
Abstract
Solanum lycopersicum auxin response factor 10 (SlARF10) is post-transcriptionally regulated by Sl-miR160. Overexpression of a Sl-miR160-resistant SlARF10 (mSlARF10) resulted in narrower leaflet blades with larger stomata but lower densities. 35S:mSlARF10-6 plants with narrower excised leaves had greater water loss, which was in contrast to the wild type (WT). Further analysis revealed that the actual water loss was not consistent with the calculated stomatal water loss in 35S:mSlARF10-6 and the WT under the dehydration treatment, indicating that there is a difference in hydraulic conductance. Pretreatment with abscisic acid (ABA) and HgCl2 confirmed higher hydraulic conductance in 35S:mSlARF10, which is related to the larger stomatal size and higher activity of aquaporins (AQPs). Under ABA treatment, 35S:mSlARF10-6 showed greater sensitivity, and the stomata closed rapidly. Screening by RNA sequencing revealed that five AQP-related genes, fourteen ABA biosynthesis/signal genes and three stomatal development genes were significantly altered in 35S:mSlARF10-6 plants, and this result was verified by qRT-PCR. The promoter analysis showed that upregulated AQPs contain AuxRE and ABRE, implying that these elements may be responsible for the high expression levels of AQPs in 35S:mSlARF10-6. The three most upregulated AQPs (SlTIP1-1-like, SlPIP2;4 and SlNIP-type-like) were chosen to confirm AuxRE and ABRE function. Promoters transient expression demonstrated that the SlPIP2;4 and SlNIP-type-like AuxREs and SlPIP2;4 and SlTIP1-1-like ABREs could significantly enhance the expression of the GUS reporter in 35S:mSlARF10-6, confirming that AuxRE and ABRE may be the main factors inducing the expression of AQPs. Additionally, two upregulated transcription factors in 35S:mSlARF10-6, SlARF10 and SlABI5-like were shown to directly bind to those elements in an electromobility shift assay and a yeast one-hybrid assay. Furthermore, transient expression of down-regulated ARF10 or up-regulated ABI5 in tomato leaves demonstrated that ARF10 is the direct factor for inducing the water loss in 35S:mSlARF10-6. Here, we show that although SlARF10 increased the ABA synthesis/signal response by regulating stomatal aperture to mitigate water loss, SlARF10 also influenced stomatal development and AQP expression to affect water transport, and both act cooperatively to control the loss of leaf water in tomato. Therefore, this study uncovers a previously unrecognized leaf water loss regulatory factor and a network for coordinating auxin and ABA signalling in this important process. In an evolutionary context, miR160 regulates ARF10 to maintain the water balance in the leaf, thus ensuring normal plant development and environmental adaptation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Both the growth and geographic distribution of plants are greatly affected by the environment, but plants also undergo favourable genetic and phenotypic alterations in response to abiotic and biotic changes in the environment. Of the abiotic stresses, the supply of water plays a key role in determining plant growth, and a range of physiological and biochemical changes occur in plants to enable them to survive water stress (Shinozaki and Yamaguchi-Shinozaki 2007). These responses include stomatal closure, repression of cell growth and photosynthesis, and activation of respiration. Through long-term natural selection, water loss must be minimized for plant growth to adapt to drought stress, such as through thick cuticles, concave stomata, rich leaf hairs and narrow blade surface areas (Mulroy and Rundel 1977).
The leaf is the key organ involved in the evaporation of water from plants, and two factors affect the loss of water through the leaves. The stoma is one of the major factors influencing leaf water loss, and stomatal closure regulates water loss by transpiration. The stomatal conductance to water vapour in the mature leaf is predominantly determined by stomatal size and density (Franks et al. 2009), and stomatal size is important because it uniquely defines the upper limit of density (Franks and Beerling 2009). A similar measure of stomatal size, when multiplied by density, was found to be correlated with leaf hydraulic conductance (Sack et al. 2003). The cleavage of growth-regulating factor (GRF) target genes by miR396, which is involved in Arabidopsis leaf development, can affect stomatal density and thus increase endurance under drought (Liu et al. 2009).
On its way through the leaf from the xylem to the stomata, water can either move through cell walls or pass from cell to cell across different tissues. Although both pathways are likely used to some degree, there is increasing evidence that living cells contribute substantially to the overall leaf hydraulic conductance (K leaf ). Transcellular water flow is facilitated and regulated by water channels in the membranes, named aquaporins (AQPs) (Heinen et al. 2009), and they facilitate the rapid, passive exchange of water across cell membranes and are responsible for up to 95 % of the water permeability of plasma membranes (Henzler et al. 2004). The AQPs are particularly important for maintaining the large amounts of water being transported because they respond differently to water stress (Hachez et al. 2006b; Heinen et al. 2009). Several studies have shown that water stress can alter leaf AQP expression and activity, which modifies plant adaptations to water stress (Aharon et al. 2003; Lian et al. 2006).
Abscisic acid (ABA) is a plant stress hormone and one of the most important signalling molecules in the regulation of leaf water loss (Peleg and Blumwald 2011). Water stress increases the ABA plant hormone by strongly inducing the expression of a 9-cis-epoxycarotenoid dioxygenase (NCED) gene, which is a key enzyme in ABA biosynthesis (Iuchi et al. 2001). Moreover, carotenoids are precursor molecules for ABA, and the formation of GGPP is a key step in the carotenoids biosynthetic pathway. This step is catalysed by geranylgeranyl pyrophosphate synthase (GGPS). Induction of carotenoid biosynthesis contributes to ABA production upon salt stress in Arabidopsis (Ruiz-Sola et al. 2014; Li et al. 2008). ABA curtails transpirational water loss by promoting stomatal closure and inhibiting stomatal opening (Acharya and Assmann 2009); moreover, the application of the hormone could decrease the relative loss of water content in plants through decreased leaf transpiration (Raphael and Maarten 2001). ABA controls leaf water deficits by both changing the stomatal apertures (for example, the stomata of ABA-treated plants exhibit more complete closure than control plants) and by modifying the hydraulic conductivity in plants (Franks and Farquhar 2001; Hose et al. 2000; Parent et al. 2009; Thompson et al. 2007). Under water stress, stomatal closure may be mediated by ABA or the hydraulic conductivity of the xylem vessels (Brodribb and Holbrook 2003; Cochard et al. 2002). Drought-induced ABA diminishes the osmotic water permeability of bundle-sheath cells by downregulating the activity of their AQPs (Shatil-Cohen et al. 2011), and the ABA signal is also reportedly involved in water stress. ABA INSENSITIVE 1 (ABI1) and ABI2 are also crucial for ABA-mediated stomatal regulation (Gosti et al. 1999; Merlot et al. 2001), and the overexpression of ABRE-BINDING FACTOR (ABF3) or AREB2/ABF4 caused ABA hypersensitivity, reduced the transpiration rate and enhanced drought tolerance in transgenics (Kang et al. 2002). Over-expression of ABI5 could enhance plant resistance to drought stress. ABRE is known to mediate gene regulation by ABI/ABF, and AuxRE is known to mediate gene regulation by ARFs (Fricke et al. 2013). High ABA content, or a strong ABA signal, is thought to be positively correlated with plant resistance to water stress.
Potential cross-talk between auxin and ABA has been suggested by a number of studies in a variety of plants. Auxin response factor 2 (ARF2) was induced by ABA, and the arf2 mutants displayed enhanced ABA sensitivity during seed germination and primary root growth, suggesting that ARF2 represses the ABA signalling pathway (Wang et al. 2011). Auxin controls seed dormancy through the stimulation of abscisic acid signalling by ARF10 and ARF16, which induces ARF-mediated ABI3 activation in Arabidopsis (Liu et al. 2013). The differential expression of drought-related proteins, which is ABA-dependent, and the related drought-adaptive responses in auxin mutant AXR2 and/or AXR1 indicate cross-talk between auxin- and ABA-signalling in Arabidopsis (Bianchi et al. 2002). Auxin may participate in the positive regulation of drought stress resistance through the regulation of root architecture and the expression of ABA-responsive genes (Shi et al. 2014).
Recently, miR160 has been reported to play a major role in the ABA response in plants by mediating the accumulation of ARF10, 16 and 17 at the mRNA level. The negative regulation of ARF10 by miR160 affects ABA sensitivity and may play a role in auxin-ABA cross-talk (Liu et al. 2007). However, the role of ARF10/miR160 in ABA dependence and its direct role in the adaptation to water stress are still unclear.
The Sl-miR160a-resistant (mSlARF10) transgenic lines offer a good system to study the relationship between water loss in tomato leaves and the regulation of ARF10 auxin–ABA crosstalk by miRNA160. In this study, we report that 35S:mSlARF10-6 transgenic plants developed narrow leaflet blades, a theoretically highly efficient water-saving leaf type, but exhibited rapid water loss under the water stress treatment. Further, this study revealed that the density and size of individual stoma are altered in 35S:mSlARF10-6, and its water loss capacity may be determined by the stomatal factor. However, with prolonged treatment time, water loss is significantly not in accordance with the stomatal factor because mSlARF10 accumulates greater ABA content and sensitivity under water stress, which leads to more sensitive stomata closure. The use of a special AQP inhibitor in HgCl2-pretreated leaves supported our hypothesis that AQP may play an important role in the overexpression of mSlARF10 in leaf water loss. Real-time-PCR and RNA sequencing revealed that mSlARF10 overexpression could enhance expression of ABA synthesis gene, the ABA signal factor and AQP expression, and cis-element analysis indicated upregulated AQPs promoters contain ABRE and AuxRE. Transient expression analysis indicated that AuxRE majorly contributes to enhanced AQP expression, while ABRE can simultaneously enhance or depress its expression. The yeast-one hybrid assay and EMSA determined that the ARF10 could directly bind to the relevant site of the promoter sequence during the upregulation of AQP. The upregulated ABI5 in 35S:mSlARF10 could also directly bind to the upregulated AQP, which contains ABRE. To verify whether SlARF10 enhances water loss by boosting ABI5 expression, transient expression of down-regulated ARF10 or up-regulated ABI5 in tomato leaves was carried out. The results demonstrated that ‘ARF1’ could descrease the water loss rate and the expression of most AQPs, while ABI5+ could significantly decrease the water loss rate at the ealier stage of water stress, although the expression of some AQPs was indeed enhanced. Here, we show that although SlARF10 increased the ABA synthesis/signal response by regulating stomatal aperture to mitigate water loss, SlARF10 also directly influenced stomatal development and AQP expression to affect water transport, and both actions cooperated to control the loss of water in tomato leaves. In an evolutionary context, miR160 regulates ARF10 to maintain the water balance in the leaf, thus ensuring normal plant development and enviormental adaptation.
Materials and methods
Plant materials and growth conditions
Tomato cv. MP1 plants (Barg et al. 1997) were grown under standard greenhouse conditions at 24 °C/18 °C (day/night) with a photoperiod of 16 h light/8 h dark. The 35S:mSlARF10-6 transgenic plants were supplied by Arazi (Hendelman et al. 2012). All of the study plants were grown under the same conditions; whole plants were grown and watered daily for 6 weeks.The samples were all obtained from the primary leaflets of the fifth leaf in each plant.
