
Abstract
Aims
We hypothesised that a) species with greater acid soil tolerance have an increased capacity to utilise incipient rainfall; and b) liming increases the productivity and the ability of pasture species to utilise available water resources in the profile of an acid soil.
Methods
A field experiment was established on a moderately acidic yellow Kandosol and monitored over 5 years. Five perennial pasture species including lucerne (Medicago sativa L.), phalaris (Phalaris aquatica L.), chicory (Cichorium intybus L.), tall fescue (Festuca arundinacea Schreb.) and cocksfoot (Dactylis glomerata L.), were sown in monocultures with and without 2.9 t/ha lime.
Results
Both lucerne and phalaris were more persistent than chicory, tall fescue and cocksfoot under severe drought, despite both being considered sensitive to soil acidity. Surface liming increased the soil water deficit by up to 27 mm at 0.75–1.65 m under perennial pastures compared to unlimed treatments, despite lime having no physical presence at that depth. Lime increased lucerne, phalaris and cocksfoot cumulative herbage biomass by 150, 30 and 20 %, respectively, but had no significant effect on chicory or tall fescue biomass.
Conclusions
The two most acid-sensitive species, lucerne and phalaris, were more resilient under drought despite the acidic nature of the soil. We contend that species sensitive to acidity can be a valuable addition to pastures on acid soils. Lime used in conjunction with deep-rooted perennial species is likely to maximise the ability of pastures to utilise scarce available soil water reserves.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil acidity is an impediment to agricultural production across much of southern Australia (Scott et al. 2000; Bolland et al. 2002). In New South Wales (NSW) alone it affects in excess of 24 million ha of agricultural land holdings (Helyar et al. 1990) with many soils continuing to acidify (Scott et al. 2007). Beneficial effects of lime (CaCO3) in ameliorating acid soils are well documented (McLachlan 1980; Singer and Munns 1996), but there are practical limitations with incorporating it into the subsoil (Scott et al. 1997). The movement of lime into the soil profile beyond the zone of incorporation takes decades (Li et al. 2010), meaning that in many instances, lime is only a partial solution for the management of soil acidity.
The leaching of soil nitrates derived from the nitrification process to beyond the plant root zone is a primary cause of soil acidification in the farming systems of southern Australia (Helyar 1976). With their longer-lived and deeper root systems, perennial plant species have greater capacity to scavenge soil nitrate from the soil profile prior to it leaching beyond the root zone, thus reducing acidification compared to annual plant species. It was reported that the presence of perennial species has been shown to reduce surface soil acidity compared to where only annual species exist (Ridley et al. 2002; Dear et al. 2009). Thus, the use of perennial pastures in conjunction with lime is promoted as a method of sustainable agricultural production on acidic soils (Li et al. 2006a, b). In environments where drought is common, there are also potential advantages in perennial species maximising the use of both incipient rainfall and stored soil water reserves.
Much research was done in Australia in the previous decade to develop farming systems with enhanced capacity to de-water the soil profile. Inefficient use of soil water leads to various forms of soil degradation, not only soil acidity as described above, but also dryland salinity which was the catalyst for much of the recent research conducted in Australia (see Dunin and Passioura 2006). The highly variable and episodic nature of rainfall across agricultural regions of southern Australia requires a ‘dry soil buffer’ (or soil water deficit) to be in place to store excess water before it is lost below the plant root zone (Ward 2006). This ‘insurance policy’ does not eliminate natural soil degradation processes associated with the loss of water and nutrients to beyond the plant root zone, but serves to minimise the frequency and severity of leaching events. Perennial pasture species are seen as a practical means to increase the soil water deficit across the agricultural landscape in order to mitigate the risk of soil degradation whilst maintaining or improving agricultural productivity (Dear and Ewing 2008).
The challenge for agricultural landscapes across southern Australia is that very few viable alternative perennial pasture species exist which are sufficiently productive to add value to existing crop and livestock enterprises whilst also being sufficiently resilient to withstand the stresses imposed by biotic and abiotic factors. Aside from specific niches, lucerne (Medicago sativa L.) is the only perennial legume species broadly adapted to a range of cropping environments across southern Australia (Li et al. 2008). The capacity of lucerne to fix atmospheric nitrogen makes it an appealing option for livestock (Wolfe et al. 1980) and crop (Peoples and Baldock 2001) production enterprises. In addition, its ability to scavenge mineral N from the soil (Dear et al. 1999) and de-water the soil profile (Sandral et al. 2006; Hayes et al. 2010c) are appealing in the context of managing soil acidity and salinity. However, lucerne is inherently sensitive to soil acidity (Bouton 1996; Hayes et al. 2012a) which may limit its use, particularly where subsoil acidity is a constraint to production (Pinkerton and Simpson 1986).
A small number of non-legume perennial species exist which may prove to be viable alternatives to lucerne in environments across southern Australia. Chicory (Cichorium intybus L.) is a broadly adapted (Li et al. 2008) summer growing, short-lived perennial herb with a proven capacity to de-water the soil profile (Hayes et al. 2010c) and provides high quality forage for livestock (Belesky et al. 2001; Hayes et al. 2010a). Temperate perennial grasses may also prove to be useful for this purpose, particularly in the higher rainfall zones of southern Australia. Phalaris (Phalaris aquatica L.) is perhaps the best grass option due to its broad adaptation across south-eastern Australia (Oram et al. 2009), as well as its relatively high level of productivity (Hayes et al. 2010a) and deep rooting habit (McWilliam and Kramer 1968; Ridley and Simpson 1994). However, phalaris is reported as being sensitive to low pH (Edmeades et al. 1991) and to Al toxicity (Culvenor et al. 1986). Tall fescue (Festuca arundinacea Schreb.) is similar to phalaris in many respects although would seem to be generally less sensitive to soil acidity (Lattimore and McCormick 2012; Song et al. 2014), but is also less productive than phalaris (Hayes et al. 2010a), particularly at lower latitudes. Cocksfoot (Dactylis glomerata L.) is the most likely option as it is highly tolerant of soil acidity (Culvenor et al. 1986; Ridley and Coventry 1992) and is also a reasonably productive forage species (Hill 1985; Hayes et al. 2010a) although its rooting depth is generally much shallower than the other species previously discussed (Ridley and Simpson 1994). Aside from this narrow range of perennial species, there appears to be very few alternative perennial species that are sufficiently robust to survive and thrive under paddock conditions across much of the target region (Hayes et al. 2012b).
The objective of the current study was to assess the production, persistence and soil drying characteristics of lucerne, chicory, phalaris, cocksfoot and tall fescue grown with and without lime on an acidic soil, typical of south-west NSW. More specifically, we hypothesise that a) species with superior acid soil tolerance have an increased capacity to dry the profile in soil environments with an acidic subsurface; and b) the application of lime increases the productivity of perennial pasture species enabling them to better exploit stored soil water in soils with an acidic subsurface.
