
Abstract
Background and aims
In recent years, ecologists have tried to determine the importance of intraspecific trait variation (ITV) versus community composition change (CCC) in shifts in community functional composition. However, results to date have not provided generality.
Methods
A six-year nitrogen (N) fertilization experiment was conducted in species-rich alpine meadow communities. We focused on five key functional traits (height, leaf dry mass content LDMC, specific leaf area SLA, leaf nitrogen content LNC, and leaf phosphorus content LPC) that belong to three trait categories (whole-plant level trait, leaf morphology trait and leaf chemical trait) and measured biomass of each species in community. A sum of squares decomposition method was used to distinguish the relative contribution of ITV versus CCC to community weighted mean (CWM) traits.
Results
Our results showed that N fertilization led to increasing heightCWM and LNCCWM due to light competition intensification and soil nutrient enrichment. However, the responses of community-wide SLA, LDMC and LPC were highly inconsistent and depended on the balance of different opposing processes. Moreover, during short-term fertilization, ITV played a more important role in mediating functional composition changes in all traits, but the effects of CCC overwhelmed ITV and became more important in determining community-wide whole-plant level trait and leaf morphology traits (height, LDMC and SLA) in subsequent fertilization years. On the other hand, ITV always played a more important role than CCC in determining the community-wide leaf chemical traits (LNC and LPC), but CCC had a greater contribution than ITV in terms of explaining shifts in whole-plant traits (height) and leaf morphology traits (LDMC and SLA). In addition, both positive and negative covariations appeared in our study, indicating that community-wide trait shifts due to ITV and CCC may either reinforce or oppose one another, depending on different trait categories.
Conclusions
These findings highlight the importance of ITV, CCC and their covariation in mediating community functional composition, and the relative importance of ITV effects and CCC effects depended on fertilization duration and trait category. This study significantly improve our understanding of the mechanisms governing the shifts in community functional composition under N deposition scenarios.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The trait-based approach has been regarded as a useful approaches to detect the general rules of complex communities (McGill et al. 2006); therefore, studies on functional traits have significantly increased in the past two decades (Westoby and Wright 2006; Lefcheck and Duffy 2015; Carmona et al. 2017). In recent years, ecological studies exploring the extent to which intraspecific trait variation (ITV) versus community composition change (CCC) contribute to shift in community functional composition, particularly under different manipulated environment (Niu et al. 2016; Siefert and Ritchie 2016) and spatio-temporal scales (Riva et al. 2015; Volf et al. 2016), have become increasingly popular. Previous studies have noted that changes in community functional composition can be broken down into three components (Lepš et al. 2011): ITV, which is derived from phenotypic plasticity and genetic variation (Münzbergová et al. 2017); CCC, which is driven by species presence or absence and the change in relative abundance (Riva et al. 2015; Niu et al. 2016); and the covariation between ITV and CCC (de Bello et al. 2011; Lepš et al. 2011).
Recently, an increasing number of studies were undertaken in both natural and semi-natural communities to distinguish the relative contribution of ITV versus CCC on change in community functional composition (Riva et al. 2015; Niu et al. 2016; Siefert and Ritchie 2016; Volf et al. 2016). In their research on a natural forest ecosystem, Fajardo and Piper (2011) found that environmental alterations could cause trait variation without species turnover in a widespread tree species. In contrast, Riva et al. (2015) showed that variation in functional structure along soil water storage gradients in Mediterranean forest communities was mainly promoted by species turnover rather than by ITV. Studies on functional composition changes in semi-natural communities have mainly focused on mowing, fertilization and grazing habitats. In 2016, in a study including long- and short-term management, Volf et al. (2016) found that the relative contribution of ITV versus CCC to community functional composition changes in a meadow depended on both the time scale of management techniques (mowing, grazing and abandonment) and the studied traits. Meanwhile, Siefert and Ritchie (2016) conducted a three-year nutrient addition experiment and noted that the shifts in community-wide traits were driven almost entirely by ITV. In a grazed alpine meadow, Niu et al. (2016) demonstrated that functional composition responses to grazing were best captured by ITV and not changes in community composition, although ITV was consistently correlated with CCC for all traits. In short, many studies have sought to clarify the relative role of ITV and CCC in determining community functional composition; however, a definitive conclusion has not yet been drawn, perhaps because the effects of ITV and CCC change with management types, management duration and the studied traits.
