
Abstract
Background and aims
Forest composition in North America has undergone important changes since the European settlement. The effects of such alterations on soil properties remain largely unknown. This study aims to understand the long-term effects of shifts in forest composition on soil properties.
Methods
Using data from 130 plots measured over an eighty-year period, the relationships between stand composition (both current and past), parent material and current soil chemical properties were studied with redundancy analyses.
Results
Results indicated that the parent material remained the dominant factor explaining soil properties, followed by current tree species composition. No legacy effect of past forest composition was found, but shifts in forest composition explained part of the current soil properties. Specifically, an increase in balsam fir was related to higher C/N ratio in the O-horizon, while an increase in maple species was related to higher net nitrification in both the O and B-horizons, and higher extractable P in the B-horizon.
Conclusion
Our results suggest that increasing the maple component at the expense of conifers over several decades may enhance nutrient availability in the O-horizon.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The forests of northeastern North America have undergone significant modifications since the first European settlements (Whitney 1994; Danneyrolles et al. 2016). Human activities, such as increases in logging and land clearing or modifications in the fire regime (Fuller et al. 1998; Thompson et al. 2013), are the main causes for the changes in stand structure and composition (Boucher et al. 2006; Dupuis et al. 2011; Held and Jones-Held 2014; Nowacki and Abrams 2015). More specifically, anthropogenic disturbances in Eastern Canada have led to an increase in the proportion of fast-growing tree species such as sugar maple (Acer saccharum Marsh.), red maple (Acer rubrum L.) and quaking aspen (Populus tremuloides Michx.) to the detriment of shade-tolerant conifers such as spruce spp. (mainly white spruce: Picea glauca (Moench) Voss) and northern white-cedar (Thuja occidentalis L.) (Boucher et al. 2006; Dupuis et al. 2011). Many forests that were mainly dominated by conifers are now either mixed or dominated by hardwoods. Moreover, the distribution of tree species is expected to be altered by global change (Brown and Vellend 2014), notably as a result of migration beyond their current northern limits (Woodall et al. 2009). The migration process will be in part conditioned by the biotic interactions such as competitive dominance or facilitation (Svenning et al. 2014; Putnam and Reich 2017).
Few studies have documented the effects these changes have on soil properties, even though vegetation has long been known to play an important role in soil nutrient dynamics (Jenny 1941), through several factors including litter quality and quantity, nutrient uptake rates, rooting patterns and water interception (Ayres et al. 2009; Laganière et al. 2010, 2012). The rates of hardwood litter decomposition and nutrient cycling are generally higher than those of conifer litter (Augusto et al. 2015). N concentrations in the tree compartments (i.e. trunk, branches, foliage, roots) are higher in hardwoods than in conifers (Paré et al. 2013), and soil pH was found to be lower in conifer forests (Raulund-Rasmussen and Vejre 1995). Soil carbon is also more abundant and soil C/N ratio increases when ectomycorrhizal (ECM) fungi are present, which are often associated with conifer-dominated forests. In contrast, hardwood-dominated forests are more commonly colonized by arbuscular mycorrhizal (AM) fungi and associated with a lower C/N ratio (Averill et al. 2014).
Soil properties are another key determinant in regional species distribution, and could influence vegetation migratory responses (Lafleur et al. 2010). Since tree species have an important influence on soil properties, it is possible that shifts from conifer- to hardwood-dominated forests will in turn influence soil properties (Brown and Vellend 2014). In a southern boreal mixed forest, it was shown that the microbe densities and soil pH were lower under conifers than hardwood cover (Bauhus et al. 1998), and the increase in hardwood species could possibly relieve certain constraints for other species to extend (Brown and Vellend 2014).
The effects of tree species composition on soil properties have often been studied with common garden approaches (Reich et al. 2005; Vesterdal et al. 2008) from which only limited inference can be drawn to forest contexts. The plant-soil combinations of a common garden might not reflect combinations observed in the forest, for example. In addition, companion plants commonly found in the forests are rarely included in common ground experiments. On the other hand, studies conducted in forests rely on the assumptions that tree species have equal chances to grow on the same substrate, and that soil conditions were similar prior shifts in forest composition. Controlling for similarity in the parent material chemistry is thus mandatory in such cases (e.g. Finzi et al. 1998a). However, such an approach does not completely alleviate all uncertainties related to the intrinsic similarity of the soils that are studied.
We studied forest compositional changes on soil properties using a unique historical forest survey with very high spatial accuracy. More specifically, we evaluated the links between increases in hardwood species (or decrease in conifers) and chemical composition of the soil. Because soil processes are slow in the northern temperate forests, we hypothesized that 1) there are strong correlations between both current and past forest composition and soil properties and 2) that shifts in stand composition influence soil properties. Several partial redundancy analyses (partial RDA) were used to relate the chemical properties of the soil to current and past forest compositions as well as to shifts in the composition in order to test our hypotheses. We expected that modifications of the soil properties following an increase in hardwood proportion enhanced nutrient availability.