Materials and methods
Site description and treatments
The field experiment was conducted near Gerogery, southern NSW, Australia (35.9025° S, 146.9377° E; Alt. 239 m). The soil was a Yellow Kandosol (Isbell 1996) with a sandy clay loam texture in the surface 0.5 m over a light clay, with clay content increasing down the profile. The subsurface soil pHCa (1:5 soil in 0.01 M CaCl2) was as low as 4.1 (Fig. 1), even where lime was applied. The soil below 0.40 m was not acidic and had concentrations of Al and Mn that would not be toxic to plants. Prior to treatments being imposed the experimental area had previously never been limed. Previous experiments conducted at this field site showed visual symptoms of Mn toxicity in legume species which was confirmed by high Mn concentrations in plant tissue. As a result, pasture productivity was reduced where the soil acidity was not amended by lime (Hayes et al. 2008, 2012a).

The experiment was a split-plot design with the 5 species as main plots and lime treatments (limed vs unlimed) as subplots, replicated 4 times. Plot size was 6 × 4 m. The 5 species were lucerne cv. Aurora (sown at 6 kg/ha), chicory cv. Grasslands Puna (5 kg/ha), phalaris cv. Landmaster (8 kg/ha), cocksfoot cv. Currie (6 kg/ha), and tall fescue cv. Fraydo (12.5 kg/ha). All species were sown as monocultures, and lucerne seed was inoculated and lime pelleted prior to sowing.
Superfine F70 lime (CaCO3) was applied at 2.9 t/ha to the surface of the limed plots using a direct drop spreader on 14 April 2002 and incorporated 2 weeks later on 28 April 2002 with a tyned seed drill as described by (Hayes et al. 2008). The current species treatments were re-sown over existing treatments with the same species and sowing rate on 25 August 2004 when the previous study was completed. Throughout this paper we refer to 2004 as ‘year 1’, 2005 as ‘year 2’ and so on.
The site was sprayed with 2 L/ha glyphosate several weeks prior to the August 2004 sowing. It was cultivated using a small offset-disc plough immediately before sowing, taking care to ensure the movement of soil between plots was minimised. Soil pH in each plot was monitored in 2002, 2003,2007 and 2009 and results showed that lime increased the pH of the soil at 0–0.1 m, but had only small effects below that depth (Fig. 1; see also Table 5).
Drier than average seasonal conditions were experienced during the experimental period which was characterised by a series of very dry autumns leading to a late start to the growing season in each year (Table 1). Total rainfall was 722 mm in 2005, 13 % above the long term average (628), but 535 mm in 2004, 247 mm in 2006, 563 mm in 2007, and 469 mm in 2008, being 15, 61, 10 and 24 % below the long term average, respectively. Year 3 (2006) was the single driest year in 86 years of observation at Walla Walla, NSW, the weather recording station nearest the experimental site (Bureau of Meteorology 2014).
Site management
Boron was applied to the entire experimental area as a bare earth treatment in the form of Borax at 20 kg/ha prior to sowing (12 May 2004), as boron concentrations were marginally below critical concentrations for subterranean clover in the previous experiment at that site (Hayes et al. 2008). Application was through a boom-spray with 670 L/ha of water due to its low solubility. The site received 2 applications of starter fertiliser (14.9 % N, 13 % P, 10.5 % S) at 180 kg/ha in May (sown and failed) and in late August (re-sown). In year 2 the site received approximately 160 kg/ha molybdenised superphosphate (8.8 % P, 11 % S, 0.025 % Mo) and 100 kg/ha granulated urea, both applied through a direct-drop spreader on 2 March 2005. Granulated urea was again applied to all treatments at 100 kg/ha and single superphosphate (8.8 % P, 11 % S) at 150 kg/ha in year 3 on 4 August 2006 and in year 4 on 6 September 2007.
Alpha-cypermethrin was applied at 100 ml/ha approximately 1 week after sowing as a preventative measure to minimise insect damage to emerging seedlings from pests such as red-legged earth mite (Halotydeus destructor Tucker). No subsequent application of insecticide was applied to the experiment. The phalaris, cocksfoot and tall fescue treatments received Flumetsulam at 25 g/ha through a shrouded boom in year 1 on 24 September 2004 to control emerging wireweed (Polygonum aviculare L.). It was not applied to the lucerne and chicory treatments at this time as it was uncertain if these species could tolerate this herbicide at such an early stage of development. However, a second application of Flumetsulam at 25 g/ha was applied to all treatments on 22 October 2004 to suppress any remaining wireweed. In year 2 Flumetsulam was again applied to all treatments at 25 g/ha on 8 August 2005 to suppress winter broadleaved weeds such as capeweed (Arctotheca calendula L.) as well as emerging wireweed. No selective grass herbicides were applied to this experiment as the spring sowing precluded significant grass weed burden in the establishment year, and subsequent grass weed burdens were reduced with grazing. No pesticides were applied to the experiment beyond year 2.
The experiment was not grazed in the establishment year. In subsequent years the site was grazed at a high stocking rate with either sheep or cattle immediately following an assessment of herbage yield. If required the experiment would subsequently be mowed, ensuring all treatments were defoliated to a uniform height to avoid differences due to grazing selectivity. A residual of 300–500 kg/ha of biomass remained on the plots after defoliation.
Measurements
Initial seedling density of perennial species was assessed 9 weeks after sowing using two 1 m × 1 m quadrats per plot. Plant density was measured again in year 2 on 5 July 2005 at the same locations. From year 3 onwards, the relative plant density of the perennial species were taken by measuring basal frequency as many established plants were too large to reliably distinguish individuals. The basal frequency was assessed at fixed locations in year 3 (21 July), year 4 (1 June) and year 5 (10 June) by placing a 1 × 1 m quadrat of 100 × 100 mm mesh over the sampling area, and counting the percentage of squares occupied or partially occupied by the base of a sown perennial plant.
Herbage yield of each plot was assessed periodically, usually coinciding with the end of a season. A visual assessment technique was used which involved splitting each plot into 6 cells and giving each a score out of 10 for herbage mass. Scores of each cell were averaged to give one score per plot. Scores were calibrated and converted to a dry matter basis by cutting 10–12 representative quadrats (0.1 m2) at each sampling date and drying samples at 70 °C for 48 h before weighing. Of the 15 assessments of herbage yield the coefficient of determination of the calibrations (r2) ranged from 0.51 to 0.98 (P < 0.05). The proportion of sown perennial species, annual legumes and weeds was assessed at the same time using the dry weight rank method (t’ Mannetje and Haydock 1963; Jones and Hargreaves 1979) at 10 random locations in each plot. Botanical composition data was used to calculate the yields of the respective components of each sward.
