
Abstract
Background
Agroforestry systems have enhanced diversity of cultivated plants compared to monocultures, and are expected to affect associated biodiversity. Despite a growing body of literature on the importance of soil fauna, the known effects of different agroforestry types on soil fauna communities and functions have not yet been synthesized.
Scope
We scanned publications on soil fauna in agroforestry systems. Our aim was to give an overview of strengths and weaknesses of the existing data, in terms of spatial coverage and representation of diverse agroforestry types and soil fauna groups and functions.
Conclusions
Our database includes sixty-seven articles, mostly focusing on tropical regions and perennial crop agroforestry systems. Soil macrofauna are the most studied fauna group. The most common question addressed is the comparison of the effect of land use types on communities. Effects on fauna abundance and diversity are mainly positive when agroforestry is compared to cropland, and neutral or negative when compared to forests. Few publications actually measure soil fauna functions, or characterize their interactions and evolution in time and space depending on system design and management. Further work on soil fauna in agroforestry should harness ecological theory and address questions of spatial structure and scale, temporal dynamics and ecological interaction networks and how they determine ecosystem functioning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Agroforestry systems are examples of mixed species cropping systems that increase “planned biodiversity” compared to monocultures, with expected effects on “associated biodiversity” (Vandermeer et al. 1998; Malézieux et al. 2009) or “unplanned” diversity of microorganisms, plants and animals. Agroforestry, broadly defined as tree presence in agricultural landscapes is of high significance worldwide, with an estimated 43% of global agricultural land (ranging from 9% in North Africa and Western Asia to more than 95% in Central America) affected by a tree cover of more than 10% (Zomer et al. 2014). It can be expected to have specific effects on soil biodiversity (Barrios et al. 2018). Firstly, trees provide specific habitats due to their perennial above- and below-ground structures. Secondly, they have a strong impact on the heterogeneity of plot microclimate as has been widely documented (e.g. Monteith et al. 1991; Hawke and Wedderburn 1994; Singh et al. 2012). Temperature variations are buffered under tree canopies, with lower maxima due to radiation interception, higher minima and a lower average temperature than outside tree influence (Chen et al. 1999). Soil moisture and air humidity are generally higher in presence of tree cover due to reduced temperatures and therefore reduced evaporative demand, and soil cover due to tree litter and wind attenuation (Vigiak et al. 2003) that reduce soil evaporation rate. These modifications of microclimate vary seasonally according to tree phenology and morphology, and can modify the associated biodiversity and fauna activity by habitat diversification (e.g. Martin-Chave et al. 2019). Thirdly, the presence of trees, and sometimes of associated grass strips or patches, leads to local concentrations of organic resources from above-ground and below-ground litter production, leading to local increases in soil organic carbon (e.g. Cardinael et al. 2017) and nutrient contents (Palm 1995; Young 1997). Soil microbial functioning is modified according to distance from trees as shown by Guillot et al. (2019), likely due to these spatial gradients in organic resources and in their biochemical quality. Soil structure and aggregate stability can be improved under tree lines (Udawatta et al. 2008), no doubt leading to spatial heterogeneity in soil microhabitat characteristics. Biodiversity of adventitious plants and soil organisms can be expected to respond to such gradients. Finally, above-ground tree litter can also provide soil cover and associated microhabitats and refuges for epigeic soil organisms.
Soil fauna play important roles in agroecosystems, as crucial actors of the four aggregated and interrelated functions that determine soil health (Kibblewhite et al. 2008). Microbivorous microfauna (in particular nematodes, protozoa) and detritivorous meso- and macrofauna (collembola, acari, diplopoda, earthworms, various insects...) modulate and/or complement the activity of saprotrophic microorganisms to determine carbon transformations in the soil. Through the same process, these organisms are also strongly involved in nutrient cycling. The soil “ecosystem engineer” macrofauna (earthworms, ants, termites) play a major role in soil structure maintenance (Jouquet et al. 2006), while many biocontrollers from the different size categories (microbivorous and carnivorous nematodes, collembola, predatory mites, ground or rove beetles, and centipedes, for instance) are involved in biological regulation. The effects of different agricultural systems and agricultural practices on the abundance, activity, taxonomic and functional diversity of soil fauna are therefore highly relevant (Giller et al. 1997) and are being investigated by a number of soil agroecologists (Blanchart et al. 2006; Ponge et al. 2013; Pelosi et al. 2014a, b; Henneron et al. 2015). However, although a number of review articles have addressed the relationship between agroforestry practices and soil health (Schroth 1998; Hartemink 2005; Jose 2009; Lorenz and Lal 2014; Dollinger and Jose 2018), only one has focused on the effects of agroforestry on soil fauna and the functions they perform (Barrios et al. 2012).
Barrios et al. (2012) reviewed the tropical literature to synthesize the effects of agroforestry on soil fauna and functioning. Using a set of studies that compared agroforestry systems to continuous cropping in the tropics, they found that the mean density of individuals for 7 soil fauna groups belonging to the micro, meso- and macrofauna was multiplied by a value ranging from 1 to 6.1 in agroforestry. However, the small number of studies considered (7 publications) led them to preach caution in generalizing these results to tropical agroforestry in general, known for its diversity of systems including among others shaded perennial crops of cocoa and coffee, tropical homegardens, slash and mulch, shaded pastures, hedgerow alley cropping and high diversity agroforests. Additional systems in temperate regions include alley cropping with wider spacing of tree rows and mediterranean silvopastoral systems. Seven years after the Barrios et al. (2012) review, the question thus remains of how much stabilized knowledge can be extracted from the existing literature about the way soil fauna and their functions are affected by a diversity of agroforestry systems around the world.
We reviewed the published data about soil fauna in agroforestry systems, to assess the current extent of the knowledge gap about the way agroforestry shapes soil faunal communities and their roles in agroecosystem functioning. We chose to define agroforestry systems following Somarriba (1992) as satisfying three conditions, 1) including at least two plant species that interact biologically, 2) at least one of the plant species is a woody perennial, 3) at least one of the plant species is managed for forage, annual or perennial crop production. Thus, we excluded mixed-species forestry plantations. In addition, we chose to narrow our focus on systems that involve simultaneous growth of at least two strata of vegetation, excluding systems known as “sequential agroforestry” (Torquebiau 2000). Our aim was to determine how much research effort had been made in different regions of the world, on different types of thus-defined agroforestry, and whether global patterns of response arise for soil fauna and the different functions of soil fauna.
Identification of relevant literature
We used Web of Science (Clarivate Analytics) using the following keywords to define three corpus, i.e. papers written in English and dealing with invertebrates (request 1: keywords macrofauna; mesofauna; microfauna; earthworm; woodlice; slug; snails; mollusks; insect; spider; centipede; termite; ant; larvae; millipede; mite; collembola; nematode; microarthropod; arthropod; protist; ecosystem engineer; pest; natural enemy; detritivor; predator; bacterial-feeder; bacterivor*; fungivor*; dwelling), papers dealing with agroforestry (request 2: keywords alley; agroforestry; silvoarable; silvopastoral; parkland; graz* AND woodland) and those dealing with soil (request 3: keyword soil).
We then combined these three corpus and the search yielded 388 manuscripts (last search made on August 22, 2019). All the manuscripts were read to assess whether they fell in the scope of this literature review, i.e. whether they contained original datasets of soil macrofauna abundance, diversity and/or functions in agroforestry systems. For this review we chose to restrict what we considered as agroforestry systems to agricultural or pastoral systems structured in at least two strata, and including trees at all times. Among the agroforestry types identified by Torquebiau (2000), we therefore included “crops under tree cover”, “agroforests” (except for “mixed woodlots”), “agroforestry in linear arrangement”, and “animal agroforestry” and excluded “sequential agroforestry”. Mixed-species forestry plantations were also excluded. Seventy-four manuscripts were selected in this way (Supplementary data S1).
