
Abstract
The number of c-Fos protein-like immunoreactive (c-Fos-IR) neurons in the spinal dorsal horn evoked by noxious stimulation was previously shown to be increased following peripheral nerve injury, and this increase was proposed to reflect the neuropathic pain state. The aim of this study was to investigate whether anomalous convergent primary afferent input to spinal dorsal horn neurons contributed to nerve injury-induced c-Fos hyperinducibility. Double immunofluorescence labeling for c-Fos and phosphorylated extracellular signal-regulated kinase (p-ERK) was performed to detect convergent synaptic input from different branches of the sciatic nerve after injury to the tibial nerve. c-Fos expression and the phosphorylation of ERK were induced by noxious heat stimulation of the hindpaw and also by electrical stimulation (ES) of the injured tibial nerve, respectively. The number of c-Fos-IR neurons was significantly decreased 3 days after the injury. However, the number of c-Fos-IR neurons returned to the control level 14 days after the injury. P-ERK immunoreactive (p-ERK-IR) neurons were induced in the central terminal field of the tibial nerve by ES of the tibial nerve. The topographic distribution pattern and number of such p-ERK-IR neurons remained unchanged after the nerve injury. The time course of changes in the number of double-labeled neurons, that presumably received convergent primary afferent input, showed a pattern similar to that of c-Fos-IR neurons after the injury. These results indicate that convergent primary nociceptive input through neighboring intact nerves may contribute to c-Fos hyperinducibility in the spinal dorsal horn.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Peripheral nerve injury has been shown to induce abnormal pain sensations such as hyperalgesia and allodynia. In addition to pain as a direct result of the injury itself, the manifestation of aberrant chronic pain has been reported within as well as outside the peripheral territory of the injured nerve. Previous studies demonstrated that the somatotopy map of second order neurons in the spinal dorsal horn was disrupted and also that neurons disconnected from their original receptive fields began to respond to stimulation of the skin outside their original receptive field [1–4]. The transection of a peripheral nerve was shown to enhance neuronal c-Fos induction in the spinal and medullary dorsal horns in response to the stimulation of spared primary nociceptors [5, 6]. We recently reported exaggerated c-Fos expression in the medullary dorsal horn evoked by electrical stimulation (ES) at the C-fiber intensity of a spared uninjured nerve rather than the injured nerve itself [7]. These changes may be responsible for hyperalgesia expanding to the area adjacent to those innervated by the injured nerve. Although the mechanism underlying hypersensitivity in the surrounding area innervated by the injured nerve has not yet been elucidated in detail, it may be attributed to the reorganization of primary afferents in the central nervous system (CNS). We hypothesize that peripheral nerve injury activates convergent nociceptive input from spared intact nerves, and such anomalous convergence contributes to the c-Fos hyperinducibility and hyperalgesia.
Extracellular signal-regulated kinase (ERK) is a member of the mitogen-activated protein kinase (MAPK) family. The phosphorylation of ERK was previously shown to be involved in the neuronal plasticity responsible for learning and memory and also pain hypersensitivity [8, 9]. Similar to c-Fos expression, phosphorylated ERK (p-ERK) expression was shown to be induced in spinal and medullary dorsal horn neurons by noxious stimulation [10, 11]. Therefore, p-ERK expression has also been used as a marker for neuronal activation following noxious stimulation. However, p-ERK expression in dorsal horn neurons following noxious stimulation was shown to be markedly quicker and more transient than that of c-Fos [10, 12].
The aim of this study was to examine whether the development of anomalous convergent primary afferent input occurred to spinal dorsal horn neurons disconnected from their normal receptive field. Double immunofluorescence labeling for c-Fos and p-ERK was performed to detect convergent synaptic inputs after the nerve injury. c-Fos expression and phosphorylation of ERK were induced by noxious heat stimulation of the hindpaw and also by ES of the tibial nerve, respectively.