Nitrogen fertilization, one of the most common management techniques in pastures, has been widely used to improve grassland production and often induced variation in community functional composition. Several previous studies have shown that N fertilization strongly affects community functional composition via ITV, CCC, and the covariation between them (Lepš et al. 2011; Siefert and Ritchie 2016). Following N addition, the increasing light competition caused by nutrient enrichment and shading intensity (Newman 1973; Hautier et al. 2009) favours exploitative species with tall stature, high SLA, high LNC and low LDMC, while it hinders conservative species with short stature, low SLA, low LNC and high LDMC (Schellberg and Pontes 2012; Zhou et al. 2016). This leads to changes in species relative abundance and species loss. On the other hand, N fertilization could result in plastic responses in individual plants, competitive changes among different individuals, and shifts in the relative abundance of genotypic variants within species (Albert et al. 2010). Finally, the processes of CCC and ITV combine and interact (positive covariation or negative covariation) to determine functional composition in fertilization communities (Lepš et al. 2011; Siefert and Ritchie 2016).
Previous studies have also displayed that the effects of N fertilization on the relative importance of CCC and ITV further relied on fertilization duration. Several studies have demonstrated that long periods of anthropogenic perturbation should cause functional composition variation via CCC, whereas short time scales of anthropogenic perturbation should mainly influence ITV (Auger and Shipley 2013; Jung et al. 2014; Siefert and Ritchie 2016), thus ITV can be regarded as a buffer against changes in species composition. Additionally, although there is generally some CCC in response to short-term fertilization, it usually reflects variations in species relative abundance rather than shifts in the presence and/or absence of species (Smith et al. 2009; Zhang et al. 2015), but long-term fertilization often causes the loss of rare species (Clark and Tilman 2008; Isbell et al. 2013).
Recent studies have also demonstrated that the responses of ITV and CCC to fertilization depended on trait categories, such as whole-plant traits versus organ-level traits, or leaf chemical traits versus leaf morphology traits (Siefert et al. 2015). According to the assumption of the optimisation model (Marks 2007; Siefert et al. 2015), whole-plant traits (e.g., height) that are measured at higher levels of plant integration should be highly sensitive to environmental disturbance (e.g., fertilization) and thus display high ITV. In contrast, organ-level traits should be more conserved; thus, the changes in functional composition would mainly be caused by CCC (Marks 2007, Siefert et al. 2015). On the other hand, previous studies have shown that leaf chemical traits (e.g., LNC, leaf phosphorus (P) content LPC) are highly labile within species and are sensitive to nutrient enrichment, whereas leaf morphology traits (e.g., SLA, LDMC) tend to be more stable (Rozendaal et al. 2006; Kazakou et al. 2014).
In short, the variations in community functional composition in nutrient enrichment conditions are promoted via interacting processes between ITV and CCC and depended on both fertilization duration and trait category. However, studies that account for these factors, particularly in a species-rich alpine meadow community, are still scarce. In the present study, we conducted a six-year N fertilization experiment and measured five important functional traits (one whole-plant trait (height), two leaf morphology traits (SLA, LDMC) and two leaf chemical traits (LNC, LPC)) in an alpine meadow of Tibetan plateau to assess the relative contribution of ITV and CCC to functional composition in different years. Specifically, the following two hypotheses were tested:
-
i.
ITV plays a critical role in mediating community functional composition and alleviating changes to community species composition during short-term (1–3 year) fertilization, but the effects of CCC would overwhelm ITV and become more important during long-term (> 3 year) fertilization.
-
ii.
The shifts in relative importance of ITV and CCC depend on different categories of functional traits. For leaf chemical traits, ITV plays a more important role than CCC in determining functional composition. In contrast, CCC has a greater contribution to functional replacement than ITV in terms of leaf morphology traits.
Materials and methods
Study site
The study was carried out at the Alpine Meadow and Wetland Ecosystems Research Station (Maqu branch) of Lanzhou University, in the eastern Tibetan Plateau (35°580′N, 101°530′E, altitude 3500 m a.s.l.) of Gansu, China. The mean annual temperature is 1.2 °C, ranging from −10 °C in January to 11.7 °C in July, and the mean annual precipitation for the period of 1975–2010 was 620 mm, which mainly fell during the short, cool summer. The area has 2580 h of sunshine and more than 270 days of frost per year (Luo et al. 2006). The vegetation is that of a typical alpine meadow and is dominated by Kobresia graminifolia (Cyperaceae), Elymus nutans (Poaceae), Anemone rivularis (Ranunculaceae), Poa poophagorum (Poaceae), Festuca ovina (Poaceae), and Carex kansuensis (Cyperaceae). The average above-ground dry biomass is 360–560 g m−2. The dominant animals in the area include livestock (e.g., yaks, Tibetan sheep and horses), marmots (Marmota himalayana) and zokor (Myospalax spp.) .
Experimental design
The fertilization experiment was initiated in April 2011 in an enclosed area of flat alpine meadow, where grazing was conducted during the non-growing season. One fertilization treatment and one control treatment (without any fertilization) were conducted, and each treatment was replicated six times. The analytical reagent ammonium nitrate (NH4NO3) was used as fertilizer, at a fertilization dose of 10 g N / m2 per year. In early May 2011, twelve 20 × 10 m plots, each separated by 1 m, were established in a 100× 100 m area of homogeneous meadow, and each plot was divided into two 10 × 10 m subplots; one subplot was used to measure species abundance, and the other subplot was used to measure functional traits. Fertilizer was applied annually at the early May on drizzly days to avoid the need for watering.
