem ETC activity. Taking into account a rapid diffusion of PQH2 in the membrane, and a high mobility of Pc within the lumen, one can conclude that it is the reaction of electron transfer from PQH2 bound the quinone-binding portal of the Cyt b6f complex that determines the overall rate of PQ turnover between PSII and PSI (for references, see Tikhonov 2014, 2018). Indeed, over a wide range of experimental conditions (pH, ionic strength, and temperature), the processes of QB reduction to QBH2 in PSII, PQH2 dissociation from PSII and its diffusion across and along the thylakoid membrane towards the Cyt b6f complex take less time than PQH2 oxidation in the quinone-binding center Qo (Tikhonov et al. 1984). Within the Cyt b6f complex, electron transfer from the reduced iron–sulfur protein (ISPred) to Cyt f proceeds more rapidly (t1/2 ≤ 2–4 ms, Gong et al. 2001; Yan and Cramer 2003) than PQH2 oxidation (PQH2 → ISP, t1/2 ~ 10–20 ms, Stiehl and Witt 1969; Witt 1979). This suggests that namely the rate of PQH2 oxidation after the formation of the substrate–enzyme complex (PQH2-ISPox) determines the overall rate of electron transfer from PQH2 to PSI.
As we noted above, the intersystem electron transport is governed by the light-induced changes in the lumen pH (pHin). The light-induced acidification of the lumen causes the deceleration of PQH2 oxidation by the Cyt b6f complex, thereby reducing the rate of the intersystem electron transfer. The down-regulation of electron transport, caused by a decrease in pHin, is associated with the back-pressure of the intra-thylakoid hydrogen ions on the proton-coupled oxidation of PQH2 by the Cyt b6f complex (for references, see Tikhonov 2014). Within the framework of our model, we take into account the influence of pHin on the rate of PQH2 oxidation (“Appendix 3”, Fig. 13). pH-dependent regulation of the intersystem electron transport in chloroplasts manifests itself in the kinetics of the light-induced redox transients of P700 (Tikhonov et al. 1981; Vershubskii and Tikhonov 2020). Below we consider the results of modeling electron transport in class B chloroplasts in more details.
Kinetics of the light-induced redox transients of P 700
Figure 5 presents calculated time-courses of P700 redox transients induced by “Light 1” and “Light 2”. In agreement with experimental data (Tikhonov et al. 1981), “Light 1” induces oxidation of P700. Change-over from “Light 1” (exciting preferentially PSI) to “Light 2” (efficiently exciting both PSI and PSII) causes a decrease in \( \left[ {{\text{P}}_{700}^{ + } } \right] \) due to electrons donated by PSII. The steady-state concentration of \( {\text{P}}_{700}^{ + } \) and the rate of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) depend on the metabolic state of chloroplasts. The kinetic curves presented in Fig. 5 are calculated for three metabolic states: state 3 (active functioning of the ATP synthase), state 4 (without ADP), and state 5 (uncoupled chloroplasts, ΔpH = 0). The \( {\text{P}}_{700}^{ + } \) level is maximal in state 4, when the oxidation of PQH2 is retarded due to a sufficiently strong acidification of the lumen. In state 3, the \( {\text{P}}_{700}^{ + } \) level is reduced. This is because the efflux of proton through the ATP synthase causes a certain decrease in ΔpH, thereby accelerating the intersystem electron flow. In uncoupled chloroplasts (state 5, ΔpH = 0), rapid electron flow to \( {\text{P}}_{700}^{ + } \) leads to more significant decrease in \( \left[ {{\text{P}}_{700}^{ + } } \right] \). In states 3 and 5, the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) occurs much more rapidly than in state 4.

Calculated time-courses of P700 redox transients (in metabolic states 3, 4, and 5) induced by the far-red light (FRL), exciting preferentially PSI, and white light (WL), efficiently exciting both PSI and PSII. Computer simulations refer to the standard model (parameter t0 = 25 °C) and room temperature (25 °C)
Effects of temperature on redox transients of P 700
The short-term mechanism of the temperature influence on the intersystem electron transport can be realized in several ways. One of the mechanisms represents a general influence of the temperature variations on the activity of partial chemical reactions that can be approximated by the Arrhenius law. Another mechanism may be related to temperature-dependent structural changes in the thylakoid membrane: variation of temperature may affect the membrane fluidity, for example, accelerating (or slowing down) diffusion of PQ and PQH2 molecules in the lipid moiety of the membrane. The acceleration (or deceleration) of PQH2 oxidation in the quinone-binding portal within the Cyt b6f complex may be controlled by the physical state of the thylakoid membrane. PQH2 oxidation is the proton-coupled electron transport process associated with the release of two protons into the lumen. The temperature-induced acceleration of proton dissociation from PQH2 through the proton-conducting channels within the Cyt b6f complex will promote the oxidation of PQH2.
Figure 6 presents the temperature plots of the kinetic parameter τ1/2 (panel a) and the Arrhenius plots of its reciprocal value \( \tau_{1/2}^{ - 1} \) (panel b), computed for the metabolic states 3, 4, and 5. Parameter τ1/2 is the half-time of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \); its reciprocal value (\( \tau_{1/2}^{ - 1} \)) characterizes the overall rate of electron flow from PQH2 to \( {\text{P}}_{700}^{ + } \). Temperature dependences of the kinetic parameters τ1/2 and \( \tau_{1/2}^{ - 1} \) adequately describe the experimental plots (Tikhonov et al. 1980, 1984). At any given temperature, τ1/2 has the highest value in state 4 (“photosynthetic control”, maximal ΔpH), and the lowest value in state 5 (ΔpH = 0). This difference is explained by the retardation of PQH2 oxidation caused by acidification of the lumen. The half-times of \( {\text{P}}_{700}^{ + } \) reduction in the states 3 and 5 (at temperatures above 25 °C) are in the range τ1/2 ≈ 15–20 ms (Fig. 6a). We note that similar values of τ1/2 were reported for isolated spinach chloroplasts (Stiehl and Witt 1969; Haehnel 1973, 1976, 1984), and bean chloroplasts (Tikhonov et al. 1981, 1984). In Fig. 6b we also present the kinetic data in the form of the Arrhenius plot, which is traditionally used to evaluate the activation energies of biochemical reactions. Figure 6b shows that in all metabolic states the temperature dependences of electron transfer to PSI reveal characteristic inflexions at t0 ≈ 25 °C. The low-temperature and the high-temperature branches of the temperature dependence of parameter \( \tau_{1/2}^{ - 1} \) are characterized by different activation energies, \( E_{\text{a} }^{ ( 1 )} \)and \( E_{\text{a}}^{ ( 2 )} \), related to the temperature ranges below and above t0, respectively. In the first case (below t0), the exponential acceleration of electron transport with temperature is characterized by\( E_{\text{a}}^{ ( 1 )} \) ~ 60 kJ mol−1. At temperatures above t0, the stimulating effect of temperature is insignificant or absent (\( E_{\text{a}}^{ ( 2 )} \) ≤ 3 kJ mol−1). This result may be accounted for by the interplay of different temperature-dependent factors that influence the electron flow from PQH2 to PSI (via the Cyt b6f complex and Pc).

Temperature dependences of the half-time \( \tau_{1/2} \) of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) (panel a) and the Arrhenius plot of its reciprocal value \( \tau_{1/2}^{ - 1} \) (panel b) computed for metabolic states 3, 4, and 5, as indicated
The two branches of the Arrhenius plot of \( \tau_{1/2}^{ - 1} \) can be explained by the balance of two effects: (i) a general activation of the apparent rate of PQH2 oxidation by temperature, which dominates in the range of temperatures below t0, and (ii) a decrease in [PQH2] and structural transitions in the lipid domains of the membrane (above t0). Both factors are pre-determined by parametrization of the function \( k_{\text{Q}}^{{}} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ] ,T} \right) \) and the model parameters \( \xi (T ) \) and \( \kappa_{\text{PSII}}^{{}} (T ) \) (Section “Effects of temperature on the partial reactions of electron transport in thylakoids”). The rate of the oxidation of PQH2, which donates electrons for the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) centers, will be proportional, at first approximation, to the product \( \left[ {{\text{PQH}}_{ 2} } \right] \cdot \left[ {b_{ 6} f} \right]_{\text{ox}} \), where [b6f]ox stands for oxidized Cyt b6f complexes. This relationship suggests that the initial rate of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) will depend on the plastoquinol concentration ([PQH2]) at the moment of switching the light off.
Figure 7 shows the temperature influence on the steady-state concentrations of \( {\text{P}}_{700}^{ + } \) (panel a) and PQH2 (panel b). It is remarkable that, in the states 3 and 4, [\( {\text{P}}_{700}^{ + } \)] changes insignificantly with temperature, while the concentration of PQH2 drops dramatically at temperatures above 30 °C. Thus, at high enough temperatures (> 30 °C) the electron flow from PQH2 to PSI will not increase with temperature. This is the reason for relatively low \( E_{\text{a}}^{ ( 2 )} \) values (< 3.5 kJ mol−1). Note that in state 5, the Arrhenius plot of \( \tau_{1/2}^{ - 1} \) at t > 30 °C formally shows negative \( E_{\text{a}}^{ ( 2 )} \). It is highly likely that this is the manifestation of both factors: a decrease in the concentration of PQH2, and an accelerated decay of \( \left[ {{\text{P}}_{700}^{ + } } \right] \).

Steady-state concentrations of \( {\text{P}}_{700}^{ + } \) (panel a) and PQH2 (panel b), and acidification of lumen pHin (panel c), computed for metabolic states 3, 4 and 5, as indicated. Computer simulations refer to the standard model (t0 = 25 °C)
Acidification of the lumen (pHin↓) is an essential factor of the electron transport control in chloroplasts (for reviews, see Tikhonov et al. 1981; Kramer et al. 1999; Tikhonov 2012, 2013). Figure 7c displays the computed steady-state pHin values in different metabolic states. In state 4 (when the overall proton flux through the CF0 complex is virtually zero), pHin decreases more significantly than in state 3 (when the protons located in the lumen can escape via the active CF0–CF1 complexes). In both states 3 and 4, the lumen becomes less acidic with increasing the temperature. This can be explained by the temperature-dependent acceleration of the proton outflow from the lumen through the ATP synthase and passive proton flux.
Summing up the above results, we conclude that the two branches of the temperature dependence of the rate of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) may be explained by the interplay of two factors that influence the overall rate of the electron flow from PQH2 to PSI. First, at sufficiently high temperatures (≥ 20–25 °C), occurs the depletion of the pool of reduced PQH2 molecules (Fig. 7b), which serves as a source of electron donors transferred to \( {\text{P}}_{700}^{ + } \) (via the Cyt b6f complex and Pc). This causes the slowing down of the overall electron flux to PSI. Secondly, the inflexion in the plot of the temperature dependence may also reflect thermo-induced structural changes in the thylakoid membranes. The assumption about the temperature-dependent “structure–function” relationship was laid down upon the parametrization of the function \( k_{\text{Q}}^{{}} \left( { [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ] ,T} \right) \) according to (Eq. 2).