Knowledge extraction
We extracted information on (i) the general research question, (ii) the type of agroforestry under consideration, (iii) groups of soil fauna considered, (iv) the method used to detect effects of agroforestry on soil fauna, (v) effects observed on soil fauna abundance or diversity and, when available, the associated soil functions that were influenced by soil fauna.
General research questions were identified in two stages. Two main research question categories were identified prior to reading the publications:
-
1)
Characterization of a soil fauna community in an agroforestry system and
-
2)
Characterization of a function associated to soil fauna in an agroforestry system.
For both questions, 4 sub-questions were also identified:
-
a)
Effect of agroforestry, as compared to a control;
-
b)
Response to design or management of the agroforestry system;
-
c)
Seasonal or spatial heterogeneity within the agroforestry system and
-
d)
Long-term changes during agroforestry system development.
Each article was read and attributed to any number of these categories. This screening also allowed the identification of additional questions that were not expected a priori (Table 1).
Concerning the type of agroforestry under consideration, we adopted a 2-stage operational classification system inspired from Sinclair (1999). First, three major types were identified according to system components: silvoarable (defined by the presence of at least one herbaceous crop and absence of livestock grazing), silvopastoral (defined by the presence of livestock grazing), and perennial crop agroforestry systems (defined by the absence of herbaceous crops). Secondly, systems were categorized as including low, medium or high species richness of crops (2, 3–5, or > 6 species respectively; timber-producing species included), and according to their spatial structure (trees aligned in rows or not). Crop species richness was recorded when possible but in many cases the exact number of species was not given in the manuscript, which is why we used categorization. The hypotheses underlying this second stage of classification were that 1) planned crop diversity could affect soil fauna by affecting habitat and resource diversification and 2) the spatial structure of the agroforestry plot would affect intra plot heterogeneity in habitats and resources, thereby affecting the spatial structure and diversity of the soil fauna community (Ettema and Wardle 2002).
Secondly, we categorized articles as focusing on any combination of microfauna, mesofauna or macrofauna. Articles focused on a specific taxonomic group were attributed to the dominant size-group of the taxon (e.g. an article dedicated to termites was attributed to articles studying macrofauna).
Regarding the method used to detect effects of agroforestry on soil fauna, we noted whether one or several control plots were studied and which land use they represented (crop, pasture, forest), or whether the study examined a gradient of sampling sites within or between studied plots.
A subset of 51 publications provided enough information to assess the effect of agroforestry on soil fauna abundance and/or diversity. The effects were categorized as positive, neutral or negative compared to the corresponding control plot(s). A positive effect signified that the agroforestry system significantly increased faunal abundance or diversity (whether the effect is viewed as beneficial or not, e.g. an increase in the abundance of a crop pest in the agroforestry system would count as a positive effect).
Finally, only functions that were explicitly linked to soil fauna in the study were selected. We categorized soil functions according to Kibblewhite et al. (2008), into “C transformations”, “Nutrient cycling”, “Soil structure maintenance” and “Biological regulation”, and added a fifth function “Crop production”.
Main questions addressed in studies about soil fauna in agroforestry systems
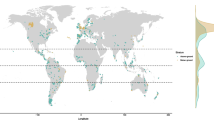
Most studies aimed to identify the effect of land use on abundance and/or diversity characteristics of a soil fauna community, either with a specific focus on the effect of agroforestry systems in particular (32 articles, Table 1), or including one or several agroforestry systems in a comparison of different land use systems of a given region (19 articles), sometimes taking into account ecological gradients due to elevation for example (e.g. Karungi et al. 2018). Some of these studies pinpointed one particular taxon, like dung beetles (e.g. Giraldo et al. 2011; De Farias et al. 2015; Gómez-Cifuentes et al. 2019) or ants (Delabie et al. 2007) with an emphasis on conservation issues, while others had a multitaxa approach on the macrofauna and/or mesofauna community (e.g. Barros et al. 2002; Rahman et al. 2012). Two studies encompassed all three size categories of soil fauna (Yeates et al. 2000; Vanhove et al. 2016). In some cases the question was broader as it aimed to evaluate the effect of land use on soil quality rather than on soil fauna community assembly (7 articles). For instance, Rousseau et al. (2013) used a detailed description of the soil macrofauna community to identify specific taxa that illustrated the effects on soil quality of the Quesungual slash-and-mulch agroforestry system in Nicaragua. Given the diversity of agroforestry systems and of soil fauna organisms, there is still room for further studies addressing these questions, particularly in under-represented regions and systems (e.g. North America and Asia, silvopastoral systems in Africa; Fig. 1).

Worldwide distribution of the agroforestry systems retained in the present literature review. The bar chart represents the number of articles related to soil fauna in the three types of agroforestry systems (perennial crop, silvopastoral, silvoarable systems) per country. The countries represented in dark green on the map are those mentioned in at least one of the 74 papers included in our analysis
Management and design of agroforestry systems and their effects on the soil fauna community were investigated in 17 studies. For example, the effects of different levels of shade on soil functioning in tropical perennial systems (coffee and cocoa agroforestry systems in particular) were studied in 4 articles. The specific effects of different shade tree species on soil fauna assemblages were reported for instance by Barros et al. (2003). A few scattered articles investigated the effect of specific management practices such as fertilization (e.g. Vohland and Schroth 1999) or pesticide application (e.g. Norgrove et al. 2009; Wielgoss et al. 2010) in agroforestry systems, or of shrub control options in silvopastoral systems (e.g. Azul et al. 2011). Two publications looked into the effects of tree addition in existing land use: establishment of tree islands in oil-palm plantations in Indonesia (Teuscher et al. 2016) or addition of shade trees in old-growth cocoa plantations (Vanhove et al. 2016).
The inherent heterogeneity of agroforestry systems was relatively poorly represented as only 8 publications provided data on seasonal and/or spatial distribution of soil fauna within agroforestry plots (most notably Price and Gordon (1998) and García-Tejero and Taboada (2016)). Finally, only three manuscripts addressed medium to long-term temporal changes in soil communities with the growth and development of agroforestry systems, two of which contained data from the same silvopastoral row agroforestry study site in New Zealand (Yeates 1988; Yeates et al. 2000), while the third focused on the alleviation by trees of long-term continuous cropping-related soil degradation in Kenya (Kamau et al. 2017).
Despite the general recognition in manuscript introductions of their importance in agroforestry, soil fauna functions were only investigated in 17 publications, among which 7 focused on the prevalence of agricultural pests (for instance slugs, in Griffiths et al. (1998) or plant parasitic nematodes, in Adekunle (2009)) and sometimes on their predators (e.g. Martin-Chave et al. 2019). Eleven of these papers compared a function in agroforestry plots and control plots, and 6 assessed its spatial and or seasonal distribution. Only 2 papers addressed effects on these functions of agroforestry design or management. No study assessed long term changes in soil fauna functions with agroforestry system development. Lastly, we noted one publication that related to anthropological issues by questioning smallholder farmers’ knowledge about soil macrofauna and its relationships with system components and functioning (Pauli et al. 2012).