Species abundance measurements
From 2011 to 2016, in the middle of August, a 0.5 × 0.5 m quadrat was harvested for replicates of each treatment, and the number of individuals of each species was counted. For clonal plants, the term individual refers to a ramet (Cheplick 1989), which are equivalent to tillers in graminoids and rosettes or rooting branches in forbs. The green, above-ground parts (stems and leaves) were then clipped and sorted by species and brought to the laboratory. The green parts were dried at 60 °C for 48 hand weighed (0.01 g) to estimate the biomass of each species.
Functional trait measurements
According to the standard method of Cornelissen et al. (2003) and Pérez-Harguindeguy et al. (2013), we chose five functional traits: height, SLA, LDMC, LNC and LPC, all of which can be easily measured and have ecological significance to our study.
In each year, these six functional traits were measured in 24 common species (Table 1) following the flowering phase. These 24 species accounted for 70–90% of the above-ground biomass, and for each species, we randomly sampled nine fully developed and undamaged leaves from the six subplots. The fresh leaves were weighed before being scanned to measure leaf area using ImageJ software (Schneider et al. 2012); the leaves were then dried at 70 °C for 48 h hand weighed using a Sartorius balance with an accuracy of 10−4 g. We calculated SLA as the ratio of leaf area to dry leaf mass, and LDMC as the ratio of dry leaf mass to fresh leaf mass. We also randomly selected 30 flowering individuals of each species to measure the species maximum height in each treatment. We then clipped the plants, dried them at 70 °C for 48 h, divided them into leaves, stems and flowers, and weighed them using the Sartorius balance. Next, the leaves were used to measure the nitrogen and phosphorus contents. In the laboratory, the leaves were first ground and oven dried at 60 °C for 48 h to a constant weight, and the N and P concentrations in the leaves were then analysed using a continuous flow-injection analyser (SKALAR, Breda, Netherlands). The N and P contents were calculated per unit leaf dry mass.
Data analysis
In the summer of 2011 and 2012, accidental grazing occurred in the experimental plots and disturbed field work; we therefore did not include the data of these 2 years in our analysis. In addition, we did not measure LNC and LPC in 2016 but used the average data (from 2013 to 2015) to replace them, because LNC and LPC were stable from 2012 to 2015.
The CWM trait, which was defined as a trait’s mean value in each species weighted by the species abundance (cover, biomass) in a given community, have been widely used to assess the shifts in functional composition in a plant community (Lepš et al. 2011; de la Riva et al. 2016). The means of height, SLA, LDMC, LNC and LPC were calculated for each species in N fertilization and control plots. The relative abundance of each species in each quadrat was calculated as the biomass of a given species divided by the total biomass of all species. The CWM of each functional trait was calculated for each plot as
where pi is the relative abundance of species i, and traiti is the mean trait value of species i in fertilization or control plots.
We estimated the fertilization response of species relative abundance with the log response ratio. The response of species relative abundance (RSRA) = log (SRAf / SRAc), where SRAf and SRAc are the mean SRA for the quadrats of fertilization and control plots, respectively. Thus, a positive RSRA value means that fertilization increases the relative abundance of a given species (Niu et al. 2010). The response of functional trait (RFT) was also estimated according to this method. Then, the relative abundance of each species between control and fertilization communities was calculated in each plot and tested using a t-test.
A two-way ANOVA was then used to detect the effects of N fertilization, fertilization years (or precipitation), and the interaction between them on CWM traits. Additionally, one-way ANOVA was performed to determine the effects of N fertilization on CWM traits in each year and post hoc comparisons among the different treatments were made using a Tukey’s honest significant difference (HSD) test. Similarly, the effects of fertilization years on CWM traits in both fertilized and control plots were also tested. All variables were tested for the assumption of normality and homogeneity of variance using a Shapiro-Wilk test and Levene’s test, respectively.
