
Abstract
Background and aims
Dissolved organic matter (DOM) is an active component of the soil organic carbon (SOC) and nitrogen (N) pool. However, the relationship between DOM chemodiversity and microbial communities, and their effects on the C and N in paddy fields under organic or inorganic fertilization, are poorly understood but important for sustainable agricultural production.
Methods
We conducted a six-year experiment in paddy fields under the following different organic and inorganic fertilizer treatments: no fertilizer (CK), chemical fertilizer (150 kg·hm−2 N; CF), 50% N from chemical fertilizer + 50% N from decomposed cattle manure (CMF50), and manure fertilizer (100% N from decomposed cattle manure; MF).
Results
CMF50 exhibited the highest yield (10.89% increase compared with CF). MF and CMF50 enriched the availability and content of soil N, which in turn exhibited a strong effect on net N mineralization and increased SOC and the amount of recalcitrant components of DOM. However, the continuous use of CF led to an increase in both soil bulk density and the number of anaerobic organisms Anaerolinea and Bellilinea. The fractions of DOM containing lignin-like and lipid-like compounds were strongly correlated with Anaerolinea and Bellilinea and contributed to net N mineralization and SOC. Continuous use of CF facilitated net N mineralization but decreased both DOC and SOC.
Conclusions
Continuous applications of CMF50 and MF improved the microbial communities and increased the recalcitrant components of DOM, thus highlighting the importance of the relationship between DOM composition and soil microbial communities while assessing the soil N and SOC.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Paddy fields are a major source of inland dissolved organic matter (DOM) (Bouman et al. 2007). DOM is an active component of the soil organic carbon (SOC) pool, playing a crucial role in biogeochemical cycles on a global scale (Li et al. 2018a; Weiwei et al. 2018). DOM provides soluble organic substrates that sustain microbial growth and activity (Guo et al. 2018). However, the complex molecular composition of DOM has made understanding its relationship with microbial communities challenging, particularly with regards to the carbon (C) and nitrogen (N) cycles in paddy fields under organic and inorganic fertilization (Li et al. 2018b; Yu et al. 2020).
Ultra-high-resolution Fourier-transform ion cyclotron resonance mass spectrometry (FT-ICR-MS) has provided unprecedented resolution of thousands of intact molecular components within DOM. This technique has been successfully applied to marine and terrestrial environments (Li et al. 2018a; Osterholz et al. 2016). However, few studies have focused on using FT-ICR-MS to analyze DOM composition and its association with microbial communities, SOC, and N mineralization in paddy fields (Li et al. 2018b, c). Chemical fertilizers can increase crop yields and soil N mineralization but can also deplete the SOC pool (Alam et al. 2019; Gai et al. 2018; Mulvaney et al. 2009). Organic fertilizer combined with chemical fertilizer may be a potential strategy to enrich soil N and SOC (Rong et al. 2016).
Soil N mineralization is generally limited by microbial N assimilation when microorganisms are exposed to substrates with N limitations (Fujii et al. 2018). In other words, increasing N-containing DOM in N-limited soil might be favorable for soil N mineralization. Therefore, DOM fractions that contain N might serve as a significant source of N to promote the net mineralization of N in paddy soil (Fujii et al. 2018; Wu et al. 2019). N fertilizer application in upland regions decreases the levels of recalcitrant lignin-like components; which may be attributed to the N-containing DOM obtained via microbial metabolism (Li et al. 2018b, d). The depletion of lignin-like components may also lead to an increase in lipid synthesis, which generally provides an energy source for microbes (Chong et al. 2018; He et al. 2022; Lee et al. 2019). However, the content of SOC and recalcitrant lignin-like components of DOM in upland soil can be increased by the long-term application of organic fertilizer (Ghosh et al. 2018; Li et al. 2018d).
N-mining bacteria decompose recalcitrant but N-rich soil organic matter (SOM) pools to obtain more mineralized N (Coonan et al. 2020; Moorhead and Sinsabaugh 2006; Yin et al. 2018). Hence, we hypothesized that the continuous application of organic fertilizer inhibits the decrease of recalcitrant lignin-like fractions, which may lead to a decrease in N-containing DOM fractions, including protein and amino sugar-like fractions that act as important N sources, and lipid-like fractions that may act as a core energy source to stimulate N mineralization by soil microbes when populations are near carrying capacity, thereby inhibiting soil net N mineralization while increasing soil C and N contents in paddy fields. We performed field experiments between 2015 and 2020 in a paddy field exposed to different organic and inorganic fertilizer treatments to test the above hypotheses.
Materials and methods
Study site
The field site was located at the experimental farm of the Foshan Institute of Agricultural Science, Guangdong Province, China (23.52886°N, 112.85709°E, Fig. S1). The region has a subtropical monsoon climate with a mean annual rainfall of 1688 mm and mean minimum and maximum temperatures in summer and winter of 28–34 °C and 13–19 °C, respectively. The soil on the farm is classified as redogenic paddy soil developed from alluvial deposits. The 2015 soil properties were: pH 5.68, organic matter content 22.3 g·kg−1, TN 1.11 g·kg−1, alkali-hydrolyzable N (AN) 133.6 mg·kg−1, available P 41.8 mg·kg−1, and available K 141.0 mg·kg−1.
Experimental design
The continuous fertilization experiment began in 2015 with two Oryza sativa L. crops per year, comprising an early season crop (sown during early to mid-March and harvested between early July and mid-July) and a late season crop (sown during mid-July to late July and harvested between late October and early November). The field site was divided into 12 plots (6 × 4.5 m) with 50 cm buffer zones. Four treatments with three replicates were established in a randomized complete block design, comprising: no fertilizer (CK), chemical fertilizer (150 kg·hm−2 N; CF), 50% N from chemical fertilizer + 50% N from decomposed cattle manure (CMF50), and manure fertilizer (100% N from decomposed cattle manure; MF). Both the early and late season crops were fertilized according to the treatments. The nutrient contents of decomposed cow manure provided are listed in Table S1. Two-thirds of the N, in the form of urea and decomposed cattle manure, was applied as the basal treatment 3 and 10 days before transplanting, respectively. The remaining N was split into two equal parts and applied at the tillering and differentiation stages. Potash and phosphate were applied to maintain the phosphorus (P) and potassium (K) contents in all fertilization treatments at the same level as those in the decomposed cattle manure treatments. A full dose of P and two-thirds of K were applied in the forms of Ca(H2PO4)2·H2O and KCl as basal treatments, with the remaining K applied at the differentiation stage. Irrigation water was applied as per the recommended agronomic practices for this area (Xu et al. 2021). Crop yield was calculated using the following formula:
Where Yieldp was the yield (kg) of rice in each plot of the field site (6 × 4.5 m).
N mineralization
The fresh soil (10 g, dry weight) of each treatment in the paddy field was collected after harvest in the late season of 2020 and placed into 100 ml centrifuge tubes, mixed with distilled water at a ratio of 1:2 (w/v). The centrifuge tube was sealed and cultured at 28 ± 1 ℃. The net N mineralization was obtained by subtracting the initial inorganic N values of the initial samples from those of the samples after 60 days of incubation.