Agroforestry system diversity
Regarding the types of agroforestry systems represented in the manuscripts, silvoarable and perennial crop agroforestry systems were the most studied with 44% and 36% of the studies respectively, compared to the relatively low number of studies focusing on soil fauna in silvopastoral systems (20% and 13 publications) (Table 2). A partial explanation could be that some silvopastoral studies are not qualified as such or as agroforestry studies, and were therefore missed by our literature survey, but an additional search with keywords “dehesa” or “montado” for instance did not yield new papers. Another explanation may be that soil biology is not regarded as a main driver of pasture ecosystem performance contrarily to soil physical quality (and mainly soil bulk density in response to trampling). It can be seen as paradoxical since for example macrofauna is known through its bioturbation activities, to be able to alleviate some of the negative effects of soil compaction (Capowiez et al. 2012).
45% of the studies dealing with soil fauna in agroforestry systems were carried out in systems based on a low planned botanical diversity with only 2 species, the crop and the tree. Fewer studies were carried out in medium or high diversity systems (28% and 27% respectively) (Table 2). This could indicate that soil fauna is considered as an interesting lever in intensive systems with low botanical diversity.
The 74 retained manuscripts report data from twenty three countries on almost all continents, reflecting the global importance of agroforestry systems (Fig. 1). Central and South America are particularly well represented in the literature, with Brazil being the only country where soil fauna has been studied in the three main types of agroforestry system. Temperate regions have no equivalent of the perennial crop agroforestry systems of South America and Central Africa, but several studies have been published on silvoarable and silvopastoral systems in western Europe.
A remarkable diversity of systems is represented. Well-represented systems include shaded coffee (10 articles from 5 countries: Ethiopia, Indonesia, Costa Rica, Uganda, Panama) and cocoa systems (13 articles from 6 countries: Costa Rica, Indonesia, Brazil, Mexico, Cameroon, Malaysia). Slash-and-mulch agroforestry systems from Central America, presented as an alternative to the traditional slash-and-burn with a shorter fallow period, are also very well represented with 7 articles. Tropical silvopastoral systems are studied in 8 papers from 4 Central- and South-American countries and 2 African countries, mirrored by 10 temperate (New Zealand, Canada, Australia) and Mediterranean (Spain, Italy, Portugal) articles. Temperate alley cropping systems with wide spacing are represented in 5 publications while Nigerian tropical hedgerow alley cropping systems are studied in 4 papers. More anecdotal systems include tropical homegardens (Rahman et al. 2012; Lakshmi and Joseph 2017), tropical multistrata successional agroforests from the Brazilian Mata Atlantica biome (Cezar et al. 2015), temperate alley cropping with market gardening (Martin-Chave et al. 2018), tropical market gardening with shade trees (Adekunle 2009; Guenat et al. 2019), and various Chinese systems like prickly ash intercropped with sweet pepper (Wang et al. 2018) or jujube intercropped with wheat (Liu et al. 2016).
Other regions are however very poorly represented, possibly due to a scarcity of agroforestry systems for example in North America, eastern Europe, and Australia, or to a lack of published studies on soil fauna in existing systems like Faidherbia albida parklands in the Sahel region of Africa. Note that some regions with significant research efforts are no doubt under-represented in this literature survey, as some studies are published in local sources and not referenced in Web of Science.
The temporal dynamics of the number of studies published showed a contrasted situation after 2005 between studies carried out under temperate or non-temperate parts of the world (Fig. 2). While the dynamics were slow and very similar up to 2005, the number of publications then increased sharply in non-temperate regions with on average 3 publications per year and in contrast remained very slow in temperate regions. This trend is possibly linked with the increasing appraisal for soil biology studies and the ecosystem services they are responsible for in tropical ecology (Marichal et al. 2014). Another explanation could be that sustainability issues related to tropical agricultural expansion have recently sparked more interest among funding agencies.

Publication dynamics on soil fauna in agroforestry systems, in mediterranean, temperate, and tropical or subtropical climates
Methods and designs used to assess the effects of agroforestry on soil fauna
In relation with the diversity of agroforestry systems, we observed a variety of designs applied to detect the effects of these systems on soil fauna. Basically, agroforestry can be seen as a mixture of crops (pasture, arable or perennial) and trees (present before the crops or intentionally planted). Thus control plots could be either (natural) forests or an open pasture or crop monoculture, or both. Most of the time the forest is considered as a positive control with higher biodiversity and low influence of anthropogenic perturbations, whereas the crop or open pasture is viewed as a negative control with reduced biodiversity and generally higher management intensity (involving tillage, pesticides, fertilizers…). In our literature review, we found that 31% of the studies did not include control plots, either because they did not test an effect of agroforestry on the soil fauna community, or because they relied on the existence of a gradient within the plot (for example distance from the trees) to do so. For example, Hauser et al. (1998) mainly focused on casting activity from the hedgerow to the middle of the alley crops. Most studies used a crop monoculture or a forest plot as control plot (respectively 35 and 25 studies, or 67% and 48% of the studies with a control), while 8 studies or 15% used a pasture. Nine studies used both crop monoculture and forest control plots, while one study used all three (Suarez et al. 2018). The choice of the relevant control plot is far from neutral; for example the abundance of some soil fauna groups like earthworms is generally lower in forests than in pasture systems with relatively low management intensity (Edwards and Bohlen 1996; Rossi and Blanchart 2005).
Within-plot heterogeneity is another characteristic of agroforestry systems that can have a large influence on soil fauna, be it intentional (trees in rows) or not (heterogeneous development of trees). In most intensive agroforestry systems with trees in rows, grassy strips are present under the line of trees where mechanization is impossible or difficult. The presence of grassy strips (managed or not) can modify the structure and abundance of soil communities, as proved by studies that focused on the effects of grassy field margins in arable land on soil fauna (Smith et al. 2008) and in particular on earthworms (Nieminen et al. 2011; Crittenden et al. 2015). We found 13 studies for which the presence of grassy strips was mentioned but only 9 in which they were taken into account to assess the effects of agroforestry systems on soil fauna. In these studies, linear transects of traps or samplings points were used between these grassy strips and crop alleys (using several distances). In some rare occasions, a double transect was used to consider as well a possible effect of trees within the grassy strips (thus using several distances between the trees). It is worth noting that all studies that mentioned grassy strips were under the silvoarable system and 75% were located under temperate climates.
A large number of soil fauna taxa are present in the database, with widely differing levels of taxonomic resolution for the presentation of results. For most faunal groups and studies, we could not reduce the number of categories by making an a posteriori attribution to a functional group, as this would require genus or sometimes species level identification. The data are furthermore scattered between a large variety of agroforestry systems throughout the world, which in many cases were imperfectly described, with precise information lacking on the plant species present and on the spatial distribution of trees and crops, as well as on grazing and tree or crop management practices. For these reasons, in addition to the relative paucity of available data, a meta-analysis would have produced inherently limited and flawed results.
Two thirds of the studies aiming to compare agroforestry systems with control plots had at least 3 true replicates of the studied treatments (as opposed to pseudo replicates), while 15 studies had no replicates and two studies had just two, making their conclusions fragile. This statistical weakness in study design of a third of the dataset may be due to the fact that it is often very difficult to have access to sufficiently large agroforestry sites (especially when effects of climate are under consideration) where true replication of study plots is possible, in particular in regions where agroforestry is still little developed.
