
Abstract
Background and aims
Soil seed bank plays a significant role in the natural recovery and the succession of forest. A large number of natural secondary forests are distributed in the Qining Mountains, China. However, it remains unclear how the soil seed banks develop and influence the recovery of secondary forests.
Methods
We explored the aboveground vegetation and soil seed bank at 27 plots along forest developmental stages of pine and oak forests in the Qinling Mountains. Species composition and abundance of aboveground vegetation as well as seed bank composition and density were surveyed and their correlations with forest developmental stages were determined.
Results
The results showed that the species richness of seed banks was the highest in middle-aged forests, as well as the aboveground vegetation. The seed density and abundance increased with forest developmental stages and varied among different forest types. The similarity between seed banks at different developmental stages of the same forest type was high, as well as aboveground vegetation. The seed bank and aboveground vegetation showed low similarity across the three developmental stages of both forest types, and the persistent soil seed bank did not reflect the changes in aboveground vegetation.
Conclusion
Our findings demonstrate that developmental stages play an important role in the composition of seed banks and aboveground vegetation. Our results also highlight the persistent soil seed bank contributes less to the forest recovery. We can assume that the natural recovery of disturbed natural secondary forests is largely dependent on transient soil seed bank and seed dispersal.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the Industrial Revolution, due to intensive human disturbance, the area of secondary forests evolved from primary forests has increased dramatically (Xiang et al. 2016). More than two billion ha of land have been identified globally as potentially suitable for either passive or active forest restoration (Crouzeilles et al. 2016). Forest restoration is a global priority and the question of how to promote the restoration of secondary forests has become a research hotspot (Cruz-Alonso et al. 2019). Forest regeneration involves the stages of seed production, seed dispersal, germination, and seedling survival (Cole et al. 2011; Calama et al. 2017), and its success depends on the physical characteristics of the surrounding landscape (Sprengelmeyer and Rebertus 2015), the intensity and frequency of disturbances (Sanou et al. 2018), and the plant colonization by propagules moving into the area (Chazdon and Guariguata 2016; Wandrag et al. 2015). Compared with plants, seeds are more resistant to stress and disturbance (Gutterman 2012), and therefore the role of soil seed bank is particularly important in the natural recovery process after vegetation destruction. However, previous studies on plant diversity have focused mainly on aboveground vegetation while ignoring the protective value of the soil seed bank (Zhao et al. 2021), making it difficult to understand how soil seed bank diversity influences aboveground plant communities (Plue et al. 2017).
Soil seed banks, as a potential source of genetic diversity for plant populations (Yang et al. 2021), not only reflect the stability of communities, but also respond to the heterogeneity of biological communities in terms of species composition, structure, function, and dynamics, as well as differences in community type, community structure, organization level, developmental stage, and habitat (Miao et al. 2016; Li et al. 2012). Transient seed banks consist of short-lived seeds that do not exhibit dormancy and are dispersed for short periods during the year. In contrast, persistent seed banks contain seeds that can remain viable in the soil for more than a year, often in a dormant state, and can germinate in response to unpredictable disturbances, playing an important role in the restoration and management of vegetation (Christoffoleti and Caetano 1998). Seeds from many genotypes from numerous generations accumulate in the soil with the regeneration of the community, creating a varied gene pool that may even be greater than the genetic variety of aboveground populations. Seeds kept in soil seed banks allow plant species to grow and establish populations under favorable conditions in unpredictable environments (Bossuyt and Honnay 2008). Additionally, soil seed banks can predict future changes and influence the direction of secondary succession following community disruption, especially persistent soil seed banks. (Koncz et al. 2010). Many previous studies have demonstrated that as part of biodiversity research, the study of soil seed banks is crucial for understanding the mechanisms that maintain population and community ecological diversity (Fengqin et al. 2017; Templeton and Levin 1979).
The Qinling Mountains, located in northwest China, have developed into a globally significant area for species and biodiversity conservation after decades of secondary succession following extensive logging in the 1960s and 1970s and the implementation of the Natural Forest Protection Program in 1998 (Chai and Wang 2016). A large area of natural secondary forest was distributed in the Qinling Mountains. Like other temperate forests, forest recovery after disturbance in this region depends on plant regeneration and soil seed bank (Barros et al. 2021; Kennard et al. 2002). Much research work on soil seed banks has been carried out worldwide, but studies on the characteristics and functions of the soil seed bank in the Qinling Mountains are rare. A study on the soil seed bank of Quercus aliena var. acuteserrata in the Qinling Mountains showed that the size of soil seed bank varied with slope aspect, being largest on semi-sunny slopes and smallest on shady slopes (Lu et al. 2019). Another study showed that the time dynamics of seed rain differed significantly among different tree species, and that the peak time of seed rain of Pinus tabulaeformis was later than that of Quercus aliena var. acuteserrata and Pinus huashanensis.