Measurement of water loss
To measure water loss, each of the 35 fully expanded, true leaves of the 6–8-week-old MP1 and 35S:mSlARF10-6 plants were detached and weighed at different times (0, 0.5, 1, 2, 6, 12 and 24 h) to determine the rate of water loss. Five leaves from each type of plant were selected at each of the time points, which were determined by consulting the literature (Kahn et al. 1993). For time course studies, leaves were weighed, and then placed on the bench to undergo dehydration for periods of 0–24 h, measured from the start of wilting. Leaf water loss ratio as a percentage of (fresh weight-dry weight)/fresh weight was calculated at the time of leaf sampling. At each time point, some leaves were frozen in liquid nitrogen and stored at −80 °C.
Leaf hydraulic conductance measurements
Leaf hydraulic conductance was measured using the evaporative flux method for three MP1 and three 35S:mSlARF10-6 plants. A filtered (0.2 μM) 20 mM KCl + 1 mM CaCl2 solution (subsequently referred to as ‘artificial xylem sap’, AXS) was used for these measurements. Flow rate through leaves was measured with a balance (model CP 224S, Sartorius, Gottingen, Germany), which logged data to a computer every 30 s. The air was adequately stirred by a fan as explained by Sack and Scoffoni (2012). Leaves were illuminated with ~1000 μmol m−2 s−1 photosynthetically active radiation (PAR) at the leaf surface by an LED worklight. Leaf temperature was monitored by a thermocouple. Leaf water potential (ψleaf) was measured using a C-52 sample chamber in a Dewpoint Water Potential System (Psypro, Wescor, Utah, USA); leaf water potential was measured after 25 min of equilibration at 25–30 °C. Four replicates were performed for each plant in this experiment. For hydrated leaves, the Kleaf was calculated as described previously (Sack and Scoffoni 2012) using the finalψleaf (ψfinal), which was determined at the end of the measurement of E (transpiration rate) which refers to the plant unit leaf area of transpiration of water within a certain amount of time. Kleaf is calculated as E/−∆ψleaf (where ∆ψleaf = ψfinal −0 MPa) and normalized by leaf area. Leaf area per square metre per hour is commonly used in transpiration water grams.
Application of ABA and HgCl2
Non-stressed leaves were perfused for 2 h with artificial xylem sap (AXS, i.e., filtered 20 mM KCl + 1 mM CaCl2 solution), AXS + 0.2 mM HgCl2 or AXS + 50 μM ABA (Laur and Hacke 2014). Solutions were introduced into the transpiring leaf by immersing the petiole in 50 mL containers. The excised leaves were then bench-dried at the different time points, and the leaves were frozen in liquid nitrogen and stored at −80 °C.
RNA extraction, reverse transcription-PCR and real-time quantitative PCR assay
Total RNA from the same plant material samples as previously described for the physiological experiments samples was extracted using TRIzol (Takara, Dalian, China) followed by an RNase-free treatment (Takara). The RNA (4 μg) was pretreated with RQ1 DNase I (Promega, Madison, WI, USA) to remove contaminating genomic DNA. Total RNA concentrations were measured using a NanoDrop unit (Thermo Fisher Scientific, Waltham, MA, USA), and RNA quality was checked using nondenaturing agarose gel electrophoresis. Total DNA-free RNA (2 μg) was used for complementary DNA (cDNA) synthesis in a total volume of 40 μL, and the resulting cDNA was used as the template for RT-PCR. The PCR for the mRNAs was carried out in 20 μL reactions containing 2 μL of cDNA synthesis reaction mixture, 400 nM of each primer, and 10.5 μL of SYBR Green PCR Master Mix (TianGen Biotech, Beijing, China) on an ABI 7500 sequence detection system (Applied Biosystems, Foster City, CA, USA). The temperature program began with 3 min at 95 °C, followed by 40 cycles of 15 s at 95 °C and 45 s at 57 °C; the melting temperature of the amplified product was determined to verify the presence of a specific product.
The main feature of mature miRNA expression detection is the RT primer, which has a stem-loop structure and a consensus sequence that effectively binds to the miRNA 3′ end. The PCR for the mature miRNAs was carried out in 25 μL reactions containing 2.5 μL of cDNA synthesis reaction mixture, 400 nM of each primer, and 12.5 μL of SYBR Green PCR Master Mix. The PCR temperature program began with 10 s at 95 °C, followed by 40 cycles of 10 s at 95 °C and 30 s at 60 °C. In addition, each measurement was repeated using three technical replicates, in which the RNA samples were mixed with three biological replicates.
To obtain accurate target mRNA expression profiles, we designed primers that flanked the cleavage sites using Primer Premier 5.0 (Premier, Canada). Sense primers were designed upstream of the miRNA cleavage sites and antisense primers were designed downstream of the cleavage sites to ensure that we would be able to detect the remaining intact mRNA levels after miRNA cleavage. For the normal genes, the sequences were obtained from the NCBI. The primer design involved selection of a specific sequence from the whole gene sequence. In addition, each measurement was repeated using three biological replicates. The primers for each gene are listed in Supplementary Table 1.
Deep sequencing
Tomato leaflet samples were collected from MP1 and 35S:mSlARF10-6 at the same stage and position on the plants, and an Illumina MiSeq library was constructed according to the manufacturer’s instructions (Illumina, San Diego, CA, USA). Magnetic beads with poly T oligos attached were used to purify the mRNA from the total RNA. Fragmentation buffer was added to cleave the mRNA into short fragments, and the fragments were used to synthesize first-strand cDNA using random hexamerprimers. The cDNA was transformed into double-stranded cDNA with RHase H and DNA polymerase I, and a paired-end library was constructed from the synthesized cDNA with a Genomic Sample Prep Kit (Illumina). Fragments of desirable lengths were purified with a QIAquick PCR Extraction Kit (Qiagen), end repaired and linked with sequencing adapters (Margulies et al. 2005). AMPureXP beads were used to remove the unsuitable fragments, and the sequencing library was the constructed by PCR amplification. After being checked with PicoGreen staining and fluorospectrophotometry and then quantified with an Agilent 2100, the multiplexed DNA libraries were mixed in equal volumes to a normalized 10 nM concentration. The sequencing library was then sequenced with the Illumina Miseq platform (Shanghai Personal Biotechnology Cp., Ltd., Shanghai, China).
Statistical analysis of the stoma characteristics
The epidermis was separated from the 20 true leaves from the MP1 and 35S:mSlARF10-6 plants to analyse stomatal density and size, respectively. The stomatal number and size from a field of view of a given area were measured under light microscopy (Zeiss Observer A1). The stomatal density for each leaf was calculated as the mean of four fields of view at 20× magnification, and the stomatal morphological parameters were measured as the mean of the 20 stomatal complexes for each leaf. Means were compared by Fisher’s LSD test at a significance level of 0.05. The stomatal morphological parameters were stomatal length, stomatal aperture width, stomata aperture area (length × width × 3.14) and stomata pore depth. The maximum stomatal conductance to water vapour (gwmax, in mol m−2 s−1) was estimated as follows (Franks and Beerling 2009; Franks and Farquhar 2001):
d = diffusivity of water in air (m2 s−1) (2.55 × 10−5); D = stomata per unit epidermal area (m−2); a = mean stomatal pore area (m2); V = molar volume of air (m3 mol−1) (22.4 × 10−3); and l = depth of the stomatal pore (approximated as W/2 for guard cells). The environmental conditions were consistent; thus, d and V are the same for both the wild-type (WT) and 35S:mSlARF10-6 plants.
Quantification of ABA
ABA was quantified using the following ELISA protocol (Yang et al. 2001). The samples were homogenized in liquid nitrogen and extracted in cold 80 % (v/v) methanol with butylated hydroxytoluene (1 mmol L−1) overnight at 4 °C. The extracts were collected after centrifugation at 10,000×g (4 °C) for 20 min and then passed through a C18 Sep-Pak cartridge (Waters, Milford, MA) and dried in N2. The residues were dissolved in PBS (0.01 mol L−1, pH 7.4) to determine the levels of ABA. Microtitration plates (Nunc) were coated with synthetic ABA-ovalbumin conjugates in NaHCO3 buffer (50 mmol L−1, pH 9.6) and left overnight at 37 °C. Ovalbumin solution (10 mg mL−1) was added to each well to block nonspecific binding, and after incubation for 30 min at 37 °C, standard ABA, samples and antibodies were added and incubated for an additional 45 min at 37 °C. The antibodies against ABA were obtained as described by Yang et al. (2001). Then, horseradish peroxidase-labelled goat anti-rabbit immunoglobulin was added to each well and incubated for 1 h at 37 °C. Finally, the buffered enzyme substrate (orthophenylenediamine) was added, and the enzyme reaction was carried out in the dark at 37 °C for 15 min and then terminated using 3 mol L−1 H2SO4. The absorbance was recorded at 490 nm. Analyses of the enzyme-immunoassay data were performed as described by Yang et al. (2001). The experiment was performed with three biological replicates, and three technical replicates.
Promoter analysis
The upstream sequence for each significantly differentially expressed tomato gene was obtained using the method by Liu et al. (2014), and we used the 2.5 kb sequence that was the 5′ of the start codon. The sequence and start codon locations were obtained from http://solgenomics.net/, and ITAG v 2.4 and v 2.5 were used for the gene models and chromosome sequences, respectively. The sequences were analysed to predict the promoter element using PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/).
Electrophoretic mobility shift assays (EMSAs)
The ARF10 was expressed as a His fusion in Rosetta cells, and ARF10 was proteolytically cleaved from the His tag during the purification. The oligonucleotides used to introduce pET30 were induced by IPTG and purified by Ni-NTP Spin Columns (Qiagen, Germany). Double-stranded 50 oligonucleotides corresponding to the sequences from the −318 to −368 Solyc06g011350.2 (GAGACA) and −1039 to −1089 Solyc05g008080.1 (TGTCTC) promoters were used as probes. Respectively, the sequences of the probes were as follows:
-
1.
F: ‘AATAGTTGATGATCTTGCAAGCTCTGTCTTGCTCTAACAAGTACCTTGAC’; R: ‘TTATCAACTACTAGAACGTTCGAGACAGAACGAGATTGTTCATGGAACTG’
-
2.
F: ‘TTTGTTGATTGTCTCGTTGATCTACAGAGGCTGGAAAAGCTAAGTATAAC’; R: ‘AAACAACTAACAGAGCAACTAGATGTCTCCGACCTTTTCGATTCATATTG’.
The oligonucleotide probes were 5′-end-labeled with biotin, 2 μL of 100 μM double-stranded probes, 0.2 μL of 50 mM EDTA, 2 μL binding buffer, 1 μL of poly (dI-dC), and 1 μL 50 % glycerol (700 ng/μL added to 2–3 μL to reach 1–2 μg). ARF10 protein was incubated for another 20 min at room temperature. The DNA–protein complexes were separated by electrophoresis through a 7 % nondenaturing polyacrylamide gel in 0.5 × TBE buffer for 40 min at 4 °C, 100 V; transmembrane: 100 V and 38 mA for 30 min. The membrane was then exposed to UV crosslinking for 2 min.
