
Abstract
Aims
The objectives of this study were to evaluate (1) the fertilizer potential of bone char, (2) the effects of wood biochar on plant-available phosphorus (P), and (3) the role of root-mycorrhizae-biochar interactions in plant P acquisition from a P-fixing soil.
Methods
Incubation and pot experiments were conducted with a P-fixing soil and maize with or without root hairs and arbuscular mycorrhizae (AM) inoculation. Olsen-, resin-P and plant P accumulation were used to estimate P availability from bone char, co-pyrolyzed bone char-wood biochar, and separate bone char and wood biochar additions produced at 60, 350 and 750 °C, and Triple Superphosphate (TSP).
Results
Maize inoculated with AM showed similar P accumulation when fertilized with either 750 °C bone char or TSP. Pyrolyzing bone did not increase extractable P in soil in comparison to unpyrolyzed bone, apart from a 67 % increase in resin-extractable P after additions of bone char pyrolyzed at 350 °C. Despite greater Olsen-P extractability, co-pyrolysis of bone with wood reduced maize P uptake. Wood biochars reduced resin-P from bone char by 14–26 %, whereas oven-dried wood increased resin-P by 23 %.
Conclusions
Bone char is an effective P fertilizer, especially if root-AM interactions are simultaneously considered. Biochar influences plant access to soil P and requires careful management to improve P availability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Managing soil phosphorus (P) availability is essential to sustain agricultural production worldwide. Phosphorus adsorption to mineral oxides results in soil fertility management challenges, especially in highly weathered acid soils (Sanchez 1976; Marschner 1995; Haynes and Mokolobate 2001; Hinsinger 2001; Raghothama and Karthikeyan 2005). In addition, rock phosphate mining for fertilizer is rapidly depleting global P reserves (Abelson 1999; Smil 2000; Gilbert 2009). Hence, there is an urgent need for developing alternative P fertilizers and sustainable management practices that optimize P availability in agro-ecosystems (Cordell et al. 2009, 2011).
Converting slaughterhouse waste into P fertilizer through pyrolysis is one way to recycle P to agricultural systems (Vassilev et al. 2013). Not only would this renewable fertilizer be a more economic option (Buss et al. 2016), bone chars also contain fewer heavy metals and are more effective in sorbing cadmium from contaminated soils than conventional fertilizer (Siebers and Leinweber 2013). Animal bones mainly consist of biological apatite, a relatively crystalline calcium phosphate structure (Wopenka and Pasteris 2003, 2005). In contrast to pyrolyzed plant materials or biochar, pyrolyzed bone contains low amounts of organic carbon. Therefore, it is not referred to as biochar but bone char. While bone char as sustainable P fertilizer has received more scientific attention over the past few years, little remains known about the effect of pyrolysis conditions on the P fertilizer efficacy of bone char. Moreover, the biological and chemical mechanisms that control plant P availability from bone char in soils are poorly understood.
Previous studies found that bone char produced at 400 °C was a more effective P fertilizer than GAFSA rock phosphate (Warren et al. 2009). Research focusing on the effect of production conditions on availability of bone char P showed that pyrolysis temperature decreases water-extractable P but increases formic acid-extractable and presumably plant-available P (Zwetsloot et al. 2015). Yet it is unclear how these results translate to actual P uptake of plants. In fact, greenhouse experiments demonstrated that bone char application to different soils and crops led to both increases and decreases in plant biomass and P concentration (Siebers et al. 2012, 2014). These findings warrant further investigation of the availability of bone char P resulting from different production conditions in order to compare bone char to conventional P fertilizers such as Triple Super Phosphate (TSP) fertilizer.
Besides testing the P fertilizer efficacy of different bone chars, managing organic matter inputs and root-mycorrhizae interactions could optimize P availability from bone char. In comparison to other nutrients, P has low mobility in soils. Especially in P-fixing soils commonly found in tropical farming systems (Sanchez 1976), plant-available P from fertilizer can be drastically reduced through its interactions with mineral oxides that chemisorb phosphate from the soil solution (Parfitt et al. 1975). Organic matter additions to P-fixing soils can improve the availability of added P by decreasing P adsorption and increasing P desorption (Singh and Jones 1976; Guppy et al. 2005). The same has been observed for biochar additions to ferrihydrite minerals (Cui et al. 2011). The presence of biochar in Ferralsol and Anthrosol soils increased total plant P uptake when P fertilizer was added (Lehmann et al. 2003). Yet other studies found that biochar only decreased P adsorption in a Ferralsol when compost was added simultaneously (Qayyam et al. 2015). It is unclear what biochar production parameters and plant growing conditions may explain these different results.
Recent research has highlighted the importance of root foraging to plant P nutrition (Ramaekers et al. 2010; Richardson et al. 2011). Increased root hair elongation, top soil lateral branching, high root:shoot ratio, root exudation, increased root P uptake kinetics and mycorrhizal symbiosis are all found to be strategies that favor P acquisition (Lynch and Beebe 1995; Bates and Lynch 2001; Vance et al. 2003; Hodge et al. 2009; Ramaekers et al. 2010; Zhu et al. 2010; Lynch 2011). Yet few studies have simultaneously examined the effect of soil amendments and nutrient acquisition strategies on plant P uptake. Especially in the case of P fertilizers with low water-solubility such as bone char, managing the foraging capacity of plants and mycorrhizal associations may significantly improve plant P acquisition. Yet we are not aware of studies examining the combined effects of plant rooting or mycorrhizae and P uptake from bone char.