Twenty grab samples of lucerne, chicory and phalaris were taken from individual plots for determination of mineral concentrations in the year following lime application and were reported in Hayes et al. (2008). Lucerne, phalaris and cocksfoot were again sampled at the conclusion of the current experiment on 9 September 2008. The herbage samples were kept cool and transported back to the laboratory for drying at 70 °C for 72 h before grinding to < 1 mm. Analysis for minerals using radial view inductively coupled plasma-optical emission spectrometry (ICP-OES) followed acid digestion of samples. Values for Mo and Co are not reported as they were below levels of accurate determination of the instrumentation. Chicory and tall fescue were not sampled due to insufficient herbage available at the time of sampling.
An aluminium access tube was inserted at the centre of each plot to enable the measurement of soil water using a neutron moisture meter (NMM) at depths of 0.15, 0.30, 0.45, 0.60, 0.75, 0.90, 1.15, 1.40, 1.65, 1.90, 2.20, 2.50 and 2.80 m. Due to the presence of a hard gravely layer, 17 of the 40 tubes inserted did not reach maximum depth. Each soil core was retained from each respective depth for the determination of gravimetric moisture content and these numbers used for the calibration of the NMM. Tubes were inserted 2 years prior to sowing and the initial NMM reading for the current experiment was taken 11 weeks prior to sowing. Subsequent readings of the NMM were generally taken about every 8 weeks throughout the experimental period.
The NMM was calibrated at field capacity at the above depths in the surface 1.65 m in March 2007. An access tube was inserted in the middle of a 4 × 4 m area of soil immediately adjacent to the experiment which was covered with black plastic and irrigated over a 6 week period using a gravity-fed dripper line. At the end of the 6 week period a NMM reading was taken as being the average of 3 counts for each depth. The soil profile adjacent to the access tube was sampled in triplicate to determine bulk density and gravimetric moisture content of the soil at each sampling depth. Soil was removed using a 150 mm-diameter auger to the appropriate depth before a 75 × 50 mm bulk density coring ring was inserted such that the midpoint of the ring was at the target sample depth (e.g., 0.15 m, 0.30 m etc.). Estimates of soil bulk density comprised the average of 3 cores at each depth and ranged from 1.55 to 1.62 g/cm3 in the surface 1.65 m, except for at 1.40 m where bulk density was 1.71 g/cm3. Bulk densities at depths beyond 1.65 m were assumed to be the average of estimates taken between 1.15 and 1.65 m (1.65 g/cm3). All bulk density and gravimetric moisture content values were converted to volumetric water content and related to the respective NMM readings. Additional dry-end calibration readings were taken in 2009 from only the lucerne and phalaris plots at each depth to 1.65 m and related to the mean of 3 NMM readings in each respective tube. All calibration data were combined to develop equations of best fit for each depth to convert NMM readings to volumetric water content (data not shown).
Soil samples were taken using a foot corer (0.02 m in diameter) from the surface 2 depths, 0–0.1 and 0.1–0.2 m, to monitor soil pH. Ten cores were taken across a transect in every plot and bulked. Samples were dried at 40 °C, sieved to < 2 mm and analysed for pH following mixing of soil for 1 h with 0.01 M CaCl2 in a 1:5 ratio. Soil was sampled on 4 occasions including twice prior to the establishment of the current experiment in August 2002 (133 days after liming) and September 2003, and twice during the current experiment in August 2007 and August 2009.
Statistical analysis
Analyses of variance were conducted using a split-plot model (Genstat version 9; VSN International Ltd) with perennial species as the main plot and lime treatment as the subplot. A linear mixed model with spatial components (AR1 × AR1) was fitted with species, lime and species × lime as fixed terms. Random terms included replicate, replicate × main plot and replicate × main plot × subplot. Row and column effects were fitted as required. The distribution of all datasets and residuals were sufficiently normal for the data not to require transformation prior to analysis. The NMM measurement on 19 October 2005, the wettest soil profile during the experimental period, was used as a covariant to compare levels of drying achieved under treatments until 9 July 2008. The repeated measures model was used with species, lime and time, and their interactions as fixed effects, and replicate and plot as random effects. All data were analysed at the 95 % significance level (P < 0.05).
Results
Plant density
All perennial species established successfully with initial seedling densities ranging from 41 to 189 plants/m2 (Table 2). Lime had no effect on the establishment of the perennial species, nor on plant density in year 2. However, in most treatments there was a marked decline in plant density between years 1 and 2. Lucerne density reduced from 47 to 31 plants/m2 (P < 0.05) over the first summer in the unlimed treatment but did not reduce (41–47 plants/m2) where lime was applied. The basal frequency of all species declined between years 3 and 5 irrespective of lime treatment. However, the extent of the decline observed in the basal frequency of lucerne (18 %) was minor compared to the average decline of the remaining species (76 %; Table 2). Lime increased the basal frequency of lucerne by up to 79 % (year 3), but differences due to lime in other species were not significant (P > 0.05; Table 2).
Aboveground biomass
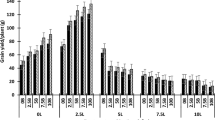
Phalaris and lucerne both produced the highest cumulative aboveground biomass during the 4 year experimental period (Aug 2004 – Sep 2008), particularly where lime was applied. Cumulative biomass of sown perennial species increased with lime application by ca. 30 and 150 % in phalaris and lucerne, respectively (Fig. 2). The cumulative biomass of the remaining sown species was not affected by lime. Averaged across lime treatments, the phalaris swards had the highest total cumulative biomass over 5 years (39.8 t/ha), followed by the lucerne (36.2 t/ha), cocksfoot (35.5 t/ha) and chicory (34.9 t/ha) swards. Tall fescue was the least productive of the perennial species (33.3 t/ha) although its total biomass was not significantly different from cocksfoot and chicory swards (P > 0.05). In most instances, ca. 50 % of total available biomass comprised ‘other’ generally undesirable volunteer weed species, such as Hordeum vulgare L. and Lolium rigidum Gaud. in winter or Polygonum aviculare L. and Pannicum capilare L. in summer. The exceptions to this were in the tall fescue swards where the tall fescue contributed only ca. 15 % to total sward biomass, and in the limed lucerne and phalaris swards in which ‘other’ species comprised only 30–40 % of total biomass (Fig. 2).