Afterwards, to distinguish the relative contribution of ITV versus CCC to variation in each CWM trait, we used a sum of squares decomposition method following Jung et al. (2014) that was originally proposed by Lepš et al. (2011). We calculated the three types of functional composition parameters: 1) specific average traits, using relative abundance and trait values of each species recorded in fertilization plots, whose changes can be caused by both CCC and intraspecific trait variability; 2) fixed average traits, using relative abundance in fertilization plots but where the trait values are measured in a control, whose variation is only due to changes in CCC; and 3) intraspecific variability effect, which is calculated from the differences between specific average traits and fixed average traits: intraspecific variability effect = specific average traits - fixed average traits. Next, the covariation between ITV and CCC was calculated via: covSS = SSspecific- SSfixed- SSintraspec.variability. If the SSfixed are positively correlated with SSintraspec.variability, then the SSspecificwill be higher than when the two effects are independent, thus the covSS will be positive. For instance, after N fertilization, the competition for light is expected to lead to dominance of tall species; meanwhile, individuals who are able grow taller will also be selected. When the two effects are negatively correlated the SSspecific will be lower than expected and the covSS will be negative. Finally, we quantified the relative contribution of ITV, CCC and the covariance between the two by variance decomposition for every functional trait in each fertilization year.
All analyses were performed in R 3.2.0 (R Development Core Team 2013).
Results
The effects of N fertilization on species composition and functional composition
In accordance with previous studies, N fertilization increased the relative abundance of grasses but decreased the relative abundance of forbs in alpine meadow communities, thus significantly changing the community composition (Table 1). There was a shift in the communities from a mixture of forbs (Anemone obtusiloba, Anemone rivularis, Oxytropis kansuensis, Thermopsis lanceolata), grasses (Poa crymophila, Poa pratensis, Elymus nutans), and sedges (Kobresia graminifolia) without N fertilization to a community dominated by two tall grass species (P. pratensis, E. nutans) with N fertilization in our study.
We also found that N fertilization significantly affected the community functional composition (Table 2). N fertilization significantly increased heightCWM and LNCCWM in each fertilization year (Fig. 1a, d). However, the effects of N fertilization on SLACWM, LDMCCWM and LPCCWM were inconsistent. After fertilization, SLACWM was slightly increased in 2013 and 2015, while it was slightly decreased in 2014 and 2016 (Fig. 1c). In contrast, N fertilization slightly decreased LDMCCWM in 2015 but significantly increased it in 2013 and 2016 (Fig. 1b). LPCCWM was significantly increased in 2015 following fertilization (Fig. 1e). The different response patterns of community level functional traits indicate that the mechanisms that influence community functional composition are complicated in fertilization habitats. If annual precipitation was exchanged for years in this analysis, the results showed that height was significantly affected by precipitation (F = 8.3, P < 0.01), rather than fertilization year (F = 1.1, P = 0.303). But the outcome of other four traits was identical to that reported in Table 2 (results not shown).

The effects of nitrogen fertilization on a CWM height, b CWM leaf dry mass content, c CWM specific leaf are, CWM leaf nitrogen content, and d CWM leaf phosphorus content in different years. Asterisks indicate significant differences (Tukey’s test, p < 0.05) for the effect of fertilization; different lowercase letters indicate significant differences (Tukey’s test, p < 0.05) for each year in the control plots; different uppercase letters indicate significant differences (Tukey’s test, p < 0.05) for each year in fertilization plots
Additionally, compared to 2013, heightCWM was significantly lower both in the control and fertilization communities in 2015 (Fig. 1a). Meanwhile, LDMCCWM in 2015 was higher than that in 2013 in fertilization communities (Fig. 1b). In contrast, SLACWM in 2015 was lower than in 2013 in control plots (Fig. 1c). The precipitation for the 2012–2015 interval was 718.6, 532.5, 610.8, and 506 mm, respectively. The precipitation in 2015 declined 17.53% from the mean precipitation over 4 years.
The influence of fertilization duration and trait categories on the relative contribution of ITV versus CCC to functional composition changes
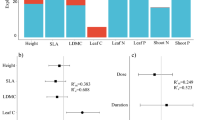
Our results showed that the relative contributions of ITV versus CCC to community functional composition after fertilization depended on fertilization duration and trait categories. For LNC and LPC, ITV always played a more important role than CCC in explaining community level trait variation in all years (Appendix Table 3, Fig. 2d, e). Meanwhile, negative covariations consistently appeared between ITV and CCC in those two traits (Appendix Table 3, Fig. 2d, e). In contrast, for LDMC and SLA, the relative extent of ITV versus CCC in determining community functional composition was dependent on fertilization duration (Appendix Table 3, Fig. 2b, c). In 2013, 3 years after fertilization, ITV explained more variation in functional composition than CCC for all traits. However, since 2014 and 2015, CCC became the dominant factor for LDMC and SLA, respectively. Similarly, for the whole-plant trait of height, ITV also played a more important role than CCC in 2013; however, in the next 3 years, CCC overwhelmed ITV and became dominant in its contribution to community functional composition change (Appendix Table 3, Fig. 2a).