Materials and methods
Study area
The study area is located in the eastern balsam fir-yellow birch bioclimatic subdomain of the province of Quebec (Fig. 1). The climate is subpolar subhumid, continental (Robitaille and Saucier 1998) with a growing season length of 160 days. Average annual rainfall ranges between 900 and 1100 mm, snow cover between 350 and 400 cm and temperature averages 2.5 °C annually. The study area is generally covered by glacial till. The water regime is mainly mesic and the mean altitude is around 200 m (Robitaille and Saucier 1998).

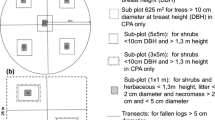
Map of the study area with location the 1930’s forest inventory plots and the schematic representation of a plot
In 1930, the Price Brothers’ Company conducted a systematic forest inventory in the study area to evaluate timber volume (Terrail et al. 2014). Approximately 5000 plots of 1012 m2 (0.5 chains or 10.06 m wide (1 chain = 20.12 m), by 5 chains or 100.58 m long) were measured along transect lines with 100 to 300 m between each plot, and approximately 120 m separating each transect. The diameter at breast height (stem diameter at 1.3 m above the ground, DBH, measured in 2 in. classes (51 mm)) was measured for each tree with a minimum DBH of 3 in. (76.2 mm) (Terrail et al. 2014). Some species were grouped at the genus level following Terrail et al. (2014) (e.g. red maple and sugar maple were grouped as “Maple”, and white spruce, red spruce (Picea rubens Sarg.) and black spruce (Picea mariana (Mill.) B. S. P.) as “Spruce”). Shrub species were not inventoried. As the inventory was conducted by land surveyors, the records were accurate enough to relocate the exact position of the plots (Terrail et al. 2014).
Forest inventory
A subset of 130 plots of the Price Brothers’ Company systematic inventory were selected using the forest maps developed by the Quebec’s Department of Forests (Ministère des Forêts, de la Faune et des Parcs du Québec 2012). The following criteria were used to select the revisited sample plots: (i) the plots had to be on public land, which represents 51% of the 22,681 km2 of forested land in the region; (ii) the stands had to have balsam fir (Abies balsamea (L.) Mill.) and/or sugar maple as part of the dominant cover (these two species are both economically and ecologically important in the study region); (iii) the mapped stand age had to be 50 years old or greater; and (iv) the plots had to be accessible by all-terrain vehicle. A total of 130 of the 300 plots that met the selection criteria were randomly selected and visited between 2012 and 2014.
In each of the revisited sample plots, the original 100 m transect was walked, and only the largest homogeneous part (hereafter designated as main plot), based on slope and vegetation, was kept in order to have consistent ecological conditions. Two variable radius subsampling plots were measured with a factor 2 prism placed at 1/3 and 2/3 of the main plot length to obtain species composition and stand basal area (BA in m2/ha). The DBH of each tree included into the subplot was measured using a DBH measuring tape (in mm). Trees with a DBH below 76.2 mm (i.e. 3 in.) were not measured, since it was the lower limit of the 1930 inventory. If the main plots were smaller than 50 m in length, only one subplot was established at the halfway mark, to avoid trees being included into two subplots. The basal area per species was obtained by averaging the species basal area of each subplot (Table 1), and was used as an indicator of current species composition. Similarly to the 1930 inventory, shrub species were ignored and some species were grouped at the genus level (e.g. Maple and spruce group). The basal area of sugar maple represented 93% of the Maple group, and red maple only 7% in our data. The changes in stand composition of each plot were calculated by subtracting the BA of each species or group measured in 2012–14 from the BA measured in 1930 (Table 1). To reduce the number of explanatory variables and develop parsimonious models, some rare species were measured but not retained in the analysis (e.g. Pinus strobus and Larix laricina: one occurrence, Fraxinus americana and Fraxinus nigra: two occurrences) and balsam poplar (Populus balsamifera L.) was grouped with quaking aspen as Populus group.
A soil pit was dug in the middle of each of the 130 main plots until the C-horizon was reached. The soil profile was described using the FAO soil reference (W.R.B. and I.W.G. 2006) and thickness of each horizon, rooting depth, presence and proportion of redox spots were measured. In the study area, soils are at the interface between orthic podzols and dystric cambisols, with frequently the presence of a partial pale coloured eluvial horizon. Samples from the O, B and C-horizons were collected. Two additional samples from the O and B-horizons were also collected at 1/4 and 3/4 of the main plot length. About 250 g of the three samples obtained from each of the O and B-horizons were pooled in airtight plastic bags and brought back to the laboratory for analysis.
Chemical analysis
The soil samples were kept frozen until thawed at room temperature during 48 h in plastic bags in order to keep their water content. They were then passed through a 4 mm sieve. The pH was measured in distilled water on the fresh samples (McKeague 1976). Inorganic nitrogen concentrations (NO3− and NH4+) of the O and B-horizons were measured on a 5 g dry weight equivalent soil sample extracted in 50 ml 2 M KCl. Samples were stirred and then, vacuum-filtered on a Whatman No. 42 filter. The first extraction was done following the 48 h thaw, and the second after 60 days of incubation at room temperature in complete darkness, with samples covered by a polyethylene film in order to limit evaporation (Légaré et al. 2005). Concentrations of nitrate (NO3−) and ammonium (NH4+) were determined by FIA (Quickchem 8500, Lachat Instruments, Loveland, Colorado) using the method described by Maynard et al. (2007). Net nitrification and net ammonification were calculated as the net changes in NO3− and NH4+ between the incubated and non-incubated samples. Because correlations among the four N variables were high (i.e. r = 0.86 for incubated-NH4+ vs. non-incubated-NH4+, and r = 0.65 for NO3−), we only considered the net nitrification (NIT) and initial NH4− as an indicator of initial available N.