Sampling and analysis
In the 2020 late-season crop harvest, each plot was harvested manually to determine grain yields. Between 2015 and 2020, paddy soil samples were collected from five randomly selected sites in each plot after harvest in the late season by obtaining cores from the surface to a depth of 20 cm. A portion of each soil sample was sieved (< 2 mm) after air-drying, and before analysis, the remaining soil was stored in the refrigerator at -20 ℃ for further biochemical and molecular analysis within a week. The SOC and TN were measured with an elemental analyzer (multi-N/C 2100/2100S, Analytikjena Jena, Jena, Germany). Inorganic N (NH4+-N + NO3−-N) was determined in the subsamples after extraction with 2 M KCl using an automatic N analyzer (FUTURA, Alliance Instruments, Frépillon, France). AN was measured as described by Xiong et al. (2008). DOC was determined as described by Jones and Willett (2006).
Following a methodology used by Li et al. (2018c), DOM was extracted by shaking 60 g of fresh soil in 100 mL Milli-Q water on a reciprocal shaker (170 rpm) at room temperature (25 ℃) for 8 h. The collecting samples and water samples were then centrifuged at 2800 × g for 10 min, and the suspension was filtered through a 0.45-μm mixed cellulose ester membrane. Before the solid-phase extraction (SPE) procedure, SPE cartridges (Bond Elut PPL, 500 mg, 6 mL, Agilent Technologies, Palo Alto, CA, USA) were activated by sequentially rinsing with pure methanol (18 mL) and 0.01 M HCl (18 mL) (Lv et al. 2016). After the DOM samples were adjusted to pH 2, they were passed through the SPE cartridges, followed by Milli-Q water. Finally, the SPE cartridges were dried completely using ultra-pure N2, after which DOM was eluted from the cartridges with methanol (mass spectrometry grade, 10 mL) and stored at -20℃ for FT-ICR-MS analysis. The molecular species of the extracted DOM were analyzed using a 9.4 T Bruker Solarix-XR FT-ICR-MS (Bruker Daltonik, GmbH, Bremen, Germany), equipped with an electrospray ionization source in negative mode, as described by Fang et al. (2017). The sampling rate used was 180 μL/h, the ion accumulation time was 0.7 s, and the detection mass ranged from 150 to 1000 (m/z). The extraction blank and water samples were analyzed and removed from the obtained DOM samples. Mass peaks with S/N (the baseline signal intensity S divided by the noise width N) > 10 were considered; however, P was not included in the formula assignment since HCl was used in the SPE process, which may interfere with the FT-ICR-MS readings for P (Li et al. 2018b). Therefore, the molecular formula assignment for DOM only included elements C, hydrogen (H), oxygen (O), N, and sulfur (S), as P was not considered due to the potential interference from HCl used in the SPE process (Li et al. 2018b).
Total genomic DNA was extracted from each 0.5 g soil sample using an E.Z.N.A.® soil DNA kit (Omega Bio-tek, Norcross, GA, USA), following the manufacturer’s protocols. The quality, integrity, and quantity of each DNA sample were determined by 1% agarose gel electrophoresis and a NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The V3-V4 region of the bacterial 16S ribosomal RNA gene was amplified by PCR using primers 341F (CCTAYGGGRBGCASCAG) and 806R (GGACTACNNGGGTATCTAAT). The PCR procedure was carried out as described by Caporaso et al. (2011). Raw sequence data were de-multiplexed, quality-filtered, and processed using QIIME software (Caporaso et al. 2012). Tags were clustered into operational taxonomic units (OTUs) at 97% sequence similarity by using VSEARCH (Rognes et al. 2016). The OTU table was used to calculate the alpha diversity and provide taxonomic profiles.
Statistics
For each DOM sample, the signal intensity of each identified molecule was normalized to the sum of all intensities of the spectrum obtained from FT-ICR-MS, and the relative intensities (relative abundance) were used for statistical analyses (Osterholz et al. 2016). Biomolecular compound groups of DOM were categorized according to the stoichiometry of their molecular formulas using the aromaticity index (AI) and elemental ratios (H/C and O/C). The labile components (H/C ≥ 1.5) of DOM consist of lipid-like fractions (H/C = 1.5–2.0, O/C = 0–0.3), protein/amino sugar-like fractions (H/C = 1.5–2.2, O/C = 0.3–0.67, N/C ≥ 0.05), and carbohydrate-like fractions (H/C = 1.5–2.0, O/C = 0.67–1.2, comprising sugars CHO and sugars CHOX). Recalcitrant components (H/C < 1.5) consist of lignin-like fractions (H/C = 0.7–1.5, O/C = 0.1–0.67, AI < 0.67, encompassing polyphenolic oligomers), tannin-like fractions (H/C = 0.5–1.5, O/C = 0.67–1.2, AI < 0.67), condensed aromatic-like fractions (H/C = 0.2–0.7, O/C = 0–0.67, AI ≥ 0.67), and unsaturated hydrocarbon-like fractions (H/C = 0.7–1, O/C = 0–0.1, comprising a mixture of aliphatic and aromatic or alkene structures with relatively little oxygen) (Ding et al. 2020; Feng et al. 2016; Huang et al. 2020; Li et al. 2018b; Šantl-Temkiv et al. 2013).
Principal coordinate analysis (PCoA) plots were utilized to evaluate the variations in the composition of microbial communities among treatments and to visualize any potential clustering of treatment effects. Cluster correlation heatmaps with signs and a correlation network (Pearson/Spearman Correlation Coefficient) were generated using the OmicStudio software (https://www.omicstudio.cn) to show correlations between DOM compositions and bacterial community compositions (where P-values < 0.05 and correlation thresholds ≥ 0.6 or ≤ -0.6 showed key linkages). Redundancy analysis (RDA) was used to identify the relationship between the soil N, C, microbial community, and DOM composition and their effects on net N mineralization. Structural equation modeling (SEM) was performed using IBM SPSS AMOS 21.0 to quantify the effects of soil environmental factors, key microorganism, and DOM composition on net N mineralization.
Results
Fertilization increased rice yield and changed soil properties
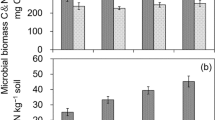
All the fertilization treatments increased the rice yield in 2020, and the continuous application of CMF50 led to a higher yield than that of the CF (10.89%) treatment (Table 1). For the soil properties, continuous application of chemical fertilizer and decomposed cattle manure to the soil in paddy fields from 2015 to 2020 produced significant changes in SOC and TN levels, with a linear relationship between SOC and TN (regression analysis, P < 0.0001, Fig. S2). SOC decreased in the CF treatment compared to the control sample in 2020; however, it increased during the study period in the CMF50 and MF treatments compared to both the control sample and CF treatment (Fig. 1a). The CMF50 and MF treatments both increased TN levels (Fig. 1b). In 2020, soil bulk density levels decreased in the MF treatment, but increased in the CF treatment, which exhibited the lowest soil pH compared to the control sample (Table 1). High-dose application of decomposed cattle manure (CMF50 and MF) decreased net N mineralization; in contrast, the CF treatment enhanced net N mineralization. NO3−-N content following treatment with CF was significantly higher than that in the control sample. The CMF50 and MF treatments showed higher AN, NO3−-N, and NH4+-N content than the control sample, and higher AN and NO3−-N content than the CF treatment.