Electron transport coupled to ATP synthesis
Here we consider the relationship between the non-cyclic electron flow around PSI and ATP synthesis in thylakoids. Figure 8a presents the temperature dependences of steady-state electron fluxes from Fd− to O2 (JFd–O2) established in the metabolic states 3, 4, and 5, as calculated for our basic model (t0 = 25 °C). These plots have bell-like shapes, with the maxima at 25 °C. The most intensive electron flow to O2 (JFd–O2) occurs in uncoupled chloroplasts (state 5, ΔpH = 0), when the intersystem electron flow is not retarded by the lumen acidification. In states 3 and 4, the JFd–O2 fluxes are reduced due to the ΔpH-dependent retardation of the intersystem electron transport. In Fig. 8b we compare the temperature dependences of VATP (the rate of ATP formation) and JFd–O2 (the rate of electron transport). The ratio VATP/JFd–O2 corresponds to the experimentally measured P/2e ratio (also termed as the ATP/O ratio), which is conventionally used as a measure for the energy coupling efficiency in bioenergetic systems (Chance and Williams 1956; Mitchell 1976; Ivanov 1993; Rigoulet et al. 1998). Experimentally determined P/2e ratio also increases with temperature, reaching the value ≈ 0.8–1.2 at 22–25 °C (depending on experimental conditions), but remains constant at higher temperatures (Timoshin et al. 1984; see also Fig. 15 in “Appendix 3”). The rise of P/2e can be explained, at least partly, by temperature-induced activation of the ATP synthase. In our model, this factor is tacitly considered by the assumption that the model parameter kATP(T) increases with temperature. The ratio P/2e depends on the conditions in which the chloroplasts are isolated and their properties are assayed (Reeves et al. 1972; Heise and Harnischfeger 1978; Ivanov 1993). It is also conceivable that the composition of the membrane lipids may affect ATP synthesis, due to their influence on the H+ ion translocation from the proton pumps to the proton sinks (the CF0 part of the ATP synthase).

Temperature dependences of steady-state electron fluxes from reduced ferredoxin (Fd−) to O2 (JFd–O2) established in metabolic states 3, 4, and 5 calculated for the basic model characterized by parameter t0 = 25 °C (panel a). In panel b we compare the temperature dependence of VATP and JFd–O2; parameter P/2e = VATP/JFd–O2 characterizes efficiency of coupling electron transport to ATP synthesis
Experimental studies show significant variability of photophosphorylation temperature dependences in chloroplasts isolated from the leaves grown under different experimental conditions (Yamori et al. 2014). Below we consider experimental data on bean chloroplasts, for which we observed the correlation between the temperature dependences of photophosphorylation and fluidity of membrane lipids (for references, see Kukushkin and Tikhonov 1988; Tikhonov and Subczynski 2005; Tikhonov 2020). In our earlier works (Tikhonov et al. 1980, 1983, 1984; Timoshin et al. 1984; Lutova and Tikhonov 1983, 1988), lipid-soluble derivatives of nitroxide radicals were used as the paramagnetic probes for thermo-induced structural changes in thylakoid membranes. Spin-labeled derivative of stearic acid, 5-SASL (Fig. 9a) is one of the most convenient probes for structural transients in the lipid domains of thylakoid membranes. Spin-probe molecules are intercalated in the membrane with the hydrophilic part in the polar headgroup region of the membrane. The radical mobility and ordering with respect to fatty acid chains of lipids depend on the position of the nitroxide radical. Alkyl chains of the lipid bilayer portion of the thylakoid membrane are very well ordered in the near polar headgroup region and fairly fluid in the membrane center. The EPR spectra of 5-SASL reflect the mobility and ordering of its nitroxide fragment in the lipid domains of the membrane (Ligeza et al. 1998). When dissolved in the thylakoid membranes, 5-SASL gives the EPR spectrum (Fig. 9b) typical of nitroxide radicals located in a hydrophobic membrane environment. The rotational mobility of the nitroxide fragment influences the spectral parameters \( 2T_{\text{II}}^{\prime } \)and \( 2T_{ \bot }^{'} \)of the EPR signal, which equal to the splitting of the “outer” and “inner” peaks (for definitions, see Fig. 9b). A degree of the nitroxide radical ordering in the lipid bilayer can be characterized by the so-called order parameter S, which can be calculated from the \( 2T_{\text{II}}^{'} \) and \( 2T_{ \bot }^{\prime } \) parameters:

Structure–function correlations in thylakoids. Panel a shows chemical structures of monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG), and the stearic acid spin-probe 5 (5-SASL) used as an indicator of the membrane physical state (fluidity). Galactolipids (MGDG and DGDG) are major components of the lipid portion of thylakoid membranes. Spin label molecules are intercalated in the membrane with the hydrophilic part (left-hand side) in the polar headgroup region of the membrane. Panel b shows the EPR spectrum of 5-SASL dissolved in the thylakoid membrane at room temperature (modified from Ligeza et al. 1998; Tikhonov 2020). Panels c, d, and e show the correlations between the temperature dependences of the structural parameter 2T ′II and the normalized rate of ATP formation (VATP) in chloroplasts isolated from different batches of bean leaves (modified from Tikhonov et al. 1983; Kukushkin and Tikhonov 1988)
where Txx, Tyy, and Tzz are the main values of the hyperfine splitting tensor, and h is the correction factor for hydrophobicity of the local surrounding of the nitroxide radical (Berliner 1976; Griffith and Jost 1976; McConnell 1976). The order parameter S decreases with depth of the radical location in the thylakoid membrane (Ligeza et al. 1998). The S value is proportional to \( 2T_{\text{II}}^{'} \), the value of which can be easily and reliably determined in a wide range of temperatures. Therefore, in most of our previous works, the spectral parameter \( 2T_{\text{II}}^{'} \)was routinely used to assay temperature-induced changes in the membrane fluidity.
In Fig. 9 (panels c, d, and e) we compare the temperature dependences of the spectral parameter \( 2T_{\text{II}}^{'} \) determined for spin-probe 5-SASL dissolved in thylakoid membranes of bean chloroplasts isolated from the leaves of different harvests (plants were grown under lower or higher temperatures; for more details, see Tikhonov et al. 1983; Kukushkin and Tikhonov 1988). All the plots of \( 2T_{\text{II}}^{\prime } \) reveal characteristic inflexions (“breaks”) of 2T ′II at different temperatures, at 25 °C (Fig. 9c), at 30 °C (Fig. 9d), and at 33 °C (Fig. 9e), suggesting that the thermo-induced “melting” of membrane lipids occurred at different temperatures. This may be caused, for example, due to different proportions of unsaturated and saturated fatty acids of the membrane lipids.
Experimental temperature dependences of ATP synthesis (VATP) show parabolic patterns: VATP increases with temperature, reaching the maximal value and then drops to zero. Note that the temperature corresponding to the inflexion in the plot of \( 2T_{\text{II}}^{\prime } \) always coincides with the temperature at which the rate of ATP synthesis is maximal. A thermo-induced increase in VATP in the range of temperatures below t0 is explained by the acceleration of electron transport and activation of the ATP synthase. At temperatures above t0, VATP decreases with the rise of temperature. This may occur (i) due to a lessening of the electron flow from PSII to PSI, and (ii) due to an increase in the passive proton leak bypassing the ATP synthase. The temperature-induced increase in the membrane fluidity will accelerate electron transport and proton pumping into the lumen, on the one hand, while the acceleration of passive proton leakage (which bypass the ATP synthase) and the enhancement of ATP hydrolysis will suppress the ATP synthesis, on the other hand. As noted above, the maximal rates of the net ATP synthesis coincide with the temperatures of inflexion points in the plots of the “structural” parameter \( 2T_{\text{II}}^{\prime } \). This observation can be considered as an evidence in favor of the regulatory role of the membrane fluidity in controlling the photosynthetic processes in thylakoids. It is highly likely that the proper balance between the gel (liquid-crystalline) and fluid phases in the thylakoid membrane, established at certain temperature, supports optimal conditions for efficient operation of photosynthetic apparatus upon fluctuations of temperature (Tikhonov 2020).
The variability of temperature dependences of VATP observed in experiment can be modeled within the framework of our model. Figure 10 depicts calculated temperature dependences of VATP for the models, characterized by parameters t0 = 20, 25, and 30 °C. Similar to experimental dependences, the calculated dependences VATP(t) have the bell-like shapes. In accordance with experimental data, the model predicts that the temperature, at which calculated JATP values is maximal, coincides with the inflexion point at temperature t0 characterizing the structural transition the thylakoid membrane (e.g., the so-called “melting” of the lipid bilayer). Thus, the model mimics the “structure–function” mechanism of temperature-dependent regulation of photophosphorylation that could be realized by modulation of the membrane fluidity.

Computed temperature dependences of the rate of ATP formation (VATP) for three models related to break points at t0 = 20, 25, and 30 °C, respectively
Discussion
In this work, we describe the results of the computer modeling of temperature-dependent regulation of photosynthetic processes in isolated thylakoids, and compare them to the relevant experimental data obtained earlier on isolated bean chloroplasts. Considering the sites of electron transport control, we have focused on the analysis of plastoquinone turnover between PSII and the Cyt b6f complex, assuming that the reaction of PQH2 oxidation is the rate-limiting step in the intersystem ETC. Rapid shuttling of PQ and PQH2 between the PSII and Cyt b6f complexes is determined by their relatively high mobility in the lipid domains of the membrane. The overall rate of the intersystem electron transfer is determined mainly by the rate of PQH2 oxidation at the quinol-binding site (Qo) localized in the interior of the Cyt b6f complex, oriented towards the lumen. Close location of PSII and granal Cyt b6f complexes, on the one hand, and sufficiently high fluidity of the membrane lipids, on the other hand, promote the formation of “substrate–enzyme” complex (PQH2-b6f), thereby accelerating the intersystem electron transport. The bifurcated reaction of PQH2 oxidation starts with the transfer of one hydrogen atom from PQH2 to the oxidized iron–sulfur protein (ISPox): PQH2-ISPox → PQH•-ISPredH. There are good reasons to believe that it is this step of PQH2 oxidation that determines the overall rate of PQ turnover inside the Cyt b6f complex (Crofts and Wang 1989; Hong et al. 1999; Crofts 2004; Crofts et al. 2000, 2013). Quantum chemical calculations (Ustynyuk and Tikhonov 2018) suggest that the activation energy of this reaction, Ea ≈ 60 kJ/mol, is close to the apparent activation energy for electron transfer from PQH2 to \( {\text{P}}_{700}^{ + } \). The effect of the membrane physical state (fluidity) on the rate of PQH2 oxidation may be accounted for by the step of the proton dissociation from PQH2 to the lumen (via the histidine residue of the ISP), which may have a restraining effect (Crofts et al. 2000).
The two branches of the temperature dependence of the post-illumination reduction \( {\text{P}}_{700}^{ + } \) described by our model reflect the interplay of several processes. The inflexion in the temperature dependence of the rate of \( {\text{P}}_{700}^{ + } \) reduction may have kinetic and structural reasons. Results of our calculations support this viewpoint. Considering the kinetic factor that determines the intersystem electron flow, we have to note that electron flow from PQH2 to \( {\text{P}}_{700}^{ + } \) (through the Cyt b6f complex) will accelerate with the concentration of reduced PQH2, i.e., \( J_{{{\text{QH}}_{2} \to {\text{P}}_{700}^{ + } }} \sim [ {\text{PQH}}_{ 2} ] \) (Tikhonov and Vershubskii 2017; Suslichenko and Tikhonov 2019). The higher is the PQH2 concentration, the faster electrons are delivered to \( {\text{P}}_{700}^{ + } \). A steady-state concentration of PQH2 is determined by the balance between two processes: (1) the PQH2 formation in PSII and (2) PQH2 oxidation by the Cyt b6f complex. The model predicts that [PQH2] gradually decreases at temperatures above 30 °C (Fig. 7b). Thus, a decrease in [PQH2] at sufficiently high temperatures, caused by the reduction of PSII activity, will hamper the intersystem electron flux through the Cyt b6f complex, which will manifest itself as an apparent decrease in E (2)a .