Materials and Methods
Animals
All surgical and experimental procedures described herein were reviewed and approved by the Animal Care and Use Committee, Okayama University, Government Animal Protection and Management Law (No. 105), Japanese Government Notification on Feeding and Safekeeping of Animals (No. 6) and the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23), revised 1996. Male Sprague–Dawley rats weighing 180–200 g at the time of surgery were used in this study. Rats were housed at 20 °C with a daily light period of 12 h, and fed food and water ad libitum. Every attempt was made to minimize the number of animals used and reduce their suffering at all stages of the study.
Surgery
A tibial nerve injury was performed under anesthesia with an i.p. injection of pentobarbital sodium (50 mg/kg) as follows. The sciatic nerve was exposed between the mid-thigh level and the popliteal fossa by a skin incision and blunt dissection through the biceps femoris muscle. The three major divisions of this nerve (the tibial, sural, and common peroneal nerves) were separated in the proximal part of this segment by their individual perineuria. The tibial nerve was ligated firmly at two separate points with 7-0 silk suture, and the nerve bundle between the two ligatures was transected with fine scissors. Sham-operated animals, in which the tibial nerve was exposed without nerve ligation or transection, were used as controls.
Double Immunofluorescence Labeling for c-Fos and p-ERK
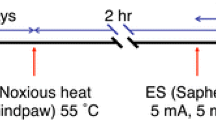
Double immunofluorescence labeling was used to detect convergent synaptic input to second order neurons. In this model, c-Fos expression and the phosphorylation of ERK were induced by noxious heat stimulation to the hindpaw and also by ES of the tibial nerve, respectively. c-Fos/p-ERK double-labeled cell profiles, which appeared after the nerve injury, were considered to have received convergent primary afferent inputs from intact and injured nerves. 3, 7 and 14 days after the tibial nerve injury or sham surgery (n = 5 in each group), re-anesthetized rats received noxious heat stimulation to the hindpaw 2 h before perfusion and the injured or uninjured tibial nerve was subsequently exposed for ES 15 min prior to perfusion (Fig. 1). The hindpaw was immersed in hot water (55 °C) for 10 s for noxious heat stimulation. Silver hock electrodes were placed beneath the isolated tibial nerve for ES, and a train of rectangular pulses (5 mA, 5 ms) was delivered at 5 Hz for 10 min. These stimulation parameters were previously determined as sufficient for exciting C-fibers [7, 13–17]. Rats were perfused transcardially with saline followed by 4 % paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4) 2 h after noxious heat stimulation and 5 min after ES (Fig. 1). The spinal cord of the 4th and 5th lumbar segment was dissected out, postfixed in the same fixative for 24 h and then immersed in 20 % sucrose in 0.02 M phosphate-buffered saline (PBS, pH 7.4) for 48 h.

Time course of surgery, noxious heat stimulation, ES of the tibial nerve, and perfusion for double immunofluorescence labeling
Frozen 10 µm-thick sections of the spinal cord were cut on a cryostat and mounted onto silane-coated slides. Sections were incubated for 48 h at RT with a mixture of the rabbit polyclonal anti-c-Fos antibody (1:8,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and mouse monoclonal anti-phospho-p44/42 MAP kinase antibody (1:2,000; Cell Signaling, Beverly, MA, USA). Alexa-488 and Alexa-568 (Molecular Probes, Eugene, OR, USA; 1:1,000)-conjugated secondary antibodies were used to visualize primary antibody binding. The primary antibody was omitted for negative controls. Sections were coverslipped with a mounting medium (DAKO Fluorescent Mounting Medium, DAKO, Carpinteria, CA, USA). The labeled sections were examined with a Nikon (Tokyo, Japan) fluorescence microscope, and images were captured with a CCD spot camera. The analysis is based on previous observations [5, 18] that medial 1/3 (tibial territory) of the superficial dorsal horn around the junction of 4th and 5th lumbar segments receives primary projection from the tibial nerve that innervate the planter surface of the hindpaw, whereas the lateral 1/4 is innervate by primary neurons supplying the posterior cutaneous nerve of the thigh. The area between them (peroneal territory) receives primary projection from the superficial peroneal and sural nerves that innervate the dorsal and lateral aspects of the hindpaw. The numbers of c-Fos protein-like immunoreactive (c-Fos-IR), p-ERK immunoreactive (p-ERK-IR), and c-Fos/p-ERK double-labeled cell profiles were counted separately for tibial and peroneal territories in laminae I and II (I/II) of the spinal dorsal horn. Five sections within 1 mm rostral or caudal to the junction of 4th and 5th lumbar segments were randomly selected for statistical analysis and the average of five sections was recorded for each rat. To confirm the reliability of double immunofluorescence labeling, a single stimulation, either noxious heat stimulation or ES to the tibial nerve, was conducted in a separate group of animals (n = 3 in each stimulation group) and processed for double immunofluorescence labeling.