Decomposition of total variation in a CWM height, b CWM leaf dry mass content, c CWM specific leaf are, CWM leaf nitrogen content, and d CWM leaf phosphorus content in each year. More details see Data analysis. White bars represent the contribution of community composition change (CCC), whereas grey bars indicate the contribution of intraspecific trait variation (ITV). Black bars denote positive covariation, whereas black bars and horizontal lines indicate negative covariation
Discussion
The effects of N fertilization on functional composition in an alpine meadow community
Our results displayed that the responsive patterns of community functional composition to N fertilization were trait-specific and different underlying processes may combine and interact to determine those patterns.
In accordance with previous studies (Li et al. 2015; Zhou et al. 2017), N fertilization led to an increase in height and LNC at the community level in our study site. After N fertilization, the intense light competition favours exploitative species with tall stature (e.g., grass) and leads to short species loss via competitive exclusion (Newman 1973; Grime 2001; Hautier et al. 2009; Schellberg and Pontes 2012), thus resulting in tall species becoming dominant and community-level height increasing. Moreover, N fertilization induced the mean height of species to substantially increase relative to the control (Appendix Table 4, Appendix Table 5), which may also enhance heightCWM. Previous studies (Yang et al. 2013) showed that community-level LNC was increased because the LNC of most species increased following the addition of N (Appendix Table 5) due to an enrichment of available N.
In our study, the responses of SLA, LDMC and LPC at the community level were highly inconsistent, and the negative covariation implied that other processes may finally determine those patterns (Fig. 2b, c, e). Generally, SLA and LDMC are two closely related functional traits (Craine et al. 2001), and they showed opposing responses to N fertilization. As many studies have demonstrated (Knops and Reinhart 2000; Schellberg and Pontes 2012), the SLA and LDMC of most species were increased and decreased, respectively, in a habitat of fertilization, but at the same time, fertilization also increased the relative abundance of grasses that had thinner, denser leaves (low SLA, high LDMC) (Weiher et al. 1999). And they may counteract one another and ultimately produce a non-significant variation in most cases. Similarly, two opposing processes may also determine the responses of LPCCWM. As the dilutive effect states, species’ LPC may decrease due to an increase in leaf area and leaf mass that is induced by N fertilization (Vicente-Chandler et al. 1959). On the other hand, LPC may increase because N limitations are alleviated after the addition of N; thus, plants invest more resource in soil P uptake (Ryser and Lambers 1995; Treseder and Vitousek 2001).
Additionally, compared to 2013, heightCWM in 2015 significantly declined both in the control and in fertilization, while LDMCCWM in fertilization was significantly decreased in 2015. These variation patterns may be caused by the drastic decline in precipitation (a decline of 17.53% from the mean precipitation over 4 years) in 2015. These results suggest that community-wide traits could change among different years due to environmental variation.
The relative contribution of ITV versus CCC is trait-specific and affected by fertilization duration
The results of our study showed that all ITV, CCC and covariation between the two must be considered when assessing the variation in functional composition after N fertilization. The relative importance of ITV effects and CCC effects depended on fertilization duration and trait categories.
In agreement with hypothesis i, ITV played a more important role in mediating functional composition during short-term fertilization (<3 year), but the effects of CCC overwhelmed ITV effects and became more important in determining three community-wide traits (height, LDMC and SLA) during long-term fertilization (>3 year). Our results agreed with the hierarchical framework that proposed by Smith et al. (2009) and implied that individual-level responses, such as increasing height and SLA and decreasing LDMC, played an important role in influencing community trait composition after short-term N fertilization. Then, with available nutrients accumulating and light competition intensifying, the relative abundance of exploitative species with tall stature (e.g., grass species, Elymus nutans, Poa pratensis) gradually increased and became dominant in the community (Yang et al. 2011). Meanwhile, the short stature conservative species (e.g., sedge species, Kobresia graminifolia; non-legume forb species, Anemone trullifolia, legume species, Oxytropis kansuensis) would be at a disadvantage in competition and their relative abundance decreased (Niu et al. 2012). Those two processes not only drove the drastic changes in species composition, but also caused CCC effects to overwhelm ITV effects and become the major factor determining community functional composition after long-term fertilization. Moreover, our results showed that more serious relative abundance changes and species loss appeared when CCC effects were dominant in the community after 4 years of N addition (Table 1). This indicated that ITV may play an important role in mediating variation in community species composition and alleviating species loss with short-term fertilization (Siefert and Ritchie 2016).