Transient expression and quantitative fluorometric β-glucuronidase assays
In the assay, the three AQP genes with the most upregulated expression (Solyc10g083880.1, ‘SlTIP1-1-like’, Soly06g011350.2, ‘SlPIP2;4’ and Solyc05g008080.1, ‘SlNIP-type like’) were selected for analysis. The promoter sequence was PCR-amplified to obtain some special elements (PIP-a, PIP-b, PIP-d, NIP-a, NIP-b, NIP-d, TIP-a, and TIP-b), and the PIP-c and NIP-c gene fragments were synthesized by the Shaibaisheng Company (Beijing, China). For the transient expression of transcripts, partial SlARF10 and full length SlABI5 were cloned, and then transferred into pB7GWIWG2(II) and pBI121 respectively. Agrobacteria harbouring the pBI101-AQPpromoters, pB7GWIWG2(II).0-ARF10 and pBI121-ABI5 were grown overnight in YEB medium and transferred to induction medium (Nimchuk et al. 2000) with 50 μM acetosyringone for 4 h until the OD600 reached 0.8–1.2. Bacteria were diluted with one volume of 10 mM MES at pH 5.6, 10 mM MgCl2, and 150 μM acetosyringone and pressure infiltrated into the leaves of 3–4-week-old tomato plants. Agrobacterium-mediated expression was performed as described in Wroblewski et al. (2005) with the EH105 Agrobacterium strain carrying the GUS-intron transgene in pBI101. At least three plants and three leaves per plant were used for each experiment.
Quantitative measurements of GUS activity were performed according to Jefferson et al. (1987). For the quantitative GUS assay, WT and 35S:mSlARF10-6 leaves were dissected and collected in 1.5 mL microfuge tubes with approximately three leaves per sample, and the GUS substrate was 4-methylumbelliferyl-β-d-glucuronide. Protein concentrations of the leaf extracts were determined by the dye-binding method. The GUS activities were calculated in relative units of fluorescence produced per μg protein per hour.
Yeast one-hybrid (Y1H) assays
Y1H assays were performed using a Matchmaker Gold Yeast One-Hybrid System Kit (Clontech) according to the manufacturer’s protocols. SlARF10 or SlABI5-like was ligated to pGADT7 to generate the AD-ARF/ABI construct. The fragments of the AQP gene promoter were ligated to the pAbAi vector to generate pAbAi-bait plasmids, which were then linearized and transformed into the Y1HGold yeast strain and selected with a plate containing selective synthetic dextrose medium lacking uracil. The AD-ARF/AD-ABI constructs were transformed into the Y1HGold strain containing pAbAi-bait and screened on an SD/-Ura/AbA plate. All transformations and screenings were performed three times.
In situ hybridizations
Templates for the generation of ribonucleic acid probes were constructed as follows. A fragment comprising nucleotides 255–554 of Solyc10g083880.1 was amplified using the primers shown in the Supplementary Information and ligated into pSPT18 (Roche). Plasmids were linearized with EcoRI or HindIII and transcribed with T7 RNA polymerase. RNA probes labelled with digoxigenin-UTP were prepared using a DIG RNA-labelling mix (Roche).
Plant material was collected from 6-week-old MP1 and 35S:mSlARF10-6, and the leaf tissue section preparation and hybridization of DIG-labelled probes were performed as described by Guerin et al. (2000). Probe quality was assessed using gel electrophoresis.
Results
Effects of miR160-resistant ARF10 overexpression on leaf morphology and water loss
The 35S:mSlARF10-6 plants displayed small leaflets with extremely narrow blades (Hendelman et al. 2012), and the leaf index (the ratio of the leaf length to the leaf width) was analysed to further assess leaf morphology. The results showed that the length of the 35S:mSlARF10-6 leaf blades was reduced only by an average of approximately 0.7 %, which was not significant; however, the width was significantly reduced by more than 45 % compared with the control (P < 0.01). As a result, the leaf index was increased from 2.05 ± 0.21 in the control plants to 6.38 ± 0.62 in the transgenic 35S:mSlARF10-6 plants (Table 1), and the length of the leaf petiole in 35S:mSlARF10-6 was also reduced relative to the control.
The shape of the leaf is a key factor for transpiration in this organ, and the leaf index is negatively related to leaf water loss (Liu et al. 2009). The WT leaves wilted with increased duration of the dehydration treatment (Fig. 1); the 35S:mSlARF10-6 also wilted with time but more seriously than the WT. The water loss percentage of the bench-dried WT leaves increased from 0 to 24 h under dehydration treatment and reached an approximate plateau (Fig. 2a). The water loss from 35S:mSlARF10-6 was also increased by dehydration treatment, but it was always higher than the WT. Therefore, overexpression of mSlARF10 may increase water loss from the leaves of transgenic plants by transpiration. The water fluxes in the leaves are regulated by the hydraulic conductivity of the leaf (K leaf) in response to many abiotic stresses (Martre et al. 2002), so the excised leaves were immediately immersed in AXS. The transpiration rate (E) and water potential (ψleaf) of the leaves were measured, yielding a calculated K leaf (ratio of E to ψleaf) for 35S:mSlARF10-6 of approximately 12.49 mM m−2 s−1 MPa−1 (Fig. 2b), which was significantly higher than that of the WT. These results showed that overexpression of mSlARF10 not only changed the leaf shape but also influenced the hydraulic conductance.

Comparison of leaf phenotypes between the WT and 35S:mSlARF10-6 plants. Detached leaves from 6-week-old plants were subjected to dehydration treatment. In each picture, the left side is the WT leaf, and the right side is the 35S:mSlARF10-6 leaf. a 0 h of dehydration treatment; b 1 h of dehydration treatment; c 2 h of dehydration treatment; d 6 h of dehydration treatment; e 12 h of dehydration treatment; f 24 h of dehydration treatment

Overexpression of mSlARF10 increased the leaf water loss. a Water loss percentages from leaves of WT and 35S:mSlARF10-6 plants. Excised leaves from the fifth leaves of 6-week-old plants were assayed for water loss. Both WT and transgenic plants were grown under normal conditions. The excised leaves were bench-dried and weighed. b The leaf hydraulic conductance (Kleaf) of WT and 35S:mSlARF10-6 was measured after the leaves were xylem perfused with AXS after 2–3 h. Data represent means ± SD of four independent experiments. *Indicates a significant difference (P ≤ 0.05), and **indicates a highly significant difference (P ≤ 0.01)
Changes in stomatal morphology and conductance in 35S:mSlARF10-6
Because the water loss and hydraulic conductivity were significantly different between 35S:mSlARF10-6 and WT, the factors related to leaf water loss were analysed. Stomatal size, aperture and density were investigated in mature WT and 35S:mSlARF10-6 leaves. Furthermore, the overexpression of mSlARF10 resulted in a significant decrease in stomatal density of approximately 30 ± 4 compared with 55 ± 4 in the WT (Fig. 3; Table 2). Stomatal density could affect water transpiration (Wang et al. 2007). The ‘stomatal index’ is defined as 100D/(DE + D), where D is the density of the stoma and DE is the number of epidermal cells per unit area (Royer 2001). The stomatal index in 35S:mSlARF10-6 was reduced to 2.94 compared with 3.07 in the WT, representing an approximate decrease of 4.3 % (Table 2). Based on the size of the stomata, the length of the WT stomata ranged from approximately 9.47 to 11.16, and the length in 35S:mSlARF10-6 ranged from 9.87 to 16.61 (Table 3). The size of the stomata was significantly larger in 35S:mSlARF10-6 than in the WT. The aperture width in the WT decreased with dehydration treatment time, and the aperture in 35S:mSlARF10-6 decreased considerably with treatment time. The stomatal aperture area decreased from 101.51 to 60.02 in the WT with dehydration treatment (40.87 %), and the area was 54.92 % higher in 35S:mSlARF10-6 than in the WT (195.19 vs. 87.99). The range of decrease of the stomatal aperture area with dehydration treatment time was also greater in 35S:mSlARF10-6 than in the WT. Before 6 h of dehydration treatment, the stomatal aperture area decreased by 20.89 % in the WT, and the stomatal aperture area decreased by 45 % in 35S:mSlARF10-6. Therefore, the 35S:mSlARF10-6 leaves were more sensitive to water stress than the WT leaves, as indicated by regulation of the stomatal aperture. Furthermore, the decrease in the area of the stomatal aperture before 6 h of dehydration treatment was greater in 35S:mSlARF10-6 than in the WT. The stomatal size, density and aperture were closely related to the stomata-dependent water loss, which was calculated according to previously established methods (Franks and Beerling 2009; Franks and Farquhar 2001). As calculated, the stomatal conductance to water vapour (gw) gradually decreased with the dehydration treatment in both the WT and 35S:mSlARF10-6 (Table 3), and gw (WT) < gw (35S:mSlARF10-6) at 0 h of treatment. Therefore, the overexpression of mSlARF10 decreased stomatal density, but the stomatal aperture increased. The stomatal conductance to water vapour was increased in 35S:mSlARF10-6 plants compared with in the WT. The gw was higher in 35S:mSlARF10-6 than in the WT before 6 h of dehydration treatment, whereas gw was lower in 35S:mSlARF10-6 after 6 h. However, the gw and actual water loss rate were not consistent; thus, there may be another water loss pathway after dehydration treatment for 6 h. The change in the speed and extent of stomatal closure in 35S:mSlARF10-6 implied that the main factor regulating stoma ABA was different from that in the WT. The stomatal morphology further implied that SlARF10 may play a function in stomatal development and the response to water stress by regulating the stomatal aperture.

Stomatal number decreased in transgenic 35S:mSlARF10-6 plants. Microscopic views of stomata in the fifth leaves of transgenic and WT plants. Six-week-old plants were used for all experiments. Bar 20 μm
miR160-resistant ARF10 overexpression increases ABA accumulation and sensitivity during drought
Drought stress triggers the production of the phytohormone ABA, which causes stomatal closure (Finkelstein et al. 2002) that may be induced by complex signal transduction (Schroeder et al. 2001). To verify that stomata-dependent water loss was altered in 35S:mSlARF10-6, the endogenous ABA content and ABA-induced stomatal closures were investigated during dehydration treatment (Table 4). The accumulated endogenous ABA was gradually increased in the WT over 0–24 h of dehydration treatment. The content of ABA was also increased in 35S:mSlARF10-6 with the dehydration treatment, and reached the peak at the 6 h, then slightly decreased finally kept the high level and was always significantly higher than in the WT over 0–24 h of dehydration treatment (P < 0.01). ABA sensitivity experiments were performed to determine whether SlARF10 is involved in ABA signalling. Non-stressed detached leaves from 35S:mSlARF10-6 and WT were perfused for 2 h in AXS + ABA solution, after which the leaves were subjected to the drought condition. ABA treatment could decrease water loss in both 35S:mSlARF10-6 and WT leaves considerably (Fig. 4). The largest decline in water loss from ABA-treated 35S:mSlARF10-6 leaves, approximately 20 %, occurred after 6 h of dehydration treatment, which was even lower than the water loss from ABA-treated WT leaves before 6 h. The speed of water loss in 35S:mSlARF10-6 was higher than in the WT, but after treatment with ABA, it was much lower than in the WT before 6 h of dehydration treatment. This result indicated that ABA had a larger inhibitory effect on water loss in 35S:mSlARF10-6 leaves than in the WT during the early dehydration treatment. Although ABA treatment could also significantly decrease the stomata aperture size in the WT, leaves from 35S:mSlARF10-6 showed greater sensitivity to the ABA treatment and exhibited a swifter and larger decrease in stomatal aperture size (Fig. 5). Only partial stomas in the WT plants were closed after 6 h of ABA treatment, and nearly all of the stomas in 35S:mSlARF10-6 were closed after dehydration treatment for 6 h. The decrease in stomata aperture size after ABA treatment demonstrated that ABA sensitivity in 35S:mSlARF10-6 was higher than in the WT. The calculated stomata-dependent water loss in the WT was higher than in 35S:mSlARF10-6 after the dehydration treatment for 6 h. However this cannot explain why the water loss in 35S:mSlARF10-6 exceeded that in the WT after ABA treatment for 6 h. This result implies another water loss pathway besides stomata.