Another reason for a decrease in \( E_{\text{a}}^{(2)} \) at t ≥ 25 °C may be associated with thermo-induced structural changes in thylakoids. Lipids provide the diffusion medium for PQH2 molecules moving towards the PQH2-binding portal located within the Cyt b6f complex. The “solidification” of membrane lipids caused by a decrease in temperature (≤ 20–25 °C) should hamper the PQH2 penetration into the quinone-binding portal in the Cyt b6f complex, decelerating the PQH2 oxidation. Otherwise, the membrane fluidization with the rise of temperature will accelerate the oxidation of PQH2. An increase in the PQH2 mobility in the membrane, caused by lipid fluidization at sufficiently high temperatures (≥ 25 °C), will promote the formation of the substrate–enzyme complex (PQH2-b6f) and accelerate the oxidation of PQH2. However, after the transition of the most part of membrane lipids into the “fluid” state, the stimulating effect of temperature on electron transport will be masked by a decrease in [PQH2]. Therefore, from the phenomenological viewpoint, a decrease in [PQH2] above sufficiently high temperatures (> t0) will appear as a decrease in the apparent activation barrier \( E_{\text{a}}^{(2)} \).
Our line of arguments in favor of temperature-dependent modulation of the membrane fluidity as the regulatory factor in chloroplasts is based on experimental data on the structure-function relationships in thylakoids. The results of spin-probe studies of thermo-induced structural transitions in thylakoid membranes are in agreement with the notion that an adaptation of the photosynthetic apparatus of higher plants to changes in environmental temperature (Boardman 1977; Berry and Björkman 1980; Gounaris et al. 1984; Allen and Ort 2001) could be realized by fitting the fluidity of the lipid domains to an optimal level. Alterations in the membrane fluidity can be controlled by temperature-induced changes in the composition of unsaturated and saturated lipids in thylakoid membranes (Moon et al. 1995; Nie and Baker 1991). There is a rather strong evidence that the thermo-induced modulation of membrane fluidity markedly affects the rates of electron and proton transport processes in photosynthetic systems of oxygenic type (see Los et al. 2013; Yamori et al. 2014; Maksimov et al. 2017; Tikhonov 2020 and references therein). The coexistence of fluid and crystalline phases in photosynthetic membranes is believed to support physiologically relevant conditions (Moon et al. 1995; Schneider and Geissler 2013). The correspondence between the temperature dependences of kinetic and structural parameters in bean chloroplasts was demonstrated by the EPR method for chloroplasts isolated from the plants of various harvests (Tikhonov et al. 1983; Kukushkin and Tikhonov 1988). Changes in growth conditions (i.e., the cultivation temperature) caused similar changes in the inflexion point positions in the plots of \( \tau_{1/2}^{ - 1} \) (the rate of \( {\text{P}}_{700}^{ + } \) reduction) and spectral parameters of lipid-soluble spin probes. When plants were grown at reduced temperatures, the inflexion points drifted towards lower temperatures; at elevated growth temperatures, the inflexion points shifted towards higher temperatures. Taking into account the reproducible correlations between “kinetic” and “structural” parameters (Fig. 9, panels c, d, and e), one can conclude that the rate of the intersystem electron transfer is affected by the membrane fluidity. It is conceivable that “melting” of the membrane lipids, when the vast majority of lipid molecules are in the “fluid” state, would accelerate the substrate–enzyme complex (PQH2-b6f) formation. The light-induced pumping of proton into the lumen supports the ATP synthesis, while the thermo-induced disordering of the membrane lipids stimulates the passive proton leakage, which bypasses the ATP synthase (for illustration, see sketch in Fig. 11). The latter factor will reduce the net rate of ATP formation at high temperatures.

A symbolic sketch qualitatively illustrating the effect of the redistribution of proton fluxes JATP and Jpass caused by temperature-induced structural changes in the physical state of the thylakoid membrane. Panel a reflects the up-regulation of the temperature dependence of ATP synthesis, VATP(T). Stimulation of ATP synthesis occurs due to a general temperature-dependent activation of the ATP synthase, including the acceleration of the cn-ring rotation in the lipid moiety with the temperature-induced fluidization of the thylakoid membrane. In this case, stimulation of ATP synthesis with temperature prevails over the loss of protons caused by their bypassing through the membrane. Panel b illustrates the down-regulation of ATP formation at sufficiently high temperatures, when temperature induces more significant leak of protons (Jpass) due to increasing the room of fluid domains in thylakoid membranes, thereby decreasing VATP(T)
Below we consider two questions: (1) how the membrane fluidity could influence the rate of PQH2 oxidation reaction, and (2) why the “fluidization” of the membrane causes an apparent decrease in the activation energy of electron transport processes (\( E_{\text{a}}^{ ( 1 )} \) > \( E_{\text{a}}^{ ( 2 )} \))? As noted above, the rate of PQ turnover is determined predominantly by the intrinsic events within the Cyt b6f complex: the penetration of PQH2 into the quinone-binding site Qo, and the subsequent oxidation of PQH2, which is accompanied by the release of protons into the bulk of the lumen. In the catalytic site Qo of the Cyt b6f complex, PQH2 is oxidized by the iron–sulfur protein (ISP), which accepts the H atom (electron + proton) from PQH2. According to the “proton-gated” model of quinol oxidation (Brandt 1996; Link 1997), in order to create a sufficiently high reducing potential, the PQH2 molecule must be deprotonated (PQH2 → PQH− + H+) before the first electron can be transferred to the oxidized iron–sulfur protein (ISPox). Both the electron and proton are transferred in concert to the Fe2S2 cluster of the ISPox and to the His residue of the ISPox (for references, see Crofts and Wang 1989; Crofts 2004; Osyczka et al. 2005; Cramer et al. 2006, 2011). Then the proton dissociates from the ISPox to the bulk of the lumen through a specific proton-conducting channel (Hasan et al. 2013a; Tikhonov 2014, 2018). Thus, taking into account that PQH2 oxidation is the proton-coupled electron transport process associated with dissociation of H+ ions into the lumen, we can suggest that the proton transfer through the membrane may serve as a factor controlling the rate of PQ turnover.
The operation of the ISP is associated with its conformational changes that might be another possible factor controlling the rate of PQH2 oxidation. Extensive crystallographic disorder of the ISP extrinsic domain indicates its conformational mobility and flexibility (Hasan and Cramer 2012; Hasan et al. 2013b). The Fe2S2 cluster serves as the recipient of an electron donated by PQH2 and then it donates the electron to Cyt f. After the reduction of the ISP, the mobile domain of the ISPred, which contains the redox center (the Fe2S2 cluster), moves from the Qo site towards the heme f. After the reduction of Cyt f, the oxidized Fe2S2 cluster returns the Qo site. The roundtrip of the mobile domain of the ISP to heme f and back to the Qo site can partly contribute to the turnover time of the Cyt b6f complex. However, the structural and kinetic data suggest that the “tethered” movement of the mobile domain of the ISP is not a rate-limiting step for electron transfer inside the Cyt b6f complex. The restricted diffusion of the ISP redox center occurs rapidly compared to the overall rate of PQH2 oxidation. Electron transfer from the reduced ISP to Cyt f proceeds more rapidly (t1/2 ≤ 2–4 ms, Gong et al. 2001; Yan and Cramer 2003) than PQH2 oxidation (t1/2 ~ 10–20 ms; Stiehl and Witt 1969; Witt 1979). This means that the rate of PQH2 oxidation is determined predominantly by the proton-coupled electron transfer after the formation of the substrate–enzyme complex (PQH2-ISPox).
Lipids may play important role in operation of the Cyt b6f complex. This complex encloses a cavity (30 Å × 15 Å × 25 Å), which serves as the portal for PQH2 binding to the catalytic center (Cramer et al. 2006). Inside the cavity, there were identified 23 lipid-binding sites per monomer of the dimeric b6f complex from Noctos PCC 7120 (Hasan and Cramer 2014). It is conceivable that the neutral lipids localized inside the quinone-binding cavity may determine enhanced conformational flexibility of the ISP mobile domain. It is essentially in this context that lipids may play important role in regulation of electron transfer through the Cyt b6f complex. In thylakoid membranes, most of the lipids are presented by galactolipids, which contain polyunsaturated fatty acids (Wada and Murata 2009; Zhou et al. 2016). Variations in the relative content of unsaturated fatty acids in thylakoid membranes were found to be one of the factors that control the photosynthetic activity of chloroplasts (Los and Murata 2004; Los et al. 2013; Maksimov et al. 2017). We may speculate that the “fluidization” of the thylakoid membrane will facilitate the proton transfer from PQH2 to the bulk of the lumen, thereby stimulating the deprotonation of reduced ISPredH+ (ISPredH+ → ISPred + \( {\text{H}}_{\text{in}}^{ + } \)) and further electron transfer from ISPred to Cyt f. A rapid proton transfer into the lumen through the proton channels of the Cyt b6f complex would accelerate the Cyt b6f turnover. Thermo-induced conformational changes in the Cyt b6f complex might also stimulate PQH2 oxidation. Thus, the temperature-dependent release of diffusion barrier for protons and concomitant acceleration of PQH2 oxidation should promote electron transfer between PSII and PSI. It is conceivable that after the temperature-dependent “melting” of the vast majority of thylakoid lipids (the “fluidization” of the membrane), the rate of the substrate–enzyme complex (PQH2-b6f) formation will depend on temperature, but not significantly.
Concluding remarks
-
1.
For modeling of the photosynthetic electron transport processes in thylakoids, we consider that the processes of PQ turnover (reduction of PQ in PSII, diffusion of PQH2 and its oxidation by the Cyt b6f complex) determine the overall rate of electron transport between PSII and PSI. The rate-limiting step of the intersystem electron transport is associated with PQH2 oxidation at the quinone-binding site of the Cyt b6f complex. The overall rate of the intersystem electron transfer is determined mainly by the rate of PQH2 oxidation at the quinol-binding site localized in the interior of the Cyt b6f complex. The feedback control of PQH2 oxidation is governed by the intra-thylakoid pH. Rapid shuttling of electrons between PSII and Cyt b6f complexes by PQH2 is determined by a high mobility of PQH2 and PQ molecules within the lipid domains of the thylakoid membrane.
-
2.
Both the PQ/PQH2 movement within the thylakoid membrane and PQH2 oxidation within the Cyt b6f complex are the temperature-dependent processes. Structural changes in the membrane are the clue factors of temperature-dependent regulation of photosynthesis. The intersystem electron transfer and related processes (e.g., proton pumping, ATP synthesis, and trans-thylakoid proton transfer) are controlled by the degree of membrane fluidity. The mechanism of fluidity-dependent regulation of photosynthetic processes is supported by correlations between the functional characteristics (electron and proton transport, ATP formation) and “structural” properties of the thylakoid membrane (i.e., membrane fluidity).
-
3.
Computer modeling of electron and proton transport processes supports the notion that PQH2 oxidation by the Cyt b6f complex and the processes of transmembrane H+ ion transfer are the basic temperature-dependent steps that determine the overall flux of electrons from PSII to molecular oxygen and the net ATP synthesis upon variations of temperature. Numerical experiments have demonstrated that the temperature dependences of these processes, which are sensitive to the physical state of the membrane, will determine the temperature-dependent pattern of PQH2 oxidation.
-
4.
The two branches of the temperature effect on the intersystem electron transport can be explained by (1) a general temperature effect on the activity of PSII, and the rate of PQH2 oxidation by the Cyt b6f complex, and by (2) thermo-induced structural changes in the thylakoid membranes, which can influence the proton transfer through the membrane, accelerating the proton leak with temperature. The low-temperature branch of activation of the intersystem electron transport (Ea ~ 60 kJ mol−1) is explained by the temperature-dependent acceleration of PQH2 oxidation. The attenuation of the activation effect at temperatures ≥ 25 °C can be explained by a gradual decrease in the concentration of PQH2 and thermo-induced structural changes in thylakoid membranes.
-
5.
The model describes the bell-like temperature dependence of ATP synthesis as resulting from the interplay of several factors: (1) the thermo-induced acceleration of electron transport through the Cyt b6f complex, (2) a deactivation of PSII photochemistry at temperatures above the structural transient, and (3) an acceleration of the passive proton outflow from the thylakoid lumen, bypassing the ATP synthase complex, which is caused by an increase in the permeability of thylakoid membranes at temperatures above the structural transient. The model also describes the temperature dependence of experimentally measured parameter P/2e, which is determined as the ratio between the rates of ATP synthesis and pseudocyclic electron transport.