Statistical Analysis
Results are presented as the mean ± SEM. Statistical analysis was performed using a one-way analysis of variance (ANOVA) followed by the post hoc Tukey–Kramer test or Student’s t test. The criterion used for significance was p < 0.05.
Results
Many dorsal horn cells exhibited immunofluorescence labeling for c-Fos and p-ERK following noxious heat stimulation to the hindpaw and ES to the injured or uninjured tibial nerve. Figure 2 shows examples of single- and double-labeled cell profiles in the spinal dorsal horn. Immunoreactivity for c-Fos was confined within the neuronal nucleus and yielded intense red fluorescence (Fig. 2a). Immunoreactivity for p-ERK spread over the neuronal cytoplasm and appeared as green labeling (Fig. 2b). Therefore, we tentatively referred to these profiles as c-Fos-IR neurons and p-ERK-IR neurons hereafter. Double-labeled neurons were distinguished by their green labeling in the cytoplasm and intense red or yellow nuclear labeling (Fig. 2c). These double-labeled neurons were considered to have responded to both noxious heat stimulation and ES because double-labeled neurons were rarely observed in sections after a single stimulation; i.e., noxious heat stimulation alone yielded red nuclear labeling for c-Fos, whereas ES of the tibial nerve only yielded cytoplasmic green labeling for p-ERK (Fig. 2d, e). In other words, immunoreactivity for c-Fos and p-ERK represented the responses to noxious heat stimulation and ES, respectively. The number of single labeled neurons for c-Fos or p-ERK after a single stimulation was similar to that after double stimulation (data not shown). These results indicate that noxious heat stimulation does not affect p-ERK expression evoked by ES to the tibial nerve and conversely, ES does not affect c-Fos expression evoked by noxious heat stimulation.

Examples of c-Fos-IR, p-ERK-IR, and double-labeled neurons. Immunofluorescence was seen for c-Fos (red; a) and p-ERK (green; b) in the dorsal horn following noxious heat stimulation of the hindpaw and ES of the tibial nerve in sham-operated animals. A merged image is shown in (c). Arrowheads indicate the nuclei of c-Fos-IR neurons. Arrows indicate the cell bodies of p-ERK-IR neurons. Double-labeled neurons are indicated by double-arrowheads. Single stimulation, either noxious heat stimulation to the hindpaw (d) or ES to the tibial nerve (e), resulted in either c-Fos or p-ERK immunoreactivity. Scale bars = 50–100 µm in (c, e), respectively
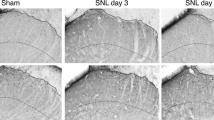
Double immunofluorescence labeling for c-Fos and p-ERK following noxious heat stimulation and ES was performed at several time points following the tibial nerve injury. Single- and double-labeled neurons were the most abundant in the medial 1/2 of laminae I/II in sham-operated or nerve-injured animals irrespective of the post-injury interval (Fig. 3). Labeled neurons were rarely found in the deeper laminae III/IV. Along the rostro-caudal axis, labeled neurons were concentrated in the caudal part of L4 and rostral part of L5 (data not shown). This distribution pattern was consistent with the previously reported central terminal field of primary neurons in the sciatic nerve [18]. The most lateral part of the dorsal horn, which corresponded to the terminal field of primary neurons supplying the posterior cutaneous nerve of the thigh, was almost devoid of immunoreactivity.

