
Abstract
Aim
The rate of nitrate (NO3 −) uptake and changes in rhizosphere properties were studied growing seedlings of two maize inbred lines differing in nitrogen use efficiency (NUE) in rhizoboxes.
Results
Changes in NO3 − uptake rates occurred in response to anion addition (induction) in seedlings grown both in hydroponic culture and in soil in rhizoboxes. The characterization of root exudate composition showed a line-specific metabolite profile, which was also affected by NO3 − availability. The induction affected respiration, nitrification, ammonification and enzyme activities of the rhizosphere. Furthermore, the composition of rhizosphere bacterial communities of the two maize lines differed suggesting the selective capacity of plants.
Conclusions
Overall, results showed a strong and fast modification of rhizospheric soil properties in response to physiological changes in plants caused by fluctuating NO3 − availability.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The main challenge in plant nutrition and crop production is the improvement of nitrogen (N) use efficiency (NUE) being currently estimated in crops less than 50% (Baligar et al. 2001; Hawkesford et al. 2012). Although the capability of a plant to acquire N from the soil depends on soil type, environment and plant species, physiological and molecular studies concerning the traits related to N acquisition and assimilation are usually carried out in simplified systems with model species (mainly arabidopsis), without considering all the components of the plant-soil-microbial system. Therefore, there is the need to study N nutrition of important crops, such as maize, in systems closer to the real situation like those of soil-grown plants.
The rhizosphere is the soil compartment profoundly modified by the release of root exudates consisting of low molecular weight organic acids, sugars and more complex chemical molecules such as polyphenols and other secondary metabolites (Hawes et al. 2003; Tomasi et al. 2008; Mimmo et al. 2014; Pii et al. 2015). Root exudates enhance plant nutrient uptake (Uren 2007; Tomasi et al. 2008), sustain a larger and more active microbial activity (Renella et al. 2007; Scagliola et al. 2016), and influence the composition of the rhizosphere microbial communities (Brimecombe et al. 2007; Lynch and Whipps 1990; Nannipieri et al. 2008; Pii et al. 2016c). It is accepted that plant species and their stage-growth, as well as soil properties and season, are the main factors influencing the microbial selection in the rhizosphere (Berg and Smalla 2009; Pii et al. 2016b).
In previous work, we reported that the two maize inbred lines with different NUE (Lo5 and T250; Balconi et al. 1997), differed for the composition of their rhizosphere microbial communities and diversity and abundance of functional genes (Baraniya et al. 2016; Pathan et al. 2015a, b; Giagnoni et al. 2016), being the maize line with the highest NUE (Lo5) able to induce the greatest modifications. However, the relation between the plant responses to N availability, particularly in terms of N uptake and root exudate profiles, and the biochemical activity and microbial community composition is still poorly understood.
Nutrient (bio)availability in soil is generally estimated by chemical extractions using H2O or saline solutions (Bremner and Mulvaney 1982). However, bioavailability depends on cell active uptake which can hardly be extrapolated by data on nutrient chemical solubility in the heterogeneous soil matrix (Barber 1995). Whole cell biosensors allow the estimation of the real bioavailable fractions of nutrients and other substances in soils because they report changes in the metabolism of living organisms upon changes in the chemical conditions surrounding microbial cells. Several whole-cell biosensors for detecting the nutrient bioavailability have been constructed with the reporter gene technique (Leveau and Lindow 2002). Among them, the lux-inserted Pseudomonas fluorescens strains, in which bioluminescence is regulated by the cellular levels of FMNH2 (Kragelund et al. 1995, 1997; Yeomans et al. 1999), have been successfully used to monitor nutrient bioavailability in different soils (Kragelund et al. 1995, 1997; Jensen and Nybroe 1999) and nutrient fluxes in the rhizosphere (Kragelund et al. 1997; Yeomans et al. 1999).
In the present study, we hypothesized that Lo5 and T250 maize inbred lines, exhibiting different transcriptome responses when exposed to NO3 − (Zamboni et al. 2014), were characterized by different root exudate profiles; this is deemed to modify the biochemical activity and the composition of the microbial communities in the rhizosphere (Fig. S1). We also hypothesized that the maize line with higher NUE was the strong competitor for inorganic N against soil microorganisms. Therefore, we studied the modifications in biochemical activities and microbial community composition of the rhizosphere of the maize inbred lines Lo5 and T250 grown in rhizoboxes during changes in NO3 − uptake rates following the NO3 − fertilization. The N bioavailability in the rhizosphere was monitored by using whole-cell biosensors and their responses were compared to the roots NO3 − uptake rates. In addition, we analysed the composition of root exudates collected from hydroponically grown plants. To date, this is the first study that analyses comprehensively the changes occurring in the plant-microbe-soil system upon fluctuation of NO3 − availability.
Materials and methods
Plant growth in hydroponic culture and collection of root exudates
Seeds of the Lo5 and T250 inbred lines of Zea mays L. were steeped in water for 24 h and then germinated over aerated deionized water in the dark at 27 °C and 90% relative humidity. After 3 d, seedlings were transferred to aerated 0.5 mM CaSO4 solution in 2.2 L-plastic pots (12 seedlings in each pot) for 1 d and then transferred in a nutrient solution (NS) containing (in mM): KH2PO4 0.175; MgSO4 0.1; KCl 0.005; FeSO4 0.002; H3BO3 0.0025; MnSO4 0.0002; ZnSO4 0.0002; CuSO4 0.00005; H2MoO4 0.00005; CaSO4 0.4 and NH4PO4 0.025. For half of the pots, the NS was integrated with the additional NO3 − 0.2 mM (induced). Plants were grown at a 16/8 h day/night regime (25 and 20 °C, respectively) and at the relative humidity of 70–75%. Since the beginning of this last treatment (growing in NS +/− NO3 −), plant roots were removed from the solutions, rinsed for 3 min in 1 mM CaSO4 and analysed for the NO3 − uptake as specified below. Based on previous results (Locci et al. 2001; Zamboni et al. 2014) and pilot experiments, net NO3 − uptake rate was determined 0, 4 and 8 h after the NO3 − addition for the Lo5 maize line and 0, 12 and 24 h after the NO3 − addition for the T 250 maize line. The whole experiment was independently repeated three times using per each NO3 − uptake assay three biological replicates (three independent intact plants).
Root exudates were collected in plastic Petri plates using plants germinated and grown as described above. In particular, 6 plants were incubated in one plate with 10 ml of the same NS used for hydroponic growth. In N- treated plates, the NO3 − was added to reach a final concentration of 200 μM. Each plate was covered with plastic film in order to avoid evaporation, roots were protected from light and the plates were incubated at 25 °C and at the relative humidity of 70–75%. Each treatment was replicated 5 times (two maize lines × 2 treatments) resulting in 20 plates and treated as true replicates. For each replicate, 1 mL of solution was sampled after 2 and 4 h for Lo5, and 5 and 12 h for T250 maize line.
Soil properties and rhizobox experiments
The rhizobox used in this study had a length of 24 cm, a width of 10.5 cm and a depth of 2.5 cm in the main rooting compartment (inner volume around 650 cm3), with a sliding and transparent front side allowing the visual observation of root development. The rhizoboxes were filled with 820 g of a sandy-clay neutral Eutric Cambisol (81% sand, 6% silt, 13% clay, 0.7% organic C, 0.07% total N, pH 6.0), previously sieved (< 2 mm). Four seedlings were transferred in each rhizobox three d after germination and the soil moisture was kept constant by regular additions of distilled water (every 2 d water consumption was estimated as weight loss of the single rhizobox). Maize plants were grown for 1 month at a 16/8 h day/night regime (25 and 20 °C, respectively) and relative humidity in the range of 70–75%. After one month of plant growth, half of the rhizoboxes were saturated with a 0.5 mM Ca(NO3)2 solution for NO3 − induction, whereas the remaining rhizoboxes were treated with NO3 − free water (control). The rhizoboxes were arranged in a complete randomized design with 18 replicates for each treatment. Based on preliminary hydroponic studies on plant responses to NO3 − induction, the Lo5 maize plants and soil were sampled after 0, 4, and 8 h whereas the T250 maize plants and soil were sampled after 0, 6, and 12 h after the N induction. The whole experiment was independently repeated three times.