Although these studies provided evidence that soil seed banks of different species vary in time and space, no study to-date has looked at the dynamics of the Qinling soil seed bank in the process of secondary forest restoration, or focused on the role of the seed bank in secondary forest restoration. The structure and composition of the forest soil seed bank change with the development age of the forest. It has been found that the diversity of plant and soil seed banks increases with the development of plantations (Kůrová 2016). Different studies have also found that younger forests have higher seed density than older forests (Zhang et al. 2014). The relationship between the density of seed bank and the age of forest has not been uniformly established. Moreover, most studies focused on transient soil seed banks, ignoring the importance of persistent soil seed banks. Qinling is a typical place to study forest vegetation restoration and more studies are needed to improve our understanding of the role of soil seed banks on restoration in temperate forests in China.
Pine forest and oak forest are the main natural secondary forest types in the mid-altitude zone of the Qinling Mountains, and the primary establishment species in the pine and oak forests are Pinus tabuliformis and Quercus aliena var. acuteserrata. In the middle of the Qinling Mountains, we carried out aboveground vegetation species surveys, sampled the persistent soil seed banks, and described the species composition, diversity, richness, and different lifespans of pine and oak forests. We also discussed how soil seed bank characteristics changed as a result of the increasing forest developmental age and succession of the forest. In addition, we studied the similarities between soil seed banks and aboveground vegetation. The objectives of this paper were therefore: (1) to determine how the density, richness, and compositional similarity of seed banks compare to aboveground communities across different ages of two Qinling forest types, and (2) to determine how soil seed banks are linked to vegetation restoration. Specifically, we hypothesized that (1) Jaccard similarity between SB at one developmental stage and SB at the next developmental stage reflects the importance of seed persistence, (2) Jaccard similarity between SB at one developmental stage and VEG at the next developmental stage reflects the potential of the soil seed bank in vegetational restoration, and (3) the richness and density of the persistent seed bank would be positively affected by increasing development age and seed banks would reflect and influence aboveground vegetation dynamics to some extent.
Materials and methods
Study area
The study area is located in the Crown Forest Farm (33°32′21″N, 108°22′26″E) in Ningshan County, Shaanxi Province, China. The Crown Forest Farm lies in the middle section of the Qinling Mountains and is the transition zone between the northern subtropical and warm temperate climate zones (He et al. 2021). The elevation ranges from 880 to 2340 m, the average annual temperature is 12.3℃, the annual frost-free period is 216 d, and the annual average precipitation is 908.0 mm, mostly concentrated in summer. The soil is brown loam with a pH of 5.96 (Han et al. 2021).
Due to the different ownership of forests, Crown Forest Farm’s natural forests are composed of a small part of collective forests (old forests) that have ceased deforestation for a long time, state-owned forests that have been controlled at the early stage of large-scale deforestation (middle-aged forests), and collective forests (young forests) that have been halted for logging after the full implementation of the Natural Forest Protection Program.
Experimental design and data collection
In early July 2021, we chose pine and oak forests with essentially the same stand conditions at four sites in Crown Forest Farm. We selected pine and oak forests at three different developmental stages (young forest (I), middle-aged forest (II), and mature forest (III)). The wood cores of trees were drilled with growth cones, and the forest plots of different developmental stages were selected according to the forest resource survey standard “Age Class and Age Class Division of Main tree Species”. For the pine forest, the age structure is young forest (30 years), middle-aged forest (31–50 years), and mature forest (61–80 years). For the oak forest, the age structure is young forest (40 years), middle-aged forest (41–60 years), and mature forest (81–120 years).
In the four sampling sites, we created five and four 20 m × 20 m plots in each of the developmental stages of pine and oak forests, respectively (in total 27 plots). The distance between the sampling sites was at least 300 m, with at least 20 m distance between two sampling plots nested in each site. We set up five shrub samples of 2 m × 2 m and 5 herb samples of 1 m × 1 m in each sample plot of 20 m × 20 m to determine the species, number, and height of aboveground shrubs and herbs (Esmailzadeh et al. 2011). The information of longitude, latitude, altitude, slope, and slope direction was also recorded (Fig. 1.)

Geographical location of the study area, four main sampling sites and 27 sampling plots
Aboveground vegetation investigation
Species composition and richness were recorded in each plot at the end of July and August 2021, during the peak of the growing season. Percentage cover of all species within each plot was recorded using visual assessment. Additionally, we estimated the abundance of each species.
Soil seed bank sampling
Since seeds are usually concentrated in the topsoil layers (Zhang et al. 2001), soil samples (0–10 cm) were collected from all plots to analyze the species composition characteristics of soil seed banks. To comprehensively study the characteristics of the persistent soil seed bank in this region, in August 2021, when the seed germination period had ended and the current-season seeds had not yet dispersed into the soil, we collected the persistent seed bank (Ma et al. 2011). Before collecting samples, we removed the litter layer to avoid the effects of partial seed rain that had fallen earlier and to more accurately measure the long-lasting seeds that accumulate in the soil over time. We used the line transect method to collect the soil seed bank samples (Tesfaye et al. 2004), a set of samples was taken every 2.5 m along the diagonal of the sample plots, 10 soil cores were collected in each sample plot and then mixed into one soil sample, each soil core had a depth of 10 cm and a width and length of 10 cm (10 cm × 10 cm × 10 cm). All the soil samples were stored in plastic bags and labeled with a code. Then they were kept in a cool place and transported to the laboratory (Fig. 1.).