The concentrations in exchangeable cations (Ca2+, K+, Mg2+, Na+, Fe3+, Mn2+, Al3+) and extractable P were obtained for both the O, B and C-horizons, following a Mehlich III extraction (Ziadi and Sen 2007) and determination by ICP-AES (Optical Emission Spectrometer, Perkin Elmer, Optima). The cation exchange capacity (CEC) was calculated as the sum of exchangeable cations (Hendershot et al. 2007). As it was later found to be mostly dependent on Ca2+ (r = 0.85) and Mg2+ (r = 0.72), we chose to use Ca2+ and Mg2+ instead of the CEC in further analyses. A summary of the soil data is presented in Table 2.
Total C and N were determined by dry combustion using a TruMac CNS analyzer (LECO Corporation, St. Joseph, MI, USA) in both O and B-horizons. Finally, the concentrations of acid-extractable elements (Ca2+, K+, Mg2+, Na+, Fe3+, Mn2+, Al3+) in the C-horizon, a proxy for total element content of the parent material, were determined by ICP-AES on the fine fraction (< 177 μm) obtained by dry sieving after heating at 550 °C for 1 h, and extraction with nitric acid (2 M) for 1 h by soaking (Thiffault et al. 2013).
Soil texture
A subsample of the C-horizon was sieved at 2 mm. Then, the hydrometer method was used to determine the soil particle size distribution on the 55 inventoried in 2013–14 (Kroetsch and Wang 2007). The statistical inference method called Dirichlet regression (Hijazi and Jernigan 2009) was then used to predict the proportion of sand, silt and clay in each of the 130 plots (see Supporting information: method S1 and data summary in Table 2). As the sum of the three types of grain size is mathematically equal to 100%, intercorrelation problems within the covariates matrix are highly probable (see section 6.3.2.6 of Borcard et al. 2011). Thus, only the percentage of silt was kept for further analysis, because preliminary results indicated it had the highest discriminating power between plots.
Data analysis
Partial redundancy analysis (partial RDA) (Legendre and Legendre 2012) were used to assess the links between soil chemistry (response matrix Y), past and present tree species composition and changes in composition between each inventory. RDA is a multivariate analysis combining multiple regression with classical ordination (Legendre et al. 2011). Details can be found in Section 11.1 of Legendre and Legendre (2012). Triplots are then used to highlight the trends in the RDA results. The significance of each canonical axis is tested and then the percentage of variance explained by each canonical axis is corrected by the R2adj of each model (Borcard et al. 2011). Symmetric scaling was used to draw the triplots. It is a compromise between distance and correlation biplots, where both sites and species scores are scaled by eigenvalues. Partial RDA decomposes the X matrices of the RDA into two groups: the variables of interest (explanatory matrix X) and variables used to control W (covariates matrix W, Borcard et al. 2011). A significance level of 5% was used when testing for the statistical significance of partial RDA equations and axis. Variation partitioning correcting by an R2adj was used to assess the importance of each group of variables (i.e. matrices X and W) in the partial RDA (Peres-Neto et al. 2006) with the ‘varpart’ function of the vegan package version 2.4–1 (Oksanen et al. 2016). Finally, multicollinearity between explanatory variables was assessed using the ‘vif.cca’ function (in vegan 2.4–1), as described by Borcard et al. (2011). Analyzes were done using the ‘vegan’ package (Oksanen et al. 2016) in the R statistical software (V. 3.3.0, R Development Core Team 2016). The variable matrices used in this analysis are summarized in Table 3.
A Hellinger transformation was applied to current and past BA, in order to give low weights to rare species (Legendre and Gallagher 2001). Changes in BA as well as the concentrations of each soil chemical element were centered and scaled prior to conducting the partial RDA.
Finally, effect of vegetation cover changes between 1930 and 2012–14 on soil properties was also tested with multivariate analysis of variance (MANOVA). The categories of vegetation cover changes were set as follows: an increase greater than 25% in conifer species, an increase greater than 25% in hardwood species or limited changes (i.e. < 25%). A Tukey HSD multiple comparison was then applied when significant differences were detected by the MANOVA.
Results
Links between soil properties and current tree species composition
A first partial RDA was used to assess the relationship between the chemical composition of the O or B-horizon (response matrix Y) and the basal area per species measured in the most recent inventory (explanatory matrix X), while the properties of the C-horizon were used as the covariate matrix (W).
