
Abstract
The development of high-quality cultivars, with durable disease resistance, is a major objective of apple breeding. The selection procedures of modern breeding programs no longer rely exclusively on phenotypic criteria but include marker-assisted breeding (MAB). Currently, molecular markers linked to several resistance genes and quantitative trait loci (QTLs) are available. In this study, we focused on markers available for resistance breeding against the major diseases scab (Venturia inaequalis), powdery mildew (Podosphaera leucotricha), and fire blight (Erwinia amylovora). One approach proposed to achieve durable resistance is the pyramiding of functionally different resistance genes against the same pathogen. This approach can be complemented with the incorporation of resistance genes against other pathogens. The resulting resistant apple cultivars would contribute considerably to low-input, sustainable, fruit production. Furthermore, apple cultivars can be developed carrying homozygous allele sets of specific resistance genes, and these genotypes can be used as parents for further crosses. Due to the ensured inheritance of the resistance genes to the progeny, MAB for these genes will become superfluous. In this study, we developed elite apple plants which are homozygous for three different scab resistance genes, Rvi6, Rvi2, and Rvi4. Furthermore, these apple selections tested positive for a resistance gene against powdery mildew (Pl1 or Pl2), and the FBF7 QTL from ‘Fiesta’ for enhanced fire blight resistance. Selected progeny plants were tested for their fire blight resistance after artificial shoot inoculation and evaluated for tree and fruit characteristics.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Intensive breeding efforts will be needed to develop high-quality, disease-resistant apple cultivars to address both the ecological concerns associated with the use of pesticides and consumer demands for high-quality fruit. To date, the most frequently used resistance gene in apple breeding has been Rvi6 (alias Vf), a single major gene conferring resistance to apple scab (Venturia inaequalis), derived from Malus floribunda 821. Breakdown of Rvi6 resistance has been observed in some geographical areas in northern Europe (Parisi et al. 2004; Parisi et al. 1993) and has progressed further in recent years. In 1994, race 7, infecting M. floribunda 821, was detected in England (Roberts and Crute 1994). In 1997, the detection of scab on a M. floribunda selection suggested the presence of race 7 in North America for the first time (Beckerman 2009). Gessler and Pertot (2012) concluded that events where Rvi6 resistance has been overcome appear to be rare and are probably due to specific V. inaequalis genotypes that originated outside of Europe or North America and have been introduced into local scab populations. The breakdown of Rvi6 demonstrates that scab problems cannot be solved in a durable and sustainable way with a single major resistance gene. To achieve durable disease resistance, several functionally distinct resistance genes and/or quantitative trait loci (QTLs) against the same pathogen should be combined in a single genotype (MacHardy et al. 2001). These genotypes can be identified and selected using the available molecular markers linked to the genes and QTLs conferring resistance to different pests and diseases (reviewed in Gessler et al. 2006; Hancock et al. 2008; Soriano et al. 2009). However, marker-assisted breeding (MAB) is currently used only in a few apple breeding programs wordwide, probably due to difficulties associated with its implementation. MAB requires both the development and application of a complex knowledge and access to well-equipped genotyping facilities. Furthermore, in some cases, molecular markers are not located sufficiently close to the genes of interest to provide accurate prediction of the phenotype. In the case of simple sequence repeat (SSR) markers, determination of marker allele size is dependent upon laboratory technique which can make selection of the appropriate allele difficult. To avoid this problem, the sizes of SSR alleles in coupling with specific disease resistance genes have been standardized (Patocchi et al. 2009). Another approach to avoid these problems is the development of elite breeding lines with homozygous genes for traits of interest. If used in crosses, such elite lines would transfer the homozygous genes of interest reliably to all progeny. Thus, marker screening of the progeny becomes superfluous for these genes. An alternative approach to using homozygous and pyramided disease resistance elite lines would be MAB with heterozygous, pyramided disease resistance elite lines. In this case, not only the parents of the cross but also all the seedlings have to be analyzed with molecular markers. However, in both approaches, overall breeding efficiency will be influenced by the fruit quality of the selected parents.
A prerequisite for the selection of homozygous genes is the use of an appropriate type of marker. SSR markers are co-dominant; thus, they can differentiate between homozygous and heterozygous states of specific loci. Sequence-characterized amplified region (SCAR) markers are often dominant, and with dominant SCAR markers, homozygous genotypes cannot be differentiated from heterozygous genotypes of the specific locus.