Cumulative available herbage biomass (t/ha) for each perennial species with and without lime measured from spring 2004 to winter 2008. Vertical bars represent LSD0.05 for the interaction between species and lime for sown perennial species and other weeds, respectively; n.s., not significant for annual legumes
Averaged across all species, lime increased available biomass of the sown perennial species by 39 % over the 5 year experimental period, with significant differences in available biomass observed on 13 out of 15 sampling times (Table 3). The maximum increase in available biomass of the mean of all sown perennial species due to lime was 69 %, observed in spring of both years 3 and 4 (Table 3). Patterns of biomass availability were generally consistent with the seasonal growth patterns of the various species. Lucerne and chicory tended to be more productive over summer and autumn whereas maximum available biomass of the grasses was generally observed in winter and spring, although this was very much dependant on the conditions of the particular season. Pasture biomass production was very low during the final 2 years of experimentation, reflecting the low rainfall received at that site during that period (Table 1) and poor persistence of most of the sown perennial species. Lucerne was the most productive species in the final 2 years of experimentation, with cumulative aboveground biomass of 4.1 and 8.9 t/ha in the nil and plus lime swards, respectively (Table 3).
Concentrations of minerals were generally highest in lucerne and lowest in phalaris herbage (Table 4). This was particularly true for B and Ca where concentrations were 10-fold and 5-fold greater than in the grasses, respectively. The exception was Na which was at more than double the concentration in phalaris herbage compared to cocksfoot and lucerne. The concentrations of all minerals other than Mn were unaffected by lime. Averaged across the three species, lime reduced Mn concentrations from 221 to 176 mg/kg (P < 0.001). Values of Mn in lucerne herbage ranged from 136 mg/kg in the limed plots to 199 mg/kg in the unlimed plots (data not shown).
Soil water
There was no significant species × lime × time interaction in soil water data at any depth. Time was a highly significant factor, reflecting the strong seasonal influence on soil water content. A significant species × time effect was observed at all depths and a lime effect was observed only in the 0.75–1.65 m (P < 0.01) zones of the profile (Fig. 3). Averaged across all species, soil water deficit on the limed treatments increased by 9 mm in the 0.75–1.65 m depth at the end of spring (November) in year 4 compared to that on unlimed treatments (P < 0.01). In the lucerne and phalaris treatments where perennial plant basal frequency was higher throughout the experiment, lime increased the apparent soil water deficit significantly (P < 0.05). The maximum difference in soil water deficit was 20 mm between unlimed and limed plots under lucerne pastures in the 0.15–0.60 m zone. In contrast, the maximum difference in soil water deficit was up to 27 mm between unlimed and limed plots under phalaris in the 0.75–1.65 m zone. No differences were found below 1.65 m (P > 0.05, data not shown).

Changes in soil water deficit (mm) at 0.15–0.60 and 0.75–1.65 m below the soil surface on plots with (Lime+) and without (Lime-) surface lime application sown to (a) lucerne; (b) chicory; (c) phalaris; (d) cocksfoot; and (e) tall fescue from October 2005 to July 2008. Vertical bars represent LSD0.05 for measurement times on the left hand side and lime effects on the right hand side of each graph. No measurement time and lime effect interaction was detected for any species; n.s., not significant
Trends in soil pH through time
Lime significantly increased soil pH by 1.04 units in the surface 0.10 m 133 days following lime application and incorporation (Table 5). There was also a small but significant increase in pH at the 0.10–0.20 m depth. There was a consistent trend of increased acidification of soil over time. For example, soil pH at 0–0.10 m depth decreased by 0.76 pH units in the limed treatment, and by 0.18 pH units in the unlimed treatment from 2002 to 2009. At the 0.10–0.20 m depth, there was a significant decrease in soil pH between 2002 and 2009 mirroring trends in the surface depth in the unlimed treatment. However, in the limed plots, pH increased slightly over time with highest values observed in 2007 (Table 5). There was no effect of species on soil pH in the top 0.20 m soil.
Discussion
There are two major findings of this study. First, surface liming at 0–0.10 m enhanced the capacity of deep-rooted perennial species to deplete water in the profile to up to 1.65 m below the soil surface. Second, the most productive and persistent perennial species under sustained drought conditions at this acidic site were the two species, lucerne and phalaris, that were previously reported to be sensitive to acid soils and that were most responsive to lime in the current study. Both findings will result in changed recommendations to the way farmers should manage acid soils.
Lime and soil water
Results showed that there was a difference in soil water deficit in the 0.75–1.65 m zone of the profile under the phalaris treatment of up to 27 mm between the unlimed and limed treatments. In its simplest form, this might be equivalent to 27 mm of ‘bonus rainfall’ implying an effective average annual rainfall of something closer to 655 mm rather than the existing value of 628 mm at this site. However, this is overly simplistic and the benefits are likely to be much greater in a paddock situation. Firstly, 27 mm of additional water in the 0.75–1.65 m zone of the profile would require far more than 27 mm of rainfall received at the surface, due to losses associated with evaporation, lateral flow and runoff and due to the drainage characteristics of individual soils which may hamper the movement of water through the profile. Secondly, additional water deep in the profile may be accessed by perennial forage species less for production and more for survival purposes. In a lysimeter study, Hoffmann et al. (2003) showed that lucerne survival increased where there was access to deep soil water, but deep soil water had little impact on lucerne production when the upper profile remained dry. One might expect a similar response in phalaris which is a species known to rely on access to deep soil water to survive periodic droughts (McWilliam and Kramer 1968). Although differences in phalaris persistence due to lime were not significant in the current study, a consistent trend was observed over the experimental period where phalaris basal frequency was numerically greater in the limed plots. A study at Goulburn, NSW, showed that lime significantly increased the basal frequency of phalaris (Hayes et al. 2010b), an effect which could be attributed to increases in either persistence or plant size. Unlike the Goulburn study, no increase in soil water at the soil surface was observed due to lime at the current site which is likely to reflect differences in soil characteristics between sites.
The increase in soil water deficit at 0.75–1.65 m below the soil surface is presumed to be an indirect effect of liming. It is highly unlikely that the lime applied had moved much beyond the initial zone of incorporation during the experimental period due its slow movement (Li et al. 2010). Indeed, there had been some lime movement over 6 years following lime application (Fig. 1), but no deeper than 0.20 m. Therefore, the explanation for the result we observed is likely to be that lime enhanced the growth of the perennial species which were then able to extract more water from the subsoil.
Further research is required to better understand the mechanisms responsible for this phenomenon. It is possible that the increased pasture growth observed during our study resulted in an increased evapotranspiration demand enabling increased water extraction from the soil profile where lime was applied. In addition, lime would be expected to increase root growth of lucerne (Hayes et al. 2011, 2012a) and phalaris (Song et al. 2014) in the soil surface by ameliorating toxicities associated with Al and Mn which may have enhanced the capacity of the perennial root systems to find more favourable channels through the subsurface and better explore the soil volume for water, as postulated by Culvenor et al. (2011). White et al. (2000) showed that lime reduced deep drainage under perennial pastures on a duplex soil, but had no effect on lateral subsurface flow. This infers that lime increased the water extraction of perennial pastures, similar to the current study. It is also possible that improved root growth of perennial species with liming is associated with improved Ca, Mo and P nutrition (Haynes 1982; Helyar et al. 1990). However, we found little evidence of changed concentrations of these nutrients due to lime (Table 4) at the conclusion of the experiment, and little evidence that any macro or micro nutrient was limiting for pasture growth, particularly in the dry years where pasture growth was only modest. Regardless of the mechanism responsible, the presumed access to increased levels of soil water has major implications for increasing production efficiencies in crop/livestock enterprises and for increasing pasture resilience under drought.