For the O-horizon, the variance partitioning showed that the current tree species composition and C-horizon explained, respectively, 8.9 and 14.9% of the total variance of the O-horizon chemical properties, and the intersection of these matrices explained 6.4% (Table 4). An ANOVA test showed that the first two ordination axes of the partial RDA were significant. The first axis explained 5.5% of the total variation and the second 1.9% (Table 4). Triplot scores for each of the constraining variables (summarized in Table 5) are used further to describe each of the triplots. The triplot of the O-horizon (Fig. 2a) showed that two groups of tree species played an important role in the dispersion of the sites along the first axis. Tree species associated with conifer-dominated stands (balsam fir and spruce group) were clearly located on the left-hand side of the triplot, while hardwood-dominated stands of the Maple group and yellow birch (Betula alleghaniensis Britton) appeared on the right-hand side. Balsam fir (−0.65 on the first axis) was positively correlated with the C/N ratio. The spruce group was also close to C/N ratio and was positively correlated with concentrations of exchangeable Al. Paper birch (Betula papyrifera Marsh.), which was close to balsam fir, was strongly correlated with exchangeable K. On the opposite side, the maple group was slightly correlated with NIT (net nitrification) and negatively correlated with balsam fir and C/N ratio. The dispersion of sites along the second axis depicted a gradient in tree species composition with stands dominated by northern white-cedar located at the bottom. On the opposite side, the dispersion of sites was mostly related to a higher prevalence of yellow birch. Northern white-cedar had a slightly shorter projection, mostly on the second axis (−0.28), and was positively correlated with exchangeable Na. Exchangeable Mg and Ca and pH formed a group of variables located between the maple group and northern white-cedar. Yellow birch had similar projections on both axes (0.53 on axis one, 0.41 on axis two) and was correlated with exchangeable Mn, extractable P and with initial NH4+. Finally, the Populus group had short projections on both axes (−0.18 and − 0.12).

a Triplot from the first partial RDA (equation 1) with O-horizon as the response matrix, current tree species composition as the explanatory matrix and C-horizon chemistry as the covariate matrix. In blue: explanatory variables with BAF: Balsam fir, PAB: Paper birch, YEB: Yellow birch, Map.: Maple group, CED: Northern white-cedar, Spr.: Spruce group, Po.: Populus group. In red: O-horizon chemistry response variables (matrix content is summarized in Table 3). b Triplots from the first partial RDA (equation 1) with B-horizon as the response matrix, current tree species composition as the explanatory matrix and C-horizon chemistry as the covariate matrix. In blue: explanatory variables with BAF: Balsam fir, PAB: Paper birch, YEB: Yellow birch, Map.: Maple group, CED: Northern white-cedar, Spr.: Spruce group, Po.: Populus group. In red: B-horizon chemistry response variables (matrix content is summarized in Table 3)
When the B-horizon was used as the response matrix, the 2012–14 species composition matrix explained 5.9% of the total variance, while the C-horizon explained 22.3%. The intersection of both matrices explained a further 7.7% of the variation (Table 4). The two first ordination axes were found to be statistically significant (Table 4) and they explained 2.9 and 1.6% of the total variation, respectively. In the B-horizon triplots (Fig. 2b), maple and balsam fir remained important variables in the dispersion of the sites along the first axis (respectively 0.52 and − 0.47). Balsam fir was correlated with the C/N ratio. The projection of the spruce group was not as important as for balsam fir, but was also correlated with C/N ratio. On the opposite side, maple was correlated with net nitrification, initial NH4+ and P. Every tree species, except maple and Populus groups were located on the left-hand side of axis 1. Paper birch had similar projections on axes one and two (−0.43 and − 0.41), similarly to northern white-cedar (−0.52 on axis one and 0.43 on axis two).
The triplot diagram (Fig. 2b) depicted some consistent relationships between tree species and chemical properties of the B-horizon. High C/N values were mostly associated with dominant balsam fir, spruce group. High values of exchangeable K were closely related to the Populus group. The links between the chemical properties of the B-horizon and tree species were not as well defined as for the O-horizon, although some links were apparent between northern white-cedar and high pH and exchangeable Ca.
Links between soil properties and past tree species composition
A second partial RDA was used to assess the relationship between the chemical composition of the O or B-horizon (response matrix Y) and the 1930 tree species composition (explanatory matrix X), but this time with both the properties of the C-horizon and the tree species composition of 2012–14 used as covariates matrices (W).
For the O-horizon, the overall model was not significant. Variance partitioning confirmed that the 1930 tree species composition explained almost none of the total variance (0.3%), while the tree species composition of 2012–14 and the C-horizon chemical composition (covariates matrices W) were found to explain respectively 8.2 and 12.4% of the variance in the chemical soil composition. The amount of variation explained by the intersection of the three matrices was also negligible (Table 4).
The partial RDA applied to the B-horizon was also not significant. The 1930 species composition explained none of the variance (0%), whereas the 2012–14 tree species composition explained 6.1% of the total variance, and the C-horizon matrix explained the largest part (21.1%). The intersection of the three matrices remained small in this case (0.15%, Table 4).