Seed germination experiment
The species composition and quantity of viable seeds in soil were quantitatively analyzed by seedling emergence method (Bourgeois et al. 2017). We sampled the seed bank before the seed rains in August and began culture tests in the laboratory in September. In September, each soil sample was cultured in a germination tray (31 cm × 31 cm × 4 cm) with a soil layer thickness of 2–3.5 cm after the stone and plant remains were removed. To avoid contamination of local seeds, seeds are germinated and diagnosed in a separate greenhouse, with sterile sand at the bottom of the germinating tray. The right temperature (25℃) and sufficient light were given to maximize germination (Birhanu et al. 2022). Water was sprinkled on the soil samples once a day at regular intervals to maintain moisture. The seedlings were recognized by their morphological traits, once they had been identified, they were deemed to be viable seeds, and their numbers were recorded, then removed from the germination tray. The remaining soil was then thoroughly mixed, giving all of the seeds an equal chance of germination. Identification and regular counting of germinated seedlings from each sample were continued until no more new seedlings emerged (for about 2 weeks) (Luo et al. 2017). The germination experiment lasted more than six months, from September 2021 to March 2022.
Statistical analyses
We summarized the seed number and species richness of the soil seed bank in each sample site and calculated the density (He et al. 2021), diversity index (Kent and Coker 1994), evenness and similarity index of the soil seed bank under different developmental stages in pine and oak forests (Smith et al. 1998; Jaccard 1912). These indices were calculated with the following equations:
Soil seed bank density:
Simpson's diversity index:
Pielou's evenness index:
Jaccard similarity index:
In these equations: N is the total number of seeds in the soil sample; A is the area of the soil sample, in square meters; S is the sum of the number of all species in the community; Pi is the relative multiplicity of species i, Pi = Ni/N; H denotes the Shannon-Wiener diversity index; c is the number of species that were found both in the plant community and in the soil seed bank; a and b indicate the number of species in plant community and soil seed bank, respectively.
The Jaccard similarity index (CJ) was used to assess the similarity between the species composition of the different components (SB-soil seed bank, VEG-aboveground vegetation). Jaccard similarity coefficients are extremely dissimilar when they reach 0.00–0.25, moderately dissimilar when they reach 0.25–0.50, moderate when they reach 0.50–0.75, and extremely similar when they reach 0.75–1.00 (Jaccard 1912). We calculated the Jaccard similarity index between the SB and the VEG at different developmental stages, as well as between SB and SB, and between VEG and VEG of pine forest and oak forest to evaluate the contribution and persistent of the soil seed bank.
We used analysis of variance (ANOVA) to determine differences between the variables of soil seed bank indicators at different development stages of the same forest type, which included density, richness, diversity, and evenness (Díaz-Villa et al. 2003). Before the ANOVA analysis, we tested the assumptions of the ANOVA, and the results showed that: (1) Normality: no obvious deviation from normal distribution was found in the histograms and Q-Q plots; (2) Homogeneity of variance: the p-values of the Levene test were all greater than 0.05, indicating that the soil seed bank indicators of different age groups had equal variances; (3) Independence: since the samples were randomly drawn, we assumed that they were independent of each other. Therefore, we concluded that the assumptions of the ANOVA were met, and we could proceed to the next step of the analysis. The least significant difference (LSD) test was used for multiple comparisons to verify whether the effect was significant or not. The results were considered statistically significant at p < 0.05. The above statistical analyses were performed using SPSS 26.0 software(SPSS Inc., Chicago, Illinois, USA), while Origin 2022 (Origin Lab Corporation, Northampton, MA, USA) was used for plotting graphs.
Results
Understory vegetation
In all the plots we studied, there were 71 species of understory plants belonging to 40 plant families in the pine forest, and 54 species of understory plants belonging to 30 families in the oak forest. Among them, herbaceous plants accounted for a large proportion, while there were only 17 and 11 shrubby plants in pine forest and oak forest, respectively. Of the two forest types, the number of understory vegetation in middle-aged forest was the highest and the understory vegetation was dominated by Rosaceae, Asteraceae, Rubiaceae and Fabaceae. Among the understory plants of all vegetation types, Rosa multiflora and Lespedeza bicolor were the most abundant shrub species, while the dominant species of herbaceous plants were Duchesnea indica and Chrysanthemum indicum (Fig. 2).

Percentage of families of soil seed bank species in pine forest and oak forest at different development stages
Soil seed bank composition
In the seed bank, a total of 17 plant species from 14 families and 17 genera were identified. The largest percentage of these species was Lysimachia christiniae, reaching 32.8% of the total seeds in the seed bank. Soil seed banks were overwhelmingly dominated by herbaceous plants. Few seeds of woody plants could be found, and the number of seeds from trees and shrubs only accounted for 0.6% and 0.4% of the total seed bank.