Water loss percentages from the leaves of WT and 35S:mSlARF10-6 plants as well as the leaves with ABA treatment, HgCl2 treatment or combined ABA and HgCl2 treatment. Both WT and transgenic plants were grown under normal conditions for 6 weeks, and then, the leaves were excised from the fifth leaves of the plants. The excised leaves were perfused for 2 h with ABA for 2 h, HgCl2 or ABA and HgCl2 and then weighed under the dehydration treatment. Data represent means ± SD of four independent experiments. *Indicates a significant difference (P ≤ 0.05), and **indicates a highly significant difference (P ≤ 0.01)

WT and 35S:mSlARF10-6 leaves were treated with ABA. Stomatal aperture with the ABA treatment decreased in transgenic 35S:mSlARF10-6 leaves more notably than in WT leaves. a WT leaves with ABA treatment after 0 h; b WT leaves with ABA treatment after 6 h; c 35S:mSlARF10-6 leaves with ABA treatment after 0 h; d 35S:mSlARF10-6 leaves with ABA treatment after 6 h. Microscopic views of stomata in the fifth leaves of transgenic and WT plants. Six-week-old plants were used for all experiments. Bar 25 μm
The advanced aquaporin activity in 35S:mSlARF10 contributes to water loss
Water movement across the leaf cell membranes is regulated by AQPs in the membranes (Heinen et al. 2009); thus, to reveal weather there is a difference in water movement in WT and 35S:mSlARF10-6, WT and 35S:mSlARF10-6 leaves were treated with HgCl2 (aquaporin inhibitor) to reveal the function of AQPs in leaf water loss (Fig. 4). The HgCl2 treatment significantly inhibited water loss in the WT, and the inhibition function became gradually stronger with increasing dehydration treatment time, especially after 6 h. In 35S:mSlARF10-6, HgCl2 showed complete inhibition of water loss after 6 h of dehydration treatment, although it did play a significant role before 6 h. After combining ABA and HgCl2 to treat the WT and 35S:mSlARF10-6 leaves, the water loss percentage was even significantly lower in 35S:mSlARF10-6 than in the WT (Fig. 4). From the Kleaf, it was also found that the water hydraulic conductance was higher in the 35S:mSlARF10-6 leaves (Fig. 2b). This suggested that the effect of AQP activity on water movement in leaves was higher in the 35S:mSlARF10-6 leaves.
ABA biosynthesis/signalling, aquaporin and stomatal development genes in 35S:mSlARF10
Next, we conducted deep sequencing using RNA from the fifth leaves of WT and 35S:mSlARF10-6 plants to determine the effects of the 35S:mSlARF10-6 leaves on global ABA biosynthesis, signalling and AQP gene expression. We identified five AQP genes that were significantly upregulated in the 35S:mSlARF10-6 and two ABA biosynthesis and twelvesignal/response genes that were also significantly upregulated or down regulated by more than twofold (P < 0.05) in 35S:mSlARF10-6 (Table 5). One transcription factor was SlABI5-like (ABSCISIC ACID-INSENSITIVE 5-like), which is known to be involved in the ABA signalling pathway, and the RT-PCR results validated the transcriptome data (Fig. S2). Furthermore, some genes related to stomatal development and regulation were significantly downregulated in 35S:mSlARF10-6 (Table 5). The transcript factors MUTE, TOO MANY MOUTHS (TMM) and LRR receptor-like serine/threonine-protein kinase ERECTA-like were significantly lower in 35S:mSlARF10-6 than in the WT.
Promoter analysis of the upregulated expression of AQP genes in 35S:mSlARF10-6
The above data demonstrated that the expression of five AQP genes was induced; thus, the promoter regions (2500-bp upstream sequences) of these genes were subjected to motif analysis using PlantCARE to analyse the cis-acting element in those upregulated AQP. The results show that among the AQPs, there were two upregulated AQP gene promoters that contained only AuxRE (TGTCTC/GAGACA); one AQP gene promoter contained only ABRE, whereas the other two had both ABRE and AuxRE (Table S2; Fig. S1).
The three most upregulated AQP genes (Solyc10g083880.1 ‘SlTIP1-1-like’, Soly06g011350.2 ‘SlPIP2;4’ and Solyc05g008080.1 ‘SlNIP-type like’) were selected to verify that AuxRE and ABRE upregulated the expression of the AQP genes, and their promoters contained ABRE or both AuxRE and ABRE, respectively, which expression levels were upregulated 21.5, 5.04 and 2.7 times in 35S:mSlARF10-6. Chimeric constructs consisting of the SlPIP2;4 or SlNIP-type-like four-promoter fragments and SlTIP1-1-like two-promoter fragments and the GUS reporter gene (Fig. 6a) were introduced into tomato leaves. The transient expression assay demonstrated that the GUS activity of the complete SlPIP2;4 promoter fragment was higher in 35S:mSlARF10-6 than in the control, and the activities of the GUS promoters containing only ABRE or AuxRE were both higher in 35S:mSlARF10-6 than in the WT (Fig. 6b). The GUS activity of the complete SlNIP-like promoter was higher in 35S:mSlARF10-6 than in the control, and the activity of GUS containing only AuxRE was also enhanced, whereas ABRE exhibited depressive effects (Fig. 6c). The GUS activity of the promoter containing the ABRE of SlTIP1-1-like was higher in 35S:mSlARF10, and the GUS activity of the promoter without the ABRE of SlTIP1-1-like was not changed significantly (Fig. 6d). These results implied that the upregulated expression of AQP is induced by AuxRE and sometimes by ABRE in 35S:mSlARF10-6.


The promoter fragments of Soly06g011350.2 (SlPIP2;4), Solyc05g008080.1 (SlNIP-type like), Solyc10g083880.1(SlTIP1-1-like) were cloned into the pBI101 plant expression vector and transformed into Agrobacterium EH105 to inject WT and 35S:mSlARF10-6 tomato leaves with the GUS reporter gene. a The promoter sequence analysis to illustrate the role of each element (AuxRE and ABRE), the complete promoter sequence with both AuxRE and ABRE (PIP-a, NIP-a), the promoter sequence with only AuxRE (PIP-b, NIP-b), the promoter sequence with only ABRE (PIP-c, NIP-c, TIP-a) and the promoter sequence without both AuxRE and ABRE (PIP-d, NIP-d, TIP-b) were cloned into the vector. b The GUS activity analysis of the promoter (PIP-a, PIP-b, PIP-c, PIP-d). c The GUS activity analysis of the promoter (NIP-a, NIP-b, NIP-c, NIP-d). d The GUS activity analysis of the promoter (TIP-a, TIP-b). Bars indicate the standard errors of three replicates. The control treatment is the background. *Indicates a significant difference (P ≤ 0.05), and **indicates a highly significant difference (P ≤ 0.01)
SlABI5-like stimulates AQP expression, and ARF10 is required to induce AQP expression
In 35S:mSlARF10-6, mSlARF10 and ABSCISIC ACID-INSENSITIVE 5-like (SlABI5-like) are two suitable candidates for the direct binding of those elements; thus, we first examined the binding of the promoter fragments of (Soly06g011350.2) ‘SlPIP2;4’ and (Solyc05g008080.1) ‘SlNIP-type like’ containing AuxRE to the SlARF10 protein. At 318–368 upstream of the start codon (ATG), in Soly06g011350.2 had an AuxRE (GAGACA), and Solyc05g008080.1 had an AuxRE (TGTCTC) at 1039–1089 bp upstream of the start codon (ATG). We performed EMSAs with an in vitro-translated ARF10 protein and the 5′ promoter region of the Soly06g011350.2 and Solyc05g008080.1 with biotin labelling. We first constructed a SlARF10 and pET30a fusion recombinant plasmid, and after sequencing, the plasmid was transferred into the ‘rosseta’ bacterial strain for expression. A SlARF10 fusion His recombinant protein was obtained by induction and purification, and the protein, which was purified by Coomassie Brilliant Blue staining, was approximately 72 kDa in size (Fig. 7a). The size was approximately the sum of ARF10 (63 kDa) and His (6 kDa) and verified by His antibody Western blotting, which indicated that this protein was indeed the His-ARF10 fusion protein. An EMSA assay indicated that the His-ARF10 protein bound the oligo DNA with molecules with the SlPIP2;4 and SlNIP-type promoter fragments, respectively (Fig. 7b), which suggests that SlARF10 could bind the promoter with TGTCTC or GAGACA to regulate the expression of the SlPIP2;4 and SlNIP-type like genes.

a In vitro-translated SlARF10 analysed by sodium dodecyl sulphate–polyacrylamide gel electrophoresis. Lane 1 protein markers. Lane 2 SlARF10-His fusion protein. b EMSA of in vitro-translated SlARF10 protein binding to the 318–368 fragment of Soly06g011350.2 (PIP2;4) promoter (GACACA) and the 1039–1089 fragment of Solyc05g008080.1 (NIP-type like) promoter (TGTCTC). #1 probe is Soly06g011350.2 (SlPIP2;4) promoter; #2 probe is Solyc05g008080.1 (SlNIP-type like) promoter. Lane 1 control; lane 2 #1 probe two times volume; lane 3 #1 probe four times volume; lane 4 control; lane 5 #2 probe two times volume; lane 6 #2 probe four times volume
A Y1H assay was also performed to test the association of SlARF10 with the promoter of the SlPIP2;4 and SlNIP-type like genes. The results showed that SlARF10 could associate with two AuxRE located at −6 bp to −1399 bp of the SlPIP2;4 and −7 bp to 1466 bp of the SlNIP-type like genes, respectively (Fig. 8), and the mutation of these binding sites could break this association, supporting the hypothesis that SlARF10 was the transcription factor associated with the AQPs. Next, further confirming the effect of the ABA signal on the AQP genes, Solyc09g009490.2 (ABSCISIC ACID-INSENSITIVE 5-like) was upregulated by two fold in 35S:mSlARF10-6 from the transcriptome. We chose the AQP gene (Solyc10g083880.1, SlTIP1-1-like), whose expression levels were upregulated 21 times from the transcriptome data, and the SlNIP-type like, which was upregulated 2.7 times, for the ABRE binding test because they could show enhanced GUS activity in transient expression. As shown in Fig. 8b, SlABI5-like bound to the ABRE containing DNA fragments (GAGACA) of the SlTIP1-1-like promoter but failed to bind to ABRE-mutated motifs (AAAAAA), and the same result was obtained for the association with SlNIP-type-like gene. The results demonstrated that upregulated AQP genes could also be directly stimulated by SlABI5-like.