Therefore, the objectives of this study were: (1) to test the potential of bone char produced at different pyrolysis temperatures as a P fertilizer in comparison to TSP; (2) to examine the effect of co-pyrolyzed and separate maple wood biochar additions on P availability from bone char in a P-fixing soil; and (3) to evaluate the role of AM and root foraging strategies in P acquisition from bone char and biochar applications.
Materials and methods
Soil characteristics
Soil was collected from the Jimma University agricultural research farm (Jimma, Ethiopia). Mehlich-III extractable P was below our detection limit of 0.01 mg P kg−1 soil. Soil pH was 4.9 in water and 4.0 in 1 M KCL. The cation exchange capacity was 279 mmolc kg−1 soil, and it contained exchangeable base cations at levels of 5.2 g Ca kg−1 soil, 2.9 g K kg−1 soil, 1.2 g Mg kg−1 soil and 0.1 g Na kg−1 soil. Total C was 31 g kg−1 and total N 3.0 g kg−1. The soil showed a substantial maximum P sorption capacity of 456 mg P kg−1 soil (Langmuir isotherm with k = 0.171, R2 = 0.98, n = 9). Soil texture was 50 % clay, 40 % silt and 10 % sand as determined by hydrometer analysis. The clay fraction mainly contained kaolinites with traces of illite, chlorite and quartz as shown by X-ray diffraction (Online Supplementary Fig. S1).
Phosphorus fertilizers and biochars
Rendered bone meal was purchased from The Espoma Company 1929 (Milville, NJ, USA). Robinson Lumber in Owego, NY, USA supplied hard wood chips (80 % red maple, 20 % sugar maple). Wood chips were oven-dried at 60 °C and ground to a particle size <2 mm with a Thomas Wiley Mill (Thomas Scientific, Swedesboro, NJ, USA). Bone meal was pyrolyzed at a heating rate of 2.5 °C min−1 and temperature was maintained at either 350 or 750 °C for 45 min in a muffle furnace swept with argon gas. Bone meal mixed with hard wood chips at a dry weight ratio of 1:1 were pyrolyzed under the same conditions. Greenkeeper’s Secret Triple Superphosphate (TSP) fertilizer (T&N, Inc., Foristell, MO, USA) supplied soluble P in the fertilizer controls. The pH, formic- and water-extractable P, total elemental P, Ca, Mg, K, Na, S, Fe and Al (Zwetsloot et al. 2015), and total elemental C, N, O and H by combustion using a Thermo Delta V Advantage Isotope Ratio Mass Spectrometer and a Temperature Conversion Elemental Analyzer (Thermo Scientific, West Palm Beach, FL were measured to characterize the bone chars, biochars and fertilizers (Table 1).
Abiotic incubation design
An abiotic incubation experiment compared 12 different P additions to P-fixing soils in an incomplete factorial design, by varying feedstock for pyrolysis, P source-biochar mixtures and pyrolysis temperatures: (a) bone meal; (b) bone meal pyrolyzed at 350 °C or 750 °C; (c) mixtures of separately oven-dried (60 °C) and pyrolyzed (at 350 °C or 750 °C) bone meal and maple wood; (d) co-pyrolyzed (at 350 °C or 750 °C) bone meal and wood; (e) TSP alone; and (f) TSP mixed separately to soil with 350 °C or 750 °C wood biochar (Supplementary Online Table S1). These were compared to unamended controls and to added wood that was either oven-dried at 60 °C or pyrolyzed at either 350 °C or 750 °C. Pyrolyzed materials were ground with a mortar and pestle to a particle size of 74–150 μm. Bone meal and dried wood chips were ground with a Thomas Wiley Mill (Thomas Scientific, Swedesboro, NJ, USA) to mesh size 60.
The treatments were applied in five replicates to 250-mL mason jars containing 5 g of soil sieved to <2 mm. Deionized water was added to saturation (4 mL jar−1). To create abiotic conditions for P sorption, HgCl2 was dissolved in the deionized water prior to application at a rate of 500 mg HgCl2 kg−1 soil (Tuominen et al. 1994). Bone and TSP treatments were applied at a rate of 360 mg total P kg−1 soil, which was based on 80 % of the maximum sorption capacity of soil. The separate maple wood biochar additions were equal to the amount of biochar applied in the co-pyrolyzed bone char-wood biochar treatments amounting to a wood application rate of 4.2 g kg−1, 1.2 g kg−1 and 0.9 g kg−1 soil for 60 °C, 350 °C and 750 °C treatments respectively. Total P content of wood biochars was less than 0.3 mg P g−1 (Table 1), confirming that the wood biochars by themselves were not a significant P nutrient source. All jars were put on a rotary shaker at 25 rpm at 30 °C for 5 weeks.
Abiotic incubation analyses
Total contents of the jars (5 g) were used for analysis. Olsen-P and resin-P extractions were used to measure plant-available P since more common extraction methods for low pH soils such as Mehlich and Bray would have overestimated P from the calcium phosphate chemical structures of bone char (Kuo 1996). In the case of Olsen-P, 100 mL of 0.5 M NaHCO3 adjusted to pH 8.5 with 1 M NaOH solution was added to the samples and put on a horizontal shaker at 100 rpm for 30 min (Kuo 1996). After filtering with both a 2 V Whatman qualitative filter paper and 0.45 μm filter paper, the filtrates received 3.25 mL 12 M HCl to lower the pH for colorimetric P determination (Murphy and Riley 1962).