Summing up the results of our study, we can state that, though the model described above is simplified, it recapitulates many of the temperature-dependent responses observed in the thylakoids in vitro (isolated class B chloroplasts). In particular, the model describes two branches of the temperature dependences of pseudocyclic electron transport and the bell-like plot of the ATP formation versus temperature. The future work on the theoretical study of temperature dependences of photosynthetic reactions in chloroplasts will need the expansion of the model presented here, including the consideration of (1) the Mitchell’s Q-cycle; (2) alternative electron transport pathways (non-cyclic/cyclic), (3) the light-induced redox regulation of the ATP synthase, and (4) the metabolism-related processes like the CBC reactions.
Abbreviations
- CBC:
-
Calvin–Benson cycle
- DGDG:
-
Digalactosyldiacylglycerol
- EPR:
-
Electron paramagnetic resonance
- ETC:
-
Electron transport chain
- Fd:
-
Ferredoxin
- FNR:
-
Ferredoxin-NADP-oxidoreductase
- ISP:
-
Iron–sulfur protein
- MGDG:
-
Monogalactosyldiacylglycerol
- ODE:
-
Ordinary differential equations
- Pc:
-
Plastocyanin
- PG:
-
Phosphatidylglycerol
- pmf :
-
Proton-motive force
- PSI and PSII:
-
Photosystem I and photosystem II, respectively
- PQ and PQH2 :
-
Plastoquinone and plastoquinol (fully reduced form of PQ), respectively
- P700 :
-
Special chlorophyll pair in PSI, primary electron donor in PSI
- P680 :
-
Special chlorophyll pair in PSII, primary electron donor in PSII
- QA and QB :
-
Primary and secondary plastoquinone molecules bound to PSII
- SQDG:
-
Sulfoquinovosyldiacylglycerol
- T :
-
Temperature in Kelvin scale
- t :
-
Temperature in Celsius scale
- τ :
-
Time
- WOC:
-
Water-oxidizing complex
- 5-SASL:
-
Spin probe 5-doxylstearate
References
Agalarov R, Brettel K (2003) Temperature dependence of biphasic forward electron transfer from the phylloquinone(s) A1 in photosystem I: only the slower phase is activated. Biochim Biophys Acta 1604:7–12
Albertsson PE (2001) A quantitative model of the domain structure of the photosynthetic membrane. Trends Plant Sci 6:349–354
Allakhverdiev SI, Kreslavski VD, Klimov VV, Los DA, Carpentier R, Mohanty P (2008) Heat stress: an overview of molecular responses in photosynthesis. Photosynth Res 98:541–550
Allen DJ, Ort DR (2001) Impact of chilling temperatures on photosynthesis in warm climate plants. Trends Plant Sci 6:36–42
Aloia RA, Boggs JM (eds) (1985) Membrane fluidity in biology. Academic Press, New York, pp 147–208
Anderson JM (1982) Distribution of the cytochromes of spinach chloroplasts between the appressed membranes of grana stacks and stroma-exposed thylakoid regions. FEBS Lett 138:62–66
Ariga T, Muneyuki E, Yoshida M (2007) F1-ATPase rotates by an asymmetric, sequential mechanism using all three catalytic subunits. Nat Struct Mol Biol 14:841–846
Arnold A, Nikoloski Z (2011) A quantitative comparison of Calvin-Benson cycle models. Trends Plant Sci 16:676–683
Asada K (1999) The water–water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Ann Rev Plant Physiol Plant Mol Biol 50:601–639
Ataullakhanov FI, Pichugin AV (1981) Modification of luciferin-luciferase method for ATP assay in erythrocytes. Biophysics (USSR) 26:81–86
Badger MR, Von Caemmerer S, Ruuska S, Nakano H (2000) Electron flow to oxygen in higher plants and algae: rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase. Philos Trans R Soc London 355:1433–1446
Bakker-Grunwald T, van Dam K (1974) On the mechanism of activation of the ATPase in chloroplasts. Biochim Biophys Acta 347:290–298
Bald D, Noji H, Yoshida M, Hirono-Hara Y, Hisabori T (2001) Redox regulation of the rotation of F1-ATP synthase. J Biol Chem 276:39505–39507
Baniulis D, Yamashita E, Zhang H, Hasan SS, Cramer WA (2008) Structure–function of the cytochrome b6f complex. Photochem Photobiol 84:1349–1358
Barber J, Ford RC, Mitchell RA, Millner PA (1984) Chloroplast thylakoid membrane fluidity and its sensitivity to temperature. Planta 161:375–380
Bendall DS, Manasse RS (1995) Cyclic photophosphorylation and electron transport. Biochim Biophys Acta 1229:23–38
Benkov MA, Yatsenko AM, Tikhonov AN (2019) Light acclimation of shade-tolerant and sun-resistant Tradescantia species: photochemical activity of PSII and its sensitivity to heat treatment. Photosynth Res 139:203–214
Benson BB, Krause Jr D (1984) The concentration and isotopic fractionation of oxygen dissolved in freshwater and seawater in equilibrium with the atmosphere. Limnol Oceanogr 29:620–632
Berliner LJ (ed) (1976) Spin labeling: theory and applications. Academic Press, New York-London
Berry J, Björkman O (1980) Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Physiol 31:491–543
Boardman NK (1977) Comparative photosynthesis of sun and shade plants. Ann Rev Plant Physiol 28:335–377
Boudiere L, Michaud M, Petroutsos D, Rebeille F, Falconet D, Bastien O, Roy S, Finazzi G, Rolland N, Jouhet J et al (2014) Glycerolipids in photosynthesis: composition, synthesis and trafficking. Biochim Biophys Acta 1837:470–480
Boyer PD (1997) The ATP synthase—a splendid molecular machine. Annu Rev Biochem 66:717–749
Brandt U (1996) Bifurcated ubihydroquinone oxidation in the cytochrome bc1 complex by proton-gated charge transfer. FEBS Lett 387:1–6
Brettel K (1997) Electron transfer and arrangement of the redox cofactors in photosystem I. Biochim Biophys Acta 1318:322–373
Brettel K, Leibl W (2001) Electron transfer in photosystem I. Biochim Biophys Acta 1507:100–114
Cardona T, Sedoud A, Cox N, Rutherford AW (2012) Charge separation in photosystem II: a comparative and evolutionary overview. Biochim Biophys Acta 1817:26–43
Chance B, Williams GR (1956) The respiratory chain and oxidative phosphorylation. Adv Enzymol 17:65–134
Cherepanov DA, Milanovsky GE, Petrova AA, Tikhonov AN, Semenov AYu (2017) Electron transfer through the acceptor side of Photosystem I: interaction with exogenous acceptors and molecular oxygen. Biochemistry (Moscow) 82:1249–1268
Clever HL, Battino R, Miyamoto H, Yampolski Y, Young CL (2014) IUPAC-NIST Solubility Data Series. 103. Oxygen and Ozone in Water, Aqueous Solutions, and Organic Liquids (Supplement to Solubility Data Series Volume 7). J Phys Chem Ref Data 43(3)
Cramer WA, Hasan SS (2016) Structure-function of the cytochrome b6f lipoprotein complex. In cytochrome complexes: evolution, structures, energy transduction, and signaling. Adv Photosynth Respiration 41:177–207
Cramer WA, Zhang H, Yan J, Kurisu G, Smith JL (2006) Transmembrane traffic in the Cytochrome b6f complex. Ann Rev Biochem 75:769–790
Cramer WA, Hasan SS, Yamashita E (2011) The Q cycle of cytochrome bc complexes: a structure perspective. Biochim Biophys Acta 1807:788–802
Crofts AR (2004) Proton-coupled electron transfer at the Qo-site of the bc1 complex controls the rate of ubihydroquinone oxidation. Biochim Biophys Acta 1655:77–92
Crofts AR, Wang Z (1989) How rapid are the internal reactions of the ubiquinol:cytochrome c2 oxidoreductase? Photosynth Res 22:69–87
Crofts AR, Guergova-Kuras M, Kuras R, Ugulava N, Li J, Hong S (2000) Proton-co;upled electron transfer at the Qo-site: what type of mechanism can account for the high activation barrier? Biochim Biophys Acta 1459:456–466
Crofts AR, Hong S, Wilson C, Burton R, Victoria D, Harrison C, Schulten K (2013) The mechanism of ubihydroquinone oxidation at the Qo-site of the cytochrome bc1 complex. Biochim Biophys Acta 1827:1362–1377
Dau H, Haumann M (2008) The manganese complex of photosystem II in its reaction cycle—basic framework and possible realization at the atomic level. Coord Chem Rev 252:273–295
Davis GA, Rutherford AW, Kramer DM (2017) Hacking the thylakoid proton motive force for improved photosynthesis: modulating ion flux rates that control proton motive force partitioning into Δψ and ΔpH. Philos Trans R Soc B 372:20160381
de Wijn R, van Gorkom HJ (2001) Kinetics of electron transfer from QA to QB in photosystem II. Biochemistry 40:11912–11922
Deamer DW (1987) Proton permeation of lipid bilayers. J Bioenerg Biomembr 19:457–479
Dekker JP, Boekema EJ (2005) Supermolecular organization of the thylakoid membrane proteins in green plants. Biochim Biophys Acta 1706:12–39
Demmig-Adams B, Cohu CM, Muller O et al (2012) Modulation of photosynthetic energy conversion efficiency in nature: from seconds to seasons. Photosynth Res 113:75–88
Díaz-Quintana A, Leibl W, Bottin H, Sétif P (1998) Electron transfer in photosystem I reaction centers follows a linear pathway in which iron–sulfur cluster FB is the immediate electron donor to soluble ferredoxin. BioChemistry 37:3429–3439
Diez M, Zimmermann B, Borsch M, Konig M, Schweinberger E, Steigmiller S, ReuterR Felekyan S, Kudryavtsev V, Seidel CA, Graber P (2004) Proton-powered subunit rotation in single membrane-bound F0F1-ATP synthase. Nat Struct Mol Biol 11:135–141
Dubinskii AYu, Tikhonov AN (1995) Mathematical simulation of the light-induced uptake of protons by chloroplasts upon various mechanisms of proton leak through the thylakoid membrane. Biophysics 40:365–371
Dubinskii AYu, Tikhonov AN (1997) Mathematical model of thylakoid as the distributed heterogeneous system of electron and proton transport. Biophysics 42:644–660
Edwards G, Walker D (1983) C3, C4: mechanisms, and cellular and environmental regulation, of photosynthesis. Univ of California Press, Berkeley
Fillingame RH, Jiang W, Dmitriev OY (2000) Coupling H+ transport to rotary catalysis in F-type ATP synthases: structure and organization of the transmembrane rotary motor. J Exp Biol 203:9–17
Ford RC, Barber J (1983) Incorporation of sterol into chloroplast thylakoid membranes and its effect on fluidity and function. Planta 158:35–41
Ford RC, Chapman DJ, Barber J, Pedersen JZ, Cox RP (1982) Fluorescence polarization and spin-label studies of the fluidity of stromal and granal chloroplast membranes. Biochim Biophys Acta 681:145–151
Foyer CH, Neukermans J, Queval G, Noctor G, Harbinson J (2012) Photosynthetic control of electron transport and the regulation of gene expression. J Exp Bot 63:637–1661
Gong X-S, Chung S, Fernandez-Velasco JG (2001) Electron transfer and stability of the cytochrome b6f complex in a small domain deletion mutant of cytochrome f. J Biol Chem 276:24365–24371
Gounaris K, Brain APR, Quinn PJ, Williams WP (1984) Structural reorganization of chloroplast thylakoid membranes in response to heat-stress. Biochim Biophys Acta 766:198–208
Griffith OH, Jost PC (1976) Lipid spin labels in biological membranes. Spin labeling: theory and applications. Academic Press, New York, pp 456–524
Gutknecht J (1987) Weak electrolyte transport across biological membranes. General principles. J Bioenerg Biomembr 19:427–455
Haehnel W (1973) Electron transport between plastoquinone and chlorophyll aI. Biochim Biophys Acta 305:618–631
Haehnel W (1976) The reduction kinetics of chlorophyll a1 as an indicator of for proton uptake between the light reactions in chloroplasts. Biochim Biophys Acta 440:506–521
Haehnel W (1984) Photosynthetic electron transport in higher plants. Annu Rev Plant Physiol 35:659–693
Hasan SS, Cramer WA (2012) On rate limitation of electron transfer in the photosynthetic b6f complex. Phys Chem Chem Phys 14:13856–13860
Hasan SS, Cramer WA (2014) Internal lipid architecture of the hetero-oligomeric cytochrome b6f complex. Structure 22:1–8
Hasan SS, Yamashita E, Baniulis D, Cramer WA (2013a) Quinone-dependent proton transfer pathways in the photosynthetic cytochrome b6f complex. Proc Natl Acad Sci USA 110:4293–4302
Hasan SS, Stofleth JT, Yamashita E, Cramer WA (2013b) Lipid-induced conformational changes within the cytochrome b6f complex of oxygenic photosynthesis. Biochemistry 52:2649–2654