As expected in hypothesis ii, our results also showed that ITV always played a more important role than CCC in determining functional composition for leaf chemical traits (LNC and LPC). However, CCC was more important than ITV to explaining shifts of whole-plant traits (height) and leaf morphology traits (LDMC and SLA) at the community-level. Previous studies examining ITV and CCC, both in natural and semi-natural communities, also found that the relative extent of ITV versus CCC varies among traits (Lepš et al. 2011; Volf et al. 2016). In line with previous studies (Rozendaal et al. 2006; Kazakou et al. 2014), our results showed that the contributions of ITV were higher in leaf chemical traits than in leaf morphology traits after N fertilization, suggested that leaf chemical traits might be highly labile and sensitive to nutrient additions (Lambers and Poorter 1992). Those patterns also suggested that leaf chemical traits tended to vary, but leaf morphology traits tended to be more conservative when responding to N fertilization in our study. These results might be attributed to the fact that the altering of chemical and physiological traits was quickly and has a lower energetic cost compared to altering of morphology traits (Lambers and Poorter 1992; Schwinning and Sala 2004; Li et al. 2016). In line with the prediction of the optimisation model (Marks 2007), the changes of two community-wide leaf morphology traits (SLA and LDMC) are mainly driven by CCC. But whole-plant traits (height) also showed a similar pattern in our study. A probable reason is that the response of plant height might be primarily determined by their stems, and the stem is a conserved organ even in fertilization habitats.
In the present study, the strengths of community trait variation were also influenced by the covariations between ITV and CCC. Both positive and negative covariations appeared after fertilization, indicating that community-level trait shifts due to ITV and CCC may either reinforce or oppose one another, and this depended on different trait categories. There was positive covariation between ITV and CCC for plant height, suggesting a unidirectional change in plant height due to changes in community composition and within species trait variability (Lepš et al. 2011). In fact, the competition for light is expected to lead to dominance of tall species; meanwhile, individuals who were able to grow taller will also be selected (Poorter and Nagel 2000; Zhou et al. 2016). In contrast, opposing community-level trait shifts due to ITV and CCC were observed in two chemical leaf traits (LNC and LPC). These patterns would be caused by N fertilization which increased the relative abundance of two grasses (Poa crymophila, Poa pratensis) that have low leaf nutrient content, but decreased the relative abundance of a legume species (Oxytropis kansuensis) that have a high leaf nutrient content (Appendix Table 4). The individuals with high leaf nutrient content within grass species were at an advantage in intraspecific competition, since LNC and LPC was closely related to photosynthetic rate and plant reproduction (Wright et al. 2004). However, within legume species, individuals with high leaf nutrients might undergo more serious toxic effects because N fertilization increases leaf manganese (Mn) concentrations and thus led to a decrease in the photosynthetic rate (Bai et al. 2015).
Lastly, we should note that the methods used to measure functional traits in this study may have underestimated the contributions of ITV: Firstly, we only sampled mature individuals and fully developed leaves but ignored the young individuals and leaves in the field. Secondly, in our study, functional traits were only measured at treatment level (control vs. fertilized) but not at plot level, thus the changes in ITV due to differentiation between replicates of plots was ignored. Thirdly, ITV, which includes plastic responses and genetic adaptation, is mainly caused by plastic responses under the short duration of fertilization. CCC is mainly driven by changes in species relative abundance because all species that appeared in this study are common species with high initial abundance; thus, species loss is infrequent.
Conclusion
In this study, we distinguished the relative contribution of ITV and CCC to variation in community functional composition following N fertilization in an alpine meadow using a classical, trait-based approach. We found that all ITV, CCC and the covariation between them should be considered when assessing the variation in functional composition, and the relative importance of ITV effects and CCC effects depended largely on fertilization duration and trait category. To our knowledge, this is the first investigation on the importance of ITV versus CCC in three functional trait categories among different fertilization years in species-rich alpine meadow communities. Our study will be helpful to understand the relative importance of ITV versus CCC in response to environmental change of nutrient-enriched community: ITV plays a critical role in alleviating changes in species composition during short-term fertilization, but the effects of CCC would overwhelm ITV and become more important in determining community functional composition for long-term fertilization.