To deal with this problem of lack of space and thus replication, other approaches can be applied to identify the effect of tree presence on soil fauna. Some authors for example studied the effect of individual trees (allowing multiples replicates) by sampling at different distances from a tree (Hauser et al. 1998; Doblas-Miranda et al. 2014; Rossetti et al. 2015; Teuscher et al. 2016); this is also the classical approach in silvopastoral systems. Others took advantage of the presence of intra-plot variability by characterizing the tree density or shading in different places of the agroforestry systems (Pauli et al. 2010; Felicitas et al. 2018). The rationale behind all these approaches is using gradients instead of replicated plots to avoid multiplying non-representative small plots. Such gradients can be used based on the intra-plot heterogeneity or can also be used and controlled between plots. For example Martin-Chave et al. (2018) used a non-replicated design but these plots represented a gradient of tree pruning (leading to a gradient of canopy openness). Another example is given by the study by Yeates (1988) using plots with different tree densities.
Beyond the comparison between systems (agroforestry versus control plots), some key questions remain unanswered: to which distance will trees have an effect? To what extent is soil biodiversity in agroforestry systems controlled by dispersal limitation and the regional species pool? Environmental heterogeneity is an inherent originality of agroforestry systems, where tree and crop compartments co-exist side by side and potentially induce specific spatial patterns and temporal dynamics. The spatial structure of soil fauna metacommunities and their interactions in agroforestry is therefore of particular interest. This further highlights the experimental designs that used intra-plot variability (Price and Gordon 1998) (intended or accidental) along with studies that focused on the effect of the existence of tree rows (Vohland and Schroth 1999; Cardinael et al. 2019). Most, if not all of these studies used sampling at different distances from individual trees or tree rows. In systems planted in rows, this question has practical importance since the answer may help to determine an optimal distance between rows, along with information about the effects of shading and root competition on crop growth and production, and of tree rows and grassy strips on adventitious plant and above-ground pest and natural enemy dissemination.
Effects of agroforestry systems on soil fauna
The number of manuscripts dealing with macrofauna (62) was much higher than those dealing with meso- or microfauna (13 and 10 respectively), and this trend was observed for the three main types of agroforestry systems (Fig. 3). This trend was not unexpected and somehow reflects the number of scientists specialized on each fauna group. It also indicates that the functions associated to macrofauna may have been first highlighted. Microfauna was only represented by nematodes (including free living and parasitic ones). Mesofauna were mainly collembola (69%), acari (54%) and enchytraeids (31% of the cases). Studies on macrofauna that explicitly mentioned certain taxonomic groups mainly focused on earthworms (20 articles), beetles (13) and then ants (6), termites (5) and spiders (4). It is worth noting that in 45% of the studies only one group of macrofauna was studied, and most of the time it was earthworms (16 articles).

Number of articles studying the effects of agroforestry on soil macro- (left), meso- (middle) and microfauna (right) depending on the type of agroforestry system and its botanical diversity (species richness)
Studied metrics were abundance, and in many cases taxonomic diversity indices of specific groups of organisms: species richness, Shannon diversity and Pielou index being the most common. Abundance data were sometimes presented for size categories (e.g. Negasa et al. 2017 for macrofauna) but most studies identified taxonomic groups, with different levels of precision. A common level of precision for studies on macrofauna and mesofauna in general was approximately class (Oligochaeta, Arachnida, Chilopoda...) to order level (for insects: Coleoptera, Heteroptera...) (15 studies). Most articles that focused on a specific taxonomic group identified to the species level (31 studies), although species-level data were generally but not always reported in the publication. A few articles grouped fauna according to ecological categories (e.g. Bhadauria et al. 2012 for earthworms), trophic or feeding groups (e.g. Sauvadet et al. 2019 for nematodes, Muvengwi et al. 2017 for termites) or eco-morphological groups (e.g. Lakshmi and Joseph 2017 for microarthropods).
Regarding the effects on soil fauna abundance or diversity, these effects were mostly positive (70% of the datasets) if the control plots were crops (Fig. 4), with response ratios mostly between 1.2 and 2 but reaching values of up to 10. Only 2 studies reported negative results compared to a crop control. These results are in line with those compiled by Barrios et al. (2012), who found response ratios varying from 1 to 6 for 6 groups of macrofauna, 2 groups of mesofauna and 2 groups of microfauna. Effects were however mostly neutral (40% of the datasets) or negative (28%) if the control plots were forests. There were too few papers dealing with pastures as control plots to draw a trend. Articles focusing on the most represented group, i.e. earthworms, reported similar results: positive effects of agroforestry compared to a crop control in 9 of 13 datasets, but also compared to a forest control in 4 of 8 datasets since earthworm abundance is generally low in forest soils.

Number of articles showing positive, neutral or negative effects of agroforestry on soil macro-, meso- and micro-fauna according to three types of reference (forest, crop or pasture)
Interestingly, the only three studies reporting negative effects compared to crop or pasture control plots were all carried out in temperate climates, while all tropical and mediterranean studies with a crop or pasture control reported positive or neutral effects of agroforestry (Fig. 5). All but one of the comparisons to a forest control plot were made in tropical or subtropical climates.

Number of articles showing positive, neutral or negative effects of agroforestry on soil fauna in tropical/subtropical (left), temperate (middle) and mediterranean systems (right), according to three types of reference (forest, crop or pasture)
We expected to find a number of articles using studies on soil fauna in agroforestry to contribute to the general debate about interactions between above-ground and below-ground diversity (Eisenhauer et al. 2011); and indeed, some reported datasets could have been interpreted in this light. However, we only found one paper that explicitly addressed this question, and in the specific context of pest regulation (Poeydebat et al. 2017). A similar number of datasets, respectively 27 and 29, concerned “high” (>5 plant species) versus “low” (2 species) diversity agroforestry systems (Fig. 3). Positive effects of agroforestry were more frequent in low diversity systems (72% of cases) than in high diversity systems (41%), but this comparison is flawed as the low diversity systems were mostly compared to a crop control while the high diversity systems were more often compared to a forest control. In fact a fairly similar proportion of low, medium and high diversity agroforestry plots showed a positive effect when they were compared to a crop control (15 in 19, 3 in 7 and 4 in 6 studies respectively).
The few studies reporting data on spatial heterogeneity in agroforestry soil fauna communities did not show systematic effects. In temperate alley-cropping systems in Canada, earthworm abundances were either higher or lower in the tree rows than in the crop alleys, depending on the tree species and on the season (Price and Gordon 1998). In similar systems in the United Kingdom, more arthropods were found in the tree rows than in the crop alleys (Park et al. 1994). Likewise in 13 French silvoarable systems, all earthworm ecological categories were more abundant in the tree rows than in the crop alley, however the authors interestingly observed that body mass was lower in the tree rows (Cardinael et al. 2019). In a Canadian site where trees were intercropped with forage crops, distance to trees did not affect oribatid mite abundance or species diversity (Doblas-Miranda et al. 2014). However sampling at different depths revealed an interesting tendency of trees to reduce depth-related differences in mite communities compared to the conventional crop.
Owing to the diversity of agroforestry types and the studied climatic regions, one of the main difficulties is to assess the genericity of the findings. It is thus important to highlight that some tools such as trait-based approaches are now more and more available, even for soil organisms (Pey et al. 2014b), that could overcome some of these limits. The use of traits has several possible advantages. First of all, it can allow diversity and functional analyses when taxonomic knowledge is fragile, which is often the case for soil organisms in many parts of the world (Marichal et al. 2014). Next, traits can help to understand how multi-trophic communities are influenced by land-use changes or soil disturbance (e.g. Hedde et al. 2012). Traits can also be used at a more integrated scale either to estimate networks of interaction between organisms (Gravel et al. 2016) or to test consistency of responses between different datasets (Pelosi et al. 2016).