An increased soil water deficit is known to reduce the risk of soil degradation attributable to deep drainage (Ridley et al. 1999; Dunin and Passioura 2006). Not only were significant increases in productivity observed due to lime, but our study suggests that the environmental sustainability of the production system would also be enhanced where lime is used in conjunction with deep-rooted perennials such as phalaris. This supports previous findings which showed perennial pastures better able to reduce acidification by taking up more N (Ridley et al. 1999, 2001). We would not expect lime to increase the dry soil deficit unless it was used in conjunction with deep-rooted perennial species on a responsive soil.
There was no evidence in the current study that phalaris and lucerne increased soil pH at the soil surface as has previously been reported (Ridley et al. 2002; Dear et al. 2009). This may be due to the relatively dry seasonal conditions experienced where deep drainage under all treatments would have been minimal, or because previous studies were comparing soil under perennial species with soil under annual pastures which were not included in the current experiment. Nevertheless, given the observed increase in dry soil deficit, it is reasonable to expect that phalaris and lucerne used in conjunction with lime on acidic soils would further reduce soil profile acidification rates over what has been previously reported.
The response to acidic soil conditions at this experimental site is further complicated by interactions with climatic conditions. Establishment of perennial species following a late-August sowing was undoubtedly assisted by above average rainfall in the November, December and February following sowing, followed by a very favourable spring in year 2 despite a poor autumn in that year (Table 1). The largely favourable seasonal conditions experienced early in the experiment may have facilitated the establishment of all perennial species irrespective of Al tolerance as suggested by Culvenor et al. (2011). Having an established root system in place prior to the three consecutive dry years undoubtedly enhanced the capacity of the perennial species to utilise soil water from deep in the profile, particularly for the two species renowned for their deep-rooting habit, lucerne (Black et al. 1981) and phalaris (McWilliam and Kramer 1968).
Perennial species
Phalaris and lucerne have both previously been described as being relatively sensitive to soil acidity justifying substantial research programs devoted to developing genotypes of both species more tolerant of soil acidity, in particular Al toxicity (Devine et al. 1976; Oram et al. 1990; Culvenor et al. 2011; Hayes et al. 2011). The perceived relative sensitivity of both species has led to advise farmers to avoid sowing these species on acidic soils (e.g., Anon. 2004). However, our study has demonstrated the value of sowing these two Al sensitive species on this acidic soil. Indeed, due to the sustained period of drought experienced at this site during the 5 year experimental period, had we not included these ‘sensitive’ species in the current experiment, by year 3 there would have been very little left to measure by way of perennial pastures (see Table 2).
The current study is the second example of which the present authors are aware where the more ‘acid-sensitive’ phalaris persisted better than cocksfoot, a species renowned for its superior adaptation to acid soils (Lolicato and Rumball 1994). Hayes et al. (2010b) conducted an experiment on an acutely acidic soil at Goulburn, New South Wales, Australia, and found that phalaris was the only species to persist beyond the first summer in the absence of lime. In both the previous and current experiments, the drought tolerance of phalaris undoubtedly contributed to its superior persistence even where soil acidity was not ameliorated by lime.
Culvenor et al. (2011) challenged the perceived thresholds at which phalaris might be expected to establish and persist in a field environment contending that contemporary recommendations are too conservative for phalaris with all cultivars expected to tolerate levels of Al saturation ≤20 % of the cation exchange capacity and pHCa of 4.2–4.5. Moreover, newer phalaris cultivars with enhanced tolerance of Al toxicity established more reliably than traditional cultivars under drought conditions. The current study using the first of the phalaris cultivars with improved Al tolerance, cv. Landmaster (Oram 1996), showed adequate phalaris persistence at this site with pHCa as low as 4.0 and up to 23 % exchangeable Al in the surface 0.20 m, but production and basal frequency were increased with lime. This collective experience of superior phalaris performance has prompted a revision of agronomic recommendations, suggesting that phalaris may be a suitable choice of species for acidic soils, particularly in environments prone to periodic drought (Hayes et al. 2015). However, as demonstrated in the current study, where phalaris is grown on acidic soils it should follow prior lime application. Applying lime is expected to enhance the reliability of phalaris establishment (such as was shown in Hayes et al. 2010b), and improve phalaris production as evidenced by the 30 % increase in cumulative biomass in the current study which is broadly consistent with responses observed in phalaris previously (Ridley and Coventry 1992; Li et al. 2006b).
The superior persistence of lucerne in the current experiment was less expected, and there are few examples in the scientific literature of lucerne persisting better than reputedly more acid-tolerant species such as phalaris and cocksfoot, particularly in the absence of lime. In the current experiment soil acidity and toxicities of Mn and Al were largely confined to the surface depths. In addition, favourable climatic conditions at establishment as discussed previously and a relatively deep soil profile (~2 m) enhanced the performance of lucerne in this experiment. However, the fact that lime increased lucerne biomass by 150 % indicates that lucerne growth was substantially constrained by acid soil-related factors, more so than the remaining species. Large responses to lime have previously been reported in lucerne when grown on an acid soil (Horsnell 1985) and interestingly, in the current study cumulative lucerne biomass did not surpass that of other species until the dry sequence of years later in the experiment (Table 3). This indicates that the relative drought tolerance of lucerne compared to the other species was an important factor influencing its overall performance. Unlike phalaris there still exists no cultivar of lucerne with enhanced tolerance of low pH or toxicities of Al or Mn (Bouton 1996; Charman et al. 2008) which would seem a worthwhile initiative to further improve the reliability of lucerne on acid soils as has been achieved with phalaris (Culvenor et al. 2011). Further research is also required to more explicitly define the characteristics of an acid soil where lucerne might reliably be expected to out-perform more tolerant perennial pasture species, as was observed in the current experiment.