The species composition and seed density of the soil seed bank varied with the developmental stage. The soil seed bank of the pine forest had a total of 16 plant species that belonged to 13 families and 16 genera. Among them, there were a considerable number of plants from the families of Fabaceae (2 species), Poaceae (2 species), and Rosaceae (2 species). The number of species counted in the soil seed bank of pine forest in stages I, II, and III was 9, 12, and 7, respectively. Five species were recorded in the soil seed bank of the pine forest at all three developmental stages (Table 1). A total of 12 plant species from 11 families and 12 genera were found in the soil seed bank of the oak forest. Of these, Rosaceae plant species occurred the most (16.67%). The number of species counted in the soil seed bank of the oak forest in stages I, II, and III was 7, 9, and 7, respectively. The two plants, Deyeuxia pyramidalis and Lysimachia christiniae were present in the soil seed bank of the oak forest at all three developmental stages (Table 1).
In pine and oak forests, the number of seeds in the soil seed bank increased gradually with developmental stages. At all developmental stages in pine and oak forests, seeds that did not appear in the aboveground vegetation were present in the soil seed bank, and the majority of the seeds recorded in established vegetation were not found in the soil seed bank. Meanwhile, there was only a weak similarity between aboveground vegetation and soil seed banks. Tree and shrub seeds were only found in individual sample seed banks, which appeared in pine forest stage II and oak forest stage I. Herbaceous plant seeds accounted for more than 98% of the total seed bank and were mostly perennial herbs, with only a small proportion of tree and shrub seeds(Fig. 3).

Concept figure of the soil seed bank density and lifespan over three development stages in pine and oak forest. □:Species that occur both in the soil seed bank and above-ground; ■:Species that only occur in the seed bank; the lifespan of the seeds in the soil seed bank.
 :herbs;
:herbs;
 :trees;
:trees;
 :shrubs
:shrubs
Biodiversity indices and density of soil seed bank
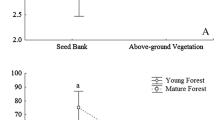
The density, richness, Simpson's diversity index, and Pielou’s evenness index of the soil seed banks of pine forest were overall higher than those of oak forest. The characteristics of soil seed banks varied according to the different developmental stages, with stage II in pine and oak forests having the highest seed bank richness among the three developmental stages (Fig. 4d). However, the other characteristics index values of stage II weren't the highest, indicating that there was the emergence of absolutely dominant species in stage II of the soil seed bank. This situation was especially obvious in oak forests, where the highest seed bank richness was found in stage II, but the Simpson's diversity index and Pielou’s evenness index in stage II were significantly lower than those in stages I and III, and the difference was significant (P < 0.05), which indicates that the species distribution of the soil seed bank in oak forest stage II was extremely uneven. The median of each index of the pine forest seed bank was at the highest level in the box line plot of stage III, indicating that the diversity and evenness of the pine forest soil seed bank increased with developmental age. In the oak forest, the density of the soil seed bank did not significantly change with developmental age (Fig. 4a), and the seed richness did not significantly differ among the three developmental stages (Fig. 4).

Soil seed bank richness, density, Simpson’s diversity, and Pielou’s evenness for the overall seed bank from three developmental stages of the two forest types. Error bars represent standard errors. Significant differences are indicated by * (p < 0.05)
Comparisons of the Jaccard index of soil seed banks
According to the analysis of the similarity between the components (SB, VEG) of the two forests, whether it is a pine forest or oak forest, the similarity between SB and SB, and the similarity between VEG and VEG are significantly greater than those between SB and VEG. The high similarity between SB and SB indicates that persistent seeds continue to accumulate in the soil with increasing developmental age. The similarity between SB and SB, as well as the similarity between VEG and VEG in adjacent developmental stages was higher than those in interval developmental ages. However, the similarity between SB in one developmental stage and VEG in the next developmental stage was low in both pine forest and oak forest, and the Jaccard maximum value was 0.114, indicating that they were extremely dissimilar. (Fig. 5).

Jaccard similarity for pine forest and oak forest between soil seed banks and aboveground vegetation. Numbers and color depth on the squares represent correlation coefficients, which can be referenced on the side axis
Discussion
As expected, soil seed banks differed between forest types and at different developmental stages of the same forest type. Our results agree with previous work on the Czech Republic and Loess Plateau forest seed banks (Wang et al. 2020; Kůrová 2016). In our study, the number of species in the soil seed bank of pine forest was higher than that of oak forest, which is consistent with the results obtained by Cheng et al. (2012) on the Ziwuling pine and oak forests. This may be related to the richness of aboveground vegetation and the intensity of its seed rain dispersal, with aboveground vegetation surveys finding higher species richness in pine forests than in oak forests (Kang et al. 2017). However, in contrast with our original hypothesis, the number of species stored in the soil seed bank did not increase with the extension of developmental age, and the number of species in the middle-aged forest seed bank was the highest. As we all know, soil seed banks are derived directly from aboveground vegetation. The difference mentioned above may be caused by the richness of the understory vegetation which is the most abundant in the middle-aged forest. Li et al. (2014) found that the density of soil seed bank increased with the increase of forest community restoration years in a certain age range, and the soil seed bank density of pine forest in this study showed that stage III > stage II > stage I, and there was a significant difference between stage III and stage I, which was similar to the findings of Kůrová (2016). This shows that the number of species in the soil seed bank does not necessarily increase with the increase of the developmental stage, but the number of stored seeds is directly proportional to the developmental stage.