Merged images of double immunofluorescence for c-Fos (red) and p-ERK (green) in the spinal dorsal horn following noxious heat stimulation to the hindpaw and ES of the tibial nerve 3, 7, and 14 days after a tibial nerve injury and sham surgery. Representative data from one animal in each group are shown. Arrowheads indicate double-labeled neurons in the spinal dorsal horn. c-Fos-IR and double-labeled neurons decreased in number, especially in the tibial territory of the dorsal horn, 3 and 7 days after the injury. However, a marked recovery in the number of c-Fos-IR and double-labeled neurons was seen 14 days after the injury. Tibial, tibial territory; Peroneal, peroneal territory; I/II, laminae I and II; III/IV, lamimae III and IV. Scale bar = 100 µm
We counted c-Fos-IR, p-ERK-IR and double-labeled neurons in laminae I/II at the level of the L4 and L5 junction. Double-labeled neurons were included in the populations of both c-Fos-IR and p-ERK-IR neurons. The number of c-Fos-IR neurons 3 and 7 days after the nerve injury was smaller than that in sham-operated rats. The c-Fos-IR neurons induced were restricted to the peroneal territory and the tibial territory was almost devoid of c-Fos-IR neurons. However, the induced c-Fos-IR neurons reappeared in the tibial territory 14 days after the nerve injury, and the number of c-Fos-IR neurons was significantly higher than that 3 days after the injury (Figs. 3, 4). ES of the tibial nerve induced a large number of p-ERK-IR neurons in the superficial dorsal horn ipsilateral to the ES, however, the number of these neurons remained unchanged after the nerve injury (Figs. 3, 4). c-Fos-IR and p-ERK-IR neurons were rarely found on the contralateral side and a change in the number of labeled neurons was not detected (data not shown). Double-labeled neurons for c-Fos and p-ERK were distributed mainly in the tibial territory in the superficial laminae of the spinal dorsal horn on the ipsilateral side. The number of double-labeled neurons was significantly lower in rats 3 and 7 days after the nerve injury than that in sham-operated controls. However, a large number of double-labeled neurons reappeared 14 days after the nerve injury. The number of double-labeled neurons was significantly higher 14 days after the injury than 3 days after (Figs. 3, 4). A small number of double-labeled neurons were also observed in the peroneal territory, however, the number of these neurons remained unchanged after the nerve injury (Figs. 3, 4).

The number per section of c-Fos-IR (top), p-ERK-IR (middle), and double-labeled neurons (bottom) in the tibial territory (left column) and the peroneal territory (right column) in the superficial laminae of the spinal dorsal horn following noxious heat stimulation to the hindpaw and ES of the tibial nerve 3, 7, and 14 days after a tibial nerve injury and sham surgery. c-Fos-IR and double-labeled neurons decreased in number 3 and 7 days after the injury. However, a significant increase was observed in the number of c-Fos-IR and double-labeled neurons 14 days after the injury. Each bar represents the mean value ± SEM of five individual experiments. The asterisk at each bar indicates a significant difference from the sham-operated group (***p < 0.001, ANOVA with the post hoc Tukey–Kramer test). Sharps indicate significant differences from the day 3 group (## p < 0.01, ANOVA with the post hoc Tukey–Kramer test)
Discussion
The present study examined the spinal dorsal horn in an attempt to elucidate the mechanisms underlying the development of neuropathic pain. Injury to the tibial nerve altered the c-Fos response to noxious heat stimulation to the hindpaw; the number of c-Fos-IR neurons induced was significantly reduced 3 and 7 days after the injury, and then increased, returning to the sham-operated control level by day 14. The number of double-labeled neurons for c-Fos and p-ERK following the injury showed a temporal distribution pattern similar to that for c-Fos-IR neurons. On the other hand, the number of p-ERK-IR neurons was not affected by the injury and remained at a level similar to the control level throughout the examined post-injury period.