SOC (a) and TN (b) in paddy soil under four continuous treatments at experimental sites (2015–2020). Treatments: no fertilizer (CK); chemical fertilizer (150 kg·hm−2 N, CF); 50% N from chemical fertilizer + 50% N from decomposed cattle manure (CMF50); manure fertilizer (100% N from decomposed cattle manure; MF). Different lowercase letters indicate significant differences among treatments at P < 0.05
Change in DOM and bacterial composition
DOC varied among the different treatments in 2020. Soil DOC decreased in the CF treatment, whereas it increased significantly in the CMF50 and MF treatments, with the highest DOC value observed after the application of MF (Fig. 2a). The application of MF considerably increased the total number of compounds among DOM, but no significant differences were observed between the CMF50, CF, and control treatments (Fig. 2b). High-dose applications of decomposed cattle manure (CMF50 and MF) decreased the labile DOM compound levels (Fig. 2c); however, CF application decreased levels of recalcitrant DOM components. In each treatment, DOM compound groups were dominated by lignin-like fractions, followed by lipid-like fractions (Fig. 2d). Application of CF decreased levels of condensed aromatic-like fractions while the application of CMF50 decreased levels of carbohydrate-like fractions. Application of MF decreased levels of carbohydrate-like and unsaturated hydrocarbon-like fractions, but increased levels of lignin-like fractions compared with the control sample. Application of CMF50 decreased levels of lipid-like and protein/amino sugar-like fractions, and application of MF decreased levels of unsaturated hydrocarbon-like, protein/amino sugar-like, and lipid-like fractions. However, the application of CMF50 and MF significantly increased levels of lignin-like fractions compared to the CF treatment.

Changes in DOC, DOM composition, and chemodiversity in paddy soils (2020) after six years of continuous experimental treatments. Treatments: no fertilizer (CK); chemical fertilizer (150 kg·hm−2 N, CF); 50% N from chemical fertilizer + 50% N from decomposed cattle manure (CMF50); manure fertilizer (100% N from decomposed cattle manure, MF). (a): DOC; (b): cumulative number of DOM compounds; (c): relative abundance of labile and recalcitrant components of DOM; (d): relative abundance of DOM compound groups. Different lowercase letters indicate significant differences among treatments at P < 0.05. *Indicates significant differences at P < 0.05, t-test
Bacterial communities were influenced by different fertilization treatments in 2020 (Fig. 3a). Application of decomposed cattle manure enhanced the alpha diversity of the soil bacterial community (Shannon index), and the highest values occurred with the application of MF (Fig. S3). The dominant phyla among the treatments were Proteobacteria (20.0%), Chloroflexi (18.87%), Planctomycetes (11.61%), and Acidobacteria (8.85%). The CMF50 and MF application treatments increased the relative abundances of Proteobacteria, Firmicutes, Acidobacteria, Actinobacteria, Candidate.division.NC10 (in MF), and Bacteroidetes (in MF), but decreased the relative abundance of Chloroflexi and Verrucomicrobia compared with the control or CF (Fig. 3b and Table S2).

Distribution of bacterial communities under different fertilizer management treatments in paddy soils (2020) after six years of continuous experimental treatments. (a) Principal coordinate analysis of the structure of bacterial communities. (b) Bacterial composition at phylum level in different treatments. (c) Heat map showing the trend in changes in genera under different fertilization management treatments. Treatments: no fertilizer (CK); chemical fertilizer (150 kg·hm−2 N, CF); 50% N from chemical fertilizer + 50% N from decomposed cattle manure (CMF50); manure fertilizer (100% N from decomposed cattle manure; MF)
At the genus level, 35 abundant genera were present at varying levels in the different treatments after six years (Fig. 3c). CF treatment increased the abundance of Anaerolinea (Chloroflexi) compared with the control (Fig. 3c and Table S3). MF treatment decreased the abundance of Anaerolinea, Bellilinea, and Desulfomonile, but increased the abundance of Anaeromyxobacter, Haliangium, Bryobacter, MND1, Sideroxydans, Pasteuria, Candidatus Nitrotoga, Occallatibacter, Gaiella, and Pedomicrobium compared with CF treatment and the control sample. The CMF50 treatment increased the abundance of Anaeromyxobacter, Bryobacter, and Occallatibacter, compared with the CF treatment and the control sample, but decreased the abundance of Anaerolinea and Bellilinea compared with the CF treatment.
The linkage between DOM composition and bacterial communities
To explore the correlations between DOM and bacterial taxonomic composition, the cut-off |R|≥ 0.6, P < 0.05 was selected to identify the important correlations (Fig. 4). Significant correlations were identified between the OTUs of bacterial phyla and the DOM compound groups. Lipid-like and unsaturated hydrocarbon-like fractions were negatively correlated with Firmicutes, Acidobacteria, Bacteroidetes, and Proteobacteria, and unsaturated hydrocarbon-like fractions were positively correlated with Chloroflexi. Protein/amino sugar-like fractions were negatively correlated with Proteobacteria, Acidobacteria, and Firmicutes, but positively correlated with Nitrospirae and Chloroflexi. Tannin-like fractions were positively correlated with Rokubacteria. Lignin-like fractions were significantly positively correlated with Actinobacteria, Proteobacteria, Firmicutes, Acidobacteria, and Bacteroidetes but negatively correlated with Chloroflexi. The OTUs of bacteria at the genus level showed opposite relationships between labile compounds and recalcitrant DOM components (Fig. 4b). Bryobacter, Pasteuria, Haliangium, Sideroxydans, Occallatibacter, Candidatus Nitrotoga, Gaiella, Steroidobacter, Pseudolabrys, Opitutus, and Pedomicrobium were positively correlated with lignin-like fractions but negatively correlated with labile DOM compounds (such as protein/amino sugar-like and lipid-like fractions). Anaerolinea and Bellilinea were strongly negatively correlated with lignin-like fractions but positively correlated with unsaturated hydrocarbon-like, lipid-like, and protein/amino sugar-like fractions (Anaerolinea).