For the resin-P extraction (Tiessen and Moir 1993), anion-exchange resin (AER) strips (Ionac MA-7500, LANXESS Sybron, Birmingham, NJ, USA) were cut to 20 by 60 mm strips, rinsed four times with deionized water, soaked in 0.5 M NaHCO3 (25 strips L−1) overnight, and rinsed with deionized water. One AER strip together with 150 mL deionized water was added to each jar and placed on a rotary shaker at 100 rpm for 24 h. AER strips were removed from the jars and placed in centrifuge tubes containing 40 mL 0.5 M HCl. After shaking the centrifuge tubes at 200 rpm for 20 h, AER strips were taken out and 0.5 M HCL extracts were colorimetrically analyzed for P (Murphy and Riley 1962). Olsen-P and resin-P extracts from the TSP treatments were analyzed by inductively coupled plasma atomic emission spectrometry (ICP-AES Thermo Jarrel Ash 166 Trace Analyzer, Thermo Jarrell Ash Corporation, Franklin, MA, USA).
Pot trial design and management
The pot trial followed a completely randomized block design. Phosphorus source additions were sieved to <2 mm and included TSP fertilizer, bone char, and bone char-wood biochar combination pyrolyzed at 750 °C in comparison to a zero-P control. Because previous studies showed higher formic-extractable and therefore presumably higher plant-available P contents from bone char with an increase in pyrolysis temperature (Zwetsloot et al. 2015), bone char produced at 750 °C was used for this trial. To examine the interaction of plant’s P foraging ability with these P sources, we both increased and decreased the P uptake efficiency of the single maize variety B73 (Zea mays L.). Phosphorus foraging capacity was decreased by using a B73 mutant with no root hairs (−RH) and increased by inoculating unmodified B73 with AM (+RH + AM). The unmodified B73 variety was also planted (+RH). B73 maize seeds were supplied by the USDA Genetic Resource Information Network (Beltsville, MD, USA). The root hairless rth1–1 mutant (115 A) seeds (Wen and Schnable 1994, Hochholdinger et al. 2008) were obtained from the Maize Genetics Coop Stock Center (Urbana, IL, USA). Glomus clarum (strain WV325, INVAM, West Virginia University, USA) was used for AM inoculation. To ensure higher P foraging ability of treatments inoculated with G. clarum, we measured AM colonization of maize roots with trypan blue staining and microscopic analysis (Koske and Gemma 1989).
To stagger plant harvests, the five replicates of the experiment were planted on successive days and placed in a 2.40 m by 1.35 m growth chamber at the Guterman Bioclimatic Laboratory and Greenhouse Complex at Cornell University, Ithaca, NY, USA. Model CP512 tree pots (volume = 5 L, height = 0.305 m, width = 0.127 m) were used (Stuewe and Sons, Tangent, OR, USA). Pots were filled with 4 L of soil (dry weight = 3713 g) with a control or G. clarum inoculum in the middle 1250 mL layer at a volume ratio of 1:25. Inoculant carrier material was Agsorb attapulgite (Oildri, Chicago, IL, USA) with dried roots and spores from a low-P sorghum-sudangrass pot culture. As a control, carrier material without inoculant was added to treatments without AM. TSP fertilizer, bone char and co-pyrolyzed bone char-wood biochar were added at the same total P application rate of 100 kg P ha−1, which equaled to 43.4 mg P kg−1 soil or 161.3 mg P pot−1. The abiotic incubation used a very high P addition rate in order to study chemical mechanisms controlling plant-available P. We lowered the application rate for the pot trial to be more in line with field conditions. The wood biochar application in the bone char-wood biochar treatment amounted to 0.01 % by dry soil weight or 0.4 g biochar pot−1. The total P content of 750 °C wood biochar was 0.28 mg P g−1 biochar (Table 1). Hence, additional P supply through biochar totaled 0.1 mg pot−1 and was ruled out as relevant P source for maize. Three B73 maize seeds were placed 50 mm above the middle layer. Three days after germination, the two weakest seedlings were removed. Treatments without root hairs only received one B73 115 A seed due to limited seed availability.
Pots were watered by weight to 50 % water-filled pore space. Every other day plants were watered with 100 mL of nutrient solution without P: 1.5 mM Ca(NO3)2, 5 mM NH4NO3, 3 mM KNO3, 2 mM (NH4)2SO4, 2 mM MgSO4, 0.024 mM Fe-EDTA, 0.05 mM KCL, 0.025 mM H3BO4, 0.007 mM MnSO4, 0.006 mM ZnSO4, 0.002 mM CuSO4 and 0.0005 mM (NH4)6Mo7O24. On day 10, all pots received an additional 100 mL of 15 mM K2SO4.
Pot trial analyses
At harvest, 5 weeks after seeding, shoots were cut and pots were sliced vertically into two halves. One half was carefully washed with water and sampled for intact crown roots that were stained (0.167 g L−1 neutral red dye) and used for morphological analyses. The remaining roots were washed with water. The shoots and roots were dried at 60 °C for 4 days and weighed. Dried biomass was ground with a Thomas Wiley Mill mesh size 60 (Thomas Scientific, Swedesboro, NJ, USA).