Links between soil properties and shifts in tree species composition
The link between the shifts in stand composition and the soil properties of both the O and B-horizons (response matrix Y) was established with another partial RDA using the differences in species composition as the explanatory matrix X and C-horizon properties as the covariates matrix W. For the O-horizon, the shifts in stand composition explained 4.2% of the observed variance, while the properties of the C-horizon accounted for 15.1% of the variance (Table 4). A further 5.6% was explained jointly by the two matrices. In the case of the B-horizon, the shifts in stand composition explained almost the same proportion of the observed variance (3.7%, Table 4). The properties of the C-horizon however explained more of the observed variance of the chemical composition of the B-horizon (23.4%), when compared to the O-horizon, and 7.5% was explained jointly by the two matrices. For both analyses, only the first ordination axis was statistically significant, and thus the triplots are not presented. In the case of the O-horizon, the only significant axis represented 2.5% of the total variation. The dispersion of the sites along this axis was mainly influenced by the shifts from balsam fir (−0.74) to maple (0.60) (Table 5). The shifts in balsam fir basal area were correlated with the C/N ratio, and those of the maple group were correlated with net nitrification. In case of an increase in hardwood species, MANOVA results indicated that NIT and exchangeable Mn in the O-horizon were significantly higher and C/N ratio was significantly lower than for the two other categories of changes in forest cover (Fig. 3). Exchangeable P was higher in the case of an increase in hardwoods when compared to an increase in conifers, unlike Al and K which were higher when conifer species increased. No significant differences between shifts in canopy composition were observed for pH, initial NH4+ and exchangeable Na, Mg, Ca, Fe. Finally, Na was higher in the case of limited changes when compared to an increase in hardwoods.

Variation of O-horizon properties by categories of vegetation cover changes between 1930 and 2012–14. Different Letters indicate a significant difference (Tukey HSD, p < 0.05). NIT is the net nitrification, CN is the C/N ratio and NH4 is the initial NH4+
For the B-horizon, the first axis explained 1.8% of the total variation, which was mostly related to changes in maple group basal area (0.66). This shift was correlated with net nitrification and slightly with extractable P.
Discussion
Our results provided evidence that while parent material and current vegetation explain much of the variation in current soil properties, shifts in forest composition over eight decades also contribute to the observed variation. Many studies have shown that North American forests have undergone an increase in the rate of disturbances since the European settlement (Fuller et al. 1998; Dupuis et al. 2011; Nowacki and Abrams 2015). With the use of multivariate RDA models, this study investigates how the current vegetation-soil relationships are affected by the previous stand composition, and by the shifts in vegetation composition these stands have undergone since eighty years.
All partial RDA models underlined the importance of the C-horizon characteristics on the O- an B-horizons properties, as observed by Finzi et al. (1998a). The current vegetation composition followed as the next important driver of the O- and B-horizons properties, while the link with past vegetation was almost non-existent. The chemical compositions of the B- and C-horizons were more similar than those of the O- and C-horizons. The B-horizon is composed of C-horizon materials that have undergone pedological processes of weathering and organic matter accumulation. The O-horizon, being mostly composed of organic material, is greatly influenced by the current vegetation, especially through litter decomposition.
Despite the fact that texture is an important property of the soil, there is no clear consensus on its effect on the plant – soil relationships. For example, Raulund-Rasmussen and Vejre (1995) found a stronger species effect on loamy sites, whereas Ste-Marie et al. (2007) reported a stronger species effect on coarse-textured soil. In our dataset, soil texture varied only slightly between-sites so this variable did not contribute to our soil-vegetation models. However, the use of smoothed predicted values based on the 55 measured soil texture plots could lead to a slight underestimation of inter-plot variability, compared to a case in which soil texture would have been measured on all 130 plots.
This study has provided evidence that shifts in stand composition between 1930 and today explained a small part of the variability in the properties of the O and B-horizons (4.2 and 3.7%, respectively). Overall, the correlations between shifts in species composition and soil properties for both horizons were not as marked as for the correlation of the soil properties with the composition of the current vegetation (8.9 and 5.9% for O and B-horizons respectively). These last results are however in agreement with what is known about the current vegetation – soil relationship.
A strong correlation between the increase in balsam fir and the C/N ratio in both O and B-horizons was observed. A high C/N ratio is an indication of a low rate of organic matter decomposition (Finzi et al. 1998b). Balsam fir’s litter has high tannin concentrations which can inhibit microbial activity (Lorenz et al. 2000), and more specifically nitrification (Baldwin et al. 1983). Therefore, a decrease in available N is expected. Tannins also reduce available N by precipitating some forms of N (Adamczyk et al. 2011), which overall may have a great influence on the regulation of C and N compounds transformation (Smolander et al. 2012). MANOVA results confirmed that with an increase in hardwood species, net nitrification is higher and C/N ratio is lower. However, unlike the RDA, the MANOVA does not consider the parent material in the analysis and thus must be interpreted carefully.
Our results indicate that an increase in maple species was positively correlated with net nitrification in the O-horizon. High concentrations of NO3− in the mineral soil associated with maples could in part be due to a low demand for N by mature sugar maple (Foster et al. 1989). Furthermore, sugar maple is known to have low soil C/N and high level of NO3− production and loss (Lovett et al. 2004), which supports our observations. In the B-horizon, an increase in sugar maple was found to be positively correlated with extractable P. Sugar maple provides litter that is overall lower in base cations than that of other hardwood species (Côté and Fyles 1994; Bal et al. 2015), and soil P is mainly governed by soil parent material (Vitousek et al. 2010). P is an essential nutrient for sugar maple growth and vigor (St.Clair et al. 2008), which could explain a better development of maple on P-rich soils.