For soil analysis, plants were carefully removed, and soil adhering to the root surface was considered as rhizosphere soil, whereas the soil portion not colonized by roots was considered as bulk soil. Rhizosphere soil was gently removed by the root surface by using a sterile spatula, placed into sterile 13-mL tubes and separated into different aliquots: the first aliquot was immediately stored at −20 °C for the analysis of the microbial communities, the second aliquot was pre-incubated at 25 °C for 7 d prior to determination of double-stranded DNA (dsDNA) content and enzyme activities, the third aliquot was air-dried for chemical analyses. The bulk soil was sampled, and split in aliquots and stored in the same way as the rhizosphere soil.
Nitrate uptake and PM H+-ATPase activity
The analysis of NO3 − uptake was performed with three biological replicates using, for the hydroponic experiments intact plants and, for those with rhizoboxes roots, carefully removed from the front side of the rhizoboxes. The experiments were independently repeated three times using plants/tissues of three independent growth.
The assay was carried out transferring the plants in 40 ml of a solution containing 0.2 mM KNO3. The net NO3 − uptake rates were determined by sampling 0.2 mL from the solutions every 2 min for 10 min and measuring the NO3 − depletion as described by Pinton et al. (1999).
Root plasma membrane (PM) vesicles were isolated using roots of seedlings grown in rhizoboxes as described by Tomasi et al. (2009). The analysis was performed with three biological replicates using plants grown in three independent experiments.
Briefly, roots were homogenized within 20 mL of ice-cold extraction buffer containing 250 mM sucrose, 2 mM MgSO4, 2 mM ATP, 10% (v/v) glycerol, 10 mM glycerol-1-phosphate, 0.16% (w/v) bovine serum albumin, 2 mM ethylene glycol tetraacetic acid, 2 mM DTT, 5.7% (w/v) choline-iodide, 1 mM phenylmethylsulfonylfluoride, 20 μg mL−1 chymostatin, 10 nM okadeic acid and 25 mM MES-1,3-bis[tris(hydroxymethyl)-methyloamino] (BTP) at pH 7.6 using a mortar. The homogenates were filtered through four layers of cheesecloth and the suspensions were subjected to differential centrifugation steps at 2 °C: 1500 g for 5 min (pellets discarded); 9800 g for 20 min (pellets discarded), 83,400 g for 30 min (pellets recovered), and a final step at 83400 g for a further 30 min. The microsomes, resuspended in 1.2 mL of ice-cold extraction buffer were loaded onto a discontinuous sucrose gradient made by layering 2 mL of sucrose solution (1.13 g cm−3) onto a 3 mL sucrose (1.17 g cm−3) cushion and centrifuged at 107600 g for 2 h. The sucrose solutions were prepared in 5 mM MES-BTP at pH 7.4 and contained all chemicals of the extraction buffer. The vesicles migrating to the 1.13–1.17 g cm−3 interface were harvested, appropriately diluted with the extraction buffer, and centrifuged at 122400 g for 30 min. The pellets were resuspended in a medium containing 250 mM sucrose, 10% (v/v) glycerol, 1 mM dithiothreitol, 50 μg mL−1 chymostatin and 2 mM MES-BTP at pH 7.0, immediately frozen in liquid nitrogen, and stored at −80 °C until use.
The PM protein content was determined according to Bradford (1976), using BSA as standard, after solubilizing membrane vesicles with 0.5 M NaOH (Gogstad and Krutnes 1982).
The PM H+-ATPase activity was measured at 38 °C in a 0.6 mL reaction buffer [50 mM MES-BTP pH 6.5, 5 mM MgSO4, 100 mM KNO3, 600 μM Na2MoO4, 1.5 mM NaN3, 5 mM ATP-BTP (pH 6.5), 0.01% (w/v) Brij 58 (polyoxyethylene 20 cetyl ether), plus or minus 100 μM V2O5. The vanadate-dependent activity was 89 ± 4%. The reaction was started by adding membrane vesicles containing 0.5 μg of total protein. After 30 min released Pi was determined as described by Forbush (1983).
Analysis of the root exudates
The composition of maize root exudates was analysed by an untargeted metabolomic approach. The solutions containing fresh root exudates were filtered and immediately analysed by HPLC-ESI-MS, using a Beckman Coulter Gold 127 HPLC system (Beckman Coulter, Fullerton, CA) equipped with a C18 guard column (7.5 × 2.1 mm) and an analytical Alltima HP C18 column (150 × 2.1 mm, particle size 3 μm) (Alltech Associates Inc., Derfield, IL). Two solvents were used: 0.5% (v/v) formic acid, 5% (v/v) acetonitrile in water (solvent A), and 100% acetonitrile (solvent B). The solvent alternation of gradient and isocratic phases were established as follows: from 0 to 10% B in 2 min, from 10 to 20% B in 10 min, from 20 to 25% B in 2 min, from 25 to 70% B in 7 min, 70% B isocratic elution for 5 min, from 70 to 90% B in 1 min, 90% B isocratic elution for 4 min. Each sample was analysed in duplicate, using an injection volume of a 100 μL and a flow rate of 200 μL min−1. The HPLC system was coupled on-line with a Bruker ion trap mass spectrometer Esquire 6000, equipped with an electrospray ionization (ESI) source. MS data were collected using the Bruker Daltonics Esquire 5.2- Esquire Control 5.2 software, and processed by the Bruker Daltonics Esquire 5.2-Data Analysis 3.2 software (Bruker Daltonik GmbH, Bremen, Germany). Alternate negative-positive ion mass spectra were recorded in the range 50–3000 m/z (full scan mode, 13,000 m/s). For molecule identification, ms/ms and ms3 spectra were recorded both in a negative and positive mode in the range 50–3000 m/z, with the fragmentation amplitude set at 1 V. Nitrogen was used as the nebulizing and drying gas and helium as the collision gas.
Metabolites were identified by comparing m/z values, fragmentation patterns (ms/ms and ms3) and retention times of each m/z fragment with those of an “in-house” library of spectra of pure standards. When these standards were not available, fragmentation patterns were compared to those available in the literature and in databases, such as MassBank (www.massbank.jp/en/database.html) and Human Metabolome Database (www.hmdb.ca). The extraction and alignment of chromatographic data were carried out using MZmine software (http://mzmine.sourceforge.net), and the resulting data were imported into a Microsoft Excel datasheet for being analysed with SIMCA-P+ version 12.0 (Umetrix AB, Umeå, Sweden). The obtained data matrix was normalized according to the weight of roots releasing the produced exudates and this was done for each signal. Pareto scaling was applied to all analytical methods. The data matrix was initially analysed by principal component analysis (PCA) and then by orthogonal projections to latent structures discriminant analysis (O2PLS-DA) in which the maize lines, the treated or control samples were imposed as Y classes. The resulting O2PLS-DA models were cross-validated using a permutation test (200 permutations). To determine which metabolites contributed to the class separation, we used the pq(corr) parameter, i.e. the correlation between p (based on the X component of the model, the metabolites) and q (based on the Y component of the model, the sample classes). The significance of each variable was also determined by univariate analysis, using the t-test.
Nutrient bioavailability in rhizosphere and bulk soil
Nutrient bioavailability in rhizosphere and bulk soil was determined by the bacterial biosensor consisting of Pseudomonas fluorescens strains inserted with the promoterless lux CDABE from Vibrio fischeri. The N-reporter biosensor was the Pseudomonas fluorescens DF57 N3 strain, the C-reporter biosensor was the Pseudomonas fluorescens 10,586 pUCD607 strain, and the P-reporter biosensor was the Pseudomonas fluorescens DF57 P9 strain. The bioluminescence of these bacterial biosensors was regulated by intracellular levels of FMNH2, was maximum under N, C and P starvation and was proportionally reduced during nutrient assimilation (Kragelund et al. 1995, 1997). All bacterial biosensors were harvested at the late exponential growth phase in Luria Bertani plus kanamycin broth and then led into an N, C and P starvation phase in a mineral N- C- and P-free minimal medium (DMM, Difco) containing kanamycin for 2 h at 25 °C. The soils were moistened to 80% of the water holding capacity using sterile distilled water and, then, centrifuged at 4000 x g at 2 °C for 20 min with a final collection of soil pore water. One-hundred mL of soil pore water were put in contact with 900 mL of bacterial biosensor suspension and then 5 μL of n-decylaldheide were immediately added. The bioluminescence was measured by a Sirius luminometer (Titertek Berthold, Pforzheim, Germany) 15 min after the injection of n-decylaldehyde into the solution. The bioluminescence response was calibrated using sterile solutions of NH4NO3 (0.01–10 mM) for bioavailable N, glucose (0.001–1 mM) for bioavailable C and NaH2PO4 (0.001–1 mM) for bioavailable P.