Silvertown and Charlesworth (2009) found that forest soil seed bank density is generally 102–103 seeds/m2. In this experiment, the results were within the range of Sitvertown, the soil seed bank density of pine and oak forests ranged from 840–1610 grains/m2 and 430–470 grains/m2, respectively. However, the soil seed bank densities in our study were occasionally lower than those reported for other temperate forests, for example, in northwestern Switzerland where soil seed bank densities in temperate forests ranged from 1000–4500 seeds/m2 (Amrein et al. 2005). This phenomenon may be caused by the difference between the study site and the forest type. Our results are similar to Wang's findings in pine and oak forests in northern China (Wang et al. 2020).
There were almost no seeds of woody plants in the soil seed bank, we highlight three possible explanations. Firstly, This is related to the fact that our research object is a pure forest, and there are not many woody species on the ground. Secondly, the seeds of Pinus tabulaeformis and Quercus aliena var. acuteserrata have a single dispersal mode, and they face the predation pressure of larger animals (Huo et al. 2019). Thirdly, the seeds of woody plants do not usually form a long-term and lasting seed bank (Thompson et al. 1993). We found that Rhus potaninii, as one of the common tree species of aboveground vegetation and one of the most successful pioneer species in forest succession in the research region, was the only tree species that appeared in the soil seed bank. Rhus potaninii was only found in the early developmental stage. We hypothesize that these seeds were ingested by birds and excreted with feces, but did not germinate immediately due to unfavorable environmental conditions, but accumulated in the soil, and germinated only when given suitable conditions (Chen et al. 2022). However, we have not determined how woody plant seeds change in persistent soil seed banks in our study. Given the relative importance of woody plants in the composition of soil seed banks, we conclude that future studies should consider the role of woody plants in the formation of transient soil seed banks.
Soil seed bank diversity
The Simpson diversity index of the pine forest soil seed bank did not differ significantly among the three developmental stages, and the Pielou evenness index showed stage III > stage II > stage I, indicating that the distribution of species in the soil seed bank of pine forest became more homogeneous with the increasing developmental age, similar to the findings of Kůrová (2016). The Pielou evenness index and Simpson diversity index of the oak forest soil seed bank were highest at stage I, then at stage III, which might be related to the fact that the young forest is rich in shrubs and grasses, with little interspecific competition, and the dominant species are not yet prominent (Yuan et al. 2009), but also may be due to the fact that with the developmental of the oak forest, the forest canopy increases, the first appearance of numerous light-loving pioneer species gradually eliminates, shade-tolerant species become the dominant species, so that the understory vegetation gradually monoculture, species uniformity decreases. As each species reaches a balanced constraint, the community enters a stable equilibrium stage in the mature forest stage, increasing species variety and evenness (Wang et al. 2017). Zhao et al. (2021) found that the species richness of the soil seed banks in the plantations was significantly higher than that in the natural restoration types. This is because compared with planted forests, secondary forests are more affected by changes in the canopy in the early development of the forest. In order to adapt to the changes in the canopy, many species emerged in the natural secondary forest at the early development stage. However, as forest development progresses, the natural secondary forest creates a close crown that reduces the available light resources, thus limiting the growth of understory plants and ultimately reducing seed production and spread (Dang et al. 2018; Erfanzadeh et al. 2020). Our study demonstrated that the richness of understory vegetation and soil seed bank of oak forest was more susceptible to age-induced canopy changes.
The feedback between vegetation composition and soil seed bank
The similarity between the soil seed bank and aboveground vegetation can measure plant community stability and predict the development trend of this community (Chang et al. 2001). A study by Bossuyt and Honnay (2008) showed that the species similarity between the soil seed bank and aboveground vegetation in forest ecosystems is about 30%. In this study, the similarity coefficient between soil seed banks of pine forest and oak forest at one developmental stage and aboveground vegetation at the next developmental stage was less than 0.1, which is similar to the findings of Godefroid et al. (2006). The similarity between the aboveground vegetation and the seed bank species was low in the study area, and there were differences among the different vegetation types. There may be several causes for the low similarity between aboveground species and persistent soil seed banks. First, not all aboveground vegetation can produce persistent seeds that survive in the soil for a long time (Shiferaw et al. 2018.). Second, there may be more seeds in the soil seed bank that are in a dormant state and have not germinated under suitable conditions (Long et al. 2015). Third, this may be related to the fact that the reproductive strategy of perennial herbaceous plants makes them contribute less to the diversity of the soil seed bank, which reduces the compositional similarity between the seed bank and the local standing (Gomes et al. 2019).
The majority of aboveground species were not represented in the soil seed bank; however, species that were absent from aboveground vegetation were present in the soil seed bank, which confirms previous research (Devlaeminck et al. 2005). The absence of aboveground plant species in the seed bank suggests that most tree species were renewed without maintaining persistent propagules in the soil seed bank and that most species' seeds only survived for less than one year on the forest floor (Gomes et al. 2019; Shiferaw et al. 2018). The soil seed banks at adjacent developmental stages had a medium similarity in pine and oak forests, with a Jaccard coefficient of 0.50–0.75, as well as the similarity between aboveground vegetations. The representative families (Asteraceae, Fabaceae, Rosaceae) of understory vegetation also appeared in seed banks. These results indicate that the importance of various understory vegetation species changes during forest development. Some species have disappeared from the understory vegetation, but their seeds may still be stored in the seed bank (Devlaeminck et al. 2005), some plants can survive the entire development of the forest, and their seeds can also be permanently stored in the seed bank.