Previous studies demonstrated that the distribution of c-Fos-IR neurons in the spinal and medullary dorsal horns following noxious mechanical and thermal stimulation of the skin was somatotopically organized and also that the number of such neurons increased in a stimulus intensity-dependent manner [6, 19–23]. In support of these findings, the distribution of c-Fos-IR neurons following noxious heat stimulation in sham-operated control rats was largely restricted to the medial half of the dorsal horn around the L4 and L5 junction. This distribution resembled the central terminal field of the primary neurons constituting the sciatic nerve [18]. The c-Fos-IR neurons induced were restricted to the peroneal territory 3 days after the injury to the tibial nerve, while those in the tibial territory (the medial part of the sciatic territory) were rare. We consider this to be the basal level of c-Fos induction by the nociceptive signal transmitted by the primary thermonociceptors of the peroneal nerve [5]. However, a significant number of induced c-Fos-IR neurons reappeared in the tibial territory 14 days after the injury, and the number of c-Fos-IR neurons in the tibial territory returned to a level similar to that in sham-operated rats. The time course of changes in the number of c-Fos-IR neurons was consistent with our recent study showing nocifensive responses to mechanical and noxious heat stimulation were attenuated 3 days after the nerve injury, recovered at 7 days, and were further augmented at 14 days [24]. Similarities in the time course of changes strongly suggest that the injury-induced increase in the c-Fos response reflected neuropathic pain including allodynia and thermal hyperalgesia. Since injured tibial nerve axons were prevented from regenerating by a tight ligature on the proximal nerve stump, the apparent recovery in the number of induced c-Fos-IR neurons should be considered to reflect the exaggerated response of secondary neurons to the nociceptive signal conveyed by the spared nerve. These changes may be involved in the expansion of hyperalgesia to the area adjacent to the denervated area. Consistent with these findings, our recent study showed that the local application of a selective adenosine A1 receptor agonist to the common peroneal nerve attenuated the c-Fos response evoked by noxious heat stimulation to the hindpaw and behavioral hyperalgesia after a tibial nerve injury [24].
MAPKs play an important role in intracellular signal transduction during the activation of neurons following various noxious stimuli [11, 25–27]. ERK was previously shown to be phosphorylated following noxious peripheral stimuli in an intensity-dependent manner [11, 26–29]. The phosphorylation of ERK was markedly increased within a few minutes, peaked after 5 min, and returned to control levels 2 h after noxious stimulation [10, 12]. Based on these findings, we utilized the phosphorylation of ERK as the second marker for the activation of nociceptive dorsal horn neurons. We observed a large number of induced p-ERK-IR neurons in the medial 1/2 of the spinal dorsal horn 5 min after ES of the tibial nerve. The number of p-ERK-IR neurons induced by ES remained unchanged after the nerve injury. These results are consistent with a previous finding in which noxious C-fiber intensity stimulation of a chronically injured nerve did not induce extraneous c-Fos-IR neurons beyond normal levels in the spinal or medullary dorsal horn [7, 15]. Therefore, a nerve injury may not enhance the transmission of a nociceptive signal from injured primary nociceptors.
Double-labeled neurons were considered to have responded to both noxious heat stimulation and ES of the tibial nerve. Both noxious heat stimulation to the hindpaw as well as ES to the tibial nerve were expected to have excited primary neurons in the tibial nerve of sham-operated controls. A large number of double-labeled neurons were found in the spinal dorsal horn. The number of double-labeled neurons was markedly decreased 3 and 7 days after the injury, and reappeared after 14 days. Since peripheral axons in the injured tibial nerve were prevented from reinnervating the skin receptive field, double labeling in nerve-injured rats was considered to represent the convergence of sensory signals from both tibial and peroneal nerves. The time course of changes in the number of double-labeled neurons matches that for c-Fos-IR neurons after the injury, suggesting that convergent nociceptive input may contribute to the c-Fos hyperinducibility after nerve injury. A possible explanation for this result may be that a nociceptive spinal dorsal horn neuron originally had convergent collateral primary input, which is normally insufficient to induce c-Fos up-regulation. Such a neuron may become responsive to a previously “silent” synaptic input when it loses the dominant input. Such “unmasking” of a previously silent input could result in an increase in c-Fos-IR neurons induced by noxious stimulation, and may also cause the referral and spread of pain. Indeed, chronic partial deafferentation by single dorsal rhizotomy induced c-Fos hyperinducibility in spinal dorsal horn neurons in response to convergent primary nociceptive input through neighboring intact dorsal root fibers [30].