To determine plant P concentration and total P uptake, shoot and root biomass were digested in a mixture of HNO3 and 30 % H2O2 (Benton Jones 2001). Digests were analyzed by inductively coupled plasma atomic emission spectrometry (ICP-AES Thermo Jarrell Ash 166 Trace Analyzer, Thermo Jarrell Ash Corporation, Franklin, MA).
Four replicates were used for root morphological analysis with WinRHIZO Pro 2007d (Regent Instruments Inc., Québec, Canada). Red stained crown roots from harvest were scanned and analyzed for number of first-order root tips (Supplementary Online Fig. S2).
Statistical analyses
JMP Pro 10 was used for all statistical analyses (SAS, Cary, NC, USA). Student’s t-tests were used to determine differences between treatments. A linear regression between resin-P and Olsen-P, total plant P uptake and resin-P, as well as total plant P uptake and Olsen-P established the effectiveness of the abiotic incubation extraction methods to predict total plant P uptake. Total plant biomass, plant P uptake, and plant calcium (Ca) uptake were transformed using a natural logarithm before statistical analyses.
Results
Abiotic incubation
Olsen-P extractions showed that increasing pyrolysis temperature decreased extractable P (p < 0.05) from rendered bone (Fig. 1a). In comparison to bone char pyrolyzed alone at either 350 °C and 750 °C, co-pyrolyzing bone with wood at 350 °C and 750 °C led to a 13–19 % increase in Olsen-P (p < 0.05). Olsen-P values from unpyrolyzed (60 °C) bone meal and bone meal pyrolyzed at 350 °C were 14–28 % higher than Olsen-P from TSP (p < 0.05).

Olsen-extractable P (a) and resin-extractable P (b) from P-fixing soil after a five-week incubation with different P sources and wood additions oven-dried at 60 °C or pyrolyzed at 350 °C or 750 °C. Treatments include rendered bone meal (RB), rendered bone meal and wood individually added after pyrolysis (RB + W), rendered bone meal and wood mixed before pyrolysis (RBW), Triple Super Phosphate (TSP) fertilizer, and TSP and wood biochar individually added (TSP + W) (n = 5; LSD is the least significant difference at p < 0.05)
Resin-P and Olsen-P were only weakly correlated (R2 = 0.13, p < 0.05). Resin-P was a better predictor for maize P uptake than Olsen-P, with R2 = 0.80 and R2 = 0.52, respectively (Supplementary Online Table S2). Pyrolyzing bone at 350 °C resulted in 67 % higher resin-P compared to unpyrolyzed bone (p < 0.05, Fig. 1b); however, this effect disappeared at 750 °C. Both the separate addition of wood biochar or co-pyrolysis of bone and wood decreased resin-P by 14–26 % in comparison to bone char alone (p < 0.05). Similar decreases in resin-P were observed for all TSP and wood biochar treatments. Adding fresh biomass to the unpyrolyzed bone led to a 23 % increase in resin-P (p < 0.05).
Pot trial
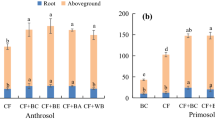
Plant P uptake by maize inoculated with AM was the same when receiving TSP or bone char fertilizer (Fig. 2a, p > 0.05), but significantly lower when fertilized with co-pyrolyzed bone char-wood biochar (p < 0.05). Without AM but with root hairs, TSP outperformed bone char fertilizer: maize fertilized with either bone char or co-pyrolyzed bone char-wood biochar showed both significantly lower P uptake than plants with TSP additions (p < 0.05).

Biomass and nutrient uptake of Z. mays after 5 weeks of growth fertilized with bone char produced at 750 °C (RB750), bone char-wood biochar co-pyrolyzed at 750 °C (RBW750), Triple Superphosphate (TSP), and no P additions (0-P) means and standard error are given (n = 5 except for -RH with n = 2–3): (a) plant P uptake, and (b) total plant Ca uptake, (c) total plant biomass, and (d) root:shoot ratio. On the x-axis, foraging strategy of maize is altered through the absence of root hairs (−RH), the presence of root hairs (+RH) and the presence of root hairs and addition of AM inoculants (+RH + AM). Different capital letters indicate significant differences (p < 0.05) between treatments as determined by student’s t-test
Plant Ca uptake followed similar trends as P uptake with two exceptions: the maize without root hairs showed no significant differences among additions and AM inoculation did not increase Ca uptake in co-pyrolyzed bone char-wood biochar or without additions (Fig. 2b, p > 0.05). Whole-plant P concentration was highest in maize inoculated with AM for all P additions, while the reverse was true for Ca concentration (Supplementary Online Table S3).
The presence of root hairs and AM inoculation also increased total plant biomass (p < 0.05) for all P sources (Fig. 2c). Unlike trends observed for P uptake, TSP application resulted in higher total plant biomass for maize with root hairs and maize with root hairs and AM in comparison to other P additions (p < 0.05). Maize with root hairs had higher total biomass when fertilized with bone char than with co-pyrolyzed bone char-wood biochar (p < 0.05), while maize without root hairs did not respond to P additions (p > 0.05).
The root:shoot ratios of maize without root hairs and with both root hairs and AM were significantly higher than those of maize without root hairs (p < 0.05, Fig. 2d). When maize was inoculated with AM, the root:shoot ratios after bone char additions were greater than those after TSP additions (p < 0.05). Without P application, root:shoot ratios of maize with or without root hairs were significantly lower than when other P sources were applied (p < 0.05).