Heise K-P, Harnischfeger G (1978) Correlation between photosynthesis and plant lipid composition. Z Naturforsch 33:537–546
Hirano M, Satoh K, Katoh S (1981) The effect on photosynthetic electron transport of temperature-dependent changes in the fluidity of the thylakoid membrane in a thermophilic blue-green alga. Biochim Biophys Acta 635:476–487
Hong SJ, Ugulava N, Guergova-Kuras M, Crofts AR (1999) The energy landscape for ubihydroquinone oxidation at the Qo-site of the bc1 complex in Rhodobacter sphaeroides. J Biol Chem 274:33931–33944
Hope AB (2000) Electron transfers amongst cytochrome f, plastocyanin and photosystem I: kinetics and mechanisms. Biochim Biophys Acta 1456:5–26
Horton P (2012) Optimization of light harvesting and photoprotection: molecular mechanisms and physiological consequences. Phil Trans R Soc B 367:3455–3465
Hu S, Ding Y, Zhu C (2020) Sensitivity and responses of chloroplasts to heat stress in plants. Front Plant Sci 11:375. https://doi.org/10.3389/fpls.2020.00375
Igamberdiev AU (2011) Computational models of photosynthesis. BioSystems 103:113–314
Inout H (1978) Break points in Arrhenius plots of the Hill reaction of spinach chloroplast fragments in the temperature range from -25 to 25°C. Plant Cell Physiol 19:355–363
Ivanov BN (1993) Stoichiometry of proton uptake by thylakoids during electron transport in chloroplasts. In: Abrol YP, Mohanty P, Govindjee G (eds) Photosynthesis: Photoreactions to Plant Productivity. Springer, Dordrecht, pp 108–128
Johnson MP, Ruban AV (2014) Rethinking the existence of a steady-state Δψ component of the proton motive force across plant thylakoid membranes. Photosynth Res 119:233–242
Junge W, Nelson N (2015) ATP synthase. Annu Rev Biochem 83:631–657
Junge W, Lill H, Engelbrecht S (1997) ATP synthase: an electrochemical transducer with rotatory mechanics. Trends Biochem Sci 22:420–423
Karavaev VA, Kukushkin AK (1993) Theoretical model of the light and dark stages of photosynthesis: the regulation problem. Biophysics 38:958–975
Kern J, Zouni A, Guskov A, Krauβ N (2009) Lipid in the structure of photosystem I, photosystem II and the cytochrome b6f complex. In: Wada H, Murata N (eds) Lipids in Photosynthesis: Essential and Regulatory Functions. Springer, Dordrecht, pp 203–242
Kirchhoff H (2008) Significance of protein crowding, order and mobility for photosynthetic membrane functions. Biochem Soc Trans 36:967–970
Kirchhoff H (2014) Diffusion of molecules and macromolecules in thylakoid membranes. Biochim Biophys Acta 1837:495–502
Kirchhoff H, Horstmann S, Weis E (2000) Control of the photosynthetic electron transport by PQ diffusion microdomains in thylakoids of higher plants. Biochim Biophys Acta 1459:148–168
Kirchhoff H, Hall C, Wood M, Herbstová M, Tsabari O, Nevo R, Charuvi D, Shimoni E, Reich Z (2011) Dynamic control of protein diffusion within the granal thylakoid lumen. Proc Natl Acad Sci USA 108:20248–20253
Kraayenhof R, Katan MB, Grunwald T (1971) The effect of temperature on energy-linked functions in chloroplasts. FEBS Lett 19:5–10
Kramer DM, Sacksteder CA, Cruz JA (1999) How acidic is the lumen? Photosynth Res 60:151–163
Kukushkin AK, Tikhonov AN (1988) Lectures on Biophysics of Photosynthesis in Higher Plants. Moscow University Press, Moscow (in Russian)
Kumamoto J, Raison JK, Lyons JM (1971) Temperature “breaks” in Arrhenius plots: a thermodynamic consequence of a phase change. J Theor Biol 31:47–51
Kuvykin IV, Vershubskii AV, Ptushenko VV, Tikhonov AN (2008) Oxygen as an alternative electron acceptor in the photosynthetic electron transport chain of C3 plants. Biochemistry (Moscow) 73:1063–1075
Laisk A, Nedbal N, Govindjee (eds) (2009) Photosynthesis in silico. Understanding complexity from molecules to ecosystems. Springer, Dordrecht
Lazár D, Schansker G (2009) Models of chlorophyll a fluorescence transients. In: Laisk A, Nedbal L, Govindjee S (eds) Photosynthesis in silico: understanding complexity from molecules to ecosystems. Advances in photosynthesis and respiration, vol 29. Springer, Dordrecht, pp 85–123
Lee AJ (1977) Lipid phase transitions and phase diagrams. I. Lipid phase transitions. Biochim Biophys Acta 472:237–281
Li Z, Wakao S, Fischer BB, Niyogi KK (2009) Sensing and responding to excess light. Annu Rev Plant Biol 60:239–260
Ligeza A, Tikhonov AN, Hyde JS, Subczynski WK (1998) Oxygen permeability of thylakoid membranes: electron paramagnetic resonance spin labeling study. Biochim Biophys Acta 1365:453–463
Link TA (1997) The role of the “Rieske” iron sulfur protein in the hydroquinone oxidation (Qp) site of the cytochrome bc1 complex: the “proton-gated affinity change” mechanism. FEBS Lett 412:257–264
Los DA, Murata N (2004) Membrane fluidity and its role in the perception of environmental signals. Biochim Biophys Acta 1666:142–157
Los DA, Mironov KS, Allakhverdiev SI (2013) Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth Res 116:489–509
Lubitz W, Chrysina M, Cox N (2019) Water oxidation in photosystem II. Photosynth Res 142:105–125
Lutova MI, Tikhonov AN (1983) Aftereffect of high temperature on photosynthesis and electron transport in wheat leaves. Biophysics 28:284–287
Lutova MI, Tikhonov AN (1988) Comparative study of temperature effects on the mobility of lipid- soluble spin label in thylakoid membranes of melon and cucumber chloroplasts. Biophysics 33:460–464
Luzikov VN, Novikova LA, Tikhonov AN, Zubatov AS (1983) Correlation between the rate of proteolysis of mitochondrial translation products and fluidity of the mitochondrial inner membrane of Saccharomices cervisiae. Biochem J 214:785–794
Luzikov VN, Novikova LA, Zubatov AS, Tikhonov AN (1984) Physical state of the mitochondrial inner membrane as a factor controlling the proteolysis of mitochondrial translational products in yeasts. Biochim Biophys Acta 775:22–30
Maksimov EG, Mironov KS, Trofimova MS, Nechaeva NL, Todorenko DA, Klementiev KE, Tsoraev GV, Tyutyaev EV, Zorina AA, Feduraev PV, Allakhverdiev SI, Paschenko VZ, Los DA (2017) Membrane fluidity controls redox-regulated cold stress responses in cyanobacteria. Photosynth Res 133:215–223
Malenkova IV, Kuprin SP, Davidov RM, Blumenfeld LA (1982) pH-jump-induced ADP phosphorylation in mitochondria. Biochim Biophys Acts 682:179–183
Mamedov M, Govindjee Nadtochenko V, Semenov A (2015) Primary electron transfer processes in photosynthetic reaction centers from oxygenic organisms. Photosynth Res 125:51–63
Margolis LB, Tikhonov AN, Vasilieva EYu (1980) Platelet adhesion to fluid and solid phospholipid membranes. Cell 19:189–194
McConnell HM (1976) Molecular motion in biological membranes. Spin labeling: theory and applications. Academic Press, New York, pp 525–561
Melnichenko NA, Koltunov AM, Vyskrebentsev AS, Bazhanov AV (2008) The temperature dependence of the solubility of oxygen in sea water according to the pulsed NMR data. Russ J Phys Chem 82:746–752
Meyer B, Schlodder E, Dekker JP, Witt HT (1989) O2 evolution and Chl a +II (P-680+) nanosecond reduction kinetics in single flashes as a function of pH. Biochim Biophys Acta 974:36–43
Milanovsky GE, Petrova AA, Cherepanov DA, Semenov AY (2017) Kinetic modeling of electron transfer reactions in photosystem I complexes of various structures with substituted quinone acceptors. Photosynth Res 133:185–199
Mitchell P (1976) Possible molecular mechanisms of the protonmotive function of cytochrome systems. J Theor Biol 62:327–367
Mizusawa N, Wada H (2012) The role of lipids in photosystem II. Biochim Biophys Acta 1817:194–208
Möbius K, Savitsky A (2009) High-field EPR spectroscopy on proteins and their model systems: characterization of transient paramagnetic states. RSC Publishing, London
Moon BY, Higashi S, Gombos Z, Murata N (1995) Unsaturation of the membrane lipids of chloroplasts stabilizes the photosynthetic machinery against low-temperature photoinhibition in transgenic tobacco plants. Proc Natl Acad Sci USA 92:6219–6223
Morales A, Yin X, Harbinson J, Driever SM, Molenaar J, Kramer DM, Struik P (2018) In silico analysis of the regulation of the photosynthetic electron transport chain in C3 plants. Plant Physiol 176:1247–1261
Moser CC, Keske JM, Warncke K, Farid RS, Dutton PL (1992) Nature of biological electron transfer. Nature 355:796–802
Murata N, Fork DC (1977) Temperature dependence of the light-induced spectral shift of carotenoids in Cvanidium caldarium and higher plant leaves. Evidence for an effect of the physical phase of chloroplast membrane lipids on the permeability of the membranes to ions. Biochim Biophys Acta 461:365–378
Nagle JF (1987) Theory of passive proton conductance in lipid bilayers. J Bioenerg Biomembr 19:413–426
Nelson N, Yocum CF (2006) Structure and function of photosystems I and II. Annu Rev Plant Biol 57:521–565
Nie GY, Baker NR (1991) Modifications to thylakoid composition during development of maize leaves at low growth temperatures. Plant Physiol 95:184–191
Nievola CC, Carvalho CP, Carvalho V, Rodrigues E (2017) Rapid responses of plants to temperature changes. Temperature 4:371–405
Nishimura M, Ito T, Chance B (1962) Studies on bacterial photophosphorylation III. A sensitive and rapid method of determination of photophosphorylation. Biochim Biophys Acta 59:177–182
Niu Y, Xiang Y (2018) An overview of biomembrane functions in plant responses to high-temperature stress. Front Plant Sci 9:915
Nolan WG (1980) Effect of temperature on electron transport activities of isolated chloroplasts. Plant Physiol 66:234–237
Nolan WG (1981) Effect of temperature on proton efflux from isolated chloroplast thylakoids. Plant Physiol 67:1259–1263
Nolan WG, Smillie RM (1976) Multi-temperature effects on Hill reaction activity of barley chloroplasts. Biochim Biophys Acta 440:461–475
Nolan WG, Smillie RM (1977) Temperature-induced changes in Hill activity of chloroplasts isolated from chilling-sensitive and chilling-resistant plants. Plant Physiol 59:1141–1145
Ogawa S, Shen C, Castillo CLA (1980) NMR study of the cross-membrane pH gradient induced by ATP hydrolysis in mitochondria. Biochim Biophys Acta 590:159–169
Ort DR, Baker NR (2002) A photoprotective role for O2 as an alternative electron sink in photosynthesis? Curr Opin Plant Biol 5:193–198
Osyczka A, Moser CC, Dutton L (2005) Fixing the Q cycle. Trends Biochem Sci 30:176–182
Page CC, Moser CC, Chen X et al (1999) Natural engineering principles of electron tunnelling in biological oxidation-reduction. Nature 402:47–52
Quinn PJ, Williams WP (1978) Plant lipids and their role in membrane function. Progr Biophys Molec Biol 34:107–173
Razeghifard MR, Klughammer C, Pace RJ (1997) Electron paramagnetic resonance kinetic studies of the S states in spinach thylakoids. Biochemistry 36:86–92
Reeves SG, Hall DO, West J (1972) Correlation of the stoichiometry of photophosphorylation with the integrity of isolated spinach chloroplasts. In: Forti G, Avron M, Melandri A (eds) Photosynthesis, two centuries after its discovery by Joseph Priestley. Springer, Dordrecht
Rigoulet M, Leverve X, Fontaine E, Ouhabi R, Guérin B (1998) Quantitative analysis of some mechanisms affecting the yield of oxidative phosphorylation: dependence upon both fluxes and forces. Mol Cell Biochem 184:35–52
Riznichenko GY, Belyaeva NE, Kovalenko IB, Rubin AB (2009) Mathematical and computer modeling of primary photosynthetic processes. Biophys 54:10–22
Romanovsky YuM, Tikhonov AN (2010) Molecular energy transducers of the living cell. Proton ATP synthase: a rotating molecular motor. Phys Usp 53:893–914
Rubin A, Riznichenko G (2014) Mathematical biophysics. Series: biological and medical physics, biomedical engineering, XV