Soil acidity, Mn and Al toxicity
Previous experimentation at this site had demonstrated evidence of Mn toxicity affecting lucerne growth and of increased pasture productivity due to lime. The concentration of Mn in lucerne herbage in unlimed plots was higher (918 mg/kg) in the year following lime application (Hayes et al. 2008) compared to an adjacent experiment sampled in 2008 where values ranged from 77 to 430 mg/kg in the absence of lime (Hayes et al. 2012a). The concentration of Mn in lucerne herbage in the current study (136–199 mg/kg) falls within the latter range and is substantially lower than values reported initially. A detailed study of Mn concentrations in soil from immediately adjacent to the present experiment showed that levels of easily reducible Mn declined sharply below 0.20 m (Hayes et al. 2012a), consistent with levels of exchangeable Mn depicted in Fig. 1. This indicates that the source of Mn toxicity at this site was largely in the surface 0.20 m. Given the known negative correlation between pH and Mn2+ in soil (Hayes et al. 2012a) and the fact that soil pH continued to decline over the life of the experiment from the initial lime application in 2002 (Table 5), it might have been expected that Mn concentrations in lucerne herbage would increase with time rather than decrease as was observed. The explanation for this is likely associated with the growth stage of lucerne and the relatively low levels of easily reducible Mn below 0.20 m. In 2002 when lucerne was establishing, it is likely that a large proportion of its root system was exposed to the toxic levels of Mn in the surface 0.20 m, resulting in high concentrations of Mn in the herbage. By contrast, in 2008 a greater proportion of the plant root system was below the toxic 0.20 m zone increasing its capacity to reduce exposure to Mn toxicity in the soil.
The positive response in pasture growth to lime may have been partly associated with amelioration of Mn toxicity, but is also likely due to increased pH and the amelioration of Al toxicity. It is not possible in a field experiment such as this to fully separate the factors and attribute response to lime as being due to the amelioration of one or other of the known acid soil factors. In the absence of significant differences in nutrient concentrations between lime+ and lime- treatments (Table 4), unequivocal proof of nutrient-related lime response is still lacking. Furthermore, concentration of nutrients in lucerne and cocksfoot appeared to be within previously established thresholds (Reuter and Robinson 1997) except for Ca and Mg in cocksfoot particularly, which were lower than the references whereas Mn was higher than the reference. It is also unlikely that the response to lime observed was entirely due to the amelioration of Mn toxicity, given that there was a strong response to lime in the grasses and species such as phalaris are known to be highly tolerant of Mn toxicity (Culvenor 1985). Low pH (Edmeades et al. 1991) and Al toxicity also likely reduced the growth of phalaris (Culvenor et al. 2011) and lucerne (Hayes et al. 2011). Similar to Mn toxicity, with pHCa of 4.7 and negligible levels of exchangeable Al in the 0.20–0.40 m depth, there is very little evidence that pH or Al below 0.40 m constrained production, even in the most sensitive species. The shallow depth of acidity at the current site undoubtedly enhanced the capacity of the acid soil-sensitive species to persist even in the absence of lime. For example, Culvenor et al. (2004) found that phalaris persistence was excellent on a soil with an average pH of 4.0, AlCa (CaCl2 extractable Al) of 16 mg/kg and Al saturation of 41 % were restricted to the upper 0.30 m with pH increasing to 4.5–5.2 from 0.30 to 0.60 m depth. However, persistence was lower on a soil with less available Al in the upper 0.30 m but in which strong acidity extended to at least 0.60 m (pH 4.0, AlCa 16 mg/kg, Al saturation 40 %). The current site was most similar to the least acidic site, Bookham, used by Culvenor et al. (2011) who evaluated the response of phalaris cultivars to Al toxicity, and so the relative stress imposed on pasture species at the current site might be considered to be only moderate. This is further supported by fact that Mn concentrations in mature lucerne herbage (199 mg/kg) at the end of the experiment were only marginally above the critical threshold of 180 mg/kg set by De Marco et al. (1995) which assessed lucerne seedlings < 40 days old. More research is required to confirm critical thresholds of Mn toxicity in mature lucerne plants.
Conclusion
Surface applied lime used in conjunction with deep rooted perennial pasture species increased the pasture productivity, improved persistence of perennial species and utilised more soil water deep in the profile. Lucerne and phalaris, previously reported to be the two species most sensitive to acid soil-related toxicities, were shown to be the most productive and persistent species at this acid soil site despite a prolonged period of unfavourable seasonal conditions. Sowing lucerne and phalaris in conjunction with lime on sufficiently deep soils with moderate levels of acidity will enhance the productivity and resilience of pastures in environments prone to drought. Lime used in conjunction with these deep-rooted perennial species will also likely help mitigate the risk of further soil degradation attributable to deep drainage due to the increased capacity to extract water from the subsoil. In the case of phalaris in particular we concur with previous studies that existing recommendations may need to be revised to reflect the fact that phalaris is adapted to acidic or high-Al soil environments. Further breeding and selection of lucerne for enhanced acid soil tolerance would be a worthwhile endeavour to further improve the adaptation of this species to acid soil environments.
References
Anon (2004) Phalaris Phalaris aquatica. Agnote DPI-284, third edition. NSW Department of Primary Industries. http://www.dpi.nsw.gov.au/agriculture/pastures/pastures-and-rangelands/species-varieties/factsheets/phalaris
Belesky DP, Turner KE, Fedders JM, Ruckle JM (2001) Mineral composition of swards containing forage chicory. Agron J 93:468–475
Black AL, Brown PL, Halvorson AD, Siddoway FH (1981) Dryland cropping strategies for efficient water-use to control saline seeps in the Northern Great Plains, U.S.A. Agric Water Manag 4(1–3):295–311
Bolland MDA, Allen DG, Rengel Z (2002) Response of annual pastures to applications of limestone in the high rainfall areas of south-western Australia. Aust J Exp Agric 42(7):925–937
Bouton JH (1996) Screening the alfalfa core collection for acid soil tolerance. Crop Sci 36:198–200
Bureau of Meteorology (2014) Climate data online (Australia). Walla Walla Post Office; station 074117. http://www.bom.gov.au/climate/data/. Accessed 9 July 2014.