a The association of SlABI5-like with the promoter of Soly06g011350.2 ‘SlPIP2;4’ and Solyc10g083880.1 ‘SlTIP1-1-like’ by a Y1H assay. b The association of SlARF10 with the promoter of Soly06g011350.2 ‘SlPIP2;4’, Solyc05g008080.1 ‘SlNIP-type like’. Four AQP gene promoter elements were mutated and interacted with SlABI5-like and SlARF10. Each fragment of the AQP gene promoter was ligated to a pAbAi vector to generate pAbAi-bait plasmids, and AD-ABI5-like or AD-ARF10 was transformed into the Y1HGold strain holding pAbAi-bait and screened again on a plate with selective synthetic dextrose medium lacking leucyl/AbA. An empty pGADT7 vector (AD) was transformed as a negative control
The effect of transient expression of SlARF10 and SlABI5 on leaf water loss
We have shown that SlABI5 and SlARF10 bind on the AQPs promoters. For further confirmation of roles of SlABI5 and SlARF10 in leaf water loss, SlABI5 overexpression and SlARF10 silencing vectors were transiently overexpressed in tomato leaves in an Agrobacterium mediated process. Three days after the injection, the expression levels of SlABI5 and SlARF10 were confirmed. The results showed that the expression levels of SlARF10 in containing pB7GWIWG2(II)-ARF10 tomato leaves were decreased more than three times (Fig. S9). Furthermore, the expression levels of SlABI5 were increased more than five times in the tomato leaves containing pBI121-ABI5 (Fig. S9). Next we performed a water loss experiment with the transient expression leaves. The results showed that transient silencing of SlARF10 significantly decreased the water loss rate compared to CK especially after the 6 h dehydration treatment (Fig. 9a), and the expression levels of the most of the AQP family members were decreased, especially Solyc10g083880.1 ‘SlTIP1-1-like’ and Solyc05g008080.1 ‘SlNIP-type-like’ (Fig. S9). The transient overexpression of SlABI5 showed that the water loss rate was significantly lower than CK before the 6h dehydration treatment,but after 6h, the water loss rate was almost the same as CK (Fig. 9b). The expression levels of some of the AQP family members were increased especially Solyc10g083880.1 ‘SlTIP1-1-like’ and Solyc05g008080.1 ‘SlNIP-type like’ whose promoters were confirmed to be bound by SlABI5 through a Y1H assay (Fig. S9), but the expression levels of other AQP family members were largely not increased. It was also found that the AQP genes with the greatest increases in expression were those whose promoters contained ABRE elements (Fig. S10).

Water loss percentages from CK leaves and transient expression leaves containing pB7GWIWG2(II).0-ARF10 and pBI121-ABI5. a The water loss of leaves that were injected with the Agrobacterium harbouring pB7GWIWG2(II).0-ARF10 or pB7GWIWG2(II).0. b The water loss of leaves that were injected with the Agrobacterium harbouring pBI121-ABI5 and pBI121. The excised leaves were bench-dried and weighed. *Indicates a significant difference (P ≤ 0.05), and **indicates a highly significant difference (P ≤ 0.01)
AQP expression is tissue specific in the upper epidermis of the tomato leaf
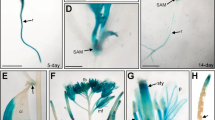
In situ hybridization experiments were performed to further reveal the distribution and accumulation of up-expression of AQP mRNA in 35S:mSlARF10-6. SlPIP2;4 (Solyc06g011350.2) was selected because its promoter contains both AuxRE and ABRE and is the second most highly up-expressed AQP in 35S:mSlARF10-6. A single-stranded RNA complement to the SlPIP2;4 gene produced a weak signal that was mainly observed in the leaf epidermal cells and neighbouring midrib mesophyll cells in the WT (Fig. 10b). The probe produced stronger chromogenic signals in 35S:mSlARF10-6 leaves from the epidermis to midrib cells (Fig. 10c), and greater SlPIP2;4 expression was detected in 35S:mSlARF10-6 leaf epidermis than in the WT.

Induction of Soly06g011350.2 ‘SlPIP2;4’ in 35S:mSlARF10-6 in the leaf epidermis. Detection of SlPIP2;4 expression in leaves. a Control cross-sections probed with a SlPIP2;4 sense probe; b the cross-sections of wild type leaves with a DIG-labelled ssRNA probe complementary to a SlPIP2;4 probe; c the cross-sections of 35S:mSlARF10-6 leaves with a DIG-labelled ssRNA probe complementary to SlPIP2;4. Bar 50 μm
SlARF10 links both auxin and ABA signalling in the regulatory mechanism to mediate leaf water loss
The results of this study could be used to build a network to illustrate the precise role of miR160 in the regulation of ARF10 in plant water stress (Fig. 11). Thus, we proposed a model of the effect of the ABA-auxin interaction on the control of water loss. When auxin signalling was activated, Aux/IAA was degraded, and ARF10 was freed at a high transcription level. Although ARF10 could enhance ABA content through active ABA synthesis gene expression and promote the ABA signal through ABI5 expression, which resulted in stomatal closure under drought. SlARF10 overdose enhanced the leaf water loss rate by modifying stomatal development and AQP expression. And SlABI5 increased the AQPs which induced the cell osmotic in leaf. The rational ARF10 expression levels mediated by miRNA160 may play an important role in maintaining leaf water balance during leaf development and adaptation to water stress.

A proposed model for the role of miR160-regulated ARF10 in response to leaf water loss. When auxin signalling was activated, AUX/IAA degraded, and ARF10 was freed at a high transcription level. miR160 also regulates the expression levels of ARF10, which directly influences stomatal development. ARF10 increases ABA content and promotes ABA signal ‘ABI5-like’ expression, resulting in sensitive stomatal closure under drought. ARF10 increased the leaf water loss rate by directly increasing AQP expression to enhance the hydraulic conductance. Stomatal-dependent and AQP-dependent hydraulic conductance both influence the water loss in the leaf. → and — indicate positive and negative modes of regulation, respectively; [ABA] indicates ABA concentration
Discussion
Overexpression of mSlARF10 resulted in narrower leaves but a higher rate of water loss
An earlier article showed that overexpression of mSlARF10 could alter leaf size to result in leaflets with extremely narrow blades (Hendelman et al. 2012). Narrower leaves typically have lower water loss rates due to the low transpiration of the small leaf area and are selected during breeding as a trait to enhance drought resistance (Liu et al. 2010). In 35S:mSlARF10 tomatoes, the overexpression of mSlARF10 resulted in greater water loss from the narrower leaves; however, 35S:SlARF10 which also had narrow leaves, demonstrated a lower rate of water loss than 35S:mSlARF10-6, and a significantly higher rate of water loss than WT (Fig. S6). A reasonable explanation for this result is that the expression levels of SlARF10 were lower than 35S:mSlARF10-6 but higher than WT. Thus, the regulation of miR160 on SlARF10 maintains the balance of the expression levels of ARF10 to mediate the leaf water loss rate. There is higher water loss efficiency in 35S:mSlARF10 tomato leaves than in the WT. The 35S:mSlARF10 tomato has a higher Kleaf than the WT, which supports this hypothesis.
The leaf is a hydraulic bottleneck that accounts for a large part of plant resistance to drought (Brodribb and Cochard 2009), and water loss is restricted by the hydraulic and stomatal conductance of the leaf (Pou et al. 2013). The leaf hydraulic conductance (Kleaf) is determined by the multiple components of the water transport pathways; water moves through the petiole and vein xylem, as well as bundle sheath and mesophyll tissue, before evaporating through the stomata. The decline in Kleaf may be caused by losses in conductivity in one or more of these components (Johnson et al. 2009; Scoffoni et al. 2011), and among them, stomatal closure is the major regulator of plant water loss. When stomata open for photosynthesis, water is lost via transpiration, and Kleaf must remain high to prevent the tissue water potential from declining sufficiently far to trigger a decline in stomatal conductance (Tyree and Zimmermann 2002). However, stomatal closure may be decreased under water stress in response to a decline in leaf hydraulic conductance (Sack and Holbrook 2006). Overexpression of mSlARF10 resulted in a larger individual stoma size but lower stomatal density in the leaf, and the transcriptome data demonstrated that the transcript factors regulating stomatal development were decreased significantly in 35S:mSlARF10-6. In 35S:SlARF10, the stomatal parameters were between the 35S:mSlARF10-6 and WT plants and did not obviously change (Table S3; Fig. S7). ARF10 regulated by miR160 was also important for the regulation of stoma development. The overexpression of MUTE has been implicated in changing leaf epidermis cells into stomata (Pillitteri et al. 2007; Serna 2007), and the leucine-rich repeat domain (LRR) receptor-like protein and TMM have been reported to be involved in stomatal regulation (Hunt and Gray 2009). Theoretical water loss based on calculated stomatal-dependent water loss was confirmed to be consistent with the actual water loss in 35S:mSlARF10 and the WT under normal conditions. However, the water loss rate does not precisely adhere to the calculated stomatal-dependent water loss in 35S:mSlARF10 and the WT after water stress for 6 h, which indicates that there may be a complex water loss pathway in 35S:mSlARF10-6.
In contrast, reducing AQP activity and potentially emptying the water-filled cell wall pores in the bundle sheath and mesophyll is another important contributor to the decline in Kleaf (Pieruschka et al. 2010; Shatil-Cohen et al. 2011). AQPs have been implicated in the determination of internal conductance to water in the leaf (Heinen et al. 2009), and reduced/enhanced AQP activity is associated with AQP activation and/or expression inhibition (Cochard et al. 2007; Nardini et al. 2005; Voicu et al. 2008, 2009). The downregulation of AQPs under water stress and coordination with stomatal conductance represent a mechanism for the coregulation of water loss at the cellular level (Pou et al. 2013).
Differences in water loss between 35S:mSlARF10 and the WT that occur when plants are treated with ABA (stomatal opening inhibitor) and HgCl2 (special AQP inhibitor) further support the conclusion that stomatal closure and AQP activity contribute more to water loss in 35S:mSlARF10 than in the WT. Treatment with only ABA for less than 6 h significantly decreased water loss in 35S:mSlARF10 to an even greater degree than in the WT treated with ABA, which indicated that 35S:mSlARF10 has greater water loss efficiency through the stomata. In the next 18 h, 35S:mSlARF10 treated with ABA exhibited a lower water loss rate than WT, revealing that the stomata are not the only reason for the more rapid water loss in 35S:mSlARF10. Using a special AQP inhibitor, HgCl2, to treat the leaf had little effect before 6 h, but it significantly inhibited the water loss in 35S:mSlARF10 after 6 h. Furthermore, in potatoes, a significant decrease in stomatal conductance may occur at an early stage of drying, even before a significant decrease in leaf water potential (Jefferies and Mackerron 1989). Combining the ABA and HgCl2 treatments could further depress water loss in 35S:mSlARF10; it was even lower than that in treated WT throughout the duration of water stress. These results reveal that the overexpression of mSlARF10 could alter leaf water loss by modifying stomatal closure and AQP activity during drought. We also examined the influence of ABA or HgCl2 treatments on 35S:SlARF10 leaf water loss (Fig. S8). Both demonstrated little inhibition of the water loss in 35S:SlARF10 leaves. The degree of inhibition in 35S:SlARF10 was between the 35S:mSlARF10-6 and MP1 by ABA or HgCl2.