Sanderson DG, Anderson LB, Gross EL (1986) Determination of the redox potential and diffusion coefficient of the protein plastocyanin using optically transparent filar electrodes. Biochim Biophys Acts 852:269–278
Santabarbara S, Redding KE, Rappaport F (2009) Temperature dependence of the reduction of P +700 by tightly bound plastocyanin in vivo. Biochemistry 48:10457–10466
Sarcina M, Murata N, Tobin MJ, Mullineaux CW (2003) Lipid diffusion in the thylakoid membranes of the cyanobacterium Synechococcus sp.: effect of fatty acid desaturation. FEBS Lett 553:295–298
Sawada S, Miyachi S (1974) Effects of growth temperature on photosynthetic carbon metabolism in green plants I. Photosynthetic activities of various plants acclimatized to varied temperatures. Plant Cell Physiol 15:111–120
Schneider AR, Geissler PL (2013) Coexistance of fluid and crystalline phases of proteins in photosynthetic membranes. Biophys J 105:1161–1170
Schuurmans JJ, Kraayenhof R (1983) Energy-regulated functional transitions of chloroplast ATPase. Photochem Photobiol 37:85–91
Seelert H, Poetsch A, Dencher NA, Engel A, Stahlberg H, Müller DJ (2000) Structural biology. Proton-powered turbine of a plant motor. Nature 405:418–419
Setif PQY, Bottin H (1994) Laser flash absorption spectroscopy study of ferredoxin reduction by Photosystem I in Synechocystis sp. PCC 6803: Evidence for submicrosecond and microsecond kinetics. Biochemistry 33:8495–8504
Shelaev IV, Gostev FE, Mamedov MD, Sarkisov OM, Nadtochenko VA, Shuvalov VA, Semenov AYu (2010) Femtosecond primary charge separation in photosystem I. Biochim Biophys Acta 1797:1410–1420
Shneyour A, Raison JK, Smillie RM (1973) The effect of temperature on the rate of photosynthetic electron transfer in chloroplasts of chilling-sensitive and chilling-resistant plants. Biochim Biophys Acta 292:152–161
Sigfridsson K (1998) Plastocyanin, an electron transfer protein. Photosynth Res 57:1–28
Siggel U (1976) The function of plastoquinone as electron and proton carrier in photosynthesis. Bioelectrochem Bioenerg 3:302–318
Staehelin LA (2003) Chloroplast structure: from chlorophyll granules to supra-molecular architecture of thylakoid membranes. Photosynth Res 76:185–196
Stiehl HH, Witt HT (1969) Quantitative treatment of the function of plastoquinone in photosynthesis. Z Naturforsch B 24:1588–1598
Stirbet A, Govindjee G (2016) The slow phase of chlorophyll a fluorescence induction in silico: origin of the S-M fluorescence rise. Photosynth Res 130:193–213
Stirbet A, Riznichenko GY, Rubin AB, Govindjee G (2014) Modeling chlorophyll a fluorescence transient: relation to photosynthesis. Biochemistry (Moscow) 79:291–323
Stirbet A, Lazár D, Guo Y, Govindjee G (2019) Photosynthesis: basics, history, and modeling. Ann Bot. https://doi.org/10.1093/aob/mcz171
Strand DD, Fisher N, Kramer DM (2016) Distinct energetics and regulatory functions of the two major cyclic electron flow pathways in chloroplasts. In: Kirchhoff H (ed) Chloroplasts: current research and future trends. Caister Academic Press, Norfolk, pp 89–100
Suslichenko IS, Tikhonov AN (2019) Photo-reducible plastoquinone pools in chloroplasts of Tradescentia plants acclimated to high and low light. FEBS Lett 593:788–798
Tietz S, Puthiyaveetil S, Enlow HM, Yarbrough R, Wood M, Semchonok DA, Lowry T, Li Z, Jahns P, Boekema EJ, Lenhert S, Niyogi KK, Kirchhoff H (2015) Functional implications of Photosystem II crystal formation in photosynthetic membranes. J Biol Chem 290:14091–14106
Tikhonov AN (2012) Energetic and regulatory role of proton potential in chloroplasts. Biochemistry (Moscow) 77:956–974
Tikhonov AN (2013) pH-Dependent regulation of electron transport and ATP synthesis in chloroplasts. Photosynth Res 116:511–534
Tikhonov AN (2014) The cytochrome b6f complex at the crossroad of photosynthetic electron transport pathways. Plant Physiol Biochem 81:163–183
Tikhonov AN (2015) Induction events and short-term regulation of electron transport in chloroplasts: an overview. Photosynth Res 125:65–94
Tikhonov AN (2016) Modeling electron and proton transport in chloroplasts. In: Kirchhoff H (ed) Chloroplasts: current research and future trends. Caister Academic Press, Norfolk, pp 101–134
Tikhonov AN (2017) Photosynthetic electron and proton transport in chloroplasts: EPR study of ΔpH generation, an overview. Cell Biochem Biophys 75:421–432
Tikhonov AN (2018) The cytochrome b6f complex: biophysical aspects of its functioning in chloroplasts. In: Harris JR, Boekema EJ (eds) Membrane protein complexes: structure and function, subcellular biochemistry, vol 87. Springer, Singapore, pp 287–328
Tikhonov AN (2020) Structure-function relationships in chloroplasts: EPR study of temperature-dependent regulation of photosynthesis, an overview. In: JR Shen, K Satoh, SI Allakhverdiev (eds) Photosynthesis: molecular approaches to solar energy conversion.
Tikhonov AN, Blumenfeld LA (1985) Hydrogen ions concentration in subcellular systems: physical meaning and the methods for determination. 30:527–537
Tikhonov AN, Subczynski WK (2005) Application of spin labels to membrane bioenergetics (photosynthetic systems of higher plants), chapter 8. In: Eaton SS, Eaton GR, Berliner LJ (eds) Biological magnetic resonance, vol 23. Biomedical EPR—part A: free radicals, metals, medicine, and physiology. Kluwer Academic Publishers, New York, pp 147–194
Tikhonov AN, Vershubskii AV (2014) Computer modeling of electron and proton transport in chloroplasts. BioSystems 121:1–21
Tikhonov AN, Vershubskii AV (2017) Connectivity between electron transport complexes and modulation of photosystem II activity in chloroplasts. Photosynth Res 133:103–114
Tikhonov AN, Khomutov GB, Ruuge EK (1980) Electron spin resonance study of electron transport in photosynthetic systems. IX. Temperature dependence of the kinetics of P700 redox transients in bean chloroplasts induced by flashes with different duration. Mol Biol (Moscow) 14:157–172
Tikhonov AN, Khomutov GB, Ruuge EK, Blumenfeld LA (1981) Electron transport control in chloroplasts. Effects of photosynthetic control monitored by the intrathylakoid pH. Biochem Biophys Acta 637:321–333
Tikhonov AN, Timoshin AA, Blumenfeld LA (1983) Electron transport kinetics, proton transfer, photophosphorylation in chloroplasts, and their relation to thermo-induced structural changes in the thylakoid membrane. Mol Biol (Moscow) 17:1236–1248
Tikhonov AN, Khomutov GB, Ruuge EK (1984) Electron transport control in chloroplasts. Effects of magnesium ions on the electron flow between two photosystems. Photobiochem Photobiophys 8:261–269
Tikhonov AN, Agafonov RV, Grigor’ev IA, Kirilyuk IA, Ptushenko VV, Trubitsin BV (2008) Spin-probes designed for measuring the intrathylakoid pH in chloroplasts. Biochim BiophysActa 1777:285–294
Timoshin AA, Tikhonov AN, Blumenfeld LA (1984) Thermoinduced structural changes in ATP-synthase as a factor of energy transduction control in chloroplasts. Biophysics 29:338–340
Torres-Pereira J, Mehlhorn R, Keith AD, Packer L (1974) Changes in membrane lipid structure of illuminated chloroplasts—studies with spin labeled and freeze-fractured membranes. Arch Biochem Biophys 160:90–99
Tremmel IG, Kirchhoff H, Weis E, Farquhar GD (2003) Dependence of plastoquinol diffusion on the shape, size, and density of integral thylakoid proteins. Biochim Biophys Acta 1603:97–109
Trubitsin BV, Tikhonov AN (2003) Determination of a transmembrane pH difference in chloroplasts with a spin label tempamine. J Magn Reson 163:257–269
Turina P, Petersen J, Gräber P (2016) Thermodynamics of proton transport coupled ATP synthesis. Biochim Biophys Acta 1857:653–664
Ustynyuk LYu, Tikhonov AN (2018) The cytochrome b6f complex: DFT modeling of the first step of plastoquinol oxidation by the iron-sulfur protein. J Organomet Chem 867:290–299
Varco-Merth B, Fromme R, Wang M, Fromme P (2008) Crystallization of the c14-rotor of the chloroplast ATP synthase reveals that it contains pigments. Biochim Biophys Acta 1777:605–612
Vershubskii AV, Tikhonov AN (2020) pH-Dependent regulation of electron and proton transport in chloroplasts in situ and in silico. Biochem (Moscow) Suppl Ser A 14:154–165
Vershubskii AV, Kuvykin IV, Priklonsky VI, Tikhonov AN (2011) Functional and topological aspects of pH-dependent regulation of electron and proton transport in chloroplasts in silico. Biosystems 103:164–179
Vershubskii AV, Trubitsin BV, Priklonskii VI, Tikhonov AN (2017) Lateral heterogeneity of the proton potential along the thylakoid membranes of chloroplasts. Biochim Biophys Acta 1859:388–401
Vershubskii AV, Nevyantsev SM, Tikhonov AN (2018) Modeling of electron and proton transport in chloroplast membranes with regard to thioredoxin-dependent activation of the Calvin-Benson cycle and ATP synthase. Biochem(Moscow) Suppl Ser A 12:287–302
Vlasov AV, Kovalev KV, Marx S-H et al (2019) Unusual features of the c-ring of F1FO ATP synthases. Sci Rep 9(1):1–11
Vollmar M, Schlieper D, Winn M, Büchner C, Groth G (2009) Structure of the c14 rotor ring of the proton translocating chloroplast ATP synthase. J Biol Chem 284:18228–18235
Wada H, Murata N (eds) (2009) Lipids in Photosynthesis: Essential and Regulatory Functions. Springer, Dordrecht
Walker JE (2013) The ATP synthase: the understood, the uncertain and the unknown. Biochem Soc Trans 41:1–16
Wallis JG, Browse J (2002) Mutants of Arabidopsis reveal many roles for membrane lipids. Prog Lipid Res 41:254–278
Witt HT (1979) Energy conversion in the functional membrane of photosynthesis. Analysis by light pulse and electric pulse methods. Biochim Biophys Acta 505:355–427
Yamamoto Y (2016) Quality control of photosystem II: the mechanisms for avoidance and tolerance of light and heat stresses are closely linked to membrane fluidity of the thylakoids. Front Plant Sci 7:1136. https://doi.org/10.3389/fpls.2016.01136
Yamamoto Y, Nishimura M (1976) Characteristics of light-induced H+ transport in spinach chloroplasts at lower temperatures I. Relationship between H+ transport and physical changes of the microenvironment in chloroplast membranes. Plant Cell Physiol 17:11–16
Yamamoto Y, Ford RC, Barber J (1981) Relationship between thylakoid membrane fluidity and the functioning of pea chloroplasts. Effect of cholesteryl hemisuccinate. Plant Physiol 67:1069–1072
Yamori W, Hikosaka K, Way DA (2014) Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation ant temperature adaptation. Photosynth Res 119:101–117
Yan J, Cramer WA (2003) Functional insensitivity of the cytochrome b6f complex to structure changes in the hinge region of the Rieske iron–sulfur protein. J Biol Chem 278:20926–20933
Yan K, Chen P, Shao H, Shao C, Zhao S, Brestic M (2013) Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS ONE 8(5):e62100
Zaks J, Amarnath K, Kramer DM, Niyogi KK, Fleming GR (2012) A kinetic model of rapidly reversible nonphotochemical quenching. Proc Natl Acad Sci USA 109:15757–15762
Zhou Y, vom Dorp K, Dörman P, Hölzl G (2016) Chloroplast lipids. In: Kirchhoff H (ed) Chloroplasts: current research and future trends. Caister Academic Press, Norfolk, pp 1–24
Zhu XG, Wang Y, Ort DR, Long SP (2013) e-Photosynthesis: a comprehensive dynamic mechanistic model of C3 photosynthesis: from light capture to sucrose synthesis. Plant Cell Environ 36:1711–1727
Acknowledgements
We thank Dr. V.I. Priklonskii for the adjustment of the computer program in order to parameterize the constants of temperature dependences of the partial reactions of electron transport. Experimental studies on bean chloroplasts, used in this work for parametrization of the model parameters, were performed earlier by one of the authors in collaboration with Professor E.K. Ruuge, Professor G.B. Khomutov, and Dr. A.A. Timoshin. We are greatly thankful to all our colleagues and collaborators.