Soil microbial biomass, respiration, enzyme activities and nitrification and ammonification potential
The soil microbial biomass was estimated by the content of the double-stranded DNA (dsDNA) according to Marstop and Witter (1999). The soil respiration was measured by placing 100 g (DW) of soil in 100 mL air-tight glass flasks provided with 3-way valves and incubated at 25 °C in the dark for 4 h. The evolution of CO2-C was measured by sampling the head-space and injecting the gas samples into a gas-chromatograph (Hewlett-Packard 6890) equipped with a packed column (Porapack Q) and a thermal conductivity detector (Blackmer and Bremner 1977). Empty flasks served as blanks accounting for the CO2-C background concentration. The acid and alkaline phosphomonoesterase activities were assayed according to Tabatabai and Bremner (1969) and the β-glucosidase activity according to Tabatabai (1982). The concentration of p-nitrophenol (p-NP) released by these hydrolase activities was determined at 400 nm by spectrophotometry (Lambda 2, PerkinElmer, Waltham, MA, USA). The urease activity was determined according to Nannipieri et al. (1974) and the protease activity by using N-benzoyl-argininamide (BAA) as a substrate, according to Ladd and Butler (1972). The NH4 + released by the urease and protease assays was extracted with 1 M KCl and quantified at 660 nm after reaction with the Nessler reagent. The efficiency of NH4 + recovery was evaluated by standard additions of NH4 + to soil slurries at concentrations in the range of those released by urease and protease activities and varied from 95% to 102% for all soil fractions. The concentrations of NH4 + released by urease and protease activities were determined by a flow injection analyser (FIAS 5000, Foss-Tecator, Fisher Scientific, Waltham, MA, USA) associated to a spectrophotometer (Lambda 2, PerkinElmer, Waltham, MA, USA).
The nitrification potential was determined by measuring the NO2 − concentration after soil incubation with 2 mM of (NH4)2SO4 for 4 h and stopping the NO2 − oxidation to NO3 − by adding 4 mM NaClO3, according to Sauvé et al. (1999). The quantification of NO2 − production allowed determining nitrification activity in soils treated with Ca(NO3)2. The soil ammonification potential was determined by incubating soils with a 0.2% of arginine solution for 3 h at 25 °C (Alef and Kleiner 1986). The NO2 − and NH4 + produced by the nitrification and ammonification activities, respectively, were extracted with 1 M KCl and quantified colorimetrically by the flow injection technique described above. The NH4 +, NO2 − and NO3 − produced by the measurements of nitrification and ammonification potential were determined by a flow injection technique described above.
Analysis of the composition of soil bacterial communities by 454 pyrosequencing
This analysis was carried out by 454 pyrosequencing at the Research Unit Environmental Genetics at the Helmholtz Zentrum München. The DNA of rhizosphere and bulk soil of both maize cultivars grown in rhizoboxes sampled before the NO3 − treatment was extracted by the phenol-chloroform-extraction protocol according to Griffiths et al. (2000), modified by Töwe et al. (2011). The extracted DNA was amplified by PCR using 50 ng of template DNA, quantified by a Quant-iT™ Pico Green® dsDNA assay Kit (Invitrogen, Carlsbad, USA). The amplification solution contained the 1× Fast start HiFi Polymerase Buffer (Roche, Basel, Switzerland), 0.05 U μL−1 Fast start HiFi Polymerase (Roche, Basel, Switzerland), 0.2 mM of dNTP, 0.2 μM of each fusion primer and 3% BSA. The PCR conditions were: 22 cycles of 1 min denaturation at 94 °C, 1 min annealing at 50 °C and 1 min elongation at 72 °C, preceded by an initial denaturation step of 5 min at 95 °C, and terminated by a final elongation step of 10 min at 72 °C. The bacterial 16S rRNA gene primer 926f (5′-AAACTYAAAKGAATTGACGG-3′) (Lane et al. 1985) or 630R (5′-CAKAAAGGAGGTGATCC-3′) (Juretschko et al. 1998) were extended with a 4-base key and a specific 10-base barcode, which was different for each soil sample allowing bidirectional sequencing, according to www.my454.com by a 21-base 454-Adaptor (adaptor A or adaptor B). The PCR products were purified with AMPure Beads (Agencourt, Beckman Coulter, Krefeld, Germany) according to the Amplicon Library Preparation Method Manual (www.my454.com) and quantified on DNA7500 chips (Agilent Technologies, Santa Clara, CA, USA). All PCR products had a peak size of approximately 730 bp.
The equimolar amounts of PCR Products were pooled, and the amplicon pools were amplified by emulsion PCR according to the Titanium protocol (454 Life Sciences, Roche Diagnostics and loaded onto a GS FLX Titanium PicoTiterPlate). The 16S rRNA amplicons were run on a second-generation pyrosequencer (454 GS FLX Titanium, Roche, Basel, Switzerland). The raw sequences were filtered using the automatic amplicon pipeline of the 454 GS Run Processor supplied by Roche with an adjusted filter, removing failed reads, adaptor sequences and low-quality reads. The raw reads of the twelve sequenced amplicon libraries were passed through the respective automatic pipeline of the 454 GS Run Processor.
The MOTHUR programme (Schloss et al. 2011) was used to remove chimeras and other unwanted DNA sequences (e.g. from mitochondria, archaea, chloroplasts and unknowns), thereby generating high-quality reads. The sequences were clustered into operational taxonomic units (OTUs) at 95% and 97% similarity.
We identified OTUs responsible for observed differences using samr with the false discovery rate cut-off of 0.15.
The similarity among microbial community groups was generated by the Newick-formatted trees and clustered using the UPGMA algorithm. The distance between communities showed that there were significant differences between the rhizosphere and bulk soil for both the maize lines. The microbial communities of the rhizosphere of the Lo5 and T250 maize line were not significantly different, either using the forward or reverse datasets.
Results
Nitrate uptake and PM H+ −ATPase activity in root of the Lo5 and T250 maize lines grown in hydroponic solution and in rhizobox.
For a long time, it has been demonstrated by experiments run in hydroponic solutions, that plant roots exposed to NO3 − increase the rate of the anion uptake (induction; Hawkesford et al. 2012). The Lo5 line showed a much faster response to the NO3 − exposure than the T250 line, after growth either in hydroponic solution or in rhizobox (Fig. 1a and b) respectively, while that of T250 happened after 12 h in both conditions. At the peaks, the rates of NO3 − uptake were similar in the two maize lines in both conditions (Fig. 1a and b). Concerning the rhizobox experiment, the NO3 − uptake rate profile paralleled the root PM H+-ATPase activity as already shown for maize plants grown in hydroponics (Fig. 1c).