Overexpression of mSlARF10 affects ABA synthesis and signalling to regulate stomatal closure
The plant hormone ABA accumulates under water deficit conditions and plays a major role in dehydration response and tolerance (Wan et al. 2004). A high ABA concentration around guard cell results in stomata closure, and the expression of AQPs and the depression of their activities help plants conserve water. In 35S:mSlARF10, high ABA content and sensitivity increase stomatal closure and the response to ABA. In ABA biosynthesis, analysis of RNA sequences did not reveal differences in the expression of key ABA biosynthesis genes (NCED) but did identify differential expression of geranylgeranyl pyrophosphate synthase, which is a key enzyme in the carotenoid biosynthetic pathway, with significant up-regulation in 35S:mSlARF10. Two pathways have been proposed for the biosynthesis of ABA. It has been shown that the ABA produced from the oxidative cleavage of carotenoids is the main pathway (Seo and Koshiba 2002). In this pathway, IPP is converted to a C20 product by geranylgeranyl pyrophosphate (GGPP). It is known that GGPP is a precursor of carotenoids (Cunningham et al. 1993). This procession is also reported to be involved in plant responses to the abiostresses such as salt stress and drought stress (Ruiz-Sola et al. 2014). During dehydration, the plant ABA content would show a continues increased trend to improve the adaption (Seo and Koshiba 2002). In this study, the ABA content of 35S:mSlARF10-6 with dehydration treatment have a maximum at 12 h and then decrease but maintain a high level. The reasonable explanation is that the ABA content reached highest level at 50 % relative water content during dehydration (Di Blasi et al. 1998). In our data, the 35S:mSlARF10-6 relative water content at 12 h dehydration treatment is around 50 %, even the longer treatment would make the 35S:mSlARF10-6 relative water content smaller, the ABA accumulation is not increased. In addition to the ABA content, the ABA signal is also reportedly involved in the adaptive responses of plants to drought stress. There are three core components: the ABA receptors PYR/PYL/RCARs (hereafter the ‘PYLs’), the negative regulators named PP2Cs (type 2C protein phosphatases), and positive regulators termed SnRK2s (subfamily 2 of SNF1-related kinases) (Ma et al. 2009; Park et al. 2009). The expression of the CYP707A gene encoding ABA 8′-hydroxlase could increase in response to dehydration stresses as well as high ABA levels (Saito et al. 2004). Water stress alters the ABA signal by modifying the core gene expression, and expression analysis indicated that most SlPYL genes were downregulated, whereas most SlPP2C and SlSnRK2 genes were upregulated by dehydration in tomato leaf (Sun et al. 2011). ABI3 is sufficient to change ABA responsiveness in vegetative tissues, and ABI1 is required for ABI3-regulated processes (Bonetta and McCourt 1998). The altered ABA signal would modify the ABA response during water stress, and overexpression of miR160 resulted in reduced sensitivity to ABA during germination. Transcriptome analysis of germinating mARF10 seeds indicated that the typical ABA-responsive genes expressed during seed maturation were overexpressed in germinating mARF10 seeds (Liu et al. 2007), and further study has revealed that the highly similar ARF10 and ARF16 proteins could both control the expression of ABI3 during seed germination but without directly binding to the ABI3 promoter (Liu et al. 2013).
In 35S:mSlARF10 leaves, both the ABA content and the ABA sensitivity (stomatal closure rate response to exogenous ABA) were significantly enhanced. RNA sequencing showed that two ABA synthesis genes and twelveABA response/signal transduction genes containing ABI5 were significantly changed in 35S:mSlARF10 leaves, and this could indicate why ABA content and sensitivity are enhanced in 35S:mSlARF10. It has been reported that HvABI5 is indispensable for ABA induction of gene expression, which is an ABI5 orthologue in barley (Zhou et al. 2013). In addition to modulating the adaptive stress response, OsABI5 also plays crucial roles in plant fertility (Zou et al. 2007, 2008), and the rice ABI5-like gene has been shown to be an important node in the cross-talk of ABA–auxin signalling (Yang et al. 2011). However, ARF10 may not directly bind to the ABI5 promoter because it lacks the typical AuxRE element, but ABI5 promoter-containing ABRE elements and high ABA content and signal exits in 35S:mSlARF10 imply that ARF10 could be a positive ABI5 regulator via an indirect effect.
High leaf water loss by overexpression of mSlARF10 might be mediated directly by ARF10 function and not through ABI5
Overexpressed mSlARF10 acts as a transcription activator to directly induce the expression of AQPs to increase the hydraulic conductance. The ARFs regulate the expression of a large set of auxin-responsive genes by binding to the auxin response elements (AuxREs) in their promoters (Guilfoyle and Hagen 2007), and the ARF protein sequences contained in the middle regions function as activation or repression domains (Ulmasov et al. 1999). There is a conflict in the literature over SlARF10 transcription activity; Wu et al. (2011) consider it to be a repressor, whereas Kumar et al. (2011) classify it as an activator. Alternatively, the ARFs may activate or repress transcription in selected cell types that contain a different set of coactivators or corepressors (Ulmasov et al. 1999). Promoter analysis showed that the significantly upregulated AQP genes in 35S:mSlARF10 contain AuxRE, and transient expression confirmed that overexpression of mSlARF10 could induce the promoter GUS activity. The Y1H assay and EMSA demonstrated direct binding between the SlARF10 protein and DNA sequence containing AuxRE. Additionally, the ‘ARF10-’ transient expression demonstrated that the water loss rate of leaves was decreased, and most of the AQP gene expression levels were decreased. Furthermore, in 35S:mSlARF10-6, overexpression of mSlARF10 always induce es GUS promoter activity for AQPs promoters containeing AuxRE. However, ‘ABI5+’ transient expression indicated that the ABI5 did not induce the increased leaf water loss, but induce some AQP genes expression levels increased.
AQP expression has been described in all of the leaf tissues in which high transmembrane water movement is required; however, AQPs can still facilitate other functions. During water stress, most plasma membrane intrinsic proteins (PIPs) and tonoplast intrinsic proteins (TIPs) show reduced expression (Alexandersson et al. 2005), and dehydration treatment rapidly decreases levels of PIP1;5, PIP2;2, PIP2;3 and PIP2;6 transcripts to one-tenth of their normal levels (Jang et al. 2004). Furthermore, post-translational regulation of PIPs has been implicated in reduced water transport under water deficit conditions. Finally, there is a negative correlation between AQP abundance and transpiration rate (Morillon and Chrispeels 2001). Plant AQPs may divide into two “classes” during dehydration. The first would be primarily involved in the maintenance of cellular water (osmotic) status, not the stress response mechanisms, and the second class of AQPs would be specifically expressed and/or regulated in the appropriate organs following stresses to compensate for the altered water potential (stress-responsive isoforms). Mutation of these isoforms would markedly alter the overall resistance of the plants to these stresses (Hachez et al. 2006a), and this information implies that the expression of only some special AQPs is related to water loss. The responses of AQPs to water stress may involve either the up- or downregulation of gene expression or no change (Tyerman et al. 2002).
It is reasonably deduced that AQPs directly regulated by SlARF10 play the major role in responding to water stress, while AQPs regulated by SlABI5 may have roles in maintaining the cell osmotic balance for plants to better adopt to the environment, e.g. stomatal closure to avoid excessive water loss (Raschke 1987; Zeevaart and Yang 2005). Other reports also indicated that high expression of ABI5 is coupled with enhanced adaptation to stress. ABI5 expression is highest in young seedlings exposed to dehydrating stress (Finkelstein and Lynch 2000; Lopez-Molina et al. 2001). Furthermore, ABI5 overexpressing plants are hypersensitive to abscisic acid and retain water more efficiently than WT plants (Lopez-Molina et al. 2001). The similar strategy identified for ABA can be involved in triggering the expression of AQP-like genes to achieve development and drought response (Ingram and Bartels 1996).
Overexpression of mSlARF10 induced AQP expression in the leaf epidermis
Plant aquaporins localize in all subcellular compartments forming or derived from the secretory pathway. Special expression of a PIP and a NIP homolog on the distal side of root exo- and endodermal cells has been described in maize and rice, respectively. Such cell polarization is consistent with the uptake and centripetal transport of water. (Hachez et al. 2006a; Ma et al. 2006; Maurel et al. 2008). Our in situ hybridization results demonstrated that the abnormal upregulated AQPs in 35S:mSlARF10 were mainly expressed in the epidermis of the leaves, and the AQPs at this position might constitute a plausible reason for high water conductance and loss in 35S:mSlARF10 plants.
miR160-regulated SlARF10 mediated auxin and ABA signalling in the leaf in response to water loss
During normal leaf development in wheat, the water loss rate of the excised leaf first increased and then decreased with increasing leaf age (Zhao et al. 2009; Yang et al. 2002). The increase in the residual transpiration rate with leaf age is due to the duration of exposure to the rigors of the external environment (Clarke and Richards 1988). Direct evidence for the important role of AQPs in leaf growth was provided by (Uehlein et al. 2003); in tobacco, NtAQP1 overexpression increased the leaf growth rate, an effect that can be attributed to NtAQP1 acting as both a water channel and CO2 transporter. The expression levels of ARF10 and miR160 were investigated in the different leaf development processes (Fig. S4); ARF10 first increased in the tender, mature, old leaves and then decreased in the yellow leaves. miR160 expression levels first decreased and then increased, which is contrary to ARF10 expression panters. It is reasonably deduced that ARF10/miR160 are involved in leaf development via coordination of water metabolism. Moreover, miR160 and ARF10 are involved in mediating auxin sensitivity. In Arabidopsis, ARF10 mediates the link between ABA and auxin responsiveness during and after seed germination (Liu et al. 2007). In soybean, ectopic expression of miR160 results in auxin hypersensitivity while miR160 appears to promote auxin activity by suppressing the levels of the ARF10/16/17 family of repressor ARF transcription factors (Nizampatnam et al. 2015; Turner et al. 2013). ARF10 expression is typically highly mediated by miR160, and in the overexpressed ARF10 line, the upregulated expression of miR160 was also observed to result in cleavage of ARF10 to maintain stable expression level (Hendelman et al. 2012). This fine tuning mechanism plays an important role in auxin sensitivity.This miRNA-mediated auxin signalling in the regulation of leaf water loss is likely an evolutionary mechanism that prevents the auxin response factor from inducing leaf water loss to maintain normal development, and it could also be crucial for improving drought stress tolerance. There are many studies indicating that miRNAs regulate ARFs to influence leaf and root development to affect drought stress and tolerance (Ding et al. 2013). The functions of ARF10 in leaf water loss may have an evolutionary benefit. ARF10 was observed to act as a positive regulator of AQP-related genes and to contribute to leaf water loss in tomato, which elucidates a crucial mechanistic role of auxin in this well-studied biological process.
In addition to its higher degree of water loss in leaves, 35S:mSlARF10-6 may be less resistant to drought. The rosseta strain containing the SlARF10-expressed protein showed lower growth than the control on the PEG-treated plate (Fig. S3). An earlier report found slower growth and developmental defects in young Arabidopsis mARF10 seedlings, which did not survive when directly grown in soil (Liu et al. 2007); thus, we assumed that an ARF10-mediated water loss mechanism may also exist in Arabidopsis. In Arabidopsis, AtARF10 and AtARF16 function as the activators of ABI3 transcription to positively regulate the ABA signal pathway, which contributes to the emerging map of the integration of hormone signalling in plant development and environmental adaption. In this study, we uncovered a previously unrecognized role of auxin in leaf water loss, namely, the promotion of leaf hydraulic and stomatal conductance through SlARF10. Therefore, our study establishes a molecular link between two important hormone pathways to water stress adaptation. miR160 would regulate ARF10 to maintain the normal leaf development and senescence process through the regulation of water status.