For scab, a number of molecular markers are available which allow qualitative resistance genes such as Rvi2, Rvi4, Rvi5, Rvi6, Rvi11, Rvi13, Rvi15, and other apple scab resistance genes to be detected in resistant parents and their progenies (Galli et al. 2010; Gessler et al. 2006). In this study, we focused on Rvi2, Rvi4, and Rvi6. Different defense reaction symptoms are observed in genotypes carrying these three different apple scab resistance genes. This suggests that their resistance mechanisms may also be different. The resistance gene Rvi4 (Vh4) from Russian apple R12740-7A (Malus Mill. sp.) elicits a classic hypersensitive response visible as small pinpoint pits which can be seen on foliar tissue as early as 3 days after inoculation (DAI) (Bus et al. 2000). Rvi2 (Vh2), also from R12740-7A, causes a more delayed response which occurs after about 4 to 6 DAI, visible as stellate necrosis (Bus et al. 2005). In contrast, R-gene Rvi6, derived from M. floribunda 821, displays a range of symptoms (7-10 DAI): no visible symptoms, pinpoint pits, flecks or necrotic lesions with no sporulation, necrotic lesions with some sparse sporulation, and chlorosis and necrosis associated with restricted sporulation (Chevalier et al. 1991). The three resistance genes Rvi2, Rvi4, and Rvi6, causing three different types of reaction, may be good candidates for gene pyramiding resulting in durable scab resistance.
Similar to scab, qualitative resistance genes Pl1, Pl2, Pld, and Plw conferring resistance to powdery mildew caused by Podosphaera leucotricha have been mapped, and molecular markers linked to them have been identified (Dunemann et al. 2007; James and Evans 2004; Seglias and Gessler 1997; Markussen et al. 1995). In this study, we have focused on Pl1 and Pl2 resistance. Caffier and Parisi (2007) reported that the major resistance gene Pl2, present in genotypes of the fourth generation after introgression of Pl2 from the crab apple Malus zumi, has been overcome. However, pyramiding of these genes may be durable in regions where Pl2 is not overcome.
Fire blight, caused by the bacterium Erwinia amylovora, is a major disease problem in many apple production areas. Although QTLs conferring high levels of resistance have been identified in wild apples such as the ornamental ‘Evereste’, M. floribunda 821 (Durel et al. 2009), and Malus × robusta 5 (Peil et al. 2007), these QTLs cannot readily be used in breeding programs due to the quantity of undesired traits present in wild accessions. To date, the only major QTL identified in commercial cultivars is the FBF7 QTL, identified in ‘Fiesta’ on linkage group 7 (Calenge et al. 2005; Khan et al. 2006). It explained about 40 % of the observed phenotypic variation. Two SCAR markers, AE10-375 and GE-8019, have been developed for this QTL (Khan et al. 2007) and were applied in this study. A pedigree analysis tracked the FBF7 QTL allele of ‘Fiesta’ back to ‘Cox’s Orange’ (Khan et al. 2007).
In this paper, we describe the development of elite breeding lines, homozygous for three scab resistance genes, Rvi6, Rvi2, and Rvi4. Furthermore, some of the lines tested positive for a powdery mildew resistance gene (Pl1 or Pl2) and the FBF7 QTL for enhanced fire blight resistance. The phenotypic scab resistance responses of progenies were compared to their scab resistance genotype. Similarly, progeny plants selected for scab resistance were also evaluated for their fire blight resistance, and the fire blight resistance level was compared to the presence of the FBF7 QTL predicted with markers.
Material and Methods
Plant Material and Crosses
Elite breeding lines were derived from nine full-sib crosses (Table 1) made in spring 2007 by bagging mother trees with insect-proof tissue and hand pollinating the flower pistils with a brush containing pollen of the father selection. Seeds of all crosses were harvested from the fruits in autumn and stratified for 2 months at 2 °C in humid sand. In February 2008, the progeny seedlings were raised in trays (Quick-Pot 35 T (QP52) from gvz_rossat, Otelfingen, Switzerland) in the greenhouse and individually labeled.
Molecular Analysis
Leaf samples were taken from the seedlings at the four leaf stage for DNA extraction prior to V. inaequalis inoculation. Marker screening was accomplished following the protocol of Frey et al. (2004) using multiplex PCRs with fluorescently labeled primers. Data analysis was done with GeneMapper™ Software (Applied Biosystems). The following loci with their corresponding markers were used to select for scab resistance:
-
Locus Rvi6: to assess the presence of Rvi6 located on linkage group 1 (LG1), the SSR marker CH-Vf1 (Vinatzer et al. 2004) was used. In our study, the size of the SSR CH-Vf1 marker allele in coupling with Rvi6 was 166 bp.
-
Locus Rvi2: for the molecular selection of the genotypes carrying Rvi2 (on LG2), the molecular markers OPL19 and CH02b10 were used. According to the revised mapping of Rvi2 done by Jänsch et al. (2015), these two markers flank the resistance gene; CH02b10 was mapped at 1.6 cM upstream, while OPL19 was mapped 1.2 cM downstream Rvi2. In this study, the alleles in coupling with the markers OPL19 and CH02b10 had the following sizes: OPL19 430 bp and CH02b10 125 bp. Only when both alleles were amplified Rvi2 was considered present.