AM inoculation significantly increased AM colonization of roots (p < 0.05) and did not vary among different P additions (Fig. 3a). For maize with both root hairs and AM, the degree of root branching measured as number of root tips was significantly higher for bone char and TSP additions in comparison to co-pyrolyzed bone char-wood biochar and zero-P additions (Fig. 3b, p < 0.05). Diameter, average root length and surface area of first-order roots did not demonstrate relevant trends among treatments (Fig. S3).

a Proportion of AM colonization of roots (means and standard error, n = 5, except for -RH treatments where n = 2–3) and b Number of root tips (means and standard error, n = 4 except for -RH treatments where n = 2–3). Phosphorus sources include no P application (0-P), rendered bone char produced at 750 °C (RB750), rendered bone char-wood biochar co-pyrolyzed at 750 °C (RBW750) and Triple Super Phosphate (TSP). On the x-axis, rooting strategy of maize is altered through the absence of root hairs (−RH), the presence of root hairs (+RH) and the presence of root hairs and AM inoculants (+RH + AM). Different capital letters indicate significant differences (p < 0.05) between treatments as determined by student’s t-test
Discussion
Bio-availability of phosphorus in bone char
Bone char proved to be an effective P fertilizer in highly weathered, acid soils. When inoculated with AM, maize plants acquired similar amounts of P from TSP and bone char pyrolyzed at 750 °C. Both resin-P and Olsen-P from soil incubated with bone char produced at 350 °C even indicated a higher P availability than when soils were incubated with TSP. While the incubation showed no difference in resin-P and Olsen-P between TSP and 750 °C bone char, maize plants without AM acquired more P from TSP in the pot trial. Unlike bone char, TSP is highly water-soluble and therefore becomes less available to plant roots over time through adsorption reactions with mineral oxide surfaces (Kucey et al. 1989). In the pot experiment, roots and AM may have taken up soluble P before chemisorption to soil minerals, explaining the higher P availability of TSP in comparison to measurements in soil incubation experiment. Given that resin-P contents were higher when bones were pyrolyzed at 350 °C than 750 °C in the incubation trial, P acquisition by maize from bone char may have been even higher if produced at lower pyrolysis temperatures than 750 °C bone char used in the pot trial. Since high soil acidity facilitates the continued dissolution of apatite through the release of structural OH− (Rajan et al. 1996), the effectiveness of bone char as fertilizer may be less in neutral or alkaline soil environments than observed in the studied acid soil.
Greater resin-extractable P from bone char produced at 350 °C in comparison to unpyrolyzed bone meal additions to soil may be explained by the increase in hydroxyl apatite-like crystals through pyrolysis, which reduces water-extractable P but increases formic-extractable P (Zwetsloot et al. 2015), representing plant-available P. Hydroxyl apatite may be more soluble than other calcium phosphates in solutions with low pH (Brown et al. 1975; Matsumoto et al. 2002), improving P availability in acid soil environments. In addition, pyrolysis between 250 and 500 °C frees up the organic bone constituents (Deydier et al. 2005). Cleaving organic P bonds could enhance extractable P (DeLuca et al. 2009). At pyrolysis temperatures greater than 500 °C, free P from organic bone constituents may become less available again through its incorporation into hydroxyl apatite crystal lattice structures. When added to soil, bone char produced at 750 °C indeed results in lower resin-P than bone char produced at 350 °C (Fig. 2b) despite its greater amounts of formic-acid extractable P reported previously (Zwetsloot et al. 2015).
The reduction in Olsen-P with an increase in pyrolysis temperature can also be explained by an increase in crystallinity. While the OH− anions from the Olsen solution (pH = 8.5) increase P solubility through complexing or precipitating aluminum (Al3+) and iron (Fe2+), the increase in soil solution pH might have limited the dissolution of calcium phosphate. Rather than the formic-P fraction, Olsen-P relies more on the water-P fraction of bone char, which decreases with greater calcium phosphate crystallization. Likewise, the increase in Olsen-P when rendered bone is co-pyrolyzed with wood biomass is likely attributed to a decrease in crystal formation and increase in water-P (Zwetsloot et al. 2015) rather than a reduction in soil P-fixation due to biochar.
Biochar effects on P availability
Mixing maple wood biochar with a P source before or after pyrolysis did not increase P availability in this study. This result was observed in both the abiotic incubation and pot experiment, suggesting that largely chemical interactions between P source, biochar and soil were responsible. The decrease in resin-P from TSP and bone fertilizer when mixed with biochar additions may be explained by ash minerals from biochar precipitating P out of solution (Hollister et al. 2013). In addition, biochar may have served as an adsorbent for organic acids already present in the soil (Cornelissen et al. 2005) increasing P fixation to soil minerals. Low molecular weight organic acids from root exudates and dead plant material are known to decrease P-fixation through competition with P for adsorption surfaces or complexing with Fe2+ and Al3+, thereby increasing the P concentration in the soil solution (Bolan et al. 1994; Jones 1998; Hinsinger 2001; Antelo et al. 2007). In contrast to pyrolyzed biomass, adding oven-dried wood biomass increased resin-P from bone meal possibly generating leachates that contain additional low-molecular weight organic acids that reduced P-fixation as often shown for plant residues (Earl et al. 1979; Haynes and Mokolobate 2001; Hunt et al. 2007). The importance of this competitive adsorption of low-molecular weight acids to biochar for soil P availability would need to be directly quantified in future experiments.