Clustering correlation heatmap showing significant correlations between DOM composition and bacterial taxa in paddy soils after six years of continuous experimental treatments: (a) phylum level, (b) genus level. *: P < 0.05, **: P < 0.01
Due to the significant correlations between compound groups and bacterial genera, the interconnections between the compositions of each DOM compound group and genera taxa (OTUs) were explored using correlation network analysis (Fig. 4S). The 35 most abundant genera and the 100 most abundant DOM molecules of each compound group were considered for the correlation network analysis. At |R|≥ 0.6, P < 0.05, there were 52 candidate top DOM molecules of lignin-like fractions that were significantly correlated with 13 top genera. Anaerolinea and Bellilinea were correlated with the 40 top molecules, accounting for 80% of the total top molecules of lignin-like fractions. Interestingly, most of the lignin-like molecules with high numbers of C atoms (C ≥ 15) were negatively correlated with the top genus Anaerolinea but positively correlated with Halliangium and Pasteuria. Most of the lignin-like molecules with several C atoms less than 14 were positively correlated with Anaerolinea but negatively correlated with Haliangium, Pasteuria, Sideroxydans, and Bryobacter (Fig. S4a). Unsaturated hydrocarbon-like molecules containing N, with a high double bond equivalence minus oxygen value (DBE-O), showed strong positive correlations with Anaerolinea and Bellilinea, but strong negative correlations with Bryobacter (Fig. S4b and Table S4). Of the labile DOM compounds, protein/amino sugar-like molecules showed negative correlations with the 10 top genera, especially Haliangium, Pasteuria, and Sideroxydans. However, 16 of the 31 protein/amino sugar-like molecules were positively correlated with Anaerolinea (Fig. S4c). The top 35 lipid-like molecules exhibited negative correlations with Bryobacter, Pasteuria, Occallatibacter, Gaiella, and Steroidobacter, whereas 23 of these 35 top molecules exhibited strong positive correlations with Bellilinea, especially Anaerolinea (Fig. S4d).
Soil properties and bacterial communities with respect to net N mineralization
The redundancy analysis (RDA) plot revealed a negative correlation between net N mineralization and NO3−-N, NH4+-N, TN, AN, DOC, and SOC, but a positive correlation with soil bulk density (Fig. 5). Unsaturated hydrocarbon-like fractions and labile DOM compounds, including carbohydrate-like, protein/amino sugar-like, and most notably lipid-like fractions, showed positive effects on net N mineralization; however, the recalcitrant DOM components, especially the lignin-like fractions with a strong positive correlation with SOC, presented a negative correlation with net N mineralization. At the phyla level, Nitrospirae, Verrucomicrobia, and especially the dominant phylum Chloroflexi (which was positively correlated with soil bulk density), exerted a positive effect on net N mineralization, while Actinobacteria, Bacteroidetes, Firmicutes, and the dominant phyla Acidobacteria and Proteogacteria, exerted a negative effect on net N mineralization (Fig. 5a). At the genera level, Anaerolinea and Bellilinea, both with a strong positive correlation with soil bulk density and unsaturated hydrocarbon-like, protein/amino sugar-like and lipid-like fractions, were strongly positively correlated with net N mineralization, while Haliangium, Candidatus Nitrotoga, Pasteuria, Bryobacter, and Occallatibacter were strongly negatively correlated with net N mineralization (Fig. 5b).

RDA of soil properties and bacterial communities with respect to net N mineralization in paddy soils after six years of continuous experimental treatments: (a) phylum level, (b) genus level
To quantify the relative importance of the soil bulk density, DOM composition, SOC, soil N, and key microorganism, SEM analysis was performed to simultaneously assess and quantify the relative contributions of major factors on the net N mineralization (Fig. S5). Soil TN and AN indirectly or directly affected the abundance of genera Anaerolinea and Bellilinea among Chloroflexi. Notably, the soil bulk density played the most important role in affecting the abundance of genera Anaerolinea and Bellilinea, which affected the accumulation of different compositions of DOM. Anaerolinea and Bellilinea, with a direct negative effect (-0.74) on the levels of lignin-like fractions, exhibited direct positive effects on the levels of lipid-like (0.88), protein/amino sugar-like (0.54), and unsaturated hydrocarbon-like fractions (0.62), but showed no significant direct effects on the levels of carbohydrate-like fractions (0.24). Compared with carbohydrate-like fractions, lignin-like, lipid-like, protein/amino sugar-like, and unsaturated hydrocarbon-like fractions exhibited stronger direct effects on net N mineralization. Notably, lipid-like (0.84) and lignin-like (-0.74) fractions exhibited a dominant effect on net N mineralization. Net N mineralization exhibited a strong effect (-0.77 to -0.60) on the content of SOC, which in turn exhibited a strong effect (0.79 to 0.86) on the content of TN.
Discussion
Variation of soil properties and rice yield
Continuous application of CMF50 and MF increased TN (ranging from 1.07 to 1.41 g·kg−1 and 1.08 to 1.61 g·kg−1, respectively) and SOC (ranging from 11.12 to 14.21 g·kg−1 and 11.90 to 15.95 g·kg−1, respectively) in paddy soil considerably during the study period (Fig. 1). These findings are consistent with those of previous studies that reported that organic fertilizer, either alone or combined with chemical fertilizer, continuously enhanced the capacity of soil to retain carbon and supply N in paddy fields (Bai et al. 2018; Coonan et al. 2020; Gai et al. 2018). Moreover, the application of high doses of decomposed cattle manure (CMF50 and MF) enhanced the AN and NO3−-N content of soil compared with the control sample and CF treatment (Table 1). A possible explanation is that the addition of decomposed cattle manure reduced N losses (Hou et al. 2018). These findings also explained why the CMF50 exhibited a higher rice yield than the CF treatment.
Continuous application of CF decreased the soil pH, which was consistent with the findings of Jin et al. (2019). The application of CF increased net N mineralization but decreased SOC in paddy soils, which might be attributed to microbial degradation in native SOM to obtain more mineralized N (Alam et al. 2019; Roba 2018). The application of MF decreased soil bulk density, but the application of CF increased soil bulk density. This finding might be associated with organic components, which have a dilution effect on bulk density (Celik et al. 2010).
Effects on DOM and bacterial composition
DOM is the most active part of the SOC pool. The application of CMF50 and MF significantly increased the content of soil DOC; by 2020, the application of MF notably enhanced the cumulative number of identified DOM compounds (Fig. 2a and b). This can be attributed to the decomposed cattle manure increasing the total organic matter reservoirs, resulting in higher DOM composition (Li et al. 2018d, 2019). However, the decrease in DOC and recalcitrant DOM components, including condensed aromatic-like fractions, in the CF treatment might result from the higher mineralization of organic materials compared with the control sample (Alam et al. 2019).