-
Locus Rvi4: for the selection of this resistance gene, the SSR marker CH02c02a was used. According to Jänsch et al. (2015), this SSR maps 0.5 cM downstream from Rvi4. In this study, the allele in coupling with Rvi4 had a size of 183 bp.
To select for mildew resistance, the following markers were used: for Pl1, the dominant SCAR marker AT20 (allele in coupling 450 bp) (Markussen et al. 1995), and the marker CH04h02 for Pl2 (allele in coupling was 186 bp) (Frey et al. 2004).
To select for fire blight resistance, two dominant SCAR markers, AE10-375 and GE-8019 (alleles in coupling: 373 and 396 bp, respectively), flanking the FBF7 QTL were used (Khan et al. 2007). Only plants amplifying both marker alleles were considered carrying the QTL.
V. inaequalis Inoculation
Phenotypic scab resistance responses of the seedlings were evaluated under glasshouse conditions. The seedlings were sprayed at the four-leaf stage with a conidial suspension of V. inaequalis originating from infected leaves of earlier seedling screenings in the glasshouse and from susceptible, non-treated (fungicide) orchard trees from different areas of Switzerland. The concentration of the suspension was 3.6 × 105 conidia ml−1. The temperature was kept at approximately 20 °C. High humidity was established by building a plastic tent above the seedling trays and using humidifiers (Defensor 3001, Axair AG, Pfäffikon SZ, Switzerland). During the first 48 h after inoculation, relative humidity was kept as close to 100 % as possible to prevent evaporation of the droplets containing the V. inaequalis conidia. For the following 5 days, humidity was reduced to ~80 %, subsequently raised again close to 100 % for 2 days to promote sporulation, and then again reduced to ~80 %. The (resistance) responses were evaluated 14 days after inoculation (DAI) using the scale of Chevalier et al. (1991): class 0 = no visible symptoms; class 1 = pinpoint pits; class 2 = flecks or necrotic lesions and no sporulation; class 3a = necrotic lesions, some with sparse sporulation; class 3b = restricted sporulation; and class 4 = abundantly sporulating lesions. Chevalier classes 1 to 3b were considered resistant.
E. amylovora Inoculation
Plant material preparation of selected elite apple plants and inoculation, in a cabin of the quarantine glasshouse at Agroscope Resarch Station in Waedenswil, was conducted as described by Khan et al. (2006). Five to 12 replicate trees of each selection, showing active shoot growth and a minimal size of 10 cm, were punctured with a syringe at the shoot tip with an E. amylovora solution (Swiss strain FAW610; 109 cfu/ml). Cultivar ‘Gala’ was included as a susceptible control, and ‘Enterprise’ as a resistant control. The length of necrotic lesion (cm) was measured 7, 14, and 21 DAI. Shoot length was measured 7 DAI. The susceptibility of the selections and control cutivars was expressed in percentage lesion length (PLL) of the shoot.
Statistical Analysis
Chi-square tests were performed to determine if there were significant differences between observed frequency of desired genotype (e.g., Rvi6Rvi6) in progeny plants, compared to expected proportions based on Mendel’s laws.
Phenotypic data of the fire blight inoculation experiment (PLL 21 DAI) was tested for normal distribution (p > 0.05) using the Kolmogorov-Smirnov test (statistical software SPSS statistical software, version 22.0 for Windows, SPSS Inc., USA). As data of 5 out of 16 genotypes was not normally distributed (p < 0.05), in the following, non-parametric tests were applied. A Kruskal-Wallis test was used to check if there was a difference between the different genotypes. Since the Kruskal-Wallis test was significant, differences between the genotypes were tested based on Mann-Whitney U tests.The percent lesion length on control cultivars ‘Enterprise’ and ‘Gala’ was compared to the PLL on all other tested genotypes. To test for the effect of the FBF7 fire blight resistance QTL, percent lesion length (21 DAI) was compared between genotypes testing positive for FBF7 and genotypes without FBF7 using a Mann-Whitney U test.