Managing soil pH is an important strategy for diminishing P adsorption to mineral surfaces and may also influence the extent to which biochar changes P fixation (Cui et al. 2011). Some biochars themselves can significantly increase soil pH thereby increasing the availability of essential plant nutrients such as P (Van Zwieten et al. 2009; Atkinson et al. 2010). Yet a significant biochar effect on P availability through a change in soil pH seems unlikely to explain variations in plant-available P in this experiment. Soil pH in the pot trial was unaffected by different bone char, TSP and biochar additions (Online Supplementary Table S4). While soil pH significantly increased from 4.9 to 5.1 by bone meal and bone char additions in the abiotic incubation (Online Supplementary Table S5), these changes did not correspond with higher levels of resin-P (r 2 = −0.19, p < 0.05 and Olsen-P (r 2 = 0.08, p > 0.05). Moreover, wood biochar by itself did not lead to a significant change in soil pH in comparison to the control (Online Supplementary Table S5). Since the biochar application rates in this study were relatively low, higher rates may need to be used in order to manage soil pH for enhancing plant-available P.
Root and AM exploitation of P sources
Bone char presents a patchier P distribution in soil and takes longer to dissolve into solution than TSP. This may explain why only maize inoculated with AM demonstrated similar P uptake under TSP and bone char fertilizer additions. AM aid plants in foraging for unevenly distributed nutrients by increasing the soil volume that is explored for P (Cui and Caldwell 1996) and also facilitate mineral weathering of apatite (Smits et al. 2012), such as bone char. Therefore, roots without AM were less successful in mining P from bone char explaining lower P acquisition in comparison to plants fertilized with TSP where P was more evenly distributed throughout the soil volume. In order to optimize plant P nutrition from bone char, it is important to consider plant foraging ability and AM inoculation.
When receiving P fertilizer, root branching appeared to be a strategy to enhance AM infection rather than P acquisition by roots. In comparison to plants with only root hairs, maize roots associated with AM had a higher number of first-order root tips with additions of either bone char or TSP. Previous studies have also reported increased branching in response to AM inoculation (Berta et al. 1993; Kaldorf and Ludwig-Muller 2000; Akiyama et al. 2005). Larger diameter and longer laterals in response to AM were not detected here, but have been observed in other studies (Mosse 1962; Vierheilig 2004).
On the other hand, co-pyrolysis of wood with bone led to no change in number of root tips and reduced plant P uptake in maize inoculated with AM. Yet AM colonization was unaffected by biochar additions in comparison to bone char (Fig. 3b), suggesting that biochar effects on plant P uptake are primarily chemical rather than biological in the present experiment. As shown by the abiotic soil incubation, biochar can reduce soil P extractability. This could explain the similar trends in number of root tips shown with biochar or no additions. Nevertheless, biological mechanisms for a reduction in root branching under biochar application cannot be excluded. Other studies have suggested that biochar may interfere with AM-plant communication through sorption of signaling and allelopathic compounds (Warnock et al. 2007) leading to a reduction in root thickening and specific root length of plants colonized by AM (Vanek and Lehmann 2015). In these cases, biochar also demonstrated a positive effect on AM colonization (LeCroy et al. 2013; Vanek and Lehmann 2015). Yet biochar-induced changes in root morphology were not observed in this study (Supplementary Online Fig. S3).
Conclusion
Bone char proved to be as effective for P fertilization as commercial TSP fertilizer in a highly P-fixing soil. Simultaneously managing soil P inputs and AM-root interactions improved plant nutritional status. However, co-pyrolyzing maple wood with bones decreased P availability in both the abiotic incubation and pot experiment. This suggests that chemical mechanisms were partly responsible for biochar-induced changes in plant-available P. Along with a reduction in P acquisition by maize, co-pyrolyzed maple biochar also decreased the number of root tips when inoculated with AM, despite greater extractability using Olsen-P soil test and no change in AM colonization. The mechanisms by which biochar affects root branching and P acquisition of AM-inoculated plants warrant further research. In addition, future greenhouse experiments and field studies should focus on the fertilizer efficacy of bone char with different pyrolysis temperatures in other soil environments and over longer periods of time.