In summary, we demonstrated that injury to the tibial nerve augmented c-Fos expression in the spinal dorsal horn induced by noxious heat stimulation to the hindpaw. Increased double immunofluorescence labeling for c-Fos and p-ERK suggested that collateral synaptic input to second order spinal dorsal horn neurons become active after the nerve injury and disconnection from their dominant receptive field. These results indicate that convergent nociceptive input through neighboring intact nerves may, at least in part, contribute to c-Fos hyperinducibility in the spinal dorsal horn and the pathogenesis of neuropathic pain following peripheral nerve injury.
Abbreviations
- ANOVA:
-
Analysis of variance
- CNS:
-
Central nervous system
- ES:
-
Electrical stimulation
- c-Fos-IR:
-
c-Fos protein-like immunoreactive
- MAPK:
-
Mitogen-activated protein kinase
- p-ERK:
-
Phosphorylated extracellular signal-regulated kinase
- p-ERK-IR:
-
p-ERK immunoreactive
References
Devor M, Wall PD (1978) Reorganisation of spinal cord sensory map after peripheral nerve injury. Nature 276(5683):75–76
Hylden JL, Nahin RL, Dubner R (1987) Altered responses of nociceptive cat lamina I spinal dorsal horn neurons after chronic sciatic neuroma formation. Brain Res 411(2):341–350
Lisney SJ (1983) Changes in the somatotopic organization of the cat lumbar spinal cord following peripheral nerve transection and regeneration. Brain Res 259(1):31–39
Markus H, Pomeranz B, Krushelnycky D (1984) Spread of saphenous somatotopic projection map in spinal cord and hypersensitivity of the foot after chronic sciatic denervation in adult rat. Brain Res 296(1):27–39
Sugimoto T, Ichikawa H, Hijiya H, Mitani S, Nakago T (1993) c-Fos expression by dorsal horn neurons chronically deafferented by peripheral nerve section in response to spared, somatotopically inappropriate nociceptive primary input. Brain Res 621(1):161–166
Nomura H, Ogawa A, Tashiro A, Morimoto T, Hu JW, Iwata K (2002) Induction of Fos protein-like immunoreactivity in the trigeminal spinal nucleus caudalis and upper cervical cord following noxious and non-noxious mechanical stimulation of the whisker pad of the rat with an inferior alveolar nerve transection. Pain 95(3):225–238
Fujisawa N, Terayama R, Yamaguchi D, Omura S, Yamashiro T, Sugimoto T (2012) Fos protein-like immunoreactive neurons induced by electrical stimulation in the trigeminal sensory nuclear complex of rats with chronically injured peripheral nerve. Exp Brain Res 219(2):191–201. doi:10.1007/s00221-012-3078-8
Ji RR, Kohno T, Moore KA, Woolf CJ (2003) Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci 26(12):696–705
Ji RR, Woolf CJ (2001) Neuronal plasticity and signal transduction in nociceptive neurons: implications for the initiation and maintenance of pathological pain. Neurobiol Dis 8(1):1–10. doi:10.1006/nbdi.2000.0360
Ji RR, Baba H, Brenner GJ, Woolf CJ (1999) Nociceptive-specific activation of ERK in spinal neurons contributes to pain hypersensitivity. Nat Neurosci 2(12):1114–1119. doi:10.1038/16040
Noma N, Tsuboi Y, Kondo M, Matsumoto M, Sessle BJ, Kitagawa J, Saito K, Iwata K (2008) Organization of pERK-immunoreactive cells in trigeminal spinal nucleus caudalis and upper cervical cord following capsaicin injection into oral and craniofacial regions in rats. J Comp Neurol 507(3):1428–1440. doi:10.1002/cne.21620