Results
Determination and/or Verification of the Presence of Resistance Genes in the Parents
Ten selections of the Agroscope apple breeding program with marketable fruit quality and pyramided disease resistance markers in a heterozygous state were chosen as parents for breeding elite lines (Table 1). The presence of disease resistance alleles in the parents of the genotypes 'Ariwa', ACW 8259, 'Regia', and 'Reka' were confirmed by DNA amplification of molecular markers linked to specific resistance alleles (Table 1). The presence of the Rvi6 apple scab resistance allele was confirmed in all 10 parents using the CH-Vf1 marker (Vinatzer et al. 2004). Eight parents were tested positive for the Rvi2 resistance gene, and in all eight cases, both marker alleles of CH02b10 SSR and OPL19 SCAR in coupling with Rvi2 were amplified. These eight parents were either derived from ‘Regia’ or ‘Reka’, both known to carry Rvi2. Six parents (ACW 16102, ACW 16111, ACW 16121, ACW 16124, ACW 16126, and ACW 16146) derived from a 'Ariwa' × 'Regia' cross tested positive for the CH02e02a marker linked to Rvi4, as well as for markers linked to Rvi6, Rvi2, and Pl1. Additionally, four of them (ACW 16111, ACW 16121, ACW 16124, and ACW 16126) tested positive for both markers (AE10-375 and GE-8019) flanking the FBF7 fire blight QTL. Two selections (ACW16437 and ACW 16446) derived from the ACW 8259 × ACW 11537 cross tested positive for markers linked to Rvi6, Pl2, and the FBF7 fire blight resistance QTL. The two selected parents (ACW 16252 and ACW 16254) derived from ‘Ariwa’ × ‘Reka’ tested positive for markers of Rvi6, Rvi2, and in the case of ACW 16252 also for the flanking markers for the FBF7 fire blight resistance QTL. Both ‘Reka’ and ‘Regia’ have ‘Cox’s Orange’ in their pedigree, and according the molecular analysis, they both carry the favorable FBF7 allele.
Progeny Genotypes
Progeny sizes ranged between 35 individuals (cross I; ACW 16437 × ACW 16446) to 287 individuals (cross B; ACW 16126 × ACW 16124) (Table 2). Outcrossers were detected by amplification of SSR marker alleles not present in the parents. This occurred in 20 individuals belonging to four different progenies (data not shown). Two individuals of cross B were detected as being triploid, due to repeated detection of three SSR marker alleles at one locus. All outcrossers and triploid plants were removed from the analyses.
The average number of individuals testing positive for SSR markers in coupling and predicting resistance in a homozygous state (Rvi6Rvi6, Rvi2Rvi2, Rvi4Rvi4, or Pl2Pl2) was close to the expected 1:4 Mendelian ratio (Table 2). We found 273 plants with genotype Rvi6Rvi6 out of the 1041 plants tested with the CH-Vf1 marker for Rvi6. Based on Mendel’s laws, 260 plants (25 %) with genotype Rvi6Rvi6 and 781 plants (75 %) with other genotypes would have been expected. Similarly, observed and expected proportions of genotypes Rvi2Rvi2, Rvi4Rvi4, Pl1, Pl2Pl2, and FBF7 were tested with chi-square tests. When comparing chi-square tests of observed frequency of desired genotypes in progeny plants with expected proportions based on Mendel’s laws, we found for all analyses p > 0.05, indicating a non-significant difference between observed and expected proportions of genotypes.
The lowest frequency was observed in the case of Pl2Pl2 (20 %), whereas for Rvi6Rvi6, Rvi2Rvi2, and Rvi4Rvi4, the average frequencies ranged between 24 and 26 %. The average frequency of individuals positive for SCAR markers (homozygous and heterozygous state) was close to 75 %, except for marker AE (FBF7) with 100 % frequency in crosses B and C (ACW 16121 × ACW 16111). This implies the presence of homozygous alleles of AE in at least one of the parents of these two crosses. Because 72 % of the progenies of cross E (ACW 16124 × ACW 16121) amplified the marker AE, these two parents must both carry this marker in a heterozygous state, and therefore, ACW 16111 and ACW 16126 are predicted to carry the marker in the homozygous state.
For each progeny, the expected and observed frequency of the individuals per desired combination of R-genes has been calculated (Table 3). The observed frequencies ranged from 1 to 8 %. Out of 1041 plants from nine crosses, 25 plants displayed the desired pyramided and homozygous allelic status.
Comparison Between Phenotypic and Genotypic Scab Resistance Evaluation
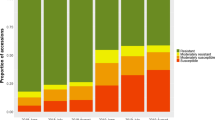
The results of the phenotypic evaluation of scab resistance performed in the greenhouse indicated that more than 94 % of all nine progenies had some degree of scab resistance (classes 1–3b), as was expected from the genotype of the parents (Fig. 1). The majority (57–89 %) of each progeny was assigned to reaction class 2, showing flecks or necrotic lesions and no sporulation. Individuals of seven progenies (all crosses with Rvi4 except cross F (ACW 16146 × ACW 16121), and crosses G (ACW 16254 × ACW 16252) and H (ACW 16252 × ACW 16254) without Rvi4) simultaneously showed both symptoms of scab infection class 2 and pinpoint pits of class 1. These individuals were classified as class 1 + 2 (0–28 % of individuals per progeny). A small proportion (0–6 %) of each progeny was assigned to class 4 (scab susceptible) showing abundantly sporulating lesions.