Regarding their possible use in agroforestry systems for soil organisms, different traits could be used or have been used such as body size (Cardinael et al. 2019), or preference for temperature or humidity and diurnal vs. nocturnal activity (Martin-Chave et al. 2018). These approaches rely heavily on existing and fully available databases of traits, the use of standardized methods (Moretti et al. 2017) and the existence of a stabilised thesaurus (Pey et al. 2014a). A prerequisite for a re-analysis of published data with a trait-based approach is that the data be accessible at the species level. This was the case for roughly half of the articles in our dataset, but only for a restricted number of taxa.
Finally, no study was found that analysed the effect of agroforestry on the soil community in terms of interaction networks, despite their potential usefulness in soil ecology (Morales-Castilla et al. 2015). This is however an interesting question, i.e. whether the juxtaposition of tree-related and crop-related soil communities leads to effective interactions between the two, depending on spatial design and possibly climate.
Effects of agroforestry systems on soil functions associated to soil fauna
It is important to note that we specifically addressed cases where the studied soil functions were associated with soil fauna (causal links only). This is the main explanation regarding the low percentage of studies where soil functions were assessed (Table 3). Among the main functions addressed we found in 36% of the cases “Soil structure and maintenance”, and mainly in silvoarable systems. This further emphasizes the higher percentages of studies focusing on macrofauna and especially earthworms (and termites). This means that the bioturbation activities of these ecosystem engineers (Jouquet et al. 2006) were often investigated with a strong focus on earthworm casting activities either in response to the presence of trees (Hauser et al. 1998; Pauli et al. 2010) but also as a specific soil feature that could enhance soil fertility (Kang and Ojo 1996). Other kinds of bioturbation activities such as sheeting or nest building by termites (Cheik et al. 2019) or burrow creation by earthworms (Bastardie et al. 2005) have not really been investigated so far.
Comparatively, “C transformations” or “Nutrient cycling” were only rarely assessed, in a total of 6 studies, which illustrates the difficulty to investigate the mainly indirect relationships between macro- or mesofauna and soil chemical fertility. Hauser (1993) studied earthworm casting activity and cast N, K, Ca, and Mg concentrations in an alley cropping system to determine their role in returning nutrients to the soil, and concluded that the earthworm-mediated flux was 3 times higher than in control plots without trees. In contrast, in an experimental study with field-incubated microcosms with or without P. corethrurus earthworms, Fonte et al. (2010) observed a very minor effect of the slash-and-mulch agroforestry system compared to slash-and-burn, but a deleterious effect of the worms on C and N dynamics. These results possibly reflect the inherent difficulty in setting up relevant field experiments for evaluation of fauna functions.
Few papers (8) explicitly studied “Crop production” or “Biocontrol” which was not unexpected regarding the fact that only a minor part of the pests have living stages below or on the ground in comparison with pests present in the vegetation. Interestingly, the biocontrol function was always associated with macrofauna (Araneae and Carabidae), thus generalist predators, whereas production (here damages) was associated with pests (slugs and nematodes). Griffiths et al. (1998) found higher abundance and diversity of slugs in temperate alley cropping than in monocultures, particularly in grass strips but also in crop alleys. They also found more damage to crops near tree rows compared to other areas studied (mid row and control plots). Kang et al. (1999) reported that alley cropping in Nigeria had no effect on plant parasitic nematodes, but increased non-parasitic nematode abundance. In a kale crop in Kenya, shade trees increased the abundance of predatory beetles and spiders, but had no direct effect on aphid predation rate, caterpillar predation rate by birds or invertebrates, or parasitism rate (Guenat et al. 2019). Finally, Martin-Chave et al. (2018) observed that the difference in predation potential between agroforestry and control plots depended on the time of year. One should note that only the two latter authors actually evaluated a predation function by using the sentinel prey technique, while the other studies used pest or natural enemy abundance and diversity as a proxy for the biocontrol function.
The relatively small number of studies actually estimating soil functions associated to soil fauna is somehow paradoxical since most of the authors justified their relevance by linking the studied fauna to their assumed functional roles in soil. This may illustrate the possible negative effects of the concept of “soil ecosystem engineer” (Lavelle et al. 2006), where assumed postulates such as “earthworms increase soil water infiltration” (Spurgeon et al. 2013) prevent the authors from actually measuring soil functions. We believe it is premature to use these postulated links, in particular for very complex systems such as agroforestry systems with strong temporal and spatial heterogeneity. We call for more studies not only characterizing soil fauna communities in different, well-described agroforestry situations, but also actually measuring the functions they supposedly perform. Such studies are vital to the development of effect trait-based approaches aiming to predict the effect of community changes on agroecosystem functioning.
Most of the studies reviewed here focused on a single soil function; just two articles reported effects on three functions simultaneously (Fonte and Six 2010; Kang and Ojo 1996). Very few links were explicitly made between biodiversity metrics and soil function intensity, although such links could be very interesting to analyse in the case of agroforestry. Agroforestry systems offer different situations of spatial associations of trees and crop/pasture and the distinct faunal communities they support, which can be expected to induce specific spatial structures of soil biodiversity and also of interaction networks. Relevant soil ecology questions in agroforestry therefore include: how does the spatial structure of soil biodiversity and its interactions influence ecosystem function? and how can we integrate within and between spatial scales to assess functions?
Conclusions
In agroforestry systems, soil ecologists have questioned whether soil fauna communities are affected by agroforestry and, a little, how soil fauna communities affect ecosystem functioning. Agroforestry systems have in many cases been analysed taxon by taxon, with a deterministic point of view, at plot scale. This agrees with the fact that soil fauna is mostly considered by empiricists and generally underrepresented in theoretical work and models (Briones 2018), both in community ecology and ecosystem ecology.
The effect of agroforestry on soil fauna abundance and diversity has generally been found to be positive, when it has been compared to a cropping system without trees. However the diversity of agroforestry types, study focus, pedoclimatic regions and ways of representing soil fauna communities still makes this conclusion vague and fragile. Trait-based approaches could be a useful tool to increase the genericity of further studies.
In generic terms, the specificity of an agroforestry system consists in associating trees with a perennial or annual crop, or a pasture: i.e. vegetation types that typically harbour different soil faunal communities. How do these different communities evolve when they are juxtaposed as a metacommunity? What is their level of interaction? How do these interactions depend on spatial design, climate, plant species identity, grazing pressure? What are the functional consequences of these interactions for the agroecosystem? These questions require more work on environmental heterogeneity, interaction networks and fauna functions; they are crucial for soil ecologists in general (Sutherland et al. 2013), and agroforestry systems offer a diversity of relevant situations to address them.