Rate of net NO3 − uptake in roots of Lo5 and T250 plants grown in hydroponic culture (a), and in rhizobox (b), and PM H+-ATPase activity of vesicles isolated from roots of plants grown in rhizobox (c). Symbols –N and +N indicate Control and NO3 − -treated plants. Data are the means ± s.e.; n = 3
Root exudate profiles of the Lo5 and T250 maize lines
The characterization of root exudates was carried out for both genotypes on plants grown in hydroponics without (control) or after NO3 − exposure. The collection times (2 and 4, 5 and 12 h for Lo5 and T250, respectively) were chosen on the bases of the different behavior of NO3 − uptake rate shown by the two lines (Fig. 1a). The chromatographic profiles of root exudates (controls) of both lines were complex (Fig. S2). The first part of the chromatograms, especially that of the Lo5 line, was dominated by low molecular weight metabolites including N organic molecules as indicated by even m/z values. Some of the N compounds were putatively identified as benzoxazinones and benzoaxazinone glucosides (Tables S1 and S3). In the less hydrophilic part of chromatograms, some signals were putatively identified as hydroxylated fatty acids (oxylipins), as free hydroxycinnamic acids (coumaric and ferulic acid) and coumaroyl tyramine (Table S1). To obtain a more comprehensive picture of differences between the metabolites of root exudates released by the two maize lines independently from time and treatment (e.g all root exudate samples), we analysed the raw chromatographic data using the M/Zmine software, and the resulting data matrix was investigated by multivariate analyses (PCA and O2PLS-DA). The analysis showed broad differences between the two genotypes (Fig. 2a). By considering the arbitrary threshold of pq(corr) (i.e. the relationship between metabolites, the X variables, and the maize lines Lo5 or T250, Y variables) values of 0.6, 45 m/z fragments were detected in root exudates of Lo5, and 203 in T250, respectively (Table S2). However, when data were analysed by univariate analyses (t-test) 38 and 214 additional m/z features were detected for Lo5 and T250, respectively (Table S2). The Lo5-specific metabolites included most of the detected benzoxazinones while the T250 typical m/z features were all unidentified except for ferulic acid.

PCA score plot (a) and O2PLS-DA S-loading plot (b) produced using all exudate samples of controls and NO3 − treated plants of both maize line (Lo5 controls sampled at 2 h, Lo5 NO3 − treated sampled at 2 h, Lo5 controls sampled at 4 h, Lo5 NO3 − treated sampled at 4 h, T250 controls sampled at 5 h, T250 NO3 − treated sampled at 5 h, T250 controls sampled at 12 h, T250 NO3 − treated sampled at 12 h; three replicates for each sample). c O2PLS-DA S-loading plot of Lo5 maize line (Lo5 controls sampled at 2 h, Lo5 NO3 − treated sampled at 2 h, Lo5 controls sampled at 4 h, Lo5 NO3 − treated sampled at 4 h; three replicates for each sample). d O2PLS-DA S-loading plot of T250 maize (T250 controls sampled at 5 h, T250 NO3 − treated sampled at 5 h, T250 controls sampled at 12 h, T250 NO3 − treated sampled at 12 h; three replicates for each sample)
The O2PLS-DA models were also applied to identify metabolite modifications occurring in response to NO3 − treatments irrespective of the time of sampling. Both NO3 − treated maize lines were characterized by a decreased level for several metabolites as compared to controls (Table S2, Fig. 2c and d).
Only a few of these metabolites could be putatively identified: the benzoaxazinone (TRIBOA) and its glucoside derivative in root exudates from the Lo5 line, and the hydroxylated fatty acid 18:1-3OH and the benzoaxazinone (HM2BOA) in root exudates from T250. Only in root exudates of the latter line, few metabolites increased their concentration in response to NO3 − treatment but they could not be identified.
Nutrient bioavailability in rhizosphere and bulk soil
A large increase in the N availability was detected by bacterial biosensors in the rhizosphere and bulk soil of both Lo5 and T250 maize lines immediately after the NO3 − addition to soil as compared to the not amended soils (Fig. 3a). A considerable N depletion in the rhizosphere, up to levels comparable to those of the not NO3 − -amended, soils was observed after 4 and 12 h for the Lo5 and T250 maize lines, respectively, whereas no such a depletion was observed in the respective bulk soils (Fig. 3a). A significant increase in bioavailable C concentration was observed in the rhizosphere of both Lo5 and T250 maize lines during the NO3 − induction phase (Fig. 3d), whereas a significant increase of P bioavailability was observed in the rhizosphere of the Lo5 and T250 maize lines after 8 and 6–12 h since the beginning of the induction phase, respectively (Fig. 3e). No significant changes in the C and P bioavailability were observed in the rhizosphere of the non-induced plants and in the bulk soils of both induced and non-induced plants (Fig. 3d and e).

Bioavailability of total N (a), NO3 − (b), exchangeable NH4 + (c), C (d) and P (e) in the rhizosphere and bulk soils of the Lo5 and T250 maize lines grown in rhizobox during the NO3 − induction period. Symbols R, B indicate the rhizosphere and bulk soil, respectively, with (+N) or without (−N) the addition of NO3 −. Data are the means ± s.e.; n = 3. Statistical significance was evaluated by one-way ANOVA followed by Tukey’s test using the GraphPad InStat Program (version 5.0). Different letters denote significant differences at P ≤ 0.05
Soil microbial biomass, respiration, ammonification and nitrification potentials during the NO3 − induction phase.
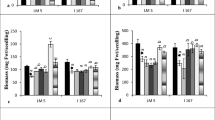
The dsDNA content of rhizosphere and bulk soil was not significantly changed by the NO3 − induction in both Lo5 and T250 maize lines (Fig. 4a), whereas the soil respiration rate was significantly enhanced in the rhizosphere of induced Lo5 and T250 maize lines after 8 and 24 h of induction, respectively (Fig. 4b). The nitrification potential rates were significantly higher in the rhizosphere of induced Lo5 and T250 maize lines than those of the rhizosphere of not induced plants at all times (Fig. 4c). Nevertheless, the ammonification potential was significantly higher in the rhizosphere of induced Lo5 and T250 lines after 8 and 12–24 h since the induction, respectively (Fig. 4d). An increase in the ammonification potential was also observed in the NO3 − amended bulk soil of both Lo5 and T250 maize lines, but it was only significant in the bulk soil of the Lo5 line at the end of the induction period (Fig. 4d).

Double stranded DNA content (a), basal respiration (b), nitrification potential (c) and ammonification potential (d) of the rhizosphere and bulk soil of Lo5 and T250 maize lines grown in rhizobox during the NO3 − induction period. Symbols R, B indicate the rhizosphere and bulk soil, respectively, with (+N) or without (−N) the addition of NO3 −. Data are the means ± s.e.; n = 3. Statistical significance was evaluated by one-way ANOVA followed by Tukey’s test using the GraphPad InStat Program (version 5.0). Different letters denote significant differences at P ≤ 0.05
The NO3 − addition to soil significantly increased the acid and alkaline phosphomonoesterase (Fig. 5a and b) and the urease (Fig. 5d) activities of the rhizosphere of the NO3 − induced plants as compared to the rhizosphere of the not induced plants and of the bulk soils of both the NO3 − amended and not amended soils. The protease activity (Fig. 5e) was significantly higher in the rhizosphere soil of the NO3 − induced Lo5 maize line but not in the rhizosphere soil of the NO3 − induced T250 maize line. The β-glucosidase activity did not change regardless of the rhizosphere and bulk soil, maize line and NO3 − induction (Fig. 5c).

Enzyme activities of the rhizosphere and bulk soil of Lo5 and T250 plants grown rhizobox during the NO3 − induction period. Alkaline phosphomonoesterase (a); acid phosphomonoesterase (b); ß-glucosidase (c); urease (d); protease (e) activities. Symbols R, B indicate the rhizosphere and bulk soil, respectively, with (+N) or without (−N) the addition of NO3 −. Data are the means ± s.e.; n = 3. Statistical significance was evaluated by one-way ANOVA followed by Tukey’s test using the GraphPad InStat Program (version 5.0). Different letters denote significant differences at P ≤ 0.05
Microbial community composition of rhizosphere and bulk soil
The analysis of microbial community composition was carried out on rhizosphere and bulk soils sampled before the NO3 − treatment for both maize lines. The dataset generated by using the forward primer, resulted after quality filtering and removal of chloroplast sequences in 80.289 high-quality reads, with an average read length of 521 bp. For statistical reasons, the reads per sample were subsampled to 5400 for all samples. The rarefaction curves indicated a good coverage of the total bacterial diversity at OTU97 for rhizosphere samples, whereas for bulk soil the plateau has not be reached (data not shown).
The calculation of indicated α-diversity based on the number of obtained OTUs (OTU97) indicated like expected an overall higher diversity in the bulk soil samples compared to rhizosphere. If in bulk soil on average more than 300 different OTUs were detected, in the rhizosphere samples the OTU richness was reduced almost by 50%. Interestingly α-diversity in the rhizosphere of T250 was significantly lower (130 OTUs) compared to Lo5 (240 OTUs; Fig. 6). Significant differences in α-diversity in the bulk soil samples from the two maize lines were not detected, due to the high variability found in samples from Lo5.