Based on microbial substrate preferences and trophic strategy, the increase in Proteobacteria, Acidobacteria, Firmicutes, and Bacteroidetes (in MF) in the CMF50 and MF treatments were shown previously to conform with the characteristics of fast-growth microorganisms and r-strategists (Jeewani et al. 2020; Mickan et al. 2019). Application of CMF50 and MF increased the alpha diversity of the soil bacterial community (Fig. S3) and especially increased the abundance of genera among the r-strategists Proteobacteria and Actinobacteria, such as MND1, Sideroxydans, Haliangium, Pedomicrobium, and Candidatus Nitrotoga in MF, and Anaeromyxobacter, Bryobacter, and Occallatibacter in CMF50 and MF, respectively. Therefore, r-strategists need to take advantage of labile C to maintain their fast growth rate and metabolism, thus resulting in the reduced relative abundance of labile DOM compounds (such as carbohydrate-like fractions) after the application of MF and CMF50 compared with the control sample (Ling et al. 2021; Razanamalala et al. 2018). However, the abundance of unsaturated hydrocarbon-like fractions containing N (Fig. 2d and S4) decreased in the MF treatment, suggesting that unsaturated hydrocarbon-like fractions among recalcitrant DOM components might be relatively vulnerable to microbial decomposition. It has been reported that Chloroflexi, as K-strategists, can grow on lignocellulosic substrates and are replete with aromatic compound degradation genes to degrade recalcitrant DOM compounds (Colatriano et al. 2018; Wong et al. 2016). Application of CMF50 and MF suppressed the growth of Chloroflexi, in particular, the anaerobic organisms Anaerolinea and Bellilinea, which led to higher levels of recalcitrant DOM components in paddy soils compared with CF application or the control sample (Fig. 3b). This finding indicates that Anaerolinea and Bellilinea could be the dominant genera among Chloroflexi that cause recalcitrant C degradation. We suggest that stoichiometrically-balanced, supplementary, decomposed cattle manure, either alone or combined with CF, could cut fertilizer use while enhancing the sequestration of C into SOM in paddy soil.
We observed an increase in the abundance of protein/amino sugar-like, lipid-like, and unsaturated hydrocarbon-like (compared with MF) fractions and a decrease in the abundance of lignin-like fractions following continuous application of CF, compared with the application of CMF50 and MF. This might be due to continuous chemical-N input allowing the growth of K-strategist bacteria, which in turn may increase the consumption of recalcitrant lignin-like fractions while synthesizing and supporting more lipid-like, protein/amino sugar-like or unsaturated hydrocarbon-like substrates when populations are near carrying capacity (Devi et al. 2012; He et al. 2022; Li et al. 2018b; Shi et al. 2017; Yin et al. 2018). Therefore, the protein/amino sugar-like, unsaturated hydrocarbon-like, and lipid-like fractions might act as important N sources or energy for K-strategists.
Responses of bacterial communities to DOM composition
We found correlations between DOM molecular distribution and bacterial taxonomy under various continuous fertilization treatments (Fig. 4). Recalcitrant lignin-like fractions showed strong negative correlations with Chloroflexi, consistent with the findings of Li et al. (2021). Chloroflexi can utilize recalcitrant C, such as aryl-C, which is found in lignin (Boye et al. 2017; Colatriano et al. 2018). Anaerolinea and Bellilinea were correlated with 80% of the 52 top lignin-like molecules (Fig. 4Sa), which suggests that they may be the dominant genera decomposing lignin-like fractions among all Chloroflexi in the paddy soil. Interestingly, most lignin-like molecules with high numbers of C atoms showed negative correlations with the relative abundance of Anaerolinea, while having a positive correlation with lignin-like molecules with < 14 C atoms. This indicates that Anaerolinea might prefer to decompose high-molecular-weight lignin-like molecules into low-molecular-weight molecules. Unsaturated hydrocarbon-like fractions containing N and labile DOM compounds, including lipid-like and protein/amino sugar-like fractions, showed positive correlations with both Anaerolinea and Bellilinea (Fig. 5b, c, e, and S4). This could be attributed to Anaerolinea and Bellilinea favoring lignin-like decomposition, and thus taking advantage of labile C and organic N as their energy and growth sources, respectively (Kirk et al. 1976; Moorhead and Sinsabaugh 2006). Although Actinobacteria, Proteobacteria, Firmicutes, Acidobacteria, and Bacteroidetes have been characterized as r-strategists, some strains within these phyla are well known for their ability to decompose or grow in lignin (Lee et al. 2019). Bryobacter, Occallatibacter (Acidobacteria), Pasteuria (Firmicutes), Haliangium, Sideroxydans, Candidatus Nitrotoga (Proteobacteria), Gaiella (Actinobacteria), Steroidobacter, Pseudolabrys, Opitutus, and Pedomicrobium (Proteobacteria) were positively correlated with lignin-like fractions, reflecting that some genera within Actinobacteria, Proteobacteria, Firmicutes, and Acidobacteria are capable of growing in lignin-like fractions (Li et al. 2019; Ling et al. 2021). Our results suggested that DOM composition influenced the change of bacterial composition; thus, labile and recalcitrant C utilization by bacteria.
Soil properties and bacterial communities associated with net N mineralization
RDA and SEM analyses showed that soil N availability and TN exhibited a strong effect on net N mineralization and was strongly positively correlated with SOC, DOC, lignin-like fractions, and some bacterial communities. These findings revealed that soil N in paddy soils might be an important factor influencing the microbial community by utilizing C as an energy source to mineralize DOC and obtain inorganic N (Coonan et al. 2020; Craine et al. 2007). Notably, soil bulk density was one of the dominant factors that influenced the bacterial community; specifically, Anaerolinea, Bellilinea (anaerobic organisms, Chloroflexi), Haliangium, Candidatus Nitrotoga (Proteobacteria), Pasteuria (Firmicutes), Bryobacter, and Occallatibacter (Acidobacteria) were found to have strong correlations with soil bulk density and net N mineralization. The net N mineralization exhibited a strong direct effect (− 0.77 to − 0.60) on the content of SOC, which exhibited a strong direct effect on the content of TN (0.79 to 0.86). These findings indicated that the soil N and C content were influenced by the net N mineralization condition in the paddy fields.
Lignin-like fractions were the dominant component of recalcitrant C under anoxic conditions (paddy fields) and exhibited a dominant negative effect (direct effect: -0.74) on net N mineralization (O'Donnell et al. 2016; Wang et al. 2016). These findings indicated the lignin-like fractions might be the dominant component of DOM that inhibit the net N mineralization in paddy soils. However, the lignin-like fractions showed a negative correlation with protein/amino sugar-like, unsaturated hydrocarbon-like, and most notably lipid-like fractions, which was consistent with the findings of Ly and Hur (2018). These findings indicated that the decrease in lignin-like fractions could lead to an increase in protein/amino sugar-like, unsaturated hydrocarbon-like, and lipid-like fractions. Genera Anaerolinea and Bellilinea exhibited a strong direct effect on protein/amino sugar-like and unsaturated hydrocarbon-like fractions. Both of them exhibited a strong effect on the net mineralization of soil N. However, the levels of NO3−-N, NH4+-N, TN, and AN exhibited a negative effect on the net mineralization of soil N. These findings indicated that protein/amino sugar-like, and unsaturated hydrocarbon-like fractions containing N (Fig. S4d), might be important sources of organic N, while the content and availability of soil N were the main factors stimulating bacteria to favor soil organic N (Ekenler and Tabatabai 2004; Fujii et al. 2018). Anaerolinea and Bellilinea exhibited no significant direct effects (0.24) on carbohydrate-like fractions but showed a strong direct effect (0.88) on lipid-like fractions. Carbohydrate-like fractions exhibited a weak positive effect (direct effect: 0.47) on net N mineralization, whereas lipid-like fractions showed a strong positive contribution (direct effect: 0.84) to net N mineralization. These findings indicated that lipid-like fractions might become a major source of C, acting as an energy source for N-mining bacteria (Chong et al. 2018; Sebastián et al. 2016). Our data supports the microbial N mining hypothesis, where the availability and content of soil N cause a priming effect to decompose recalcitrant DOM components but allow more energy sources and the availability of more N-rich DOM for N mining by bacteria.