Determination of scab infection classes of cross progeny according to the scale of Chevalier et al. (1991): class 0 = no visible symptoms; class 1 = pinpoint pits; class 2 = flecks or necrotic lesions and no sporulation; class 3a = necrotic lesions, some with sparse sporulation; class 3b = restricted sporulation; and class 4 = abundantly sporulating lesions
The scab infection classes, observed in the glasshouse test, on plants with different combinations of scab resistance genes Rvi6, Rvi2, and Rvi4 for progeny of crosses A to F are shown in Table 4. None of the plants with one or more scab resistance gene were classified in class 4 (abundantly sporulating lesions, susceptible plants). Most plants (68 %) were scored as class 2 (flecks or necrotic lesions and no sporulation). Seedlings testing positive for Rvi4 did not show the hypersensitive response (pinpoint pits, class 1) expected for this gene. Ninety-six percent of the seedlings carrying only scab resistance Rvi2 were assigned to reaction class 2, showing flecks or necrotic lesions and no sporulation.
Phenotypic Fire Blight Screening in Selected Progeny Plants
The level of fire blight resistance was examined in 14 selected progeny plants. Plants were selected based on their molecular setup (desired progeny genotype; see Table 3) as well as phenotypic scab resistance and tree characteristics. Glasshouse screening of the selected progeny plants with a shoot inoculation test for fire blight resistance indicated differences, among the tested genotypes, in susceptibility toward E. amylovora (Fig. 2). According to a Kruskal-Wallis test, these differences in the percent lesion length 21 DAI between the tested genotypes were significant (chi-square 47.131, p < 0.001). The susceptible control cultivar ‘Gala’ reached 97.0 % of mean lesion length in percent of total shoot length 21 DAI (PLL). Resistant control cultivar ‘Enterprise’ was with 19.5 % PLL as expected among the most resistant genotypes. According to a Mann-Whitney U test, six genotypes (G14, H40, H59, G165, H60, and B232) did not differ significantly (p > 0.05) from control cultivar ‘Enterprise’, and two genotypes (H7 and B232) did not differ significantly from control cultivar ‘Gala’. Genotypes tested positive for FBF7 did not differ significantly from plants without FBF7 (Mann-Whitney U test: N1 = 9, N2 = 7, U = −1.111, p = 0.266). G14 was the most resistant plant and was tested positive for the FBF7 fire blight resistance QTL.

Mean lesion length in percent of total shoot length (with 95 % confidence interval) for selected progeny plants compared to ‘Enterprise’ and ‘Gala’ at 7, 14, and 21 days after inoculation. In brackets, number of inoculated plants and presence of FBF7 QTL for enhanced fire blight resistance are indicated
Discussion
In the present study, elite apple lines carrying homozygous, pyramided resistances against apple scab have been developed. Some of these elite parents additionally carried the resistance genes Pl1 or Pl2 toward powdery mildew and the FBF7 QTL related to enhanced fire blight resistance. To our knowledge, this is the first report on the development of apple plants with pyramided, homozygous scab resistances. These elite selections have the potential to increase breeding efficiency and can be used as parents in future crosses.
The pre-requisite of such a pyramiding approach was the development of molecular markers closely linked to the specific resistance genes. Servin et al. (2004) previously described the two stages of the gene-pyramiding scheme. The first part, which they called “pedigree”, is aimed at the accumulation of one copy of all target genes in a single genotype, called the “root genotype”. The second part, which they called the “fixation step”, is aimed at fixing the target genes into a homozygous state. Apple breeding is a long-term, labor-intensive, and therefore cost-intensive process. The combination of phenotypic and molecular techniques allows for an efficient early selection of desired genotypes in apple breeding (Kellerhals et al. 2009). By developing and using elite selections with pyramided resistance genes in a homozygous state, the efficiency of evaluating the progeny of individual crosses will be increased. Selections with genes in a homozygous state should ensure the transfer of resistance loci to all progeny plants and thereby theoretically eliminate the need to either phenotype or genotype the progeny for the trait of interest. However, it would be necessary to develop a range of such elite selections in order to avoid narrowing the genetic basis of the breeding program.
To achieve durable resistance, combining genes or QTLs with different mechanisms of resistance is a promising strategy. Major genes are interesting for resistance breeding if loss of avirulence leads to fitness costs for the pathogen (Leach et al. 2001). Pyramiding of major resistance genes in rice has shown that durable resistance can be achieved by combining major genes (Correa-Victoria and Martinez 1995). In addition to pyramiding several major resistance genes in the same genetic background, it might be useful to additionally integrate polygenic quantitative resistance against the same pathogen, which is also considered to increase durability. For apple scab, this would be possible as broad spectrum QTLs have been identified by Liebhard et al. (2003) and Soufflet-Freslon et al. (2008).