a α-Diversity of bacterial communities in the rhizosphere (R) and bulk soil (B) of the two maize lines Lo5 and T250 based on the number of obtained OTUs (similarity level 97%) after direct DNA extraction from the samples and partial amplification of the 16S rRNA genes followed by next generation sequencing. Data are the means ± s.e.; n = 3. b Abundance of microbial communities in the rhizosphere (R) and bulk soil (B) of the two maize lines Lo5 and T250 based on the number of obtained OTUs (similarity level 97%) after direct DNA extraction from the samples and partial amplification of the 16S rRNA genes followed by next generation sequencing. Data are the means ± s.e.; n = 3
The different composition of bacterial communities between the two different maize lines was probably supported when shared OTUs between rhizosphere samples between both lines were calculated. From the 96 OTUs found in all samples from Lo5 rhizosphere, only 52 were also detected in T250 rhizosphere.
The most abundant OTUs in all samples could be linked to Proteobacteria, ranging from 58 to 72%. As expected in both rhizosphere samples significant higher relative abundance values were found compared to bulk soil. The second most dominant group of OTUs were characterized as Firmicutes, which were more frequently found in samples from bulk soil. Interestingly differences were found in the abundance of Firmicutes when the rhizosphere samples from both maize lines were compared, with higher abundances for Firmicutes in samples obtained from T250. In contrast, OTUs related to Planctomycetes, Gemmatimonadetes and Chloroflexi were more abundant in rhizosphere samples from Lo5, whereat the share of Chloroflexi was below 5% of all reads obtained (Fig. 6b). Overall the obtained results did not differ when the reads obtained from PCR products using the reverse primer were analyzed (data not shown).
Discussion
Characterization of responses to NO3 − availability performed on the two maize inbred lines hydroponically grown confirmed the previous observations on the faster induction of the high-NUE maize line Lo5 than T250 (Locci et al. 2001; Zamboni et al. 2014). The results showed that this behavior was also maintained by plants grown in soil, with the shorter time for the NO3 − uptake peak in the Lo5 maize inbred line (Fig. 1b). Furthermore, the rate of NO3 − uptake of the plant grown in not NO3 −-fertilized rhizoboxes did not show significant differences throughout the experimental time (Fig. 1b). Nitrate uptake rates also coincided with the PM H+-ATPase activity, confirming the previous results by Pii et al. (2016a). Confirmation of results obtained in hydroponics for NO3 − uptake by the maize lines with those grown in soil is noteworthy, because it demonstrates that soil factors playing important roles in plant nutrition such as nutrient concentrations, soil structure, water retention, abundance, activity, and composition of fungal and bacterial communities in the rhizosphere (Badalucco and Nannipieri 2007), may not substantially limit the plant genetic response to NO3 − availability.
Nitrogen bioavailability determined by whole-cell biosensors indicated that the bioavailable N was significantly depleted in the rhizosphere during the induction phase, whereas no significant depletion in rhizosphere of not induced plants and in bulk soil was observed (Fig. 3a, b and c). Therefore, even with different timing, both the Lo5 and T250 maize lines were proven to be both the stronger competitors than bacteria for the bioavailable N during the NO3 − induction phase. In fact, the N bioavailability in the rhizosphere of induced plants paralleled the NO3 − uptake and PM H+-ATPase activity for both maize lines, since the timing of N microbial starvation and N plant uptake is a mirror image of each other (Fig. 1b and c). The stronger competitive capability of microorganisms as compared to plants roots on the short term has been mainly attributed to their higher affinity for N. This was demonstrated by the studies on the N mobilization/immobilization turnover (MIT) of showing that the N immobilization in soil after N addition was very fast (Barraclough 1997), and by the calculation of Michaelis–Menten kinetics of NO3 −, NH4 +, and amino acids uptake by roots and microorganisms (Kuzyakov and Xu 2013). The latter study also demonstrated that although microorganisms are stronger competitors shortly after soil N amendment, plants are stronger competitors in N acquisition on the long term mainly due to the mass flow from soil to plant roots. Our results of N bioavailability in the rhizosphere resembled the lower NO3 − bioavailability in the wild oat rhizosphere observed by DeAngelis et al. (2005) using bacterial biosensors. Both nitrification and ammonification activities were increased in the rhizosphere of the Lo5 and T250 maize lines (Fig. 4c and d). However, Giagnoni et al. (2016) showed a decrease in both exchangeable NH4 + and NO3 − concentrations in the rhizosphere soils of the two maize lines with time: interestingly both concentrations were generally lower in the rhizosphere of the Lo5 than T250 maize line. A steep gradient of decrease of nitrification potential from the rhizosphere to bulk soil has been shown (Li et al. 2007). Generally, root exudation of carboxylates, mainly malate and citrate, occurs by increasing NO3 − concentrations in the culture medium (Neumann and Romheld 2007). The observed increase in nitrification activity in the rhizosphere paralleled that observed by Watson et al. (1994) after addition of Ca-ammonium nitrate to soil. It cannot be excluded that the increase in nitrification activity of the rhizosphere soils might have been triggered by signaling molecules released by the plant in the rhizosphere or changes in the composition of root exudates with a decrease in the concentration of compounds inhibiting nitrification (Subbarao et al. 2012). In fact, compounds such as brachialactone, isolated from root exudates of the pasture grass Brachiaria humidicola, and methyl 3-(4- hydroxyphenyl) propionate (MHPP), sorgoleone and sakuranetin exuded from sorghum (Sorghum bicolor) have proven to act as biological nitrification inhibitors in the rhizosphere (Subbarao et al. 2012). The increase in the ammonification potential in the rhizosphere of the Lo5 and T250 maize lines promoted by the NO3 − induction phase (Fig. 4d) may also depend on the high availability of NH4 + produced by the net microbial N mineralization from soil organic N (Fig. 3c). The observed increase of enzyme activities in the rhizosphere soil represents an indirect support to this hypothesis (Fig. 5).
The C and P bioavailability were increased in the rhizosphere soil during the induction phase (Fig. 3d and e); in particular, the C bioavailability showed an increasing trend during the whole plant induction phase for both the Lo5 and T250 maize lines (Fig. 3d), and this trend partially paralleled with the soil respiration rates (Fig. 4b). The nutrient bioavailability determined by whole-cell biosensors provides an estimation of the nutrient pool that can cross the cell membranes; it should be interpreted as a flux of nutrient that is capable of inducing the genetic response of biosensors (Werlen et al. 2004; Harms et al. 2006), including the eventual nutrient resupply into the soil solution from different chemical pools after plant uptake (van der van der Meer et al. 2004). Interestingly, the biosensor response to the N availability was the mirror image of NO3 − response of the two different maize lines, indicating microbial N starvation as compared to the activity of plant NO3 − and NH4 + uptake.