Conclusion
The continuous application of CMF50 for six years resulted in the highest rice yield. The applications of both MF and CMF50 increased the soil N content and availability but decreased the net N mineralization. MF and CMF50 increased SOC, whereas CF decreased SOC, which was associated with the labile and recalcitrant components of DOM. The decrease in lignin-like fractions led to an increase in the levels of lipid-like fractions and the fractions of DOM containing N, which were strongly correlated with Anaerolinea and Bellilinea and contributed to net N mineralization and SOC.
The findings of this study suggested that the composition of DOM, soil N-driven bacterial carbon-acquisition strategy, and net N mineralization in paddy fields respond to fertilization treatments. These findings have the potential to aid in predicting soil’s ability to retain C and supply N, as well as its capacity to support rice production.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AI:
-
Aromaticity index
- AN:
-
Alkali-hydrolysable N
- CF:
-
Chemical fertilizer
- CK:
-
No fertilizer
- CMF50:
-
50% N from chemical fertilizer + 50% N from decomposed cattle manure
- DBE-O:
-
Double bond equivalence minus oxygen value
- DOC:
-
Dissolved organic carbon
- DOM:
-
Dissolved organic matter
- FT-ICR-MS:
-
Fourier-transform ion cyclotron resonance mass spectrometry
- MF:
-
Manure fertilizer
- OTU:
-
Operational taxonomic units
- SOC:
-
Soil organic carbon
- RDA:
-
Redundancy analysis
- SPE:
-
Solid-phase extraction
- TN:
-
Total nitrogen
References
Alam M, Rahman M, Biswas JC, Akhter S, Maniruzzaman M, Choudhury AK, Jahan MAHS, Miah MMU, Sen R, Kamal MZU, Mannan MA, Shiragi HK, Kabir W, Kalra N (2019) Nitrogen transformation and carbon sequestration in wetland paddy field of Bangladesh. Paddy Water Environ 17:677–688. https://doi.org/10.1007/s10333-019-00693-7
Bai Z, Caspari T, Gonzalez MR, Batjes NH, Mäder P, Bünemann EK, de Goede R, Brussaard L, Xu M, Ferreira CSSF, Reintam E, Fan H, Mihelič R, Glavan M, Tóth Z (2018) Effects of agricultural management practices on soil quality: a review of long-term experiments for Europe and China. Agric Ecosyst Environ 265:1–7. https://doi.org/10.1016/j.agee.2018.05.028
Bouman M, Lampayan RM, Tuong TP (2007) Coping with water scarcity. In: Hardy B (ed) Water Management in Irrigated Rice. International Rice Research Institute, Los Baños, pp 17–22
Boye K, Noël V, Tfaily MM, Bone SE, Williams KH, Bargar JR, Fendorf S (2017) Thermodynamically controlled preservation of organic carbon in floodplains. Nat Geosci 10:415–419. https://doi.org/10.1038/ngeo2940
Caporaso JG, Lauber CL, Walters WA, Berglyons D, Huntley J, Fierer N, Owens SM, Betley J, Fraser L, Bauer M (2012) Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J 6:1621. https://doi.org/10.1038/ismej.2012.8
Caporaso JG, Lauber CL, Walters WA, Lyons DB, Lozupone CA, Turnbaugh PJ, Fierer N, Knight R (2011) Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. PNAS 108:4516–4522. https://doi.org/10.1073/pnas.1000080107
Celik I, Gunal H, Budak M, Akpinar C (2010) Effects of long-term organic and mineral fertilizers on bulk density and penetration resistance in semi-arid Mediterranean soil conditions. Geoderma 160:236–243. https://doi.org/10.1016/j.geoderma.2010.09.028
Chong G, Huang X, Di J, Xu D, He Y, Pei Y, Tang Y, Ma C (2018) Biodegradation of alkali lignin by a newly isolated Rhodococcus pyridinivorans CCZU-B16. Bioproc Biosyst Eng 41:501–510. https://doi.org/10.1007/s00449-017-1884-x
Colatriano D, Tran PQ, Guéguen C, Williams WJ, Lovejoy C, Walsh DA (2018) Genomic evidence for the degradation of terrestrial organic matter by pelagic Arctic Ocean Chloroflexi bacteria. Commun Biol 1:1–9. https://doi.org/10.1038/s42003-018-0086-7
Coonan EC, Kirkby CA, Kirkegaard JA, Amidy MR, Strong CL, Richardson AE (2020) Microorganisms and nutrient stoichiometry as mediators of soil organic matter dynamics. Nutr Cycl Agroecosys 117:273–298. https://doi.org/10.1007/s10705-020-10076-8
Craine JM, Morrow C, Fierer N (2007) Microbial nitrogen limitation increases decomposition. Ecology 88:2105–2113. https://doi.org/10.1890/06-1847.1
Devi MP, Subhash GV, Mohan SV (2012) Heterotrophic cultivation of mixed microalgae for lipid accumulation and wastewater treatment during sequential growth and starvation phases: effect of nutrient supplementation. Renew Energy 43:276–283. https://doi.org/10.1016/j.renene.2011.11.021
Ding Y, Shi Z, Ye Q, Liang Y, Liu M, Dang Z, Wang Y, Liu C (2020) Chemodiversity of soil dissolved organic matter. Environ Sci Technol 54:6174–6184. https://doi.org/10.1021/acs.est.0c01136
Ekenler M, Tabatabai M (2004) β-Glucosaminidase activity as an index of nitrogen mineralization in soils. Commun Soil Sci Plan 35:1081–1094. https://doi.org/10.1081/CSS-120030588
Fang Z, He C, Li Y, Chung KH, Xu C, Shi Q (2017) Fractionation and characterization of dissolved organic matter (DOM) in refinery wastewater by revised phase retention and ion-exchange adsorption solid phase extraction followed by ESI FT-ICR MS. Talanta 162:466–473. https://doi.org/10.1016/j.talanta.2016.10.064
Feng L, Xu J, Kang S, Li X, Li Y, Jiang B, Shi Q (2016) Chemical composition of microbe-derived dissolved organic matter in cryoconite in Tibetan plateau glaciers: insights from fourier transform ion cyclotron resonance mass spectrometry analysis. Environ Sci Technol 50:13215. https://doi.org/10.1021/acs.est.6b03971
Fujii K, Yamada T, Hayakawa C, Nakanishi A, Funakawa S (2018) Another bottleneck for nitrogen mineralization in temperate forest soils: arginine metabolism in microorganisms. Soil Biol Biochem 126:22–30. https://doi.org/10.1016/j.soilbio.2018.08.005
Gai X, Liu H, Liu J, Zhai L, Yang B, Wu S, Ren T, Lei Q, Wang H (2018) Long-term benefits of combining chemical fertilizer and manure applications on crop yields and soil carbon and nitrogen stocks in North China Plain. Agric Water Manag 208:384–392. https://doi.org/10.1016/j.agwat.2018.07.002