References
Albert CH, Thuiller W, Yoccoz NG, Soudant A, Boucher F, Saccone P, Lavorel S (2010) Intraspecific functional variability: extent, structure and sources of variation. J Ecol 98:604–613
Auger S, Shipley B (2013) Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest. J Veg Sci 24:419–428
Bai W, Guo D, Tian Q, Liu N, Cheng W, Li L, Zhang WH (2015) Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition. J Ecol 103:1570–1579
Carmona CP, Guerrero I, Morales MB, Oñate JJ, Peco B, Llorens L (2017) Assessing vulnerability of functional diversity to species loss: a case study in Mediterranean agricultural systems. Funct Ecol 31:427–435
Cheplick G (1989) Nutrient availability, dimorphic seed production, and reproductive allocation in the annual grass Amphicarpum purshii. Can J Bot 67:2514–2521
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715
Cornelissen J, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich D, Reich P, Ter Steege H, Morgan H, Van Der Heijden M (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Craine JM, Froehle J, Tilman DG, Wedin DA, Chapin IFS (2001) The relationships among root and leaf traits of 76 grassland species and relative abundance along fertility and disturbance gradients. Oikos 93:274–285
de Bello F, Lavorel S, Albert CH, Thuiller W, Grigulis K, Dolezal J, Janeček Š, Lepš J (2011) Quantifying the relevance of intraspecific trait variability for functional diversity. Methods Ecol Evol 2:163–174
de la Riva EG, Pérez Ramos IM, Tosto A, Navarro Fernández CM, Olmo M, Marañón T, Villar R (2016) Disentangling the relative importance of species occurrence, abundance and intraspecific variability in community assembly: a trait-based approach at the whole-plant level in Mediterranean forests. Oikos 125:354–363
Fajardo A, Piper FI (2011) Intraspecific trait variation and covariation in a widespread tree species (Nothofaguspumilio) in southern Chile. New Phytol 189:259–271
Grime JP (2001) Plant strategies, vegetation processes, and ecosystem properties, 2nd edn. John Wiley and Sons, Chichester 417 p
Hautier Y, Niklaus PA, Hector A (2009) Competition for light causes plant biodiversity loss after eutrophication. Science 324:636–638
Isbell F, Reich PB, Tilman D, Hobbie SE, Polasky S, Binder S (2013) Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc Natl Acad Sci 110:11911–11916
Jung V, Albert CH, Violle C, Kunstler G, Loucougaray G, Spiegelberger T (2014) Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events. J Ecol 102:45–53
Kazakou E, Violle C, Roumet C, Navas ML, Vile D, Kattge J, Garnier E (2014) Are trait-based species rankings consistent across data sets and spatial scales? J Veg Sci 25:235–247
Knops JM, Reinhart K (2000) Specific leaf area along a nitrogen fertilization gradient. Am Midl Nat 144:265–272
Lambers H, Poorter H (1992) Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv Ecol Res 23:187–261
Lefcheck JS, Duffy JE (2015) Multitrophic functional diversity predicts ecosystem functioning in experimental assemblages of estuarine consumers. Ecology 96:2973–2983
Lepš J, de Bello F, Šmilauer P, Doležal J (2011) Community trait response to environment: disentangling species turnover vs intraspecific trait variability effects. Ecography 34:856–863
Li W, Cheng JM, Yu KL, Epstein HE, Guo L, Jing GH, Zhao J, Du GZ (2015) Plant functional diversity can be independent of species diversity: observations based on the impact of 4-yrs of nitrogen and phosphorus additions in an alpine meadow. PLoS One 10:e0136040
Li H, Yu K, Ratajczak Z, Nippert JB, Tondrob D, Xu D, Li W, Du G (2016) When variability outperforms the mean: trait plasticity predicts plant cover and biomass in an alpine wetland. Plant Soil 407:401–415
Luo Y, Qin G, Du G (2006) Importance of assemblage-level thinning: a field experiment in an alpine meadow on the Tibet plateau. J Veg Sci 17:417–424
Marks CO (2007) The causes of variation in tree seedling traits: the roles of environmental selection versus chance. Evolution 61:455–469
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends Ecol Evol 21:178–185
Münzbergová Z, Hadincová V, Skálová H, Vandvik V, Bonser S (2017) Genetic differentiation and plasticity interact along temperature and precipitation gradients to determine plant performance under climate change. J Ecol 105:1358–1373
Newman E (1973) Competition and diversity in herbaceous vegetation. Nature 244:310
Niu K, Zhang S, Zhao B, Du G (2010) Linking grazing response of species abundance to functional traits in the Tibetan alpine meadow. Plant Soil 330:215–223
Niu K, Schmid B, Choler P, Du G (2012) Relationship between reproductive allocation and relative abundance among 32 species of a Tibetan alpine meadow: effects of fertilization and grazing. PLoS One 7:e35448
Niu KC, He JS, Lechowicz MJ (2016) Grazing-induced shifts in community functional composition and soil nutrient availability in Tibetan alpine meadows. J Appl Ecol 53:1554–1564
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, Urcelay C, Veneklaas EJ, Reich PB, Poorter L, Wright IJ, Ray P, Enrico L, Pausas JG, de Vos AC, Buchmann N, Funes G, Quétier F, Hodgson JG, Thompson K, Morgan HD, ter Steege H, Sack L, Blonder B, Poschlod P, Vaieretti MV, Conti G, Staver AC, Aquino S, Cornelissen JHC (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167