In the case of an increase in maple species, the links between tree species and soil chemical elements differ slightly when considering the O- or B-horizons. Interspecific differences between horizons have been reported for various species (Alban 1982; Finzi et al. 1998a). They may be attributable in a part to the fact that different tree species take up nutrients (i.e. nitrogen) at different soil depths (Houle et al. 2014).
The association of plants to either ECM or AM fungi has a significant impact on litter decomposition and soil C storage (Averill et al. 2014). Storage of soil carbon is greater in stands dominated by ECM-associated plants than in those dominated by AM fungi. In addition, the litter from AM-associated plants is known to decompose faster (Fisher et al. 2016). Similarly to what was found by Brown and Vellend (2014), the presence of balsam fir, which typically forms associations with ectomycorrhizal (ECM) fungi (Villeneuve et al. 1989; Klironomos 1995), was associated with lower pH and higher C/N ratio. In turn, these conditions may partially explain the antagonism between balsam fir and sugar maple, which is typically associated with arbuscular mycorrhizal (AM) fungi.
The tree population and the community dynamics are key drivers in nutrient cycling (Finzi et al. 1998b). The association of soil properties with stands dominated by maple or balsam fir are relevant in the context of the marked increase of maple dominance in numerous forests of Québec (Dupuis et al. 2011). Because no significant effects of the past stand cover were detected on the current soil properties, our results suggest that the legacies of past stand cover have disappeared after 80 years. This is beyond the 30 years’ time frame over which species composition influences the soil properties in a temperate climate context (Reich et al. 2005).
In the context of global change, forest ecosystem functions and attributes may be altered because tree species may migrate beyond their northern limits (Woodall et al. 2009). Other factors such as elevation, soil properties or biotic interactions are also among the key drivers of the distribution of species (Brown and Vellend 2014; Svenning et al. 2014; Putnam and Reich 2017). In this study, the range of soil conditions does not appear to be wide enough to constrain species establishment. At a regional scale, edaphic factors may have an important impact on the local distribution of species by constraining their migratory response (Lafleur et al. 2010). Soil nutrient availability could interfere in vegetation migration (Brown and Vellend 2014) and explain the time lag in the soil properties’ response (Pennington 1986). Nevertheless, our results suggest that vegetation shifts have modified soil conditions, but these changes are not as important as the combined influence of current vegetation and soil properties. The methodology described here could be used to assess the impacts of these changes, which could be particularly relevant as climate is expected to alter the soil properties over the long term (Brown and Vellend 2014). A better understanding of plant-soil relationships is an increasingly relevant issue in a species migration context.
Conclusion
Using a unique historical survey, our approach yields insight into the factors that influence soil properties in a changing environment. Our results showed that in the study area, the parent material had the largest influence on soil properties, followed by current vegetation. Although past vegetation composition had little or no influence on soil properties, our results provided evidence that shifts in vegetation over time could have a significant influence. Such changes could be related to an alteration of soil organic matter and nutrient dynamics, changes in litter quality, and changes in mycorrhizal fungi composition with vegetation shifts. These changes could lead to modifications in soil N cycling, litter decomposition and soil C storage (Averill et al. 2014; Fisher et al. 2016). In eastern Québec as in other regions, stands dominated by hardwood species such as sugar maple are gaining importance over conifer-dominated stands (Dupuis et al. 2011; Brown and Vellend 2014; Nowacki and Abrams 2015). Consequent changes in the upper soil properties may be observed, such as an increase in nitrate and a decrease in soil C/N ratio. The effects on nutrient cycling, elemental exports to watersheds, soil C storage, and further facilitation in vegetation shifts remain to be investigated.