References
Acharya BR, Assmann SM (2009) Hormone interactions in stomatal function. Plant Mol Biol 69:451–462. doi:10.1007/s11103-008-9427-0
Aharon R, Shahak Y, Wininger S, Bendov R, Kapulnik Y, Galili G (2003) Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. Plant Cell 15:439–447. doi:10.1105/tpc.009225
Alexandersson E, Fraysse L, Sjövall-Larsen S, Gustavsson S, Fellert M, Karlsson M, Johanson U, Kjellbom P (2005) Whole gene family expression and drought stress regulation of aquaporins. Plant Mol Biol 59:469–484. doi:10.1007/s11103-005-0352-1
Barg R, Pilowsky M, Shabtai S, Carmi N, Szechtman AD, Dedicova B, Salts Y (1997) The TYLCV-tolerant tomato line MP-1 is characterized by superior transformation competence. J Exp Bot 48:1919–1923. doi:10.1093/jxb/48.11.1919
Bianchi MW, Damerval C, Vartanian N (2002) Identification of proteins regulated by cross-talk between drought and hormone pathways in Araidopsis wild-type and auxin-insensitive mutants, axr1 and axr2. Funt. Plant Biol 29:55–61
Bonetta D, McCourt P (1998) Genetic analysis of ABA signal transduction pathways. Trends Plant Sci 3:231–235. doi:10.1016/S1360-1385(98)01241-2
Brodribb TJ, Cochard H (2009) Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol 149:575–584. doi:10.1104/pp.108.129783
Brodribb TJ, Holbrook NM (2003) Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol 132:2166–2173. doi:10.1104/pp.103.023879
Clarke JM, Richards RA (1988) The effects of glaucousness, epicuticular wax, leaf age, plant height, and growth environment on water loss rates of excised wheat leaves. Can J Plant Sci 68:975–982. doi:10.4141/cjps88-118
Cochard H, Coll L, Le Roux X, Améglio T (2002) Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut. Plant Physiol 128:282–290. doi:10.1104/pp.010400
Cochard H, Venisse JS, Barigah TS, Brunel N, Herbette S, Guilliot A, Tyree MT, Sakr S (2007) Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol 143:122–133. doi:10.1104/pp.106.090092
Cunningham FX, Chamovitz D, Misawa N, Gantt E, Hirschberg J (1993) Cloning and functional expression in Escherichia coli of a cyanobacterial gene for lycopene cyclase, the enzyme that catalyzes the biosynthesis of beta-carotene. FEBS Lett 328:130–138. doi:10.1016/0014-5793(93)80980-9
Di Blasi S, Puliga S, Losi L, Vazzana C (1998) S. stapfianus and E. curvula cv. Consol in vivo photosynthesis, PSII activity and ABA content during dehydration. Plant Growth Regul 25:97–104
Ding Y, Tao Y, Zhu C (2013) Emerging roles of microRNAs in the mediation of drought stress response in plants. J Exp Bot 64:3077–3086. doi:10.1093/jxb/ert164
Finkelstein RR, Lynch TJ (2000) The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell 12:599–609
Finkelstein RR, Gampala SSL, Rock CD (2002) Abscisic acid signaling in seeds and seedlings. Plant Cell 14:515–545
Franks PJ, Beerling DJ (2009) CO2-forced evolution of plant gas exchange capacity and water-use efficiency over the Phanerozoic. Geobiology 7:227–236. doi:10.1111/j.1472-4669.2009.00193.x
Franks PJ, Farquhar GD (2001) The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant Physiol 125:935–942. doi:10.1104/pp.125.2.935
Franks PJ, Drake PL, Beerling DJ (2009) Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: an analysis using eucalyptus globulus. Plant Cell Environment 32:1737–1748. doi:10.1111/j.1365-3040.2009.02031.x
Fricke J, Hillebrand A, Twyman RM, Prüfer D, Schulze Gronover C (2013) Abscisic acid-dependent regulation of small rubber particle protein gene expression in Taraxacum brevicorniculatum is mediated by TbbZIP1. Plant Cell Physiol 54:448–464. doi:10.1093/pcp/pcs182
Gosti F, Beaudoin N, Serizet C, Webb AA, Vartanian N, Giraudat J (1999) ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell 11:1897–1909. doi:10.1105/tpc.11.10.1897
Guerin J, Rossel JB, Robert S, Tsuchiya T, Koltunow A (2000) A DEFICIENS homologue is down-regulated during apomictic initiation in ovules of hieracium. Planta 210:914–920. doi:10.1007/s004250050697
Guilfoyle TJ, Hagen G (2007) Auxin response factors. Curr Opin Plant Biol 10:453–460. doi:10.1016/j.pbi.2007.08.014
Hachez C, Moshelion M, Zelazny E, Cavez D, Chaumont F (2006a) Localization and quantification of plasma membrane aquaporin expression in maize primary root: a clue to understanding their role as cellular plumbers. Plant Mol Biol 62:305–323. doi:10.1007/s11103-006-9022-1
Hachez C, Zelazny E, Chaumont F (2006b) Modulating the expression of aquaporin genes in planta: a key to understand their physiological functions? Biochim Biophys Acta 1758:1142–1156. doi:10.1016/j.bbamem.2006.02.017
Heinen RB, Ye Q, Chaumont F (2009) Role of aquaporins in leaf physiology. J Exp Bot 60:2971–2985. doi:10.1093/jxb/erp171
Hendelman A, Buxdorf K, Stav R, Kravchik M, Arazi T (2012) Inhibition of lamina outgrowth following Solanum lycopersicum AUXIN RESPONSE FACTOR 10 (SlARF10) derepression. Plant Mol Biol 78:561–576. doi:10.1007/s11103-012-9883-4
Henzler T, Ye Q, Steudle E (2004) Oxidative gating of water channels (aquaporins) in Chara by hydroxyl radicals. Plant Cell Environ 27:1184–1195. doi:10.1111/j.1365-3040.2004.01226.x
Hose E, Steudle E, Hartung W (2000) Abscisic acid and hydraulic conductivity of maize roots: a study using cell- and root-pressure probes. Planta 211:874–882. doi:10.1007/s004250000412
Hunt L, Gray JE (2009) The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr Biol 19:864–869. doi:10.1016/j.cub.2009.03.069
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in plants. Annu Rev Plant Physiol Plant Mol Biol 47:377–403. doi:10.1146/annurev.arplant.47.1.377
Iuchi S, Kobayashi M, Taji T, Naramoto M, Seki M, Kato T, Tabata S, Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K (2001) Regulation of drought tolerance by gene manipulation of 0-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J 27:325–333
Jang JY, Kim DG, Kim YO, Kim JS, Kang H (2004) An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol Biol 54:713–725. doi:10.1023/B:PLAN.0000040900.61345.a6
Jefferies RA, Mackerron DKL (1989) Radiation interception and growth of irrigated and droughted potato (Solanum tuberosum). Field Crops Res 22:101–112. doi:10.1016/0378-4290(89)90061-0
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901–3907
Johnson DM, Meinzer FC, Woodruff DR, McCulloh KA (2009) Leaf xylem embolism, detected acoustically and by cryo-SEM, corresponds to decreases in leaf hydraulic conductance in four evergreen species. Plant Cell Environ 32:828–836. doi:10.1111/j.1365-3040.2009.01961.x
Kahn TL, Fender SE, Bray EA, O’Connell MA (1993) Characterization of expression of drought- and abscisic acid-regulated tomato genes in the drought-resistant species Lycopersicon pennellii. Plant Physiol 103:597–605
Kang JY, Choi HI, Im MY, Kim SY (2002) Arabidopsis basic leucine zipper proteins that mediate stress-responsive abscisic acid signaling. Plant Cell 14:343–357. doi:10.1105/tpc.010362
Kumar R, Tyagi AK, Sharma AK (2011) Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Mol Genet Genom 285:245–260. doi:10.1007/s00438-011-0602-7
Laur J, Hacke UG (2014) The role of water channel proteins in facilitating recovery of leaf hydraulic conductance from water stress in Populus trichocarpa. PLoS One 9:e111751. doi:10.1371/journal.pone.0111751
Li F, Vallabhaneni R, Wurtzel ET (2008) PSY3, a new member of the phytoene synthase gene family conserved in the Poaceae and regulator of abiotic stress-induced root carotenogenesis. Plant Physiol 146:1333–1345. doi:10.1104/pp.107.111120
Lian HL, Yu X, Lane D, Sun WN, Tang ZC, Su WA (2006) Upland rice and lowland rice exhibited different PIP expression under water deficit and ABA treatment. Cell Res 16:651–660. doi:10.1038/sj.cr.7310068
Liu PP, Montgomery TA, Fahlgren N, Kasschau KD, Nonogaki H, Carrington JC (2007) Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages. Plant J 52:133–146. doi:10.1111/j.1365-313X.2007.03218.x
Liu D, Song Y, Chen Z, Yu D (2009) Ectopic expression of miR396 suppresses GRF target gene expression and alters leaf growth in Arabidopsis. Physiol Plant 136:223–236. doi:10.1111/j.1399-3054.2009.01229.x
Liu X, Huang J, Wang Y, Khanna K, Xie Z, Owen HA, Zhao D (2010) The role of floral organs in carpels, an Arabidopsis loss-of-function mutation in microRNA160a, in organogenesis and the mechanism regulating its expression. Plant J 62:416–428. doi:10.1111/j.1365-313X.2010.04164.x
Liu X, Zhang H, Zhao Y, Feng Z, Li Q, Yang HQ, Luan S, Li J, He ZH (2013) Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc Natl Acad Sci USA 110:15485–15490. doi:10.1073/pnas.1304651110
Liu N, Wu S, Van Houten J, Wang Y, Ding B, Fei Z, Clarke TH, Reed JW, van der Knaap E (2014) Down-regulation of AUXIN RESPONSE FACTORS 6 and 8 by microRNA167 leads to floral development defects and female sterility in tomato. J Exp Bot 65:2507–2520. doi:10.1093/jxb/eru141
Lopez-Molina L, Mongrand S,Chua NH (2001) A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Natl Acad Sci 98:4782–4787
Ma JF, Tamai K, Yamaji N, Mitani N, Konishi S, Katsuhara M, Ishiguro M, Murata Y, Yano M (2006) A silicon transporter in rice. Nature 440:688–691. doi:10.1038/nature04590
Ma Y, Szostikiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E (2009) Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324:1064–1068. doi:10.1126/science.1172408
Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen YJ, Chen Z, Dewell SB, Du L, Fierro JM, Gomes XV, Godwin BC, He W, Helgesen S, Ho CH, Ho CH, Irzyk GP (2005) Genome sequencing in microfabricated high-density picolitre reactors. Nature 437:376–338. doi:10.1038/nature03959
Martre P, Morillon R, Barrieu F, North GB, Nobel PS, Chrispeels MJ (2002) Plasma membrane aquaporins play a significant role during recovery from water deficit. Plant Physiol 130:2101–2110. doi:10.1104/pp.009019
Maurel C, Verdoucq L, Luu DT, Santoni V (2008) Plant aquaporins: membrane channels with multiple integrated functions. Annu Rev Plant Biol 59:595–624. doi:10.1146/annurev.arplant.59.032607.092734
Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J (2001) The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J 25:295–303. doi:10.1046/j.1365-313x.2001.00965.x
Morillon R, Chrispeels MJ (2001) The role of ABA and the transpiration stream in the regulation of the osmotic water permeability of leaf cells. Proc Natl Acad Sci USA 98:14138–14143. doi:10.1073/pnas.231471998