The analysis of the root exudate profiles of the Lo5 and T250 plants before and after the NO3 − induction showed that only MBOA and its glucoside derivative could be identified univocally because of the lack of the relative standards for the other unidentified compounds (Tables S1 and S3). Therefore, a complete investigation of the root exudate profiles is needed. The MBOA and some glucoside derivatives, as well as benzoaxazinone have been detected in maize (Cambier et al. 1999; Jonczyk et al. 2008; Marti et al. 2013; Meihls et al. 2013). Moreover, free hydroxycinnamic acids, such as coumaric and ferulic acid and derivatives such coumaroyl tyramine, and less hydrophilic compounds, identified as oxylipin, were also detected in maize by Marti et al. (2013). These compounds are intermediates and end-chain products in the hydroxycinnamic acid and tyramine amide biosynthesis pathways (Negrel and Javelle 1997) and have a main role in defence against pathogens, allelopathic interactions and cell wall reinforcement (Bassard et al. 2010). The benzoxazinones are non-toxic 2-O-β-D-glucosides compounds detected in several plant species including maize, rye and wheat and can be transformed into toxic aglycones by enzymatic or chemical hydrolysis; toxic effects of aglycones occurs through irreversible alkylation of relevant biological nucleophile metabolites (Dixon et al. 2012). We mainly found aglycones instead of glucosides, which are generally present as root exudates (Marti et al. 2013), probably due to chemical hydrolysis, as supported by the detection of HMPA, a malonamic acid, which is a degradation product of MBOA and HMBOA (Villagrasa et al. 2008). Glucosides and the relative intermediates can have different locations in maize plants, while the DIMBOA indolic precursors are synthesized in the plastids and the successive steps in the synthesis occur in the endoplasmic reticulum; the glucosides of DIBOA and TRIBOA are released in the cytosol and then stored in vacuoles (Jonczyk et al. 2008). However, the responses of these secondary metabolites to N availability, their role in the root exudates and their effects on the activity and composition of microbial communities of the rhizosphere soil should be clarified by future research. From the results of root exudation together with the nutrient bioavailability and the biochemical activity in the rhizosphere of the two maize lines, it can be hypothesized that the release of C could cause N limiting conditions of the rhizosphere, promoting the immobilization of NH4 +, and followed by the observed increase in the ammonification rate (Fig. 4d). Future research by using either 15N enriched NH4 + or 15N enriched NO3 − for determining gross rates of N immobilization, N mineralization and nitrification by the 15N dilution technique is required to verify the validity of the above-proposed hypotheses. The increase in C bioavailability in the rhizosphere may explain the lack of the increase in the β-glucosidase activity since this enzyme activity is inhibited in soil when sufficient amounts of low molecular weight organic C are available (Renella et al. 2007). Differently, the P bioavailability showed a peak after 4 and 12 h for the Lo5 and T250 maize lines, respectively, and this may be related to increases in the acid and alkaline phosphomonoesterase activities (Fig. 5a and b), which hydrolyse organic P to inorganic P, which is taken up by plants.
The analysis of bacterial diversity suggests that plants may select the microbial communities in the rhizosphere (Fig. 6a), as already reported by Berg and Smalla (2009). The rhizosphere of the Lo5 maize line had higher species richness than that of the T250 maize line and such differences may depend on the different exudation profiles of the two maize lines. The distribution of the identified bacterial groups in soils of the two maize lines confirms what already reported (Pathan et al. 2015a; Giagnoni et al. 2016) that bacterial communities of the bulk soils were dominated by Proteobacteria, Firmicutes, Planctomycetes and Acidobacteria, and the rhizosphere soils resulted enriched in Proteobacteria. Interestingly the OUT differing between the rhizosphere of both samples was related to Geobacter, which has been mainly identified in wetland soils, and has plant growth promoting properties due to its capability to reduce oxidized Mn4+ to Mn2+, which is the Mn metabolically active form (Lovley et al. 2004). The relatively high abundance of OTUs belonging to Firmicutes could be related to the reduced oxygen content in the rhizosphere of Lo5 maize line. These OTUs were mostly linked to Clostridia, which includes several N-fixing bacteria and thus the presence of this genus may be important in supporting plant growth by releasing available N. Overall, these results confirm that Proteobacteria dominate the rhizosphere bacterial communities whereas Firmicutes diversity is reduced in the maize rhizosphere (Chelius and Triplett 2001; Peiffer et al. 2013). The decrease of line-specific benzoxazinones in exudates, known as antimicrobial compounds, during the NO3 − induction, suggests a direct effect of some root exudates on specific microbial groups. This is a novel evidence of responses of microbial communities to changes induced by roots responding to NO3 − availability in soil. Future studies are needed to get insights on the relationship between specific root exudates and specific bacterial groups in the rhizosphere microbial communities, and the relationship between plant NUE and the composition of the rhizosphere microbial communities (Rice and Tiedje 1989).
Conclusions
The Lo5 maize line with higher NUE retained its ability to increase faster the NO3 − uptake rate in comparison to T250 when grown in soil. Analysis of N bioavailability showed that the two maize lines were stronger competitors for NO3 − than soil microorganisms, inducing microbial N starvation. The studied maize lines responded to the increased NO3 − availability, probably by modifying their root exudate profiles. Although only a few metabolites were identified, we can suggest that changes in the root exudate profiles trigged a strong microbial response in terms of respiration, nitrification, ammonification, and enzyme activities, particularly in the rhizosphere of the high NUE Lo5 maize line. The composition of rhizosphere bacterial communities of the two maize lines differed, suggesting the selective capacity of plants. Further studies are needed to better elucidate the role of the identified root exudates in selecting the bacterial populations in the maize rhizosphere, and in affecting the nutrient turnover and the maize NUE.
References
Alef K, Kleiner D (1986) Arginine ammonification, a simple method to estimate microbial activity potentials in soils. Soil Biol Biochem 18:233–235. https://doi.org/10.1016/0038-0717(86)90033-7
Badalucco L, Nannipieri P (2007) Nutrient transformations in the rhizosphere. In: Pinton R, Varanini Z, Nannipieri P (eds) The rhizosphere: biochemistry and organic substances at the soil-plant interface. CRC Press, Boca Raton, pp 111–133
Balconi C, Brosio D, Motto M (1997) Analysis of nitrogen partitioning in maize. Maize Genet Coop News Lett 71:10–11
Baligar VC, Fageria NK, He ZL (2001) Nutrient use efficiency in plants. Commun Soil Sci Plant Anal 32:921–950. https://doi.org/10.1081/CSS-100104098
Baraniya D, Puglisi E, Ceccherini MT, Pietramellara G, Giagnoni L, Arenella M, Nannipieri P, Renella G (2016) Protease encoding microbial communities and protease activity of the rhizosphere and bulk soils of two maize lines with different N uptake efficiency. Soil Biol Biochem 96:176–179. https://doi.org/10.1016/j.soilbio.2016.02.001
Barber SA (1995) Soil nutrient bioavailability: a mechanistic approach, 2nd edn. John Wiley, New York
Barraclough D (1997) The direct or MIT route for nitrogen immobilization: a 15N mirror image study with leucine and glycine. Soil Biol Biochem 29:101–108
Bassard J-E, Ullmann P, Bernier F, Werck-Reichhart D (2010) Phenolamides: bridging polyamines to the phenolic metabolism. Phytochemistry 71:1808–1824. https://doi.org/10.1016/j.phytochem.2010.08.003