Funding
This work was partly supported by the Russian Foundation for Basic Research (Grant 18-04-00214).
Author information
Authors and Affiliations
Contributions
ANT: design and supervision of the work, data processing, writing of the manuscript. AVV: computer calculations, data processing, discussion of results, preparation of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1: equations and rate constants of the model
Equations (A1–A7) represent the system of ordinary differential equations (ODE), which describe the redox transitions of electron carriers (variables [Fd], [\( {\text{P}}_{ 7 0 0}^{ + } \)], [\( {\text{P}}_{ 6 8 0}^{ + } \)], [Pc], [PQ]), the acidification of the thylakoid lumen ([\( {\text{H}}_{\text{in}}^{ + } \)]), and the yield of ATP (variable [ATP]).
Here, the function \( k_{\text{Q}} ( [ {\text{PQ],[Pc],[H}}_{\text{in}}^{ + } ] ,T ) \) describes the electron transfer from PQH2 to Pc via the Cyt b6f complex [see above Eqs. (1) and (2)]. ΔpH is the trans-thylakoid pH difference, ΔpH = pHout – pHin. We assume that pHout to be constant, pHout = 8, due to sufficiently high buffer capacity of the outer medium. [ADN]0 is the total concentration of ADP and ATP. The model parameters α and β in Eqs. (A6) and (A7) are determined by the rate constants of the proton exchange with the proton-accepting groups of the ATP synthase (see Fig. 3 and the explanations below). Formulating Eqs. (A1–A7), we assume the following stoichiometry between the electron transport and ATP synthase complexes: [PSI]/[PSII]/[b6f]/[CF0-CF1] = 1/1/1/1. The relative capacity of the photo-reducible PQ pool, Fd, and Pc was taken as [PQ]0/[PSI] = 10, [Fd]0/[PSI] = 3, and [Pc]0/[PSI] = 1.5, respectively.
The model parameters L1 and L2 describe the numbers of light quanta per unit time exciting P700 and P680, respectively. Parameter m expresses the stoichiometry of proton transfer through the ATP synthase, H+/ATP; m = n/3 = 14/3 is the stoichiometry ratio, the ratio between a number n = 14 of subunits c in the cn-ring to three ATP molecules formed per one turn (360o) of the membrane rotor cn (Seelert et al. 2000; Vollmar et al. 2009). Constants marked with a subscript “0” are the maximal concentrations of the relevant variables. Constants \( k_{{{\text{P}}_{ 6 8 0} }}^{{}} \), \( k_{{{\text{P}}_{ 7 0 0} }}^{{}} \), kQ, kPc, and kMeh are the effective rate constants of the reactions shown in Fig. 1. Constant kADP governs the effective rate of ATP hydrolysis. The function kQ, which characterizes the oxidation of PQH2, depends on pHin (for details see Eqs. 1 and 2; Dubinskii and Tikhonov 1997; Vershubskii et al. 2011). The model parameters KM and Bin, characterize the buffer properties of the system. Here, KM is the equilibrium constant for the reaction of proton binding by buffer groups inside of thylakoids; the model parameter Bin is the concentration of these buffer groups. In this work, we assume the stoichiometric ratio Bin/PSI = 100 (for details, see Vershubskii et al. 2011). Note that parameters KM and Bin (the left side of Eq. A7) can influence the time-course of the system response to switching the actinic light on. The steady-state levels of all the variables of the model, however, are independent on the KM and Bin values (Dubinskii and Tikhonov 1997).
The formulation of the system of differential equations and the choice of the apparent rate constants have been considered in details in our previous works (Vershubskii et al. 2011, 2018). The rate constants for key stages of electron flow and proton transport were determined by fitting the respective experimental and simulated kinetic curves. In particular, effective rate constant for PQH2 oxidation by the Cyt b6f complex was derived by comparing calculated and experimental plots for the rates of the post-illumination reduction of \( {\text{P}}_{ 7 0 0}^{ + } \) at different pHin ( “Appendix 3”, Fig. 13). Effective rate constants of the transmembrane proton transport coupled to ATP synthesis and passive leak of protons through the thylakoid membrane were determined by the comparison of calculated and experimental data on the light-induced acidification of the thylakoid lumen (for details, see “Appendix 2”; Dubinskii and Tikhonov 1995). The values of the rate constants, which characterize different stages of the electron transfer along the ETC, from the water-splitting complex of PSII to PSI acceptors, were chosen on the basis of the literature data on the kinetics of partial reactions of electron transport in different segments of the chloroplast ETC as described above. The characteristic times of electron transfer reactions are given in Table 1.
Appendix 2: proton transport and ATP synthesis in the model
There are indications that the proton conductivity of a lipid bilayer is determined mainly by the presence of acidic groups in the membrane (Deamer 1987; Gutknecht 1987; Nagle 1987). In this work, the equations for the active (JATP) and passive (Jpass) fluxes of protons were derived from a simple model based on the assumption that the processes of the transmembrane transfer of protons occur through the acidic groups bound either to the ATP synthase or buried inside the thylakoid membrane, respectively (Dubinskii and Tikhonov 1995). According to the model considered in our work, the proton transfer through the CFo segment of the ATP synthase includes stages of protonation and deprotonation of carboxyl groups of the cm-ring: –COO– + \( {\text{H}}_{\text{in}}^{ + } \) → –COOH → –COO– + \( {\text{H}}_{\text{out}}^{ + } \). Concerning the passive trans-thylakoid transport of H+ ions, we assume that the proton first binds to the intramembrane proton-accepting group and then dissociates into the stroma. The efficient rate constants of the proton exchange with the proton-accepting group are indicated in Fig. 3b. The rate constants of the direct and reverse reactions are related by the ratio \( k_{1}^{\text{in}} /k_{ - 1}^{\text{in}} = K_{\text{M1}} \) and \( k_{2}^{\text{out}} /k_{ - 2}^{\text{out}} = K_{\text{M2}} \), where KM1 and KM2 are the effective constants of the proton equilibrium for the buffer group –COO− and hydrogen ions inside and outside of the thylakoid, respectively. It is reasonable to assume that, for the proton-accepting groups –COO− fixed in the membrane and involved in the passive transfer of protons across the membrane, the equality KM1 = KM2 ≡ KM must be true. Fitting of the rate constant parameters related to proton transfer through the membrane acidic groups has been performed by means of the comparison of calculated and experimental data on the light-induced uptake of protons by the chloroplasts (Dubinskii and Tikhonov 1995). For the ΔpH-driven flux of protons through the ATP synthase, JATP, coupled to ATP synthesis, we used the following relationship:
where ΔpH = pHout − pHin is the driving force for the operation of the ATP synthase (for details, see Tikhonov and Vershubskii 2014). Coefficients α and β are the model parameters, the values of which are determined by pKA of the acidic group –COO− of the cn-ring of the ATP synthase and the values of the efficient rate constants k1 and k2 characterizing proton transport to -COO− from the lumen and stroma, respectively; \( \alpha = 10^{ - \text{p}K_\text{A}} \left( {1 + \beta } \right) \), \( \beta = k_{ 2}^{\text{out}} /k_{1}^{\text{in}} \). In our calculations we used the model parameters pKA = 7.3 and β = 20, which values have been chosen on the basis of our previous works (Vershubskii et al. 2011, 2017, 2018; Vershubskii and Tikhonov 2020). Formula (A8) provides a sigmoid dependence of the ATP synthesis rate versus the proton-motive force ΔpH (Fig. 3c), which is typical of experimental force–flux relationships in chloroplasts (Turina et al. 2016). Note that variations of the model parameters α and β influence markedly a threshold ΔpHth, above which value the ATP synthase efficiently produces ATP. Numerical values of the model parameters, related to the trans-thylakoid proton transfer, were determined by fitting theoretical curves to relevant experimental dependences of the light-induced acidification of the lumen at different values of external pHout. Fitting of the rate constants \( k_{ 1}^{\text{in}} \), \( k_{ - 1}^{\text{in}} \), \( k_{ 2}^{\text{in}} \), and \( k_{ - 2}^{\text{in}} \) was performed earlier (Dubinskii and Tikhonov 1995).
The model parameter \( k_{{{\text{H}}^{ + } }} \), which stands in the right side of Eq. A7, determines the passive efflux of protons (Jpass) from the lumen to the outer space: Jpass = \( k_{{{\text{H}}^{ + } }} (T )\cdot ( [ {\text{H}}_{\text{in}}^{ + } ] { } - {\text{ [H}}_{\text{out}}^{ + } ] ) \). The \( k_{{{\text{H}}^{ + } }} (T ) \) values were chosen by means of fitting calculated values of Jpass to experimentally measured proton fluxes determined for chloroplasts in the metabolic state 4 (compare experimental and theoretical data in Fig. 4c).