Ghosh A, Bhattacharyya R, Meena MC, Dwivedi BS, Singh G, Agnihotri R, Sharma C (2018) Long-term fertilization effects on soil organic carbon sequestration in an Inceptisol. Soil till Res 177:134–144. https://doi.org/10.1016/j.still.2017.12.006
Guo P, Zhang C, Wang Y, Yu X, Zhang Z, Zhang D (2018) Effect of long-term fertilization on humic redox mediators in multiple microbial redox reactions. Environ Pollut 234:107–114. https://doi.org/10.1016/j.envpol.2017.10.106
He C, Wang H, Peng X, Li H (2022) Recent biotechnology advances in bio-conversion of lignin to lipids by bacterial cultures. Front Chem 393. https://doi.org/10.3389/fchem.2022.894593
Hou Y, Wei S, Ma W, Roelcke M, Nieder R, Shi S, Wu J, Zhang F (2018) Changes in nitrogen and phosphorus flows and losses in agricultural systems of three megacities of China, 1990–2014. Resour Conserv Recy 139:64–75. https://doi.org/10.1016/j.resconrec.2018.07.030
Huang M, Li Z, Chen M, Wen J, Luo N, Xu W, Ding X, Xing W (2020) Dissolved organic matter released from rice straw and straw biochar: contrasting molecular composition and lead binding behaviors. Sci Total Environ 739:140378. https://doi.org/10.1016/j.scitotenv.2020.140378
Jeewani PH, Gunina A, Tao L, Zhu Kuzyakov Y, Van Zwieten L, Guggenberger G, Shen C, Yu G, Singh BP, Pan S, Luo Y, Xu J (2020) Rusty sink of rhizodeposits and associated keystone microbiomes. Soil Biol Biochem 147:107840. https://doi.org/10.1016/j.soilbio.2020.107840
Jin Z, Chen C, Chen X, Jiang F, Hopkins I, Zhang X, Han Z, Billy G, Benavides J (2019) Soil acidity, available phosphorus content, and optimal biochar and nitrogen fertilizer application rates: a five-year field trial in upland red soil, China. Field Crop Res 232:77–87. https://doi.org/10.1016/j.fcr.2018.12.013
Jones DL, Willett VB (2006) Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol Biochem 38:991–999. https://doi.org/10.1016/j.soilbio.2005.08.012
Kirk TK, Connors WJ, Zeikus JG (1976) Requirement for a growth substrate during lignin decomposition by two wood-rotting fungi. Appl Environ Microb 32:192–194. https://doi.org/10.1128/aem.32.1.192-194.1976
Lee S, Kang M, Bae JH, Sohn JH, Sung BH (2019) Bacterial valorization of lignin: strains, enzymes, conversion pathways, biosensors, and perspectives. Front Bioeng Biotech 209. https://doi.org/10.3389/fbioe.2019.00209
Li HY, Wang H, Wang HT, Xin PY, Xu XH, Ma Y, Liu WP, Ten CY, Jiang CL, Lou LP, Arnold W, Cralle L, Zhu YG, Chu JF, Gilbert JA, Zhang ZJ (2018a) The chemodiversity of paddy soil dissolved organic matter correlates with microbial community at continental scales. Microbiome 6:1–16. https://doi.org/10.1186/s40168-018-0561-x
Li M, Sun G, Chen S, Fang Z, Yun H, Shi Q, Zhu Y (2018b) Molecular chemodiversity of dissolved organic matter in paddy soils. Environ Sci Technol 52:963–971. https://doi.org/10.1021/acs.est.7b00377
Li J, Wen Y, Li X, Li Y, Yang X, Lin Z, Song Z, Cooper JM, Zhao B (2018c) Soil labile organic carbon fractions and soil organic carbon stocks as affected by long-term organic and mineral fertilization regimes in the North China Plain. Soil till Res 175:281–290. https://doi.org/10.1016/j.still.2017.08.008
Li X, Chen Q, He C, Shi Q, Chen S, Reid BJ, Zhu Y, Sun G (2018d) Organic carbon amendments affect the chemodiversity of soil dissolved organic matter and its associations with soil microbial communities. Environ Sci Technol 53:50–59. https://doi.org/10.1021/acs.est.8b04673
Li Y, Nie C, Liu Y, Du W, He P (2019) Soil microbial community composition closely associates with specific enzyme activities and soil carbon chemistry in a long-term nitrogen fertilized grassland. Sci Total Environ 654:264–274. https://doi.org/10.1016/j.scitotenv.2018.11.031
Li Y, Heal K, Wang S, Cao S, Zhou C (2021) Chemodiversity of soil dissolved organic matter and its association with soil microbial communities along a chronosequence of Chinese Fir monoculture plantations. Front Microbiol 12:729344. https://doi.org/10.3389/fmicb.2021.729344
Ling L, Fu Y, Jeewani PH, Tang C, Pan S, Reid BJ, Gunina A, Li Y, Li Y, Cai Y, Kuzyakov Y, Li Y, Su W, Singh BP, Luo Y, Xu J (2021) Organic matter chemistry and bacterial community structure regulate decomposition processes in post-fire forest soils. Soil Biol Biochem 160:108311. https://doi.org/10.1016/j.soilbio.2021.108311
Lv J, Zhang S, Wang S, Luo L, Cao D, Christie P (2016) Molecular-scale investigation with ESI-FT-ICR-MS on fractionation of dissolved organic matter induced by adsorption on iron oxyhydroxides. Environ Sci Technol 50:2328–2336. https://doi.org/10.1021/acs.est.5b04996
Ly QV, Hur J (2018) Further insight into the roles of the chemical composition of dissolved organic matter (DOM) on ultrafiltration membranes as revealed by multiple advanced DOM characterization tools. Chemosphere 201:168–177. https://doi.org/10.1016/j.chemosphere.2018.02.181
Mickan BS, Abbott LK, Solaiman ZM, Mathes F, Siddique KH, Jenkins SN (2019) Soil disturbance and water stress interact to influence arbuscular mycorrhizal fungi, rhizosphere bacteria and potential for N and C cycling in an agricultural soil. Biol Fert Soils 55:53–66. https://doi.org/10.1007/s00374-018-1328-z
Moorhead DL, Sinsabaugh RL (2006) A theoretical model of litter decay and microbial interaction. Ecol Monogr 76:151–174. https://doi.org/10.1890/0012-9615(2006)076[0151:ATMOLD]2.0.CO;2
Mulvaney RL, Khan SA, Ellsworth TR (2009) Synthetic nitrogen fertilizers deplete soil nitrogen: a global dilemma for sustainable cereal production. J Environ Qual 38:2295–2314. https://doi.org/10.2134/jeq2008.0527
O’Donnell JA, Aiken GR, Butler KD, Guillemette F, Podgorski DC, Spencer RGM (2016) DOM composition and transformation in boreal forest soils: the effects of temperature and organic-horizon decomposition state. J Geophys Res-Biogeo 121:2727–2744. https://doi.org/10.1002/2016JG003431