References
Adamczyk B, Adamczyk S, Smolander A, Kitunen V (2011) Tannic acid and Norway spruce condensed tannins can precipitate various organic nitrogen compounds. Soil Biol Biochem 43:628–637
Alban DH (1982) Effects of nutrient accumulation by aspen, spruce, and pine on soil properties. Soil Sci Soc Am J 46:853–861
Augusto L, De Schrijver A, Vesterdal L, Smolander A, Prescott C, Ranger J (2015) Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol Rev 90:444–466
Averill C, Turner BL, Finzi AC (2014) Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 505:543–545
Ayres E, Steltzer H, Berg S, Wallenstein MD, Simmons BL, Wall DH (2009) Tree species traits influence soil physical, chemical, and biological properties in high elevation forests. PLoS One 4:e5964
Bal TL, Storer AJ, Jurgensen MF, Doskey PV, Amacher MC (2015) Nutrient stress predisposes and contributes to sugar maple dieback across its northern range: a review. Forestry: An International Journal of Forest Research 88:64–83
Baldwin IT, Olson RK, Reiners WA (1983) Protein binding phenolics and the inhibition of nitrification in subalpine balsam fir soils. Soil Biol Biochem 15:419–423
Bauhus J, Paré D, Côté L (1998) Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol Biochem 30:1077–1089
Borcard D, Gillet F, Legendre P (2011) Chapter 6: canonical ordination. In: Numerical ecology with R. Springer, New York
Boucher Y, Arseneault D, Sirois L (2006) Logging-induced change (1930–2002) of a preindustrial landscape at the northern range limit of northern hardwoods, eastern Canada. Can J For Res 36:505–517
Brown CD, Vellend M (2014) Non-climatic constraints on upper elevational plant range expansion under climate change. Proc R Soc Lond B Biol Sci 281:20141779
Côté B, Fyles JW (1994) Nutrient concentration and acid-base status of leaf litter of tree species characteristic of the hardwood forest of southern Quebec. Can J For Res 24:192–196
Danneyrolles V, Arseneault D, Bergeron Y (2016) Long-term compositional changes following partial disturbance revealed by the resurvey of logging concession limits in the northern temperate forest of eastern Canada. Can J For Res 46:943–949
Dupuis S, Arseneault D, Sirois L (2011) Change from pre-settlement to present-day forest composition reconstructed from early land survey records in eastern Québec, Canada. J Veg Sci 22:564–575
Finzi AC, Canham CD, Van Breemen N (1998a) Canopy tree-soil interactions within temperate forests: species effects on pH and cations. Ecol Appl 8:447–454
Finzi AC, Van Breemen N, Canham CD (1998b) Canopy tree-soil interactions within temperate forests: species effects on soil carbon and nitrogen. Ecol Appl 8:440–446
Fisher JB, Sweeney S, Brzostek ER, Evans TP, Johnson DJ, Myers JA, Bourg NA, Wolf AT, Howe RW, Phillips RP (2016) Tree-mycorrhizal associations detected remotely from canopy spectral properties. Glob Chang Biol 22:2596–2607
Foster NW, Hazlett PW, Nicolson JA, Morrison IK (1989) Ion leaching from a sugar maple forest in response to acidic deposition and nitrification. Water Air Soil Pollut 48:251–261
Fuller JL, Foster DR, McLachlan JS, Drake N (1998) Impact of human activity on regional Forest composition and dynamics in Central New England. Ecosystems 1:76–95
Held M, Jones-Held S (2014) Long-term Forest changes in northeastern Pennsylvania. Castanea 79:138–146
Hendershot W, Lalande H, Duquette M (2007) Ion exchange and exchangeable cations. In: Carter M, Gregorich E (eds) Soil sampling and methods of analysis, 2nd edn. CRC Press, Boca Raton, pp 173–178
Hijazi RH, Jernigan RW (2009) Modelling compositional data using Dirichlet regression models. Journal of Applied Probability & Statistics 4:77–91
Houle D, Moore JD, Ouimet R, Marty C (2014) Tree species partition N uptake by soil depth in boreal forests. Ecology 95:1127–1133
Jenny H (1941) Factors of soil formation: a system of quantitative pedology. McGraw-Hill, New York
Klironomos JN (1995) Arbuscular mycorrhizae of Acer saccharum in different soil types. Can J Bot 73:1824–1830
Kroetsch D, Wang C (2007) Particle size distribution. In: Carter M, Gregorich E (eds) Soil sampling and methods of analysis, 2nd edn. CRC Press, Boca Raton (USA)
Lafleur B, Paré D, Munson AD, Bergeron Y (2010) Response of northeastern north American forests to climate change: will soil conditions constrain tree species migration? Environ Rev 18:279–289
Laganière J, Paré D, Bradley RL (2010) How does a tree species influence litter decomposition? Separating the relative contribution of litter quality, litter mixing, and forest floor conditions. Can J For Res 40:465–475
Laganière J, Paré D, Bergeron Y, Chen HYH (2012) The effect of boreal forest composition on soil respiration is mediated through variations in soil temperature and C quality. Soil Biol Biochem 53:18–27
Légaré S, Paré D, Bergeron Y (2005) Influence of aspen on forest floor properties in black spruce-dominated stands. Plant Soil 275:207–220
Legendre P, Gallagher ED (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129:271–280
Legendre P, Legendre LFJ (2012) Chapter 11 - canonical analysis. In: Numerical ecology (3rd ed.), volume 24. Developments in environmental modelling. Elsevier, Oxford
Legendre P, Oksanen J, ter Braak CJF (2011) Testing the significance of canonical axes in redundancy analysis. Methods Ecol Evol 2:269–277
Lorenz K, Preston CM, Raspe S, Morrison IK, Feger KH (2000) Litter decomposition and humus characteristics in Canadian and German spruce ecosystems: information from tannin analysis and 13C CPMAS NMR. Soil Biol Biochem 32:779–792