It is well-known fact that the ATP synthase activity is controlled by the redox status of chloroplasts. The light-induced activation of the ATP synthase is associated with the reduction of the thiol groups in the subunit γ, rotating together with the cn-ring (Bakker-Grunwald and van Dam 1974; Bald et al. 2001). It is conceivable that the reduction of these groups may be mediated through the pigments found in the central cavity of the c14-rotor (Varco-Merth et al. 2008; Vlasov et al. 2019). In the current work, however, we ignored this effect, because we compared experimental and theoretical data on the initial phase of the light-induced ATP synthesis (during 10-s illumination), when the linear growth of ATP concentration was not affected by the light-induced modulation of the chloroplast ATP synthase. According to our previous measurements, the light-induced activation of the ATP synthase was observed after 2 min of chloroplast illumination in the presence of methylviologen (data not shown).
Appendix 3: materials and experimental methods
Plant material and preparation of chloroplasts
Class B chloroplasts were isolated from greenhouse bean leaves (Vicia faba, 2–3 weak old) as described by Tikhonov et al. (1981). Chloroplasts were suspended at a final concentration of 2–3 mg chlorophyll/ml in the medium containing 0.2 M sucrose, 2 mM MgCI2, and 10 mM Tricine-NaOH buffer (pH between 6.5 and 9.5) or Mes-HCl buffer (pH between 4.5 and 6.5). For measurements of chloroplast activity at different pH of the chloroplast suspension, we also used the medium which contained 0.2 M sucrose, 2 mM MgC12, 10 mM phosphate-citrate buffer, and 4 mM Mg-ADP. 20 μM methylviologen was used as a mediator of electron transfer from PSI to molecular oxygen. The pH value of the suspending medium was checked with a Radiometer glass electrode GK2321C.
In this work, we refer to experimental results described in earlier works of our group (Tikhonov et al. 1980, 1981, 1983, 1984; Timoshin et al. 1984; Kukushkin and Tikhonov 1988; Tikhonov and Subczynski 2005). The batches of chloroplasts used in these works were isolated from bean leaves of different harvests, including plants grown under variable experimental conditions (different seasons and concomitant changes in environmental conditions). In general, plants were cultivated at growth temperatures in the range 18–32 °C, depending on the season. We found that variability in the plant cultivation conditions could cause somewhat different temperature dependences of photosynthetic processes in isolated bean chloroplasts (compare, for example, panels c, d, and e in Fig. 9). Variability of this kind proved useful for analyzing the structure–function relationships in chloroplasts. In particular, statistically significant coincidence of the peculiar temperatures of the “structural” (membrane fluidity) and “functional” (ATP synthesis) characteristics was observed for each individual batch of chloroplasts (Fig. 9). Inflexion points in temperature dependences of the plots of “structural” and “functional” parameters coincided with a sufficiently high precision (±1 °C). In the meantime, the harvest-depending scattering of these peculiar points was more significant (for example, in the range 25–33 °C, Fig. 9). This observation allowed us to suggest that the temperature dependences of electron and proton transport processes were controlled by the physical state of thylakoid membranes.
EPR measurements of PSII activity and the intersystem electron transfer
The redox transients of P700 were monitored by measuring the light-induced changes in the amplitude of the EPR signal from \( {\text{P}}_{700}^{ + } \) (Tikhonov et al. 1980, 1981; Tikhonov 2015). The EPR measurements of \( {\text{P}}_{700}^{ + } \) were performed with a Varian EPR spectrometer (model E-4) at 4 G modulation amplitude and 10 mW microwave power. Far-red background illumination (interference filter SIF707, Karl Zeiss Jena; λmax = 707 nm, Δλl/2 = 5 nm) was applied to provide the re-oxidation of the ETC between PSII and PSI. The intensity of this light, provided by a 150 W incandescent lamp equipped with a water filter and focusing lens, was adjusted to reach maximal level of P700 oxidation. A similar light source without the interference filter (white light) was used for efficient excitation of both photosystems. Two kinds of white light pulses of were used to test PSII activity: (i) short flashes (τl/2 = 7 μs) of saturating intensity, and (ii) long flashes (τl/2 = 750 μs) were applied for multiple operation of P680. The energy released in the discharge circuit was 10 and 100 J, respectively (Tikhonov et al. 1980).
Figure 12a shows a simplified diagram illustrating electron transfer from PSII and O2, the terminal acceptor of electrons donated by PSI. Illumination of chloroplasts by the far-red (or white) light induces generation of the EPR signal from \( {\text{P}}_{700}^{ + } \) shown in Fig. 12b. After sudden shutdown of white light (WL), \( {\text{P}}_{700}^{ + } \) rapidly reduces due to electrons donated by reduced PQH2 molecules (Fig. 12c). The half-time of \( {\text{P}}_{700}^{ + } \) decay (parameter τ1/2) characterizes the rate of electron transfer from PQH2 to \( {\text{P}}_{700}^{ + } \).

Modified figures adopted from (Tikhonov et al. 1984)
Simplified diagram illustrating electron transfer from PSII and O2, the terminal acceptor of electrons donated by PSI (panel a); EPR signals of bean chloroplasts in the dark and during illumination with the far-red light, λmax = 707 nm, as indicated (panel b); the post-illumination kinetics of \( {\text{P}}_{700}^{ + } \) reduction in bean chloroplasts pre-illuminated by continuous white light (panel c)
Figure 13 shows experimental and theoretical dependencies of the half-time of \( {\text{P}}_{700}^{ + } \) reduction after switching off the white light on the intra-thylakoid pHin. Open symbols, experimental data; filled symbols, calculated data. Experimental points were obtained for the suspension of uncoupled chloroplasts (on the basis of results published by Tikhonov et al. (1980)).

Experimental and theoretical dependencies of the half-time of \( {\text{P}}_{700}^{ + } \) reduction after switching off the white light on the intra-thylakoid pHin. Open symbols, experimental data; filled symbols, calculated data. Experimental points were obtained for the suspension of uncoupled chloroplasts (on the basis of results published in Tikhonov et al. 1984)
Figure 14a shows the time-course of P700 redox changes in aerated suspension of bean chloroplasts. Illumination of chloroplasts by the far-red light (λ707), absorbed predominantly by PSI, induced oxidation of P700. Application of a short saturating pulse (τ1/2 = 7 µs) induced the reduction of \( {\text{P}}_{700}^{ + } \) due to the injection of electrons from PSII to the intersystem ETC. The reduction of \( {\text{P}}_{700}^{ + } \) is followed by the re-oxidation of P700 due to the action of the continuous far-red light. The area W1 over the kinetic curve can serve as a measure of PSII photochemical activity: in response to a short flash, each PSII donates one electron (on an average). In this case, the Mn4Ca cluster is oxidized by one electron and one electron is donated to the intersystem ETC (Cardona et al. 2012). After the action of a prolonged flash (τ1/2 = 750 µs), the area over the kinetic curve (parameter W2) increases. This occurs due to a multiple charge separations in PSII and donation of several electrons to the PQ pool (Tikhonov and Vershubskii 2017). Figure 14b shows the temperature dependences of parameters W1 and W2. The ratio f = W2/W1 is determined by the rate of electron transfer from PSII to the PQ pool (Fig. 14c). The temperature of a sample was regulated with the Varian temperature controller.

a The light-induced redox transients of P700 in bean chloroplasts induced by the far-red light (λ707) and pulses of white light of different durations. Parameters W1 and W2 are proportional to the numbers of electrons injected into intersystem electron transport chain (for other details see text and Tikhonov et al. 1980). b Temperature dependences of parameters W1 and W2 shown in the panel a. c Temperature dependence of the ratio f = W2(T)/W1(T), which determines a number of electrons donated by PSII during the action of the long flash
Potentiometry methods of assaying the chloroplasts activity
The rate of pseudocyclic electron flow JFd–O2 (H2O → PSII → PSI → MV → O2; the so-called “water-water” cycle; Asada 1999) was determined by measuring the O2 uptake (O2 + e− → O •−2 ) in an aerated suspension of chloroplasts, using a laboratory-made Clark-type electrode. In this case, the superoxide and catalase activities of chloroplasts were inhibited by the addition of small amounts of NaN3 as described earlier (Timoshin et al. 1984).
The rate of ATP formation (VATP) was routinely measured by the potentiometry method (Nishimura et al. 1962; Timoshin et al. 1984). All controls including that of adenylate kinase activity were properly made. Adequacy of potentiometric measurements of photophosphorylation (ADP + Pi → ATP + H2O) was verified independently by the use of enzymatic (Malenkova et al. 1982) and modified luciferin-luciferase (Ataullakhanov and Pichugin 1981) methods. The rate of ATP hydrolysis was also determined by measuring changes in the 31P NMR spectra of ATP, ADP, and Pi in bean chloroplast suspension according to (Ogawa et al. 1980). Results of this study, performed together with professor E.K. Ruuge, will be published elsewhere.
Having measured, under the same experimental conditions, the rates of ATP synthesis (VATP) and electron transport (JFd–O2), we could determine the ratio VATP/JFd–O2, which characterizes the efficiency of coupling of electron transport and ATP synthesis (often termed as the so-called ratio “P/2e”; Ivanov 1993). Figure 15 reproduces the temperature dependence of the ratio P/2e that was determined earlier in our work (Timoshin et al. 1984). As one can see, the ratio P/2e increases with temperature, reaching a plateau at temperatures above 25 °C.

Modified figures after (Timoshin et al. 1984)
Temperature dependences of parameter P/2e, which denotes the ratio between the rates of photophosphorylation (ADP + Pi → ATP) and pseudocyclic electron flow and ATPase activity of bean chloroplasts (closed and open symbols were obtained on different batches of chloroplasts)
Measurements of ΔpH generation
Transmembrane pH difference (ΔpH) across the thylakoid membrane was measured by three independent methods based on the use of the EPR technique: (1) the determination of the pHin-dependent rate of the intersystem electron transfer from the kinetics of the post-illumination reduction of \( {\text{P}}_{700}^{ + } \) (Tikhonov et al. 1981, 1984), (2) the use of pH-sensitive water-soluble spin probes located in the thylakoid lumen (Tikhonov et al. 2008; Tikhonov 2017; Vershubskii et al. 2017), and (3) the determination of ΔpH in chloroplasts from the partitioning of a water-soluble spin label tempamine (Trubitsin and Tikhonov 2003; Tikhonov 2017). We used experimental data on ΔpH measurements in bean chloroplasts in order to perform the final fitting of the model parameters (see, for example, Vershubskii and Tikhonov 2020).
Spin-labeling study of thermo-induced structural changes in thylakoid membranes
Thermo-induced changes in the lipid domains of the thylakoid membranes were studied with the lipid-soluble spin probes, paramagnetic derivatives of stearic acid, 5-SASL (Fig. 9b), as described in Tikhonov et al. (1980, 1983), Timoshin et al. (1984), Lutova and Tikhonov (1988), Ligeza et al. (1998), and Tikhonov and Subczynski (2005). A small aliquot (1% v/v) of 5-SASL dissolved in ethanol was added to chloroplast suspension. After 10-min incubation of thylakoids with the spin probe, the suspension of spin-labeled thylakoids was used for EPR measurements. The temperature of a sample placed into the cavity of a Varian (E-4) X-band EPR spectrometer was regulated with the Varian temperature controller, with precision up to ± 0.5 °C.
Rights and permissions
About this article

Cite this article
Tikhonov, A.N., Vershubskii, A.V. Temperature-dependent regulation of electron transport and ATP synthesis in chloroplasts in vitro and in silico. Photosynth Res 146, 299–329 (2020). https://doi.org/10.1007/s11120-020-00777-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11120-020-00777-0