We consider the strategy to pyramid homozygous resistance genes, toward the same pathogen and to integrate scab, mildew, and fire blight resistance in the same genotype as being a promising approach.
Segregation of the SSR Markers of the Scab Resistance Loci Rvi6, Rvi2, and Rvi4
Rvi6 is located on LG1 (Maliepaard et al. 1998); Rvi2 and Rvi4 are located on LG2 (Bus et al. 2005). The distance between Rvi2 and Rvi4 on LG2 is estimated to be between 40 and 50 cM. Thus, we assumed an independent segregation of all three genes. The calculated percentage of each progeny carrying homozygous pyramided resistance genes at three loci, i.e., Rvi6Rvi6 + Rvi2Rvi2 + Rvi4Rvi4, is 1.6 % (0.25 × 0.25 × 0.25). For progeny B, we found more genotypes with all three scab resistance loci in homozygous state (n = 10) compared to the expected number (n = 4). For progeny G, we found less genotypes with the scab resistance loci in homozygous state (Rvi6Rvi6 + Rvi2Rvi2) (n = 7) compared to the expected number (n = 12, Table 3). These differences might be due to chance.
Phenotypic Evaluations
The phenotypic analysis of scab symptoms in the glasshouse screening was performed 14 DAI. The symptoms development and expression were different compared to when one single resistance gene, such as Rvi6, is incorporated. Symptoms of class 2 were present in a high proportion of seedlings. According to Bus et al. (2000, 2005), the Rvi2 resistance gene conditions distinctive stellar necrosis after about 4 to 6 DAI. Rvi4 causes a hypersensitive response with pit-type lesions already visible at 3 DAI. To get a more precise description on a phenotype (reflecting a specific genetic setup), the evaluations of scab symptoms should be conducted at several time points: at 3 DAI for pinpoint pits, at 4–6 DAI for stellar necrosis, and at 14 DAI for the symptoms elicited by Rvi6 and for susceptible plants. As expected, the seedlings assigned to class 4 in our study lacked all scab resistance alleles in molecular tests. Seedlings predicted by marker information to carry only Rvi4 and not showing the hypersensitive response (pinpoint pits) could be explained by the late scoring for Rvi4. Class 1 and class 1 + 2 symptoms in crosses G, H, and I (all without Rvi4) could be due to the presence of the Rvi6 scab resistance. Chevalier et al. (1991) in fact previously reported this symptom in association with Vf (Rvi6) resistance.
Fire blight symptom development after artificial inoculation with E. amylovora in control cultivars was higher (‘Gala’ +38 % and ‘Enterprise’ +16 %) in this study in comparison to previous years (Baumgartner et al. 2011). This might be explained by variability in climatic conditions. Plants in the fire blight test with very low susceptibility (less than 25 % lesion length relative to ‘Gala’) such as G14 (Rvi6Rvi6 + Rvi2Rvi2 + FBF7) or low susceptibility (between 25 and 40 % lesion length relative to ‘Gala’) such as H40 (Rvi6Rvi6 + Rvi2Rvi2), H59 (Rvi6Rvi6 + Rvi2Rvi2 + FBF7), and G165 (Rvi6Rvi6 + Rvi2Rvi2) are promising parents for new crosses. The sample size was probably too small to show a significant difference in percent lesion length (21 DAI) between plants testing positive for the FBF7 fire blight resistance QTL, compared to plants without FBF7 as demonstrated in earlier evaluations (Baumgartner et al. 2010). Although a significant association between the presence of the FBF7 flanking markers and the level of fire blight resistance can be expected in larger progenies, the flanking markers are not a direct predictor of resistance for individual genotypes. Therefore, it is important to compare the phenotypic levels of resistance with marker, and pedigree information.
Based on the results of the molecular analysis, the phenotypic scab resistance in the artificial scab test, and the criterion “tree feature” (powdery mildew resistance and growth character of the potted seedling), 13 out of 25 possible elite parents were selected for further evaluation in the field. Further screening, especially for tree and fruit characteristics, is necessary to identify the most promising elite parents among the pyramided homozygous plants developed in this study. In years 2012 and 2013, the first eight trees with pyramided homozygous scab resistances flowered. Fruit quality (combination of good appearance, texture, juiciness, and flavor) of three of these elite parents is fair and they will be used as elite parents for crosses in the coming years.