The low number of common species between aboveground vegetation and soil seed banks in both communities indicates that the number of seeds in the soil seed bank is not sufficient to support the number of seeds needed for aboveground vegetation recovery. The persistent soil seed bank contributes less to the forest recovery process. This emphasizes the importance of transient seed banks and exotic seeds to sustain natural regeneration.
Conclusion
Our results revealed the characteristics of soil seed banks in pine and oak forests at different development ages and provided strong evidence that compared to the abrupt change in the composition of aboveground vegetation, the composition of the seed bank hardly changes in this process. In addition, forest development stages greatly affect the composition of aboveground vegetation and seed banks. Middle-aged forest always keeps higher species diversity of soil seed bank. Further analysis showed that the seed bank of pine forest had a greater contribution to aboveground vegetation than that of oak forest, but in general, aboveground vegetation composition had only a marginal effect on the composition of the persistent seed bank. Our results suggest that although new persistent seeds are accumulating in soil seed banks as forests develop, the number of these seeds is not sufficient to support the natural recovery of natural secondary forests. Long-term community dynamics aboveground do not necessarily reflect belowground dynamics and the persistent seed bank may serve as a seed source for future restoration efforts. Our study did not involve the study of transient soil seed banks and did not discuss the effects of soil physicochemical properties and environmental factors in different forest development stages on soil seed banks. In conclusion, we call on ecologists to pay more attention to a more comprehensive and in-depth study of soil seed banks.Data availability The data that support the findings of this study are available from the corresponding author upon reasonable request.
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Amrein D, Rusterholz HP, Baur B (2005) Disturbance of suburban Fagus forests by recreational activities: Effects on soil characteristics, above-ground vegetation and seed bank. Appl Veg Sci 8(2):175–182. https://doi.org/10.1111/j.1654-109X.2005.tb00643.x
Barros MF, Ribeiro EMS, Vanderlei RS et al (2021) Resprouting drives successional pathways and the resilience of Caatinga dry forest in human-modified landscapes. For Ecol Manage 482:118881. https://doi.org/10.1016/j.foreco.2020.118881
Birhanu L, Bekele T, Tesfaw B et al (2022) Soil seed bank composition and aboveground vegetation in dry Afromontane forest patches of Northwestern Ethiopia. Trees. For People 9:100292. https://doi.org/10.1016/j.tfp.2022.100292
Bossuyt B, Honnay O (2008) Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J Veg Sci 19(6):875–884. https://doi.org/10.3170/2008-8-18462
Bourgeois B, Boutin C, Vanasse A et al (2017) Divergence between riparian seed banks and standing vegetation increases along successional trajectories. J Veg Sci 28(4):787–797. https://doi.org/10.1111/jvs.12536
Calama R, Manso R, Lucas-Borja ME et al (2017) Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For Syst 26. https://doi.org/10.5424/FS/2017262-11255
Chai Z, Wang DA (2016) Comparison of species composition and community assemblage of secondary forests between the birch and pine-oak belts in the mid-altitude zone of the Qinling Mountains, China. Peerj 4:e1900. https://doi.org/10.7717/peerj.1900
Chang ER, Jefferies RL, Carleton TJ (2001) Relationship between vegetation and soil seed banks in an arctic coastal marsh. J Ecol 89(3):367–384. https://doi.org/10.1046/j.1365-2745.2001.00549.x
Chazdon RL, Guariguata MR (2016) Natural regeneration as a tool for large-scale forest restoration in the tropics: prospects and challenges. Biotropica 48(6):716–730. https://doi.org/10.1111/btp.12381
Chen X, Chen H, Zhao M et al (2022) Insect industrialization and prospect in commerce: a case of China. Entomol Res 52(4):178–194. https://doi.org/10.1111/1748-5967.12576
Cheng J, Cheng J, Shao H et al (2012) Soil seed banks and forest succession direction reflect soil quality in Ziwuling Mountain, Loess Plateau, China. Clean Soil, Air, Water 40:140–147. https://doi.org/10.1002/clen.201000377
Christoffoleti PJ, Caetano RSX (1998) Soil seed banks. Sci Agric 55:74–78. https://doi.org/10.1590/S0103-90161998000500013
Cole RJ, Holl KD, Keene CL et al (2011) Direct seeding of late-successional trees to restore tropical montane forest. For Ecol Manage 261:1590–1597. https://doi.org/10.1016/j.foreco.2010.06.038
Crouzeilles R, Curran M, Ferreira MS et al (2016) A global meta-analysis on the ecological drivers of forest restoration success. Nat Commun 7(1):11666. https://doi.org/10.1038/ncomms11666