Poorter H, Nagel O (2000) The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Funct Plant Biol 27:1191–1191
R Development Core Team (2013) R: A Language and Environment for Statistical Computing. In: R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Riva EG, Pérez Ramos IM, Tosto A, Navarro Fernández CM, Olmo M, Marañón T, Villar R (2015) Disentangling the relative importance of species occurrence, abundance and intraspecific variability in community assembly: a trait-based approach at the whole-plant level in Mediterranean forests. Oikos 125(3):354–363
Rozendaal DMA, Hurtado VH, Poorter L (2006) Plasticity in leaf traits of 38 tropical tree species in response to light: relationships with light demand and adult stature. Funct Ecol 20:207–216
Ryser P, Lambers H (1995) Root and leaf attributes accounting for the performance of fast- and slow-growing grasses at different nutrient supply. Plant Soil 170:251–265
Schellberg J, Pontes LS (2012) Plant functional traits and nutrient gradients on grassland. Grass Forage Sci 67:305–319
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH image to image J: 25 years of image analysis. Nat Methods 9:671–675
Schwinning S, Sala OE (2004) Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 141:211–220
Siefert A, Ritchie ME (2016) Intraspecific trait variation drives functional responses of old-field plant communities to nutrient enrichment. Oecologia 181:245–255
Siefert A, Violle C, Chalmandrier L, Albert CH, Taudiere A, Fajardo A, Aarssen LW, Baraloto C, Carlucci MB, Cianciaruso MV, de LDantas V, de Bello F, Duarte LDS, Fonseca CR, Freschet GT, Gaucherand S, Gross N, Hikosaka K, Jackson B, Jung V, Kamiyama C, Katabuchi M, Kembel SW, Kichenin E, Kraft NJB, Lagerström A, Bagousse Pinguet YL, Li Y, Mason N, Messier J, Nakashizuka T, Overton JM, Peltzer DA, Pérez-Ramos IM, Pillar VD, Prentice HC, Richardson S, Sasaki T, Schamp BS, Schöb C, Shipley B, Sundqvist M, Sykes MT, Vandewalle M, Wardle DA (2015) A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol Lett 18:1406–1419
Smith MD, Knapp AK, Collins SL (2009) A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecology 90:3279–3289
Treseder KK, Vitousek PM (2001) Effects of soil nutrient availability on investment in acquisition of n and p in hawaiian rain forests. Ecology 82:946–954
Vicente-Chandler J, Silva S, Figarella J (1959) The effect of nitrogen fertilization and frequency of cutting on the yield and composition of three tropical grasses. Agron J 51:202–206
Volf M, Redmond C, Albert ÁJ, Bagousse-Pinguet Y, Biella P, Götzenberger L, Hrázský Z, Janeček Š, Klimešová J, Lepš J, Šebelíková L, Vlasatá T, Bello F (2016) Effects of long- and short-term management on the functional structure of meadows through species turnover and intraspecific trait variability. Oecologia 180(4):1–10
Weiher E, van der Werf A, Thompson K, Roderick M, Garnier E, Eriksson O (1999) Challenging Theophrastus: a common core list of plant traits for functional ecology. J Veg Sci 10:609–620
Westoby M, Wright IJ (2006) Land-plant ecology on the basis of functional traits. Trends Ecol Evol 21:261–268
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas M-L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) The worldwide leaf economics spectrum. Nature 428:821–827
Yang Z, Ruijven J, Du G (2011) The effects of long-term fertilization on the temporal stability of alpine meadow communities. Plant Soil 345:315–324
Yang Z, Guo H, Zhang J, Du G (2013) Stochastic and deterministic processes together determine alpine meadow plant community composition on the Tibetan plateau. Oecologia 171:495–504
Zhang P, Zhou X, Li J, Guo Z, Du G (2015) Space resource utilisation: a novel indicator to quantify species competitive ability for light. Sci Rep 5:16832
Zhou X, Wang Y, Zhang P, Guo Z, Chu C, Du G (2016) The effects of fertilization on the trait–abundance relationships in a Tibetan alpine meadow community. J Plant Ecol 9(2):144–152
Zhou X, Guo Z, Zhang P, Li H, Chu C, Li X, Du G (2017) Different categories of biodiversity explain productivity variation after fertilization in a Tibetan alpine meadow community. Ecol Evol 7:3464–3474
Acknowledgements
We would like to thank the staff of the Research Station of Alpine Meadow and Wetland Ecosystems at Lanzhou University for providing invaluable field assistance. The study was supported by the National Nature Science Foundation of China (Grant No.31700355), the Scientific Research Startup Foundation for Doctors of Xinjiang University (Grant No. BS160260).
Author information
Authors and Affiliations
Contributions
XLZ and GZD conceived and designed the experiments. XLZ, PFZ and ZG performed the experiments. XLZ analyzed the data. XLZ wrote the manuscript; other authors provided editorial advice.
Corresponding author
Additional information
Responsible Editor: Hans Lambers
Appendices
Appendix 1
Appendix 2
Appendix 3
Rights and permissions
About this article

Cite this article
Zhou, X., Guo, Z., Zhang, P. et al. Shift in community functional composition following nitrogen fertilization in an alpine meadow through intraspecific trait variation and community composition change. Plant Soil 431, 289–302 (2018). https://doi.org/10.1007/s11104-018-3771-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-3771-x