References
Baumgartner I, Franck L, Silvestri G, Patocchi A, Duffy B, Frey J, Kellerhals M (2010) Advanced strategies for breeding fire blight resistant high quality apples. In: Proceedings of 14th international conference on organic fruit-growing, University of Hohenheim, Germany, February 2010, pp 31–37
Baumgartner IO, Patocchi A, Franck L, Kellerhals M (2011) Fire blight resistance from ‘Evereste’ and Malus sieversii used in breeding for new high quality apple cultivars: strategies and results. Acta Hort (ISHS) 895:391–397
Beckerman J (2009) A 33-year evaluation of resistance and pathogenicity in the apple scab-crabapples pathosystem. Hort Sci 44:599–608
Bus V, Gardiner S, Bassett H, Ranatunga C, Rikkerink E (2000) Marker assisted selection for pest and disease resistance in the New Zealand apple breeding programme. Acta Hort (ISHS) 538:541–547
Bus VGM, Rikkerink EHA, van de Weg WE, Rusholme RL, Gardiner SE, Bassett HCM, Kodde LP, Parisi L, Laurens FND, Meulenbroek EJ, Plummer KM (2005) The Vh2 and Vh4 scab resistance genes in two differential hosts derived from Russian apple R12740-7A map to the same linkage group of apple. Mol Breed 15:103–116
Caffier V, Parisi L (2007) Development of apple powdery mildew on sources of resistance to Podosphaera leucotricha, exposed to an inoculum virulent against the major resistance gene Pl-2. Plant Breed 126:319–322
Calenge F, Drouet D, Denancé C, van de Weg WE, Brisset MN, Paulin JP, Durel CE (2005) Identification of a major QTL together with several minor additive or epistatic QTLs for resistance to fire blight in apple in two related progenies. Theor Appl Genet 111:128–135
Chevalier M, Lespinasse Y, Renaudin S (1991) A microscopic study of the different classes of symptoms coded by the Vf gene in apple for resistance to scab (Venturia ineaqualis). Plant Pathol 40:249–256
Correa-Victoria FJ, Martinez C (1995) Genetic structure and virulence diversity of Pyriculariagrisea in breeding for rice blast resistance. In: Induced Mutations and Molecular Techniques for Crop Improvement: Proceedings of an International Symposium on the Use of Induced Mutations and Molecular Techniques for Crop Improvement Jointly Organized by the International Atomic Energy Agency and the Food and Agriculture Organization of the United Nations. International Atomic Energy Agency, Vienna, pp 133–145
Dunemann F, Peil A, Urbanietz A, Garcia-Libreros T (2007) Mapping of the apple powdery mildew resistance gene Pl1 and its genetic association with an NBS-LRR candidate resistance gene. Plant Breed 126:476–481
Durel CE, Denancé C, Brisset MN (2009) Two distinct major QTL for resistance to fire blight co-localize on linkage group 12 in apple genotypes ‘Evereste’ and Malus floribunda 821. Genome 52:139–147
Fischer C, Büttner R, Fischer M (1998) Testing of the scab resistance stability of new resistant cultivars. Erwerbs Obstbau 40:130–135
Frey JE, Frey B, Sauer C, Kellerhals M (2004) Efficient low-cost DNA extraction and multiplex fluorescent PCR method for marker-assisted selection in breeding. Plant Breed 123:554–557
Galli P, Broggini GAL, Kellerhals M, Gessler C, Patocchi A (2010) High-resolution genetic map of the Rvi15 (Vr2) apple scab resistance locus. Mol Breed 26:561–572
Gessler C, Pertot I (2012) Vf scab resistance of Malus. Trees 26:95–108
Gessler C, Patocchi A, Sansavini S, Tartarini S, Gianfranceschi L (2006) Venturia inaequalis resistance in apple. Crit Rev Plant Sci 25:473–503
Hancock JF, Luby JJ, Brown SK, Lobos GA (2008) Apples. In: Hancock JF (ed) Temperate fruit crop breeding. Springer Science+Business Media B.V, New York, pp 1–37
James CM, Evans KM (2004) Identification of molecular markers linked to the mildew resistance genes Pl-d and Pl-w in apple. Acta Hort (ISHS) 663:123–128
Jänsch M, Broggini GAL, Weger J, Bus VGM, Gardiner SE, Bassett H, Patocchi A (2015) Identification of SNPs linked to eight apple disease resistance loci. Mol Breed 35:45 doi:10.1007/s11032-015-0242-4
Kellerhals M, Spuhler M, Duffy B, Patocchi A, Frey JE (2009) Selection efficiency in apple breeding. Acta Hort (ISHS) 814:177–184
Khan MA, Duffy B, Gessler C, Patocchi A (2006) QTL mapping of fire blight resistance in apple. Mol Breed 17:299–306
Khan MA, Durel CE, Duffy B, Drouet D, Kellerhals M, Gessler C, Patocchi A (2007) Development of molecular markers linked to the ‘Fiesta’ linkage group 7 major QTL for fire blight resistance and their application for marker-assisted selection. Genome 50:568–577