Charman N, Ballard RA, Humphries AW, Auricht GC (2008) Improving lucerne nodulation at low pH: contribution of rhizobial and plant genotype to the nodulation of lucerne seedlings growing in solution culture at pH 5. Aust J Exp Agric 48(4):512–517
Culvenor RA (1985) Tolerance of Phalaris aquatica L. lines and some other agricultural species to excess manganese, and the effect of aluminium on manganese tolerance in P. aquatica. Aust J Agric Res 36(5):695–708
Culvenor RA, Oram RN, de St F, Groth C (1986) Variation in tolerance in Phalaris aquatica L. and a related species to aluminium in nutrient solution and soil. Aust J Agric Res 37(4):383–395
Culvenor RA, Wood JT, Avery AL, Dempsey W, McDonald SE, Ronnfeldt G, Veness PE (2004) Multi-site evaluation on acid soils of a Phalaris aquatica × P. arundinacea × P. aquatica backcross population bred for acid soil tolerance. Aust J Agric Res 55(6):681–692
Culvenor RA, McDonald SE, Veness PE, Watson D, Dempsey W (2011) The effect of improved aluminium tolerance on establishment of the perennial grass, phalaris, on strongly acid soils in the field and its relation to seasonal rainfall. Crop Pasture Sci 62(5):413–426
De Marco D, Li CB, Randall PJ (1995) Manganese toxicity in Trifolium balansae, T. resupinatum, T. subterraneum, Medicago murex, M. polymorpha, M. sativa, Lotus pedunculatus, and Ornithopus compressus: relative tolerance and critical toxicity concentrations. Aust J Exp Agric 35(3):367–374
Dear BS, Ewing MA (2008) The search for new pasture plants to achieve more sustainable production systems in southern Australia. Aust J Exp Agric 48:387–396
Dear BS, Peoples MB, Cocks PS, Swan AD, Smith AB (1999) Nitrogen fixation by subterranean clover (Trifolium subterraneum L.) growing in pure culture and in mixtures with varying densities of lucerne (Medicago sativa L.) or phalaris (Phalaris aquatica L.). Aust J Agric Res 50(6):1047–1058
Dear BS, Virgona JM, Sandral GA, Swan AD, Morris S (2009) Changes in soil mineral nitrogen, nitrogen leached, and surface pH under annual and perennial pasture species. Crop Pasture Sci 60(10):975–986
Devine TE, Foy CD, Fleming AL, Hanson CH, Campbell TA, McMurtrey JE, Schwartz JW (1976) Development of alfalfa strains with differential tolerance to aluminum toxicity. Plant Soil 44:73–79
Dunin F, Passioura J (2006) Prologue: amending agricultural water use to maintain production while affording environmental protection through control of outflow. Aust J Agric Res 57(3):251–255
Edmeades DC, Blamey FPC, Asher CJ, Edwards DG (1991) Effects of pH and aluminium on the growth of temperate pasture species. I. Temperate grasses and legumes supplied with inorganic nitrogen. Aust J Agric Res 42(3):559–569
Gillman G, Sumpter E (1986) Modification to the compulsive exchange method for measuring exchange characteristics of soils. Soil Res 24(1):61–66
Hayes RC, Dear BS, Orchard BA, Peoples MB, Eberbach PL (2008) Response of subterranean clover, balansa clover, and gland clover to lime when grown in mixtures on an acid soil. Aust J Agric Res 59(9):824–835
Hayes RC, Dear BS, Li GD, Virgona JM, Conyers MK, Hackney BF, Tidd J (2010a) Perennial pastures for recharge control in temperate drought-prone environments. Part 1: productivity, persistence and herbage quality of key species. N Z J Agric Res 53(4):283–302
Hayes RC, Li GD, Dear BS, Conyers MK, Virgona JM (2010b) Phalaris and lime – improving productivity on an acidic soil in a drought-prone ‘high-rainfall’ environment. In: Dove H, Culvenor RA (eds) Food Security from Sustainable Agriculture, Proceedings of 15th Agronomy Conference. Australian Society of Agronomy, Lincoln
Hayes RC, Li GD, Dear BS, Conyers MK, Virgona JM, Tidd J (2010c) Perennial pastures for recharge control in temperate drought-prone environments. Part 2: soil drying capacity of key species. N Z J Agric Res 53(4):327–345
Hayes RC, Scott BJ, Dear BS, Li GD, Auricht GC (2011) Seedling validation of acid soil tolerance of lucerne populations selected in solution culture high in aluminium. Crop Pasture Sci 62(9):803–811
Hayes RC, Conyers MK, Li GD, Poile GJ, Price A, McVittie BJ, Gardner MJ, Sandral GA, McCormick JI (2012a) Spatial and temporal variation in soil Mn2+ concentrations and the impact of manganese toxicity on lucerne and subterranean clover seedlings. Crop Pasture Sci 63(9):875–885
Hayes RC, Li GD, Hackney BF (2012b) Perennial pasture species for the mixed farming zone of southern NSW - we don’t have many options. In: Harris C, Lodge G, Waters C (eds) Driving your landscape to success - Managing a grazing business for profit in the agricultural landscape, Proceedings of the 27th annual conference of The Grassland Society of NSW Inc. The Grassland Society of NSW Inc., Orange, Wagga Wagga, pp 92–100
Hayes RC, Li GD, Culvenor RA (2015) Changed recommendations for the use of phalaris on acid soils. In: Building Productive, Diverse and Sustainable Landscapes. Proceedings of the 17th Australian Society of Agronomy Conference, 20–24 September, Hobart, Australia. www.agronomy2015.com.au
Haynes RJ (1982) Effects of liming on phosphate availability in acid soils. Plant Soil 68:289–308
Helyar KR (1976) Nitrogen cycling and soil acidification. J Aust Inst Agric Sci 42:217–221
Helyar KR, Cregan PD, Godyn DL (1990) Soil acidity in New South Wales - current pH values and estimates of acidification rates. Aust J Soil Res 28(4):523–537
Hill BD (1985) Yields of perennial grasses growing in mixtures with annual species and the effect of perennials on total yields. Aust J Exp Agric 25(4):840–849
Hoffmann JD, Eberbach PL, Virgona JM, Katupitiya A (2003) Conservative water use by lucerne, 11th Australian Agronomy Conference, “Solutions for a new environment”. Australian Society of Agronomy, Geelong
Horsnell LJ (1985) The growth of improved pastures on acid soils. 2. The effect of soil incorporation of lime and phosphorus on the growth of subterranean clover and lucerne pastures and on their response to topdressing. Aust J Exp Agric 25(1):157–163
Isbell RF (1996) The Australian Soil Classification. CSIRO, Melbourne
Jones RM, Hargreaves JNG (1979) Improvements to the dry-weight-rank method for measuring botanical composition. Grass Forage Sci 34:181–189
Lattimore MA, McCormick L (2012) Pasture varieties used in New South Wales 2012–13. New South Wales: Published jointly by NSW Department of Primary Industries and the Grassland Society of NSW Inc
Li GD, Helyar KR, Conyers MK, Castleman LJC, Fisher RP, Poile GJ, Lisle CJ, Cullis BR, Cregan PD (2006a) Pasture and sheep responses to lime application in a grazing experiment in a high-rainfall area, south-eastern Australia. II. Liveweight gain and wool production. Aust J Agric Res 57(10):1057–1066