Mulroy TW, Rundel PW (1977) Annual plants: adaptations to desert environments. Bioscience 27:109–114. doi:10.2307/1297607
Nardini A, Gortan E, Salleo S (2005) Hydraulic efficiency of the leaf venation system in sun- and shade-adapted species. Funct Plant Biol 32:953–961. doi:10.1071/FP05100
Nimchuk Z, Marois E, Kjemtrup S, Leister RT, Katagiri F, Dangl JL (2000) Eukaryotic fatty acylation drives plasma membrane targeting and enhances function of several type III effector proteins from Pseudomonas syringae. Cell 101:353–363. doi:10.1016/S0092-8674(00)80846-6
Nizampatnam NR, Schreier SJ, Damodaran S, Adhikari S, Subramanian S (2015) microRNA160 dictates stage-specific auxin and cytokinin sensitivities and directs soybean nodule development. Plant J 84:140–153. doi:10.1111/tpj.12965
Parent B, Hachez C, Redondo E, Simonneau T, Chaumont F, Tardieu F (2009) Drought and abscisic acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: a trans-scale approach. Plant Physiol 149:2000–2012. doi:10.1104/pp.108.130682
Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, Alfred SE, Bonetta D, Finkelstein R, Provart NJ, Desveaux D, Rodriguez PL, McCourt P, Zhu JK, Schroeder JI, Volkman BF (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of start proteins. Science 324:1068–1071. doi:10.1126/science.1173041
Peleg Z, Blumwald E (2011) Hormone balance and abiotic stress tolerance in crop plants. Curr Opin Plant Biol 14:290–295. doi:10.1016/j.pbi.2011.02.001
Pieruschka R, Huber G, Berry JA (2010) Control of transpiration by radiation. Proc Natl Acad Sci USA 107:13372–13377. doi:10.1073/pnas.0913177107
Pillitteri LJ, Sloan DB, Bogenschutz NL, Torii KU (2007) Termination of asymmetric cell division and differentiation of stomata. Nature 445:501–505. doi:10.1038/nature05467
Pou A, Medrano H, Flexas J, Tyerman SD (2013) A putative role for TIP and PIP aquaporins in dynamics of leaf hydraulic and stomatal conductances in grapevine under water stress and re-watering. Plant Cell Environ 36:828–843. doi:10.1111/pce.12019
Raphael M, Maarten JC (2001) The role of ABA and the transpiration stream in the regulation of the osmotic water permeability of leaf cells. Plant Biol 98:14138–14143
Raschke K (1987) Action of abscisic acid on guard cells. In: Zeiger E, Farquhar GD, Cowan IR (eds) Stomatal function. Stanford University Press, Stanford, CA, pp 253–279
Royer DL (2001) Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration. Rev Palaeobot Palynol 114:1–28. doi:10.1016/S0034-6667(00)00074-9
Ruiz-Sola M, Arbona V, Gomez-Cadenas A, Rodriguez-Concepcion M, Rodriguez-Villalon A (2014) A root specific inductio of carotenoid biosynthesis contributes to ABA production upon salt stress in Arabidopsis. PLoS One 9:e90765. doi:10.1371/journal.pone.0090765
Sack L, Holbrook NM (2006) Leaf hydraulics. Annu Rev Plant Biol 57:361–381. doi:10.1146/annurev.arplant.56.032604.144141
Sack L, Scoffoni (2012) Measurement of leaf hydraulic conductance and stomatal conductance and their responses to irradiance and dehydration using the evaporative flux method(EFM). J Vis Exp 70:e4179. doi:10.3791/4179.
Sack L, Cowan PD, Jaikumar N, Holbrook NM (2003) The ‘hydrology’ of leaves: co-ordination of structure and function in temperate woody species. Plant Cell Environ 26:1343–1356. doi:10.1046/j.0016-8025.2003.01058.x
Saito S, Hirai N, Matsumoto C, Ohigashi H, Ohta D, Sakata K, Mizutani M (2004) Arabidopsis CYP707As encode (+)-abscisic acid 8. Plant Physiol 134:1439–1449. doi:10.1104/pp.103.037614
Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D (2001) Guard cell signal transduction. Annu Rev Plant Physiol Plant Mol Biol 52:627–658. doi:10.1146/annurev.arplant.52.1.627
Scoffoni C, Rawls M, Mckown A, Cochard H, Sack L (2011) Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture. Plant Physiol 156:832–843. doi:10.1104/pp.111.173856
Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesis in plants. Trends Plant Sci 7:41–47. doi:10.1016/S1360-1385(01)02187-2
Serna L (2007) bHLH proteins know when to make a stoma. Trends Plant Sci 12:483–485. doi:10.1016/j.tplants.2007.08.016
Shatil-Cohen A, Attia Z, Moshelion M (2011) Bundle-sheath cell regulation of xylem-mesophyll water transport via aquaporins under drought stress: a target of xylem-borne ABA? Plant J 67:72–80. doi:10.1111/j.1365-313X.2011.04576.x
Shi H, Chen L, Ye T, Liu X, Ding K, Chan Z (2014) Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol Biochem 82:209–217. doi:10.1016/j.plaphy.2014.06.008
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58:221–227. doi:10.1093/jxb/erl164
Sun L, Wang YP, Chen P, Ren J, Ji K, Li Q, Li P, Dai SJ, Leng P (2011) Transcriptional regulation of SlPYL, SlPP2C, and SlSnRK2 gene families encoding ABA signal core components during tomato fruit development and drought stress. J Exp Bot 62:5659–5669. doi:10.1093/jxb/err252
Thompson AJ, Andrews J, Mulholland BJ, McKee JM, Hilton HW, Horridge JS, Farquhar GD, Smeeton RC, Smillie IR, Black CR, Taylor IB (2007) Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol 143:1905–1917. doi:10.1104/pp.106.093559
Turner M, Nizampatnam NR, Baron M, Coppin S, Damodaran S, Adhikari S, Arunachalam SP, Yu O, Subramanian S (2013) Ectopic expression of miR160 results in auxin hypersensitivity, cytokinin hyposensitivity, and inhibition of symbiotic nodule development in soybean. Plant Physiol 162:2042–2055. doi:10.1104/pp.113.220699
Tyerman SD, Niemietz CM, Bramley H (2002) Plant aquaporins: multifunctional water and solute channels with expanding roles. Plant Cell Environ 25:173–194. doi:10.1046/j.0016-8025.2001.00791.x
Uehlein N, Lovisolo C, Siefritz F, Kaldenhoff R (2003) The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature 425:734–737. doi:10.1038/nature02027
Ulmasov T, Hagen G, Guilfoyle TJ (1999) Activation and repression of transcription by auxin-response factors. Proc Natl Acad Sci USA 96:5844–5849. doi:10.1073/pnas.96.10.5844
Voicu MC, Zwiazek JJ, Tyree MT (2008) Light response of hydraulic conductance in bur oak (Quercus macrocarpa) leaves. Tree Physiol 28:1007–1015. doi:10.1093/treephys/28.7.1007
Voicu MC, Cooke JE, Zwiazek JJ (2009) Aquaporin gene expression and a poplastic water flow in bur oak (Quercus macrocarpa) leaves in relation to the light response of leaf hydraulic conductance. J Exp Bot 60:4063–4075. doi:10.1093/jxb/erp239
Wan X, Steudle E, Hartung W (2004) Gating of water channels (aquaporins) in cortical cells of young corn roots by mechanical stimuli (pressure pulses): effects of ABA and of HgCl2. J Exp Bot 55:411–422. doi:10.1093/jxb/erh051
Wang Y, Chen X, Xiang C (2007) Stomatal density and bio-water saving. J Integr Plant Biol 49:1435–1444. doi:10.1111/j.1672-9072.2007.00554.x
Wang L, Hua D, He J, Duan Y, Chen Z, Hong X, Gong Z (2011) Auxin response factor 2 (ARF2) and its regulated homeodomain gene HB33 mediate abscisic acid response in Arabidopsis. PLoS Genet 7:e1002172. doi:10.1371/journal.pgen.1002172
Wroblewski T, Tomczak A, Michelmore R (2005) Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol J 3:259–273. doi:10.1111/j.1467-7652.2005.00123.x
Wu J, Wang F, Cheng L, Kong F, Peng Z, Liu S, Yu X, Lu G (2011) Identification, isolation and expression analysis of auxin response factor (ARF) genes in Solanum lycopersicum. Plant Cell Rep 30:2059–2073. doi:10.1007/s00299-011-1113-z
Yang J, Zhang J, Wang Z, Zhu Q, Wang W (2001) Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiol 127:315–323. doi:10.1104/pp.127.1.315
Yang J, Zhang J, Wang Z, Zhu Q, Liu L (2002) Abscisic acid and cytokinins in the root exudates and leaves and their relationship to senescence and remobilization of carbon reserves in rice subjected to water stress during grain filling. Planta 215:645–652. doi:10.1007/s00425-002-0789-2
Yang X, Yang YN, Xue LJ, Zou MJ, Liu JY, Chen F, Xue HW (2011) Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. Plant Physiol 156:1397–1409. doi:10.1104/pp.111.173427
Zeevaart JAD, Yang SH (2005) Abscisic acid metabolism. Proceedings of the of the 32nd Annual Meeting of the Plant Growth Regulation Society of America, Newport Beach, CA, pp 1–5
Zhao HL, Ma RK, Jia XL, Hou LB, Liu EC (2009) Difference of water loss rate and its response to water supply of different winter wheat varieties. Acta Agri Boreali-Sinica 24:153–157
Zhou X, Yuan F, Wang M, Guo A, Zhang Y, Xie CG (2013) Molecular characterization of an ABA insensitive 5 orthologue in Brassica oleracea. Biochem Biophys Res Commun 430:1140–1146. doi:10.1016/j.bbrc.2012.12.023
Zou M, Guan Y, Ren H, Zhang F, Chen F (2007) Characterization of alternative splicing products of bZIP transcription factors OsABI5. Biochem Biophys Res Commun 360:307–313. doi:10.1016/j.bbrc.2007.05.226
Zou M, Guan Y, Ren H, Zhang F, Chen F (2008) A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol Biol 66:675–683. doi:10.1007/s11103-008-9298-4
Acknowledgments
We greatly thank T. Arazi and his colleagues from the Institute of Plant Sciences of the Agricultural Research Organization, Volcani Centre, Israel, for their kindness in providing the ‘35S:SlARF10’ and ‘35S:mSlARF10’ transgenic materials.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 31572167, 31272153 and 31171966), the Cultivation Plan for Youth Agricultural Science and Technology Innovative Talents of Liaoning Province (No. 2014051) and the Priority Development Area Foundation of the Ministry of Education of China (No. 20132103130002).
Author contributions
X. Liu, T. Xu and T. Li designed the research; X. Liu, X. Dong and Z. Liu performed experiments; X. Liu, Z. Shi, Y. Jiang and M. Qi analysed data; X. Liu and T. Xu wrote the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
We declare that we do not have any commercial or associative interests that represent a conflict of interest in connection with the work submitted.
Additional information
Xin Liu and Xiufen Dong have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Liu, X., Dong, X., Liu, Z. et al. Repression of ARF10 by microRNA160 plays an important role in the mediation of leaf water loss. Plant Mol Biol 92, 313–336 (2016). https://doi.org/10.1007/s11103-016-0514-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-016-0514-3