References
Abelson PH (1999) A potential phosphate crisis. Science 283:2015
Akiyama K, Matsuzaki K, Hayashi H (2005) Plant sesquiterpenes induce hyphal branching in arbuscual mycorrhizal fungi. Nature 435:824–827
Antelo JF, Arce F, Avena M, Fiol S, Lopez R, Macias F (2007) Adsorption of a soil humic acid at the surface of goethite and its competitive interaction with phosphate. Geoderma 138:12–19
Atkinson CJ, Fitzgerald JD, Hipps NA (2010) Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: a review. Plant Soil 337:1–18
Bates TR, Lynch JP (2001) Root hairs confer a competitive advantage under low phosphorus availability. Plant Soil 236:243–250
Benton Jones J (2001) Laboratory guide for conducting soil tests and plant analysis. CRC Press LLC, Boca Raton
Berta G, Fusconi A, Trotta A (1993) VA mycorrhizal infection and the morphology and function of root systems. Environ Exp Bot 33:159–173
Bolan NS, Naidu R, Mahimairaja S, Baskaran S (1994) Influence of low-molecular weight organic-acids on the solubilization of phosphates. Biol Fertil Soils 18:311–319
Brown WE, Patel PR, Chow LC (1975) Formation of CaHPO4 from enamel mineral and its relationship to caries mechniasm. J Dent Res 54:475–481
Buss W, Graham MC, Shepherd JG, Mašek O (2016) Suitability of marginal biomass-derived biochars for soil amendment. Sci Total Environ 547:314–322
Cordell D, Drangert JO, White S (2009) The story of phosphorus: global food security and food for thought. Glob Environ Chang 19:292–305
Cordell D, Rosamarin A, Schroder JJ, Smit AL (2011) Towards global phosphorus security: a systems framework for phosphorus recovery and reuse options. Chemosphere 84:747–758
Cornelissen G, Gustafsson O, Bucheli TD, Jonker MTO, Koelmans AA, Van Noort PCM (2005) Extensive sorption of organic compounds to black carbon, coal, and kerogen in sediments and soils: mechanisms and consequences for distribution, bioaccumulation, and biodegradation. Environ Sci Technol 39:6881–6895
Cui MY, Caldwell MM (1996) Facilitation of plant phosphate acquisition by arbuscular mycorrhizas from enriched soil patches II. Hyphae exploiting root-free soil. New Phytol 133:461–467
Cui HJ, Wang MK, Fu ML, Ci E (2011) Enhancing phosphorus availability in phosphorus-fertilized zones by reducing phosphate adsorbed on ferrihydrite using rice straw-derived biochar. J Soils Sediments 11:1135–1141
DeLuca TH, MacKanzie MD, Gundale MJ (2009) Bio-char effects on soil nutrient transformation. In: Lehmann J, Joseph S (eds) Biochar for environmental management: science and technology. Earthscan Publications Ltd, London, pp. 251–270
Deydier E, Guilet R, Sarda S, Sharrock P (2005) Physical and chemical characterisation of crude meat and bone meal combustion residue: “waste or raw material?”. J Hazard Mater 121:141–148
Earl KD, Syers JK, McLaughlin JR (1979) Origin of the effects of citrate, tartrate and acetate on phosphate soprtion by soils and synthetic gels. Soil Sci Soc Am J 43:674–678
Gilbert N (2009) The disappearing nutrient. Nature 461:33–143
Guppy CN, Menzies NW, Moody PW, Blamey FPC (2005) Competitive sorption reactions between phosphorus and organic matter in soil: a review. Aust J Soil Res 43:189–202
Haynes RJ, Mokolobate MS (2001) Amelioration of Al toxicity and P deficiency in acid soils by additions of organic residues: a critical review of the phenomenon and the mechanisms involved. Nutr Cycl Agroecosyst 59:47–63
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195
Hochholdinger F, Wen TJ, Zimmerman R, Chimot-Marolle P, da Costa e Silva O, Bruce W, Lamkey KR, Wienand U, Scnable PS (2008) The maize (Zea mays L.) roothairless3 gene encodes a putative GPI-anchored, monocot-specific, COBRA-like protein that significantly affects grain yield. Plant J 54:888–898
Hodge A, Berta G, Doussan C, Merchan F, Crespi M (2009) Plant root growth, architecture and function. Plant Soil 321:153–187
Hollister CC, Bisogni JJ, Lehmann J (2013) Ammonium, nitrate, and phosphate sorption to and solute leaching from biochars prepared from corn stover (Zea mays L.) and oak wood (Quercus spp.). J Environ Qual 42:137–144
Hunt JF, Ohno T, He Z, Honeycutt CW, Dail DB (2007) Inhibition of phosphorus sorption to goethite, gibbsite, and kaolin by fresh and decomposed organic matter. Biol Fertil Soils 44:277–288
Jones DL (1998) Organic acids in the rhizosphere - a critical review. Plant Soil 205:25–44
Kaldorf M, Ludwig-Muller J (2000) AM fungi might affect the root morphology of maize by increasing indole-3-butyric acid biosynthesis. Physiol Plant 109:58–67
Koske RE, Gemma JN (1989) A modified procedure for staining roots to detect VA-mycorrhizas. Mycol Res 92:486–505
Kucey RMN, Janzen HH, Leggett ME (1989) Microbially mediated increases in plant-available phosphorus. Adv Agron 42:199–228
Kuo S (1996) Phosphorus. In: Sparks DL (ed) Methods of soil analysis - Part 3 chemical methods. Soil Science Society of America, Inc. & American Society of Agronomy, Madison, pp. 895–997
LeCroy C, Masiello CA, Rudgers JA, Hockaday WC, Silberg JJ (2013) Nitrogen, biochar, and mycorrhizae: alteration of the symbiosis and oxidation of the char surface. Soil Biol Biochem 58:248–254
Lehmann JJ, da Silva P, Steiner C, Nehls T, Zech W, Glaser B (2003) Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: fertilizer, manure and charcoal amendments. Plant Soil 249:343–357
Lynch JP (2011) Root phenes for enhanced soil exploration and phosphorus acquisition: tools for future crops. Plant Physiol 156:1041–1049