Li GD, Helyar KR, Welham SJ, Conyers MK, Castleman LJC, Fisher RP, Evans CM, Cullis BR, Cregan PD (2006b) Pasture and sheep responses to lime application in a grazing experiment in a high-rainfall area, south-eastern Australia. I. Pasture production. Aust J Agric Res 57(10):1045–1055
Li GD, Lodge GM, Moore GA, Craig AD, Dear BS, Boschma SP, Albertson TO, Miller SM, Harden S, Hayes RC, Hughes SJ, Snowball R, Smith AB, Cullis BR (2008) Evaluation of perennial pasture legumes and herbs to identify species with high herbage production and persistence in mixed farming zones in southern Australia. Aust J Exp Agric 48:449–466
Li G, Conyers M, Cullis B (2010) Long-term liming ameliorates subsoil acidity in high rainfall zone in south-eastern Australia. In: Gilkes RJ, Prakongkep N (eds) 19th World Congress of Soil Science; Soil Solutions for a Changing World. International Union of Soil Science, Brisbane, pp 136–139
Lolicato S, Rumball W (1994) Past and present improvement of cocksfoot (Dactylis glomerata L.) in Australia and New Zealand. N Z J Agric Res 37:379–390
McLachlan KD (1980) Nutrient problems in sown pasture on an acid soil. 2. Role of lime and superphosphate. Aust J Exp Agric Anim Hus 20(106):568–575
McWilliam JR, Kramer PJ (1968) The nature of the perennial response in Mediterranean grasses. I. Water relations and summer survival in Phalaris. Aust J Agric Res 19(3):381–395
Oram RN (1996) Register of Australian herbage plant cultivars. A. Grasses. 3. Phalaris (a) Phalaris aquatica L. (phalaris) cv. Landmaster. Aust J Exp Agric 36:913–914
Oram RN, Ridley AM, Hill MJ, Hunter J, Hedges DA, Standen RL, Bennison L (1990) Improving the tolerance of Phalaris aquatica L. to soil acidity by introgression genes from P. arundinacea L. Aust J Agric Res 41:657–668
Oram RN, Ferreira V, Culvenor RA, Hopkins AA, Stewart A (2009) The first century of Phalaris aquatica L. cultivation and genetic improvement: a review. Crop Pasture Sci 60(1):1–15
Peoples MB, Baldock JA (2001) Nitrogen dynamics of pastures: nitrogen fixation inputs, the impact of legumes on soil nitrogen fertility, and the contributions of fixed nitrogen to Australian farming systems. Aust J Exp Agric 41(3):327–346
Pinkerton A, Simpson JR (1986) Responses of some crop plants to correction of subsoil acidity. Aust J Exp Agric 26(1):107–113
Reuter DJ, Robinson JB (1997) Plant Analysis an Interpretation Manual, 2nd edn. CSIRO Publishing, Collingwood
Ridley AM, Coventry DR (1992) Yield responses to lime of phalaris, cocksfoot, and annual pastures in north-eastern Victoria. Aust J Exp Agric 32(8):1061–1068
Ridley AM, Simpson RJ (1994) Seasonal development of roots under perennial and annual grass pastures. Aust J Agric Res 45(5):1077–1087
Ridley AM, Simpson RJ, White RE (1999) Nitrate leaching under phalaris, cocksfoot, and annual ryegrass pastures and implications for soil acidification. Aust J Agric Res 50(1):55–64
Ridley AM, White RE, Helyar KR, Morrison GR, Heng LK, Fisher R (2001) Nitrate leaching loss under annual and perennial pastures with and without lime on a duplex (texture contrast) soil in humid southeastern Australia. Eur J Soil Sci 52:237–252
Ridley AM, Avery AL, Oram RN, Hunter J, Shovelton JB, Mahoney GP, Muller WJ (2002) Long-term persistence of aluminium-tolerant and sensitive Phalaris lines on acidic soils and associated changes in soil acidity. Aust J Exp Agric 42(8):1033–1042
Sandral GA, Dear BS, Virgona JM, Swan AD, Orchard BA (2006) Changes in soil water content under annual- and perennial-based pasture systems in the wheatbelt of southern New South Wales. Aust J Agric Res 57(3):321–333
Scott BJ, Conyers MK, Poile GJ, Cullis BR (1997) Subsurface acidity and liming affect yield of cereals. Aust J Agric Res 48(6):843–854
Scott BJ, Ridley AM, Conyers MK (2000) Management of soil acidity in long-term pastures of south-eastern Australia: a review. Aust J Exp Agric 40(8):1173–1198
Scott BJ, Fenton IG, Fanning AG, Schumann WG, Castleman LJC (2007) Surface soil acidity and fertility in the eastern Riverina and Western Slopes of southern New South Wales. Aust J Exp Agric 47(8):949–964
Singer MJ, Munns DN (1996) Acidity and salinity. In: Singer MJ, Munns DN (eds) Soils an Introduction Third Edition. Prentice Hall, Upper Saddle River, pp 267–298
Song Y, Hayes RC, Sandral GA, McVittie BJ, Zheng W, Li GD (2014) Aluminium tolerance of 20 temperate grasses and 4 temperate legumes in solution culture. In: Patti A, Tang C, Wong V (eds) Proceedings of the Soil Science Australia National Soil Science Conference: Securing Australia’s soils - for profitable industries and healthy landscapes. Australian Society of Soil Science, Melbourne, http://www.soilscience2014.com/proceedings/Guangdi_Li.pdf
t’ Mannetje L, Haydock KP (1963) The dry-weight-rank method for the botanical analysis of pasture. J Br Grassl Soc 18:268–275
Ward PR (2006) Predicting the impact of perennial phases on average leakage from farming systems in south-western Australia. Aust J Agric Res 57(3):269–280
White RE, Helyar KR, Ridley AM, Chen D, Heng LK, Evans J, Fisher R, Hirth JR, Mele PM, Morrison GR, Cresswell HP, Paydar Z, Dunin FX, Dove H, Simpson RJ (2000) Soil factors affecting the sustainability and productivity of perennial and annual pastures in the high rainfall zone of south-eastern Australia. Aust J Exp Agric 40(2):267–283
Wolfe EC, FitzGerald RD, Hall DG, Southwood OR (1980) Beef production from lucerne and subterranean clover pastures. 1. The effects of pasture, stocking rate and supplementary feeding. Aust J Exp Agric Anim Hus 20(107):678–687
Acknowledgments
The authors gratefully acknowledge Evan Moll and family, ‘Elderslie’ Gerogery, for their enthusiastic support of this research and access to land for experimentation. Funding for this research was provided by the NSW Department of Primary Industries and Australian Wool Innovation. The senior author presented some of the results from this experiment in his Master Thesis submitted to Charles Sturt University in 2009. The insightful comments of two anonymous referees greatly improved the quality of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Ismail Cakmak.
Rights and permissions
About this article

Cite this article
Hayes, R.C., Li, G.D., Conyers, M.K. et al. Lime increases productivity and the capacity of lucerne (Medicago sativa L.) and phalaris (Phalaris aquatica L.) to utilise stored soil water on an acidic soil in south-eastern Australia. Plant Soil 400, 29–43 (2016). https://doi.org/10.1007/s11104-015-2706-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2706-z