References
Adekunle OK (2009) Population dynamics of Meloidogyne incognita and three other phytonematodes on okra cultivars planted in alleys of Leucaena leucocephala and Gliricidia sepium. Australas Plant Pathol 38:211–215
Azul AM, Mendes SM, Sousa JP, Freitas H (2011) Fungal fruitbodies and soil macrofauna as indicators of land use practices on soil biodiversity in Montado. Agrofor Syst 82:121–138
Barrios E, Sileshi GW, Shepherd K, Sinclair F (2012) Agroforestry and soil health: linking trees, soil biota, and ecosystem services. In: Soil ecology and ecosystem services, Oxford University Press. Wall, D. H. et al, pp 315–330
Barrios E, Valencia V, Jonsson M, Brauman A, Hairiah K, Mortimer PE, Okubo S (2018) Contribution of trees to the conservation of biodiversity and ecosystem services in agricultural landscapes. Int J Biodivers Sci Ecosyst Serv and Manage 14:1–16
Barros E, Pashanasi B, Constantino R, Lavelle P (2002) Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biol Fertil Soils 35:338–347
Barros E, Neves A, Blanchart E, Fernandes ECM, Wandelli E, Lavelle P (2003) Development of the soil macrofauna community under silvopastoral and agrosilvicultural systems in Amazonia. Pedobiologia 47:273–280
Bastardie F, Capowiez Y, Cluzeau D (2005) 3D characterisation of earthworm burrow systems in natural soil cores collected from a 12-year-old pasture. Appl Soil Ecol 30:34–46
Bhadauria T, Kumar P, Kumar R, Maikhuri RK, Rao KS, Saxena KG (2012) Earthworm populations in a traditional village landscape in central Himalaya, India. Appl Soil Ecol 53:83–93
Blanchart E, Villenave C, Viallatoux A, Barthès B, Girardin C, Azontonde A, Feller C (2006) Long-term effect of a legume cover crop (Mucuna pruriens var. utilis) on the communities of soil macrofauna and nematofauna, under maize cultivation, in southern Benin. Eur J Soil Biol 42:S136–S144
Briones MJI (2018) The serendipitous value of soil Fauna in ecosystem functioning: the unexplained explained. Front Environ Sci 6:149
Capowiez Y, Samartino S, Cadoux S, Bouchant P, Richard G, Boizard H (2012) Role of earthworms in regenerating soil structure after compaction in reduced tillage systems. Soil Biol Biochem 55:93–103
Cardinael R, Chevallier T, Cambou A, Béral C, Barthès B, Dupraz C, Durand C, Kouakoua E, Chenu C (2017) Increased soil organic carbon stocks under agroforestry: a survey of six different sites in France. Agric Ecosyst Environ 236:243–255
Cardinael R, Hoeffner K, Chenu C, Chevallier T, Béral C, Dewisme A, Cluzeau D (2019) Spatial variation of earthworm communities and soil organic carbon in temperate agroforestry. Biol Fertil Soils 55:171–183
Cezar RM, Vezzani FM, Schwiderke DK, Gaiad S, Brown GG, Seoane CES, Froufe LCM (2015) Soil biological properties in multistrata successional agroforestry systems and in natural regeneration. Agrofor Syst 89:1035–1047
Cheik S, Bottinelli N, Soudan B, Harit A, Chaudhary E, Sukumar R, Jouquet P (2019) Effects of termite foraging activity on topsoil physical properties and water infiltration in vertisol. Appl Soil Ecol 133:132–137
Chen J, Saunders SC, Crow TR, Naiman RJ, Brosofske KD, Mroz GD, Brookshire BL, Franklin JF (1999) Microclimate in Forest ecosystem and landscape ecology variations in local climate can be used to monitor and compare the effects of different management regimes. BioScience 49:288–297
Crittenden SJ, Huerta E, de Goede RGM, Pulleman MM (2015) Earthworm assemblages as affected by field margin strips and tillage intensity: an on-farm approach. Eur J Soil Biol 66:49–56
De Farias PM, Arellano L, Hernández MIM, Ortiz SL (2015) Response of the copro-necrophagous beetle (Coleoptera: Scarabaeinae) assemblage to a range of soil characteristics and livestock management in a tropical landscape. J Insect Conserv 19:947–960
Delabie JHC, Jahyny B, do Nascimento IC, Mariano CSF, Lacau S, Campiolo S, Philpott SM, Leponce M (2007) Contribution of cocoa plantations to the conservation of native ants (Insecta: Hymenoptera: Formicidae) with a special emphasis on the Atlantic Forest fauna of southern Bahia, Brazil. Biodivers Conserv 16:2359–2384
Doblas-Miranda E, Paquette A, Work TT (2014) Intercropping trees’ effect on soil oribatid diversity in agro-ecosystems. Agrofor Syst 88:671–678
Dollinger J, Jose S (2018) Agroforestry for soil health. Agrofor Syst 92:213–219
Edwards CA, Bohlen PJ (1996) Biology and ecology of earthworms. Springer Science & Business Media
Eisenhauer N, Migunova VD, Ackermann M, Ruess L, Scheu S (2011) Changes in plant species richness induce functional shifts in soil nematode communities in experimental grassland. PLoS One 6(9):e24087
Ettema CH, Wardle DA (2002) Spatial soil ecology. Trends Ecol Evol 17:177–183
Felicitas AC, Hervé BDB, Ekesi S, Akutse KS, Djuideu CTCL, Meupia MJ, Babalola OO (2018) Consequences of shade management on the taxonomic patterns and functional diversity of termites (Blattodea: Termitidae) in cocoa agroforestry systems. Ecol Evol 8:11582–11595
Fonte SJ, Six J (2010) Earthworms and litter management contributions to ecosystem services in a tropical agroforestry system. Ecol Appl 20:1061–1073
Fonte SJ, Barrios E, Six J (2010) Earthworm impacts on soil organic matter and fertilizer dynamics in tropical hillside agroecosystems of Honduras. Pedobiologia 53:327–335
García-Tejero S, Taboada Á (2016) Microhabitat heterogeneity promotes soil fertility and ground-dwelling arthropod diversity in Mediterranean wood-pastures. Agric Ecosyst Environ 233:192–201
Giller KE, Beare MH, Lavelle P, Izac AMN, Swift MJ (1997) Agricultural intensification, soil biodiversity and agroecosystem function. Appl Soil Ecol 6:3–16
Giraldo C, Escobar F, Chará JD, Calle Z (2011) The adoption of silvopastoral systems promotes the recovery of ecological processes regulated by dung beetles in the Colombian Andes. Insect Conserv Diver 4:115–122
Gómez-Cifuentes A, Giménez Gómez VC, Moreno CE, Zurita GA (2019) Tree retention in cattle ranching systems partially preserves dung beetle diversity and functional groups in the semideciduous Atlantic forest: the role of microclimate and soil conditions. Basic Appl Ecol 34:64–74
Gravel D, Albouy C, Thuiller W (2016) The meaning of functional trait composition of food webs for ecosystem functioning. Philos Trans R Soc B 371:20150268
Griffiths J, Phillips DS, Compton SG, Wright C, Incoll LD (1998) Responses of slug numbers and slug damage to crops in a silvoarable agroforestry landscape. J Appl Ecol 35:252–260
Guenat S, Kaartinen R, Jonsson M (2019) Shade trees decrease pest abundances on brassica crops in Kenya. Agrofor Syst 93:641–652
Guillot E, Hinsinger P, Dufour L, Roy J, Bertrand I (2019) With or without trees: resistance and resilience of soil microbial communities to drought and heat stress in a Mediterranean agroforestry system. Soil Biol Biochem 129:122–135