Lynch JP, Beebe SE (1995) Adaptation of beans (Phaseolus vulgaris L) to low phosphorus availability. Hortscience 30:1165–1171
Marschner H (1995) Mineral nutrition of higher plants. Academic Press, London
Matsumoto T, Okazaki M, Inoue M, Hamada Y, Taira M, Takahashi J (2002) Crystallinity and solubility characteristics of hydroxyapatite adsorbed amino acid. Biomaterials 23:2241–2247
Mosse B (1962) Establishment of vesicular-arbuscular mycorrhiza under aseptic conditions. J. Gen Microbiol 27:509–520
Murphy J, Riley JP (1962) A modified single solution method for determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Parfitt RL, Atkinson RJ, Smart RSC (1975) Mechanism of phosphate fixation by iron oxides. Soil Sci Soc Am J 39:837–841
Qayyam MF, Ashraf I, Abid M, Steffens D (2015) Effect of biochar, lime, and compost application on phosphorus adsorption in a Ferralsol. J Plant Nutr Soil Sci 178:576–581
Raghothama KG, Karthikeyan AS (2005) Phosphate acquisition. Plant Soil 274:37–49
Rajan SSS, Watkinson JH, Sinclair AG (1996) Phosphate rocks for direct application to soils. Adv Agron 57:77–159
Ramaekers L, Remans R, Rao IM, Blair MW, Vanderleyden J (2010) Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crop Res 117:169–176
Richardson AE, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Ryan MH, Veneklaas EJ, Lambers H, Oberson A, Culvenor RA, Simpson RJ (2011) Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 349:121–156
Sanchez P (1976) Properties and management of acid soils in the tropics. John Wiley, New York
Siebers NF, Leinweber P (2013) Bone char: a clean and renewable phosphorus fertilizer with cadmium immobilization capability. Soil Sci Soc Am J 42:405–411
Siebers NF, Godlinski F, Leinweber P (2012) The phosphorus fertilizer value of bone char for potatoes, wheat and onions: first results. Landbauforsch Volk 62:59–64
Siebers NF, Godlinski F, Leinweber P (2014) Bone char as phosphorus fertilizer involved in cadmium immobilization in lettuce, wheat, and potato cropping. J Plant Nutr Soil Sci 177:75–83
Singh BB, Jones JP (1976) Phosphorus sorption and desorption characteristics of soil as affected by organic residues. Soil Sci Soc Am J 40:389–394
Smil V (2000) Phosphorus in the environment: natural flows and human interferences. Annu Rev Energy 25:53–88
Smits MM, Bonneville S, Benning LG, Banwart SA, Leake JR (2012) Plant-driven weathering of apatite - the role of an ectomycorrhizal fungus. Geobiology 10:445–456
Tiessen H, Moir J (1993) Characterization of available P by sequential fractionation. In: Carter M (ed) Soil sampling and methods of analysis. Lewis, Boca Raton, pp. 75–78
Tuominen L, Kairesalo T, Hartikainen H (1994) Comparison of methods for inhibiting bacterial-activity in sediment. Appl Environ Microbiol 60:3454–3457
Van Zwieten L, Kimber S, Morris S, Chan KY, Downie A, Rust J, Joseph S, Cowie A (2009) Effect of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility. Plant Soil 327:235–246
Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol 157:423–447
Vanek S, Lehmann J (2015) Phosphorus availability to beans via interactions between mycorrhizas and biochar. Plant Soil 395:105–123
Vassilev N, Martos E, Mendes G, Martos V, Vassileva M (2013) Biochar of animal origin: a sustainable solution ot the global problem of high-grade rock phosphate scarcity? J Sci Food Agric 93:1799–1804
Vierheilig H (2004) Regulatory mechanisms during the plant-arbuscular mycorrhizal fungus interaction. Can J Bot 82:1166–1176
Warnock DD, Lehmann J, Kuyper TW, Rillig MC (2007) Mycorrhizal responses to biochar in soil - concepts and mechanisms. Plant Soil 300:9–20
Warren GP, Robinson JS, Someus E (2009) Dissolution of phosphorus from animal bone char in 12 soils. Nutr Cycl Agroecosyst 84:167–178
Wen TJ, Schnable (1994) Analyses of mutrants of 3 genes that influence root hair development in Zea mays (Gramineae) suggest that root hairs are dispensable. Am J Bot 81:833–842
Wopenka B, Pasteris J (2003) Biological apatites: a comparison of bone and tooth mineralization. J Vertebr Paleontol 23:112A–112A
Wopenka B, Pasteris JD (2005) A mineralogical perspective on the apatite in bone. Mater Sci Eng C 25:131–143
Zhu J, Zhang C, Lynch JP (2010) The utility of phenotypic plasticity of root hair length for phosphorus acquisition. Funct Plant Biol 37:313–322
Zwetsloot MJ, Lehmann J, Solomon D (2015) Recycling slaughterhouse waste into fertilizer: how do pyrolysis temperature and biomass additions affect phosphorus availability and chemistry? J Sci Food Agric 95:281–288
Acknowledgments
We are grateful for support from the Towards Sustainability Foundation, CARE-Cornell Impact through Innovations Fund, McKnight Foundation, Bradfield Award, Fulbright and Huygens Talent Scholarship Program. We would also like to thank Cornell Center for Materials Research for help with X-ray Diffraction Analysis under NSF award number DMR-0520404, Berhanu Belay and Gebermedihin Ambaw for support in procuring the soil, and Dawit Solomon for help with data interpretation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible Editor: Andreas Meyer-aurich.
Electronic supplementary material
ESM 1
(DOCX 424 kb)
Rights and permissions
About this article

Cite this article
Zwetsloot, M.J., Lehmann, J., Bauerle, T. et al. Phosphorus availability from bone char in a P-fixing soil influenced by root-mycorrhizae-biochar interactions. Plant Soil 408, 95–105 (2016). https://doi.org/10.1007/s11104-016-2905-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-016-2905-2