Berg G, Smalla K (2009) Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol 68:1–13. https://doi.org/10.1111/j.1574-6941.2009.00654.x
Blackmer AM, Bremner JM (1977) Gas chromatographic analysis of soil atmosphere. Soil Sci Soc Am J 41:908–912
Bradford MM (1976) Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein- dye binding. Anal Biochem 72:248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Bremner JM, Mulvaney CS (1982) Nitrogen-Total. In: Al P, Miller RH, Keeney DR (eds) Methods of soil analysis, part 2. Academic Press, New York, pp 595–624. https://doi.org/10.2134/agronmonogr9.2.2ed.c31
Brimecombe MJ, FAAM DL, Lynch JM (2007) Rhizodeposition and microbial populations. In: Pinton R, Varanini Z, Nannipieri P (eds) The rhizosphere: biochemistry and organic substances at the soil-plant interface. CRC Press, Boca Raton, pp 73–109
Cambier V, Hance T, de Hoffmann E (1999) Non-injured maizecontains several 1,4-benzoxazin-3one related compounds but only as glucoconjugates. Phytochem Anal 10:119–126. https://doi.org/10.1002/(SICI)1099-1565(199905/06)10:3
Chelius MK, Triplett EW (2001) The diversity of archaea and bacteria in association with the roots of Zea mays L. Microb Ecol 41:252–263. https://doi.org/10.1007/s002480000087
DeAngelis KM, Ji P, Firestone MK, Lindow SE (2005) Two novel bacterial biosensors for detection of nitrate availability in the rhizosphere. Appl Environ Microbiol 71:8537–8547. https://doi.org/10.1128/AEM.71.12.8537-8547.2005
Dixon DP, Sellars JD, Kenwright AM, Steel PG (2012) The maize benzoxazinone DIMBOA reacts with glutathione and other thiols to form spirocyclic adducts. Phytochemistry 77:171–178. https://doi.org/10.1016/j.phytochem.2012.01.019
Forbush B (1983) Assay of Na, K-ATPase in plasma-membrane preparations: increasing the permeability of membrane-vesicles using sodium dodecyl-sulfate buffered with bovine serum- albumin. Anal Biochem 128:159–163. https://doi.org/10.1016/0003-2697(83)90356-1
Giagnoni L, Pastorelli R, Mocali S, Arenella M, Nannipieri P, Renella G (2016) Availability of different nitrogen forms changes the microbial communities and enzyme activities in the rhizosphere of maize lines with different nitrogen use efficiency. Appl Soil Ecol 98:30–38. https://doi.org/10.1016/j.apsoil.2015.09.004
Gogstad GO, Krutnes MB (1982) Measurement of protein in cell-suspensions using the coomassie brilliant blue dye-binding assay. Anal Biochem 126:355–359. https://doi.org/10.1016/0003-2697(82)90527-9
Griffiths RI, Whiteley AS, O’Donnell AG, Bailey MJ (2000) Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA- and rRNA-based microbial community composition. Appl Environ Microbiol 66:5488–5491. https://doi.org/10.1128/AEM.66.12.5488-5491.2000
Harms H, Wells MC, van der Meer JR (2006) Whole-cell living biosensors–are they ready for environmental application? Appl Microbiol Biotechnol 70:273–280. https://doi.org/10.1007/s00253-006-0319-4
Hawes MC, Bengough G, Cassab G, Ponce G (2003) Root caps and rhizosphere. J Plant Growth Regul 21:352–367. https://doi.org/10.1007/s00344-002-0035-y
Hawkesford M, Horst W, Kichey T, Lambers H, Schjoerring J, Skrumsager Møller I, White P (2012) Functions of macronutrients. In: Marschner P (ed) Marschner’s mineral nutrition of higher plants, 3rd edn. Academic Press, London, pp 135–189
Jensen LE, Nybroe O (1999) Nitrogen availability to Pseudomonas fluorescens DF57 is limited during decomposition of barley straw in bulk soil and in the barley rhizosphere. Appl Environ Microbiol 65:4320–4328
Jonczyk R, Schmidt H, Osterrieder A, Fiesselmann A, Schullehner K, Haslbeck M, Sicker D, Hofmann D, Yalpani N, Simmons C, Frey M, Gierl A (2008) Elucidation of the final reactions of DIMBOA-glucoside biosynthesis in maize: characterization of Bx6 and Bx7. Plant Physiol 146:1053–1063. https://doi.org/10.1104/pp.107.111237
Juretschko S, Timmermann G, Schmid M, Schleifer KH, Pommerening-Röser A, Koops H-P, Wagner M (1998) Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge: Nitrosococcus mobilis and Nitrospira-like bacteria as dominant populations. Appl Environ Microbiol 64:3042–3051
Kragelund L, Christoffersen B, Nybroe O, de Bruijn FJ (1995) Isolation of lux reporter gene fusions in Pseudomonas fluorescens DF57 inducible by nitrogen or phosphorus starvation. FEMS Microbiol Ecol 17:95–106. https://doi.org/10.1111/j.1574-6941.1995.tb00134.x
Kragelund L, Hosbond C, Nybroe O (1997) Distribution of metabolic activity and phosphate starvation response of lux-tagged Pseudomonas fluorescens reporter bacteria in the barley rhizosphere. Appl Environ Microbiol 63:4920–4928
Kuzyakov Y, Xu X (2013) Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytol 198:656–669
Ladd JN, Butler JH (1972) Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol Biochem 4:19–30. https://doi.org/10.1016/0038-0717(72)90038-7
Lane DJ, Pace B, Olsen GJ, Stahl DA, Sogin ML, Pace NR (1985) Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc Natl Acad Sci U S A 82:6955–6959. https://doi.org/10.1073/pnas.82.20.6955
Leveau JH, Lindow SE (2002) Bioreporters in microbial ecology. Curr Opin Microbiol 5:259–265. https://doi.org/10.1016/S1369-5274(02)00321-1
Li YL, Zhang YL, Hu J, Shen QR (2007) Contribution of nitrification happened in rhizospheric soil growing with different rice cultivars to N nutrition. Biol Fertil Soils 43:417–425. https://doi.org/10.1007/s00374-006-0119-0
Locci G, Santi S, Monte R, Pinton R, Varanini Z (2001) Involvement of plasma membrane H+-ATPase in nitrate uptake by maize genotypes. In: Horst WJ, Schenk MK, Bürkert A, Classsen N, Flessa H, Frommer WB, Goldbach H, Olfs H-W, Römheld V, Sattelmacher B, Schmidhalter U, Schubert S, von Virén N, Wittenmayer L (eds) Plant nutrition-food security and sustainability of agro-ecosystems through basic and applied research. Kluwer Academic Publishers, Netherlands, pp 184–185. https://doi.org/10.1007/0-306-47624-X_88
Lovley DR, Holmes DE, Nevin KP (2004) Dissimilatory Fe(III) and Mn(IV) reduction. Adv Microb Physiol 49:219–286. https://doi.org/10.1016/S0065-2911(04)49005-5
Lynch JM, Whipps JM (1990) Substrate flow in the rhizosphere. Plant Soil 129:1–10. https://doi.org/10.1007/BF00011685
Marstop H, Witter E (1999) Extractable dsDNA and product formation as measures of microbial growth in soil upon substrate addition. Soil Biol Biochem 31:1443–1453. https://doi.org/10.1016/S0038-0717(99)00065-6
Marti G, Erb M, Boccard J, Glauser G, Doyen GR, Villard N, Robeet CAM, Turlings TCJ, Rudaz S, Wolfender J-L (2013) Metabolomics reveals herbivore-induced metabolites of resistance and susceptibility in maize leaves and roots. Plant Cell Environ 36:621–639. https://doi.org/10.1111/pce.12002
Meihls LN, Handrick V, Glauser G, Barbier H, Kaur H, Haribal MM, Lipka AE, Gershenzon J, Buckler ES, Erb M, Köllner TG, Jander G (2013) Natural variation in maize aphid resistance is associated with DIMBOA-Glc methyltransferase activity. Plant Cell 25:2341–2355. https://doi.org/10.1105/tpc.113.112409
Mimmo T, Del Buono D, Terzano R, Tomasi N, Vigani G, Crecchio C, Pinton R, Zocchi G, Cesco S (2014) Rhizospheric organic compounds in the soil–microorganism–plant system: their role in iron availability. Eur J Soil Sci 65:629–642. https://doi.org/10.1111/ejss.12158
Nannipieri P, Ceccanti B, Cervelli S, Sequi P (1974) Use of 0.1 M pyrophosphate to extract urease for podzol. Soil Biol Biochem 6:359–362. https://doi.org/10.1016/0038-0717(74)90044-3
Nannipieri P, Ascher J, Ceccherini MT, Guerri G, Renella G, Pietramellara G (2008) Recent advances in functional genomics and proteomics of plant associated microbes. In: Nautiyal CS, Dion P (eds) Molecular mechanisms of plant and microbe coexistence. Springer, Heidelberg, pp 215–241. https://doi.org/10.1007/978-3-540-75575-3
Negrel J, Javelle F (1997) Purification, characterization and partial amino acid sequencing of hydroxycinnamoyl-CoA:tyramine N-(hydroxycinnamoyl) transferase from tobacco cell-suspension cultures. Eur J Biochem 247:1127–1135. https://doi.org/10.1111/j.1432-1033.1997.01127.x