Hartemink AE (2005) Nutrient stocks, nutrient cycling, and soil changes in cocoa ecosystems: a review. In: Sparks DL (ed) Advances in agronomy, vol 86. Elsevier Academic Press Inc, San Diego, pp 227–253
Hauser S (1993) Distribution and activity of earthworms and contribution to nutrient recycling in alley cropping. Biol Fertil Soils 15:16–20
Hauser S, Asawalam DO, Vanlauwe B (1998) Spatial and temporal gradients of earthworm casting activity in alley cropping systems. Agrofor Syst 41:127–137
Hawke MF, Wedderburn ME (1994) Microclimate changes under pinus radiata agroforestry regimes in New Zealand. Agric For Meteorol 71:133–145
Hedde M, van Oort F, Lamy I (2012) Functional traits of soil invertebrates as indicators for exposure to soil disturbance. Environ Pollut 164:59–65
Henneron L, Bernard L, Hedde M, Pelosi C, Villenave C, Chenu C, Bertrand M, Girardin C, Blanchart E (2015) Fourteen years of evidence for positive effects of conservation agriculture and organic farming on soil life. Agron Sustain Dev 35:169–181
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76:1–10
Jouquet P, Dauber J, Lagerlöf J, Lavelle P, Lepage M (2006) Soil invertebrates as ecosystem engineers: intended and accidental effects on soil and feedback loops. Appl Soil Ecol 32:153–164
Kamau S, Barrios E, Karanja NK, Ayuke FO, Lehmann J (2017) Soil macrofauna abundance under dominant tree species increases along a soil degradation gradient. Soil Biol Biochem 112:35–46
Kang BT, Ojo A (1996) Nutrient availability of earthworm casts collected from under selected woody agroforestry species. Plant Soil 178:113–119
Kang BT, Caveness FE, Tian G, Kolawole GO (1999) Longterm alley cropping with four hedgerow species on an Alfisol in southwestern Nigeria – effect on crop performance, soil chemical properties and nematode population. Nutr Cycl Agroecosyst 54:145–155
Karungi J, Cherukut S, Ijala AR, Tumuhairwe JB, Bonabana-Wabbi J, Nuppenau EA, Hoeher M, Domptail S, Otte A (2018) Elevation and cropping system as drivers of microclimate and abundance of soil macrofauna in coffee farmlands in mountainous ecologies. Appl Soil Ecol 132:126–134
Kibblewhite MG, Ritz K, Swift MJ (2008) Soil health in agricultural systems. Philos Trans R Soc B-Biol Sci 363:685–701
Lakshmi G, Joseph A (2017) Soil microarthropods as indicators of soil quality of tropical home gardens in a village in Kerala, India. Agrofor Syst 91:439–450
Lavelle P, Decaëns T, Aubert M et al (2006) Soil invertebrates and ecosystem services. Eur J Soil Biol 42:S3–S15
Liu Y, Li X, Liu Q (2016) Soil nematode communities in jujube (Ziziphus jujuba mill.) rhizosphere soil under monoculture and jujube/wheat (Triticum aestivum Linn.) intercropping systems, a case study in Xinjiang arid region, northwest of China. Eur J Soil Biol 74:52–59
Lorenz K, Lal R (2014) Soil organic carbon sequestration in agroforestry systems. A review. Agronomy for Sustainable Development 34 (2):443-454
Malézieux E, Crozat Y, Dupraz C, Laurans M, Makowski D, Ozier-Lafontaine H, Rapidel B, de Tourdonnet S, Valantin-Morison M (2009) Mixing plant species in cropping systems: concepts, tools and models: a review. In: Lichtfouse E, Navarrete M, Debaeke P et al (eds) Sustainable Agriculture. Springer Netherlands, Dordrecht, pp 329–353
Marichal R, Grimaldi M, Feijoo MA, Oszwald J, Praxedes C, Hernan D, Cobo R, del Pilar HM, Desjardins T, da Silva Junior ML, da Silva Costa LG, Miranda IS, Oliveira MND, Brown GG, Tsélouiko S, Martins MB, Decaëns T, Velasquez E, Lavelle P (2014) Soil macroinvertebrate communities and ecosystem services in deforested landscapes of Amazonia. Appl Soil Ecol 83:177–185
Martin-Chave A, Béral C, Mazzia C, Capowiez Y (2018) Agroforestry impacts the seasonal and diurnal activity of dominant predatory arthropods in organic vegetable crops. Agrofor Syst. https://doi.org/10.1007/s10457-018-0309-4
Martin-Chave A, Béral C, Capowiez Y (2019) Agroforestry has an impact on nocturnal predation by ground beetles and Opiliones in a temperate organic alley cropping system. Biol Control 129:128–135
Monteith JL, Ong CK, Corlett JE (1991) Microclimatic interactions in agroforestry systems. For Ecol Manag 45:31–44
Morales-Castilla I, Matias MG, Gravel D, Araújo MB (2015) Inferring biotic interactions from proxies. Trends Ecol Evol 30:347–356
Moretti M, Dias ATC, de Bello F, Altermatt F, Chown S, Azcarate FM, Bell JR, Fournier B, Hedde M, Hortal J, Ibanez S, Ockinger E, Sousa JP, Ellers J, Berg MP (2017) Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Funct Ecol 31:558–567
Muvengwi J, Mbiba M, Ndagurwa HGT, Nyamadzawo G, Nhokovedzo P (2017) Termite diversity along a land use intensification gradient in a semi-arid savanna. J Insect Conserv 21:801–812
Negasa T, Ketema H, Legesse A, Sisay M, Temesgen H (2017) Variation in soil properties under different land use types managed by smallholder farmers along the toposequence in southern Ethiopia. Geoderma 290:40–50
Nieminen M, Ketoja E, Mikola J, Terhivuo J, Sirén T, Nuutinen V (2011) Local land use effects and regional environmental limits on earthworm communities in Finnish arable landscapes. Ecol Appl 21:3162–3177
Norgrove L, Csuzdi C, Forzi F, Canet M, Gounes J (2009) Shifts in soil faunal community structure in shaded cacao agroforests and consequences for ecosystem function in Central Africa. Trop Ecol 50:71–78
Palm CA (1995) Contribution of agroforestry trees to nutrient requirements of intercropped plants. In: Sinclair FL (ed) Agroforestry: science, policy and practice: selected papers from the agroforestry sessions of the IUFRO 20th world congress, Tampere, Finland, 6–12 August 1995. Springer Netherlands, Dordrecht, pp 105–124
Park J, Newman SM, Cousins SH (1994) The effects of poplar (P. trichocarpa × deltoides) on soil biological properties in a silvoarable system. Agrofor Syst 25:111–118
Pauli N, Oberthür T, Barrios E, Conacher AJ (2010) Fine-scale spatial and temporal variation in earthworm surface casting activity in agroforestry fields, western Honduras. Pedobiologia 53:127–139
Pauli N, Barrios E, Conacher AJ, Oberthür T (2012) Farmer knowledge of the relationships among soil macrofauna, soil quality and tree species in a smallholder agroforestry system of western Honduras. Geoderma 189–190:186–198
Pelosi C, Barot S, Capowiez Y, Hedde M, Vandenbulcke F (2014a) Pesticides and earthworms. A review. Agron Sustain Dev 34:199–228
Pelosi C, Pey B, Hedde M, Caro G, Capowiez Y, Guernion M, Peigné J, Piron D, Bertrand M, Cluzeau D (2014b) Reducing tillage in cultivated fields increases earthworm functional diversity. Appl Soil Ecol 83:79–87
Pelosi C, Pey B, Caro G, Cluzeau D, Peigné J, Bertrand M, Hedde M (2016) Dynamics of earthworm taxonomic and functional diversity in ploughed and no-tilled cropping systems. Soil Tillage Res 156:25–32