Lovett GM, Weathers KC, Arthur MA, Schultz JC (2004) Nitrogen cycling in a northern hardwood forest: do species matter? Biogeochemistry 67:289–308
Maynard DG, Kalra YP, Crumbaugh JA (2007) Nitrate and exchangeable ammonium nitrogen. In: Carter M, Gregorich E (eds) Soil sampling and methods of analysis, 2nd edn. CRC Press, Boca Raton
McKeague JA (1976) Manual of soil sampling and methods of analysis. Canadian Society for Soil Science, Ottawa
Ministère des Forêts, de la Faune et des Parcs du Québec - Direction des inventaires forestiers - Forêt Québec (ACRIGéo). 2012. Couche écoforestière 22C08–0201, Inventaire du quatrième décennal
Nowacki GJ, Abrams MD (2015) Is climate an important driver of post-European vegetation change in the Eastern United States? Glob Chang Biol 21:314–334
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Henry M, Stevens H, Szoecs E, Wagner H (2016) Vegan : community ecology package. R Package version 2:4–1
Paré D, Bernier P, Lafleur B, Titus BD, Thiffault E, Maynard DG, Guo X (2013) Estimating stand-scale biomass, nutrient contents, and associated uncertainties for tree species of Canadian forests. Can J For Res 43:599–608
Pennington W (1986) Lags in adjustment of vegetation to climate caused by the pace of soil development. Evidence from Britain. Plant Ecol 67:105–118
Peres-Neto PR, Legendre P, Dray S, Borcard D (2006) Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology 87:2614–2625
Putnam RC, Reich PB (2017) Climate and competition affect growth and survival of transplanted sugar maple seedlings along a 1700-km gradient. Ecol Monogr 87:130–157
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Raulund-Rasmussen K, Vejre H (1995) Effect of tree species and soil properties on nutrient immobilization in the forest floor. Plant Soil 168:345–352
Reich PB, Oleksyn J, Modrzynski J, Mrozinski P, Hobbie SE, Eissenstat DM, Chorover J, Chadwick OA, Hale CM, Tjoelker MG (2005) Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species. Ecol Lett 8:811–818
Robitaille A, Saucier JP (1998) Paysages régionaux du Québec méridional. [Québec]: Gouvernement du Québec, Ministère des ressources naturelles, Sainte-Foy
Smolander A, Kanerva S, Adamczyk B, Kitunen V (2012) Nitrogen transformations in boreal forest soils—does composition of plant secondary compounds give any explanations? Plant Soil 350:1–26
St.Clair SB, Sharpe WE, Lynch JP (2008) Key interactions between nutrient limitation and climatic factors in temperate forests: a synthesis of the sugar maple literature. Can J For Res 38:401–414
Ste-Marie C, Paré D, Gagnon D (2007) The contrasting effects of Aspen and Jack pine on soil nutritional properties depend on parent material. Ecosystems 10:1299–1310
Svenning JC, Gravel D, Holt RD, Schurr FM, Thuiller W, Münkemüller T, Schiffers KH, Dullinger S, Edwards TC, Hickler T, Higgins SI, Nabel JEMS, Pagel J, Normand S (2014) The influence of interspecific interactions on species range expansion rates. Ecography 37:1198–1209
Terrail R, Arseneault D, Fortin MJ, Dupuis S, Boucher Y (2014) An early forest inventory indicates high accuracy of forest composition data in pre-settlement land survey records. J Veg Sci 25:691–702
Thiffault E, Paré D, Guindon L, Beaudoin A, Brais S, Leduc A, Michel JP (2013) Assessing forest soil base cation status and availability using lake and stream sediment geochemistry: a case study in Quebec (Canada). Geoderma 211:39–50
Thompson JR, Carpenter DN, Cogbill CV, Foster DR (2013) Four centuries of change in northeastern United States forests. PLoS One 8:e72540
Vesterdal L, Schmidt IK, Callesen I, Nilsson LO, Gundersen P (2008) Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For Ecol Manag 255:35–48
Villeneuve N, Grandtner MM, Fortin JA (1989) Frequency and diversity of ectomycorrhizal and saprophytic macrofungi in the Laurentide Mountains of Quebec. Can J Bot 67:2616–2629
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen–phosphorus interactions. Ecol Appl 20:5–15
W.R.B, I.W.G (2006) World reference base for soil resources. In: FAO (ed) World soil resources reports no. 103. Ed, 2nd edn. FAO, Rome
Whitney GG (1994) From coastal wilderness to fruited plain: a history of environmental change in temperate North America from 1500 to the present. Cambridge University Press, New York
Woodall CW, Oswalt CM, Westfall JA, Perry CH, Nelson MD, Finley AO (2009) An indicator of tree migration in forests of the eastern United States. For Ecol Manag 257:1434–1444
Ziadi N, Sen TT (2007) Mehlich 3-extractable elements. In: Carter M, Gregorich E (eds) Soil sampling and methods of analysis, 2nd edn. CRC Press, Boca Raton
Acknowledgments
We thank the Fonds de Recherche du Québec - Nature et Technologies and the Groupe Lebel for funding. We also thank Serge Rousseau of the Laurentian Forestry Centre for the help in soil chemical analyzes, and the CEF for its support. We also sincerely thank Alain Caron for his valuable support in the redundancy analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Andrea Schnepf.
Electronic supplementary material
ESM 1
(DOCX 26.7 kb)
Rights and permissions
About this article

Cite this article
Gauthray-Guyénet, V., Schneider, R., Paré, D. et al. Influence of shifts over an 80-year period in forest composition on soil properties. Plant Soil 433, 111–125 (2018). https://doi.org/10.1007/s11104-018-3819-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-3819-y