Cruz-Alonso V, Ruiz-Benito P, Villar-Salvador P et al (2019) Long term recovery of multifunctionality in Mediterranean forests depends on restoration strategy and forest type. J Appl Ecol 56(3):745–757. https://doi.org/10.1111/1365-2664.13340
Dang P, Gao Y, Liu J et al (2018) Effects of thinning intensity on understory vegetation and soil microbial communities of a mature Chinese pine plantation in the Loess Plateau. Sci Total Environ 630:171–180
Devlaeminck R, Bossuyt B, Hermy M (2005) Inflow of seeds through the forest edge: Evidence from seed bank and vegetation patterns. Plant Ecol 176(1):1–17. https://doi.org/10.1007/s11258-004-0008-2
Díaz-Villa MD, Marañón T, Arroyo J et al (2003) Soil seed bank and floristic diversity in a forest-grassland mosaic in southern Spain. J Veg Sci 14(5):701–709. https://doi.org/10.1111/j.1654-1103.2003.tb02202.x
Erfanzadeh R, Hazhir S, Jafari M (2020) Effect of cushion plants on soil seed bank in overgrazed semi-arid regions. Land Degrad Dev 31:990–1000. https://doi.org/10.1002/ldr.3517
Esmailzadeh O, Hosseini SM, Tabari M et al (2011) Persistent soil seed banks and floristic diversity in Fagus orientalis forest communities in the Hyrcanian vegetation region of Iran. Funct Ecol Plants 206(4):365–372. https://doi.org/10.1016/j.flora.2010.04.024
Fengqin JIA, Tashpolat T, Nan WU et al (2017) Characteristics of soil seed banks at different geomorphic positions within the longitudinal sand dunes of the Gurbantunggut Desert, China. J Arid Land 9(3):355–367. https://doi.org/10.1007/s40333-017-0055-x
Godefroid S, Phartyal SS, Koedam N (2006) Depth distribution and composition of seed banks under different tree layers in a managed temperate forest ecosystem. Acta Oecol 29(3):283–292. https://doi.org/10.1016/j.actao.2005.11.005
Gomes FM, de Oliveira CC, da Rocha MR et al (2019) Relationships between soil seed bank composition and standing vegetation along chronosequences in a tropical dry forest in north-eastern Brazil. J Trop Ecol 35(4):173–184. https://doi.org/10.1017/S0266467419000130
Gutterman Y (2012) Seed germination in desert plants. Springer Science & Business Media, Berlin
Han AX, Qiu J, He C et al (2021) Spatial distribution patterns and intraspecific and interspecific associations of dominant shrub species Lonicera fragrantissima var. lancifolia in Huangguanof Qinling Mountains, China. Yingyong Shengtai Xuebao 32(08):2737–2744. https://doi.org/10.13287/j.1001-9332.202208.003
He C, Liu R, Yang Z et al (2021) Species composition and community structure of warm temperate deciduous broadleaved forests in Huangguan of Qinling Mountains, China. Yingyong Shengtai Xuebao 32(08):2737–2744. https://doi.org/10.13287/j.1001-9332.202108.001
Huo XY, Kang HB, Wang DX et al (2019) Effects of rodents on seed dispersal patterns of constructive species in the pine-oak mixed forests of the Qinling Mountains, Shaanxi Province, China. Acta Ecol Sin 39:2435–2443
Jaccard P (1912) The distribution of the flora in the alpine zone. 1. New Phytol 11(2):37–50. https://doi.org/10.1111/j.1469-8137.1912.tb05611.x
Kang H, Zheng Y, Liu S et al (2017) Population structure and spatial pattern of predominant tree species in a pine–oak mosaic mixed forest in the Qinling Mountains. China. J Plant Interact 12(1):78–86. https://doi.org/10.1080/17429145.2017.1283069
Kennard DK, Gould K, Putz FE et al (2002) Effect of disturbance intensity on regeneration mechanisms in a tropical dry forest. For Ecol Manage 162(2–3):197–208. https://doi.org/10.1016/S0378-1127(01)00506-0
Kent M, Coker P (1994) Vegetation description and analysis: a practical approach. John Willey & Sons, New York
Koncz G, Papp M, Török P et al (2010) The role of seed bank in the dynamics of understorey in an oak forest in Hungary. Acta Biol Hung 61(Supplement-1):109–119. https://doi.org/10.1556/abiol.61.2010.suppl.11
Kůrová J (2016) The impact of soil properties and forest stand age on the soil seed bank. Folia Geobot 51(1):27–37
Li SF, Liu WD, Su JR et al (2012) Dynamics of seed banks in soil at the different restoration stages of a monsoon broad-leaved evergreen forest. Sci Silvae Sin 48(9):56–61
Li X, Jiang D, Zhou Q et al (2014) Soil seed bank characteristics beneath an age sequence of Caragana microphylla shrubs in the Horqin Sandy Land region of northeastern China. Land Degrad Dev 25(3):236–243. https://doi.org/10.1002/ldr.2135
Long RL, Gorecki MJ, Renton M et al (2015) The ecophysiology of seed persistence: a mechanistic view of the journey to germination or demise. Biol Rev 90(1):31–59. https://doi.org/10.1111/brv.12095
Lu YL, Zhang WH, Yang B et al (2019) Seed rain, soil seed bank and seedling regeneration of Quercus aliena var. acureserrata in different slope directions on the middle Qinling Mountains, China. J Appl Ecol 30(6):1965–1973. https://doi.org/10.13287/j.1001-9332.201906.004