Pey B, Laporte M-A, Nahmani J, Auclerc A, Capowiez Y, Caro G, Cluzeau D, Cortet J, Decaëns T, Dubs F, Joimel S, Guernion M, Briard C, Grumiaux F, Laporte B, Pasquet A, Pelosi C, Pernin C, Ponge JF, Salmon S, Santorufo L, Hedde M (2014a) A thesaurus for soil invertebrate trait-based approaches. PLoS One 9:e108985
Pey B, Nahmani J, Auclerc A, Capowiez Y, Cluzeau D, Cortet J, Decaëns T, Deharveng L, Dubs F, Joimel S, Briard C, Grumiaux F, Laporte MA, Pasquet A, Pelosi C, Pernin C, Ponge JF, Salmon S, Santurofo L, Hedde M (2014b) Current use of and future needs for soil invertebrate functional traits in community ecology. Basic Appl Ecol 15:194–206
Poeydebat C, Tixier P, Chabrier C, de Lapeyre de Bellaire L, Vargas R, Daribo MO, Carval D (2017) Does plant richness alter multitrophic soil food web and promote plant-parasitic nematode regulation in banana agroecosystems? Appl Soil Ecol 117–118:137–146
Ponge J-F, Peres G, Guernion M, Ruiz-Camacho N, Cortet J, Pernin C, Villenave C, Chaussod R, Martin-Laurent F, Bispo A, Cluzeau D (2013) The impact of agricultural practices on soil biota: a regional study. Soil Biol Biochem 67:271–284
Price GW, Gordon AM (1998) Spatial and temporal distribution of earthworms in a temperate intercropping system in southern Ontario, Canada. Agrofor Syst 44:141–149
Rahman PM, Varma RV, Sileshi GW (2012) Abundance and diversity of soil invertebrates in annual crops, agroforestry and forest ecosystems in the Nilgiri biosphere reserve of Western Ghats, India. Agrofor Syst 85:165–177
Rossetti I, Bagella S, Cappai C, Caria MC, Lai R, Roggero PP, Martins da Silva P, Sousa JP, Querner P, Seddaiu G (2015) Isolated cork oak trees affect soil properties and biodiversity in a Mediterranean wooded grassland. Agric Ecosyst Environ 202:203–216
Rossi J-P, Blanchart E (2005) Seasonal and land-use induced variations of soil macrofauna composition in the Western Ghats, southern India. Soil Biol Biochem 37:1093–1104
Rousseau L, Fonte SJ, Téllez O, van der Hoek R, Lavelle P (2013) Soil macrofauna as indicators of soil quality and land use impacts in smallholder agroecosystems of western Nicaragua. Ecol Indic 27:71–82
Sauvadet M, den Meersche KV, Allinne C, Gay F, Filho EMV, Chauvat M, Becquer T, Tixier P, Harmand JM (2019) Shade trees have higher impact on soil nutrient availability and food web in organic than conventional coffee agroforestry. Sci Total Environ 649:1065–1074
Schroth G (1998) A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor Syst 43:5–34
Sinclair FL (1999) A general classification of agroforestry practice. Agrofor Syst 46:161–180
Singh AK, Kumar P, Singh R, Rathore N (2012) Dynamics of tree-crop interface in relation to their influence on microclimatic changes - a review. HortFlora Res Spectr 1:193–198
Smith J, Potts SG, Woodcock BA, Eggleton P (2008) Can arable field margins be managed to enhance their biodiversity, conservation and functional value for soil macrofauna? J Appl Ecol 45:269–278
Somarriba E (1992) Revisiting the past: an essay on agroforestry definition. Agrofor Syst 19:233–240
Spurgeon DJ, Keith AM, Schmidt O, Lammertsma DR, Faber JH (2013) Land-use and land-management change: relationships with earthworm and fungi communities and soil structural properties. BMC Ecol 13:46
Suarez LR, Paladines Josa YT, Astudillo Samboni EJ, Lopez Cifuentes KD, Durán Bautista EH, Salazar JCS (2018) Soil macrofauna under different land uses in the Colombian Amazon. Pesqui Agropecu Bras 53:1383–1391
Sutherland WJ, Freckleton RP, Godfray HCJ, Beissinger SR, Benton T, Cameron DD, Carmel Y, Coomes DA, Coulson T, Emmerson MC, Hails RS, Hays GC, Hodgson DJ, Hutchings MJ, Johnson D, Jones JPG, Keeling M, Kokko H, Kunin WE, Lambin X, Lewis OT, Malhi Y, Mieszkowska N, Milner-Gulland EJ, Norris K, Phillimore AB, Purves DW, Reid JM, Reuman DC, Thompson K, Travis JMJ, Turnbull LA, Wardle D, Wiegand T (2013) Identification of 100 fundamental ecological questions. J Ecol 101:58–67
Teuscher M, Gérard A, Brose U, Buchori D, Clough Y, Ehbrecht M, Hölscher D, Irawan B, Sundawati L, Wollni L, Kreft H (2016) Experimental biodiversity enrichment in oil-Palm-dominated landscapes in Indonesia. Front Plant Sci 7:1538
Torquebiau EF (2000) A renewed perspective on agroforestry concepts and classification. CR Acad Sci III - Vie 323:1009–1017
Udawatta RP, Kremer RJ, Adamson BW, Anderson SH (2008) Variations in soil aggregate stability and enzyme activities in a temperate agroforestry practice. Appl Soil Ecol 39:153–160
Vandermeer J, van Noordwijk M, Anderson J, Ong C, Perfecto I (1998) Global change and multi-species agroecosystems: concepts and issues. Agric Ecosyst Environ 67:1–22
Vanhove W, Vanhoudt N, Van Damme P (2016) Effect of shade tree planting and soil management on rehabilitation success of a 22-year-old degraded cocoa (Theobroma cacao L.) plantation. Agric Ecosyst Environ 219:14–25
Vigiak O, Sterk G, Warren A, Hagen LJ (2003) Spatial modeling of wind speed around windbreaks. Catena 52:273–288
Vohland K, Schroth G (1999) Distribution patterns of the litter macrofauna in agroforestry and monoculture plantations in Central Amazonia as affected by plant species and management. Appl Soil Ecol 13:57–68
Wang S, Pan K, Tariq A, Zhang L, Sun X, Li Z, Sun F, Xiong Q, Song D, Olatunji OA (2018) Combined effects of cropping types and simulated extreme precipitation on the community composition and diversity of soil macrofauna in the eastern Qinghai-Tibet plateau. J Soils Sediments 18:3215–3227
Wielgoss A, Tscharntke T, Buchori D, Fiala B, Clough Y (2010) Temperature and a dominant dolichoderine ant species affect ant diversity in Indonesian cacao plantations. Agric Ecosyst Environ 135:253–259
Yeates GW (1988) Earthworm and enchytraeid populations in a 13-year-old agroforestry system. NZ J Forestry Sci 18:304–310
Yeates GW, Hawke MF, Rijkse WC (2000) Changes in soil fauna and soil conditions under Pinus radiata agroforestry regimes during a 25-year tree rotation. Biol Fertil Soils 31:391–406
Young A (1997) Agroforestry for soil management. CAB International, New York
Zomer RJ, Trabucco A, Coe R, Place F, van Noordwijk M, Xu JC. 2014. Trees on farms: an update and reanalysis of agroforestry’s global extent and socio-ecological characteristics. Working paper 179. Bogor, Indonesia: world agroforestry Centre (ICRAF) Southeast Asia regional program
Acknowledgments
We very much thank Eric Blanchart and three anonymous reviewers for their helpful comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Claire Chenu.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 119 kb)
Rights and permissions
About this article

Cite this article
Marsden, C., Martin-Chave, A., Cortet, J. et al. How agroforestry systems influence soil fauna and their functions - a review. Plant Soil 453, 29–44 (2020). https://doi.org/10.1007/s11104-019-04322-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-019-04322-4