Leach JE, Vera Cruz CM, Bai J, Leung H (2001) Pathogen fitness penalty as a predictor of durability of disease resistance genes. Annu Rev Phytopathol 39:187–224
Liebhard R, Kellerhals M, Pfammatter W, Jermini M, Gessler C (2003) Mapping quantitative physiological traits in apple (Malus × domestica Borkh.). Plant Mol Biol 52:511–526
MacHardy WE, Gadoury DM, Gessler C (2001) Parasitic and biological fitness of Venturia inaequalis: relationship to disease management strategies. Plant Dis 85:1036–1051
Maliepaard C, Alston FH, van Arkel G, Brown LM, Chevreau E, Dunemann F, Evans KM, Gardiner S, Guilford P, van Heusden AW, Janse J, Laurens F, Lynn JR, Manganaris AG, den Nijs APM, Periam N, Rikkerink E, Roche P, Ryder C, Sansavini S, Schmidt H, Tartarini S, Verhaegh JJ, Vrielink-van Ginkel M, King GJ (1998) Aligning male and female linkage maps of apple (Malus pumila Mill.) using multi-allelic markers. Theor Appl Genet 97:60–73
Markussen T, Krüger J, Schmidt H, Dunemann F (1995) Identification of PCR-based markers linked to the powdery mildew resistance gene Pl1 from Malus robusta in cultivated apple. Plant Breed 114:530–534
Parisi L, Lespinasse Y, Guillaumes J, Krüger J (1993) A new race of Venturia inaequalis virulent to apples with resistance due to the Vf gene. Phytopathol 83:533–537
Parisi L, Fouillet V, Schouten HJ, Groenwold R, Laurens F, Didelot F, Evans K, Fischer C, Gennari F, Kemp H, Lateur M, Patocchi A, Thissen J, Tsipouridis C (2004) Variability of the pathogenicity of Venturia inaequalis in Europe. Acta Hort (ISHS) 663:107–114
Patocchi A, Frei A, Frey JE, Kellerhals M (2009) Towards improvement of marker assisted selection of apple scab resistant cultivars: Venturia inaequalis virulence surveys and standardization of molecular marker alleles associated with resistance genes. Mol Breed 24:337–347
Peil A, Garcia-Libreros T, Richter K, Trognitz C, Trognitz B, Hanke MV, Flachowsky H (2007) Strong evidence for a fire blight resistance gene of Malus robusta located on linkage group 3. Plant Breed 126:470–475
Roberts AL, Crute IR (1994) Apple scab resistance from Malus floribunda 821 (Vf) is rendered ineffective by isolates of Venturia inaequalis from Malus floribunda. Norw J Agr Sci Suppl 17:403–406
Seglias N, Gessler C (1997) Genetics of apple powdery mildew resistance from Malus zumi (Pl2). IOBC (WPRS) Bull: Integrated Control of Pome Fruit Diseases, Croydon 1996. 20:195–208
Servin B, Martin OC, Mézard M, Hospital F (2004) Toward a theory of marker-assisted gene pyramiding. Genetics 168:513–523
Soriano JM, Joshi SG, van Kaauwen M, Noordijk Y, Groenwold R, Henken B, van de Weg WE, Schouten HJ (2009) Identification and mapping of the novel apple scab resistance gene Vd3. Tree Genet Genome 5:475–482
Soufflet-Freslon V, Gianfranceschi L, Patocchi A, Durel CE (2008) Inheritance studies of apple scab resistance and identification of Rvi14, a new major gene that acts together with other broad-spectrum QTL. Genome 51:657–667
Vinatzer BA, Patocchi A, Tartarini S, Gianfranceschi L, Sansavini S, Gessler C (2004) Isolation of two microsatellite markers from BAC clones of the Vf scab resistance region and molecular characterization of scab-resistant accessions in Malus germplasm. Plant Breed 123:321–326
Acknowledgments
The authors are grateful to Teresa Koller, Bettina Gleichauf, Beatrice Frey, Markus Spuhler, Markus Oggenfuss, Pierre-Marie Le Roux, Brion Duffy, Jürgen Krauss, Reto Leumann, and all Agroscope for scientific and technical assistance, to Eve Silfverberg for proofreading, and to Claude Renaux for advice on statistics. Fire blight inoculation tests were financially supported by the Swiss Federal Office for Agriculture project “ZUEFOS” (breeding of fire blight tolerant fruit varieties). Part of this work was conducted within the framework of the European research network COST864.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Baumgartner, I.O., Patocchi, A., Frey, J.E. et al. Breeding Elite Lines of Apple Carrying Pyramided Homozygous Resistance Genes Against Apple Scab and Resistance Against Powdery Mildew and Fire Blight. Plant Mol Biol Rep 33, 1573–1583 (2015). https://doi.org/10.1007/s11105-015-0858-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11105-015-0858-x