Shimizu K, Asano M, Kitagawa J, Ogiso B, Ren K, Oki H, Matsumoto M, Iwata K (2006) Phosphorylation of extracellular signal-regulated kinase in medullary and upper cervical cord neurons following noxious tooth pulp stimulation. Brain Res 1072(1):99–109. doi:10.1016/j.brainres.2005.12.040
Devor M, Govrin-Lippmann R (1983) Axoplasmic transport block reduces ectopic impulse generation in injured peripheral nerves. Pain 16(1):73–85
Hughes AS, Averill S, King VR, Molander C, Shortland PJ (2008) Neurochemical characterization of neuronal populations expressing protein kinase C gamma isoform in the spinal cord and gracile nucleus of the rat. Neuroscience 153(2):507–517. doi:10.1016/j.neuroscience.2008.01.082
Molander C, Hongpaisan J, Grant G (1992) Changing pattern of c-Fos expression in spinal cord neurons after electrical stimulation of the chronically injured sciatic nerve in the rat. Neuroscience 50(1):223–236
Shortland P, Molander C (1998) The time-course of abeta-evoked c-Fos expression in neurons of the dorsal horn and gracile nucleus after peripheral nerve injury. Brain Res 810(1–2):288–293
Tokunaga A, Kondo E, Fukuoka T, Miki K, Dai Y, Tsujino H, Noguchi K (1999) Excitability of spinal cord and gracile nucleus neurons in rats with chronically chronically injured sciatic nerve examined by c-Fos expression. Brain Res 847(2):321–331
Swett JE, Woolf CJ (1985) The somatotopic organization of primary afferent terminals in the superficial laminae of the dorsal horn of the rat spinal cord. J Comp Neurol 231(1):66–77. doi:10.1002/cne.902310106
Strassman AM, Vos BP (1993) Somatotopic and laminar organization of fos-like immunoreactivity in the medullary and upper cervical dorsal horn induced by noxious facial stimulation in the rat. J Comp Neurol 331(4):495–516
Strassman AM, Vos BP, Mineta Y, Naderi S, Borsook D, Burstein R (1993) Fos-like immunoreactivity in the superficial medullary dorsal horn induced by noxious and innocuous thermal stimulation of facial skin in the rat. J Neurophysiol 70(5):1811–1821
Bullitt E (1990) Expression of c-Fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J Comp Neurol 296(4):517–530
Hunt SP, Pini A, Evan G (1987) Induction of c-Fos-like protein in spinal cord neurons following sensory stimulation. Nature 328(6131):632–634
Terayama R, Nagamatsu N, Ikeda T, Nakamura T, Rahman OI, Sakoda S, Shiba R, Nishimori T (1997) Differential expression of Fos protein after transection of the rat infraorbital nerve in the trigeminal nucleus caudalis. Brain Res 768(1–2):135–146
Yamaguchi D, Terayama R, Omura S, Tsuchiya H, Sato T, Ichikawa H, Sugimoto T (2014) Effect of adenosine A1 receptor agonist on the enhanced excitability of spinal dorsal horn neurons after peripheral nerve injury. Int J Neurosci 124(3):213–222. doi:10.3109/00207454.2013.842566
Dai Y, Iwata K, Fukuoka T, Kondo E, Tokunaga A, Yamanaka H, Tachibana T, Liu Y, Noguchi K (2002) Phosphorylation of extracellular signal-regulated kinase in primary afferent neurons by noxious stimuli and its involvement in peripheral sensitization. J Neurosci 22(17):7737–7745
Ji RR, Befort K, Brenner GJ, Woolf CJ (2002) ERK MAP kinase activation in superficial spinal cord neurons induces prodynorphin and NK-1 upregulation and contributes to persistent