Luo X, Cao M, Zhang M et al (2017) Soil seed banks along elevational gradients in tropical, subtropical and subalpine forests in Yunnan Province, southwest China. Plant Divers 39(5):273–286. https://doi.org/10.1016/j.pld.2017.10.001
Ma M, Zhou X, Du G (2011) Soil seed bank dynamics in alpine wetland succession on the Tibetan Plateau. Plant Soil 346:19–28. https://doi.org/10.1007/s11104-011-0790-2
Miao C, Li X, Jia M et al (2016) Spatial structure and species composition of soil seed banks in moving sand dune systems of northeast China. J For Res 27(1):75–80. https://doi.org/10.1007/s11676-015-0166-1
Plue J, De Frenne P, Acharya K et al (2017) Where does the community start, and where does it end? Including the seed bank to reassess forest herb layer responses to the environment. J Veg Sci 28(2):424–435. https://doi.org/10.1111/jvs.12493
Sanou L, Zida D, Savadogo P et al (2018) Comparison of aboveground vegetation and soil seed bank composition at sites of different grazing intensity around a savanna-woodland watering point in West Africa. J Plant Res 131(5):773–788
Shiferaw W, Demissew S, Bekele T (2018) Ecology of soil seed banks: implications for conservation and restoration of natural vegetation: a review. Biodivers Conserv 10:380–393. https://doi.org/10.5897/IJBC2018.1226
Silvertown J, Charlesworth D (2009) Introduction to plant population biology. John Wiley & Sons, New York
Smith RL, Smith TM, Hickman GC et al (1998) Elements of ecology. Menlo Parie, New York
Sprengelmeyer EE, Rebertus AJ (2015) Seed bank dynamics in relation to disturbance and landscape for an ant-dispersed species. Plant Ecol 216(3):371–381
Templeton AR, Levin DA (1979) Evolutionary consequences of seed pools. Am Nat 114(2): 232-249. https://www.jstor.org/stable/2460220
Tesfaye G, Teketay D, Assefa Y et al (2004) The impact of fire on the soil seed bank and regeneration of Harenna forest, Southeastern Ethiopia. Mt Res Dev 24(4):354–361. https://doi.org/10.1659/0276-4741(2004)024[0354:TIOFOT]2.0.CO;2
Thompson K, Band SR, Hodgso JG (1993) Seed size and shape predict persistence in soil. Funct Ecol 7:236–241. https://doi.org/10.2307/2389893
Wandrag EM, Dunham AE, Miller RH et al (2015) Vertebrate seed dispersers maintain the composition of tropical forest seedbanks. AoB Plants 7:plv130. https://doi.org/10.1093/aobpla/plv130
Wang N, Jiao J, Jia Y et al (2017) Influence of afforestation on the species diversity of the soil seed bank and understory vegetation in the hill-gullied loess plateau, China. Int J Environ Res Public Health 14:1285. https://doi.org/10.3390/ijerph14101285
Wang N, He X, Zhao F et al (2020) Soil seed bank in different vegetation types in the Loess Plateau region and its role in vegetation restoration. Restor Ecol 28:A5–A12. https://doi.org/10.1111/rec.13169
Xiang WH, Zhou J, Ouyang S et al (2016) Species-specific and general allometric equations for estimating tree biomass components of subtropical forests in southern China. Eur J Forest Res 135:963–979
Yang X, Baskin CC, Baskin JM et al (2021) Global patterns of potential future plant diversity hidden in soil seed banks. Nat Commun 12(1):1–8. https://doi.org/10.1038/s41467-021-27379-1
Yuan B, Li D, Li J et al (2009) Studies on characteristics of soil seed banks in natural vegetation restoration of the Loess hilly gully regions. Agric Res Arid Area 27(06):215–222
Zhang HW, Yu F, Wang DX et al (2014) Changes of species composition and diversity in the process of community succession of pine oak forests on the south-facing slopes in Qinling Mountains. Acta Botan Boreali-Occiden Sin 34(1):169–176
Zhang ZQ, Shu WS, Lan CY et al (2001) Soil seed bank as an input of seed source in revegetation of lead/zinc mine tailing. Restor Ecol 9(4):378–385. https://doi.org/10.1046/j.1526-100X.2001.94007.x
Zhao Y, Li M, Deng J et al (2021) Afforestation affects soil seed banks by altering soil properties and understory plants on the eastern Loess Plateau, China. Ecol Indic 126:107670. https://doi.org/10.1016/j.ecolind.2021.107670
Acknowledgements
National Natural Science Foundation of China (grant number 32271861) financially supported this work.
Author information
Authors and Affiliations
Contributions
RW, XH analyzed the experimental data and wrote the manuscript; YW collected and analyzed the experimental data; RW, ZL performed the field experiments; DW designed the study; RW, XH, YW, ZL, DW corrected the manuscript.
Corresponding author
Additional information
Responsible Editor: Xinhua He.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Runqin Wu and Xueying Huo contributed equally to this work and should be considered co-first authors.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wu, R., Huo, X., Wang, Y. et al. Soil seed bank characteristics at different developmental stages in pine and oak forests and its potential in vegetation restoration. Plant Soil 497, 535–548 (2024). https://doi.org/10.1007/s11104-023-06410-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06410-y