Osterholz H, Singer G, Wemheuer B, Daniel R, Simon M, Niggemann J, Dittmar T (2016) Deciphering associations between dissolved organic molecules and bacterial communities in a pelagic marine system. ISME J 10:1717–1730. https://doi.org/10.1038/ismej.2015.231
Razanamalala K, Razafimbelo T, Maron P, Ranjard L, Chemidlin N, Lelièvre M, Dequiedt S, Ramaroson VH, Marsded C, Becquer T, Trap J, Blanchart E, Bernard L (2018) Soil microbial diversity drives the priming effect along climate gradients: a case study in Madagascar. ISME J 12:451–462. https://doi.org/10.1038/ismej.2017.178
Roba T (2018) Review on: the effect of mixing organic and inorganic fertilizer on productivity and soil fertility. OALib J 5:1–11. https://doi.org/10.4236/oalib.1104618
Rognes T, Flouri T, Nichols B, Quince C, Mahe F (2016) VSEARCH: a versatile open source tool for metagenomics. PeerJ 4:e2584. https://doi.org/10.7717/peerj.2584
Rong Y, Su Y, Tao W, Qin Y (2016) Effect of chemical and organic fertilization on soil carbon and nitrogen accumulation in a newly cultivated farmland. J Integr Agr 15:658–666. https://doi.org/10.1016/S2095-3119(15)61107-8
Šantl-Temkiv T, Finster K, Dittmar T, Hansen BM, Thyrhaug R, Nielsen NW, Karlson UG (2013) Hailstones: a window into the microbial and chemical inventory of a storm cloud. PLoS One 8:e53550. https://doi.org/10.1371/journal.pone.0053550
Sebastián M, Smith AF, González JM, Fredricks HF, Van Mooy B, Koblížek M, Brandsma J, Koster G, Mestre M, Mostajir B, Pitta P, Postle AD, Sánchez P, Gasol JM, Scanlan DJ, Chen Y (2016) Lipid remodelling is a widespread strategy in marine heterotrophic bacteria upon phosphorus deficiency. ISME J 10:968–978. https://doi.org/10.1038/ismej.2015.172
Shi K, Gao Z, Shi TQ, Song P, Ren L, Huang H, Ji X (2017) Reactive oxygen species-mediated cellular stress response and lipid accumulation in oleaginous microorganisms: the state of the art and future perspectives. Front Microbiol 8:793. https://doi.org/10.3389/fmicb.2017.00793
Wang J, Xiong Z, Kuzyakov Y (2016) Biochar stability in soil: meta-analysis of decomposition and priming effects. Glob Chang Biol 8:512–523. https://doi.org/10.1111/gcbb.12266
Weiwei L, Xin Y, Keqiang S, Baohua Z, Gao G (2018) Unraveling the sources and fluorescence compositions of dissolved and particulate organic matter (DOM and POM) in Lake Taihu, China. Environ Sci Pollu R 26:4027–4040. https://doi.org/10.1007/s11356-018-3873-2
Wong MT, Wang W, Lacourt M, Couturier M, Edwards EA, Master ER (2016) Substrate-driven convergence of the microbial community in lignocellulose-amended enrichments of gut microflora from the Canadian beaver (Castor canadensis) and North American moose (Alces americanus). Front Microbiol 7:961. https://doi.org/10.3389/fmicb.2016.00961
Wu H, Du S, Zhang Y, An J, Zou H, Zhang Y, Yu N (2019) Effects of irrigation and nitrogen fertilization on greenhouse soil organic nitrogen fractions and soil-soluble nitrogen pools. Agr Water Manag 216:415–424. https://doi.org/10.1016/j.agwat.2019.02.020
Xiong Y, Xia H, Li ZA, Cai XA, Fu S (2008) Impacts of litter and understory removal on soil properties in a subtropical Acacia mangium plantation in China. Plant Soil 304:179–188. https://doi.org/10.1007/s11104-007-9536-6
Xu Y, Feng J, Li H (2021) How intercropping and mixed systems reduce cadmium concentration in rice grains and improve grain yields. J Hazard Mater 402:123762. https://doi.org/10.1016/j.jhazmat.2020.123762
Yin L, Dijkstra FA, Wang P, Zhu B, Cheng C (2018) Rhizosphere priming effects on soil carbon and nitrogen dynamics among tree species with and without intraspecific competition. New Phytol 218:1036–1048. https://doi.org/10.1111/nph.15074
Yu Q, Hu X, Ma J, Ye J, Sun W, Wang Q, Lin H (2020) Effects of long-term organic material applications on soil carbon and nitrogen fractions in paddy fields. Soil Till Res 196:104483. https://doi.org/10.1016/j.still.2019.104483
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 42107050), the Talent Introduction Project of Guangdong Academy of Agricultural Sciences (Grant No. R2021YJ-YB2003), the Guangzhou Basic Research Project (Grant No. 2023A04J0788), the Science and Technology Planning Project of Guangdong Province (Grant No. 2017A020208024), the National Key Research and Development Program of China (Grant No. 2016YFD0200402), and the “Jinying Star” discipline leader training plan of the Guangdong Academy of Agricultural Sciences (Grant No. R2020PY-JX021).
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 42107050), the Talent Introduction Project of Guangdong Academy of Agricultural Sciences (Grant No. R2021YJ-YB2003), the Guangzhou Basic Research Project (Grant No. 2023A04J0788), the Science and Technology Planning Project of Guangdong Province (Grant No. 2017A020208024), the National Key Research and Development Program of China (Grant No. 2016YFD0200402), and the “Jinying Star” discipline leader training plan of the Guangdong Academy of Agricultural Sciences (Grant No. R2020PY-JX021).
Author information
Authors and Affiliations
Contributions
Yanggui Xu: Methodology, Formal analysis, Investigation, Writing—Original Draft, Writing—Review & Editing. Zhiping Peng: Conceptualization, Validation, Investigation, Resources. Yuting Tu: Formal analysis, Investigation. Jichuan Huang: Conceptualization, Validation, Resources, Writing—Review & Editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Hans Lambers.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Xu, Y., Peng, Z., Tu, Y. et al. Combining organic and inorganic fertilization increases rice yield and soil nitrogen and carbon: dissolved organic matter chemodiversity and soil microbial communities. Plant Soil 492, 557–571 (2023). https://doi.org/10.1007/s11104-023-06203-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06203-3