Neumann G, Romheld V (2007) The release of root exudates as affected by the plant physiological status. In: Pinton R, Varanini Z, Nannipieri P (eds) The rhizosphere: biochemistry and organic substances at the soil-plant interface. CRC Press, Boca Raton, pp 23–72. https://doi.org/10.1201/9781420005585.ch2
Pathan S, Ceccherini MT, Pietramellara G, Puschenreiter M, Giagnoni L, Arenella M, Varanini Z, Nannpieri P, Renella G (2015a) Enzyme activity and microbial community structure in the rhizosphere of two maize lines differing in N use efficiency. Plant Soil 387:413–424. https://doi.org/10.1007/s11104-014-2306-3
Pathan SI, Ceccherini MT, Hansen MA, Giagnoni L, Ascher J, Arenella M, Sørensen SJ, Pietramellara G, Nannipieri P, Renella G (2015b) Maize lines with different nitrogen use efficiency select bacterial communities with different β-glucosidase-encoding genes and glucosidase activity in the rhizosphere. Biol Fertil Soils 51:995–1004. https://doi.org/10.1007/s00374-015-1045-9
Peiffer JA, Spor A, Koren O, Jin Z, Green Tringed S, Dangle JL, Buckler ES, Ley RE (2013) Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc Natl Acad Sci USA 110:6548–6553. https://doi.org/10.1073/pnas.1302837110
Pii Y, Mimmo T, Tomasi N, Terzano R, Cesco S, Crecchio C (2015) Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition processes. A review. Biol Fertil Soils 51:403–415. https://doi.org/10.1007/s00374-015-0996-1
Pii Y, Alessandrini M, Dall’Osto L, Guardini K, Prinsi B, Espen L, Zamboni A, Varanini Z (2016a) Time-resolved investigation of molecular components involved in the induction of NO3 − high affinity transport system in maize roots. Front Plant Sci 7:1657. https://doi.org/10.3389/fpls.2016.01657
Pii Y, Borrusso L, Brusetti L, Cesco S, Mimmo T (2016b) How do plants-having different exudation patterns-shape a similar microbial community? Res Rev J Bot Sci 5:61–64
Pii Y, Borruso L, Brusetti L, Crecchio C, Cesco S, Mimmo T (2016c) The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome. Plant Physiol Biochem 99:39–48. https://doi.org/10.1016/j.plaphy.2015.12.002
Pinton R, Cesco S, Iacolettig G, Astolfi S, Varanini Z (1999) Modulation of NO3 −-uptake by water-extractable humic substances: involvement of root plasma membrane H+ ATPase. Plant Soil 215:155–161. https://doi.org/10.1023/A:1004752531903
Renella G, Landi L, Valori F, Nannipieri P (2007) Microbial hydrolase activity after release of low molecular weight organic compounds by a model root surface in a clayey and a sandy soil. Appl Soil Ecol 36:124–129. https://doi.org/10.1016/j.apsoil.2007.01.001
Rice CW, Tiedje JM (1989) Regulation of nitrate assimilation by ammonium in soils and in isolated soil microorganisms. Soil Biol Biochem 21:597–602. https://doi.org/10.1016/0038-0717(89)90135-1
Sauvé S, Dumestre A, McBride M, Gillett JW, Berthelin J, Hendershot W (1999) Nitrification potential in field-collected soils contaminated with Pb or Cu. Appl Soil Ecol 12:29–39. https://doi.org/10.1016/S0929-1393(98)00166-8
Scagliola M, Pii Y, Mimmo T, Cesco S, Ricciuti P, Crecchio C (2016) Characterization of plant growth promoting traits of bacterial isolates from the rhizosphere of barley (Hordeum vulgare L.) and tomato (Solanum lycopersicon L.) grown under Fe sufficiency and deficiency. Plant Physiol Biochem 107:187–196. https://doi.org/10.1016/j.plaphy.2016.06.002
Schloss PD, Gevers D, Westcott SL (2011) Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS One 6:e27310. https://doi.org/10.1371/journal.pone.0027310
Subbarao GV, Sahrawat KL, Nakahara K, Ishikawa T, Kishii M, Rao IM, Hash CT, George TS, Srinivasa Rao P, Nardi P, Bonnett D, Berry W, Suenaga K, Lata JC (2012) Chapter six- biological nitrification inhibition - a novel strategy to regulate nitrification in agricultural systems. Adv Agron 114:249–302. https://doi.org/10.1016/B978-0-12-394275-3.00001-8
Tabatabai MA (1982) Soil enzymes. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis, part 2. Academic Press, New York, pp 903–947
Tabatabai MA, Bremner JM (1969) Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol Biochem 1:301–307. https://doi.org/10.1016/0038-0717(69)90012-1
Tomasi N, Weisskopf L, Renella G, Landi L, Pinton R, Varanini Z, Nannipieri P, Torrent J, Martinoia E, Cesco S (2008) Flavonoids of white lupin roots participate in phosphorus mobilization from soil. Soil Biol Biochem 40:1971–1974. https://doi.org/10.1016/j.soilbio.2008.02.017
Tomasi N, Kretzschmar T, Espen L, Weisskopf L, Fuglsang AT, Palmgren MG, Neumann G, Varanini Z, Pinton R, Martinoia E, Cesco S (2009) Plasma membrane H-ATPase-dependent citrate exudation from cluster roots of phosphate-deficient white lupin. Plant Cell Environ 32:465–475. https://doi.org/10.1111/j.1365-3040.2009.01938.x
Töwe S, Wallisch S, Bannert A, Fischer D, Hai B, Haesler F, Kleineidam K, Schloter M (2011) Improved protocol for the simultaneous extraction and column-based separation of DNA and RNA from different soils. J Microbiol Methods 84:406–412. https://doi.org/10.1016/j.mimet.2010.12.028
Uren NC (2007) Types, amounts and possible functions of compounds released into the rhizosphere by soil-grown plants. In: Pinton R, Varanini Z, Nannipieri P (eds) The rhizosphere: biochemistry and organic substances at the soil-plant interface. CRC Press, Boca Raton, pp 1–21. https://doi.org/10.1201/9781420005585.ch1
van der Meer JR, Tropel D, Jaspers M (2004) Illuminating the detection chain of bacterial bioreporters. Environ Microbiol 6:1005–1020. https://doi.org/10.1111/j.1462-2920.2004.00655.x
Villagrasa M, Guillamón M, Navarro A, Eljarrat E, Barceló D (2008) Development of a pressurized liquid extraction-solid-phase extraction followed by liquid chromatography-electrospray ionization tandem mass spectrometry method for the quantitative determination of benzoxazolinones and their degradation products in agricultural soil. J Chromatogr A 1179:190–197. https://doi.org/10.1016/j.chroma.2007.11.087
Watson CJ, Kilpatrick DJ, Cooper JE (1994) The effect of increasing application rate of granular calcium ammonium nitrate on net nitrification in a laboratory study of grassland soils. Fert Res 40:155–161. https://doi.org/10.1007/BF00750101
Werlen C, Jaspers MCM, van der Meer JR (2004) Measurement of biologically available naphthalene in gas and aqueous phases by use of a Pseudomonas putida biosensor. Appl Environ Microbiol 70:43–51. https://doi.org/10.1128/AEM.70.1.43-51.2004
Yeomans CV, Porteous F, Paterson E, Meharg AA, Killham K (1999) Assessment of lux-marked Pseudomonas fluorescens for reporting on organic carbon compounds. FEMS Microbiol Lett 176:79–83. https://doi.org/10.1111/j.1574-6968.1999.tb13645.x
Zamboni A, Astolfi S, Zuchi S, Pii Y, Guardini K, Tononi P, Varanini Z (2014) Nitrate induction triggers different transcriptional changes in a high and a low nitrogen use efficiency maize inbred line. J Integr Plant Biol 56:1080–1094. https://doi.org/10.1111/jipb.12214
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Philip John White.
Electronic supplementary material
ESM 1
(PDF 4694 kb)
ESM 2
Chromatographic profiles of root exudates collected by roots of seedling not treated with NO3- (control) of Lo5 and T250 lines. (PDF 39 kb)
ESM 3
ID, m/z (-) value (in negative ionization mode) m/z (+) value (in positive ionization mode), retention time (rt), putative identification of signal. (XLSX 14 kb)
ESM 4
Row ID, Row m/z (in negative ionization mode), retention time, putative identification, p1 and pq(corr)1 and p-value of t-test of multivariate and t-test analysis performed of each comparisons or samples. (XLSX 631 kb)
ESM 5
ID, m/z (-) value (in negative ionization mode), m/z (+) value (in positive ionization mode), retention time (rt), putative identification, ms/ms value and ms3 value for each fragment. I %= percentage of intensity of the signal compared to the higher signal (=100%). (PDF 162 kb)
Rights and permissions
About this article

Cite this article
Varanini, Z., Cesco, S., Tomasi, N. et al. Nitrate induction and physiological responses of two maize lines differing in nitrogen use efficiency: effects on N availability, microbial diversity and enzyme activity in the rhizosphere. Plant Soil 422, 331–347 (2018). https://doi.org/10.1007/s11104-017-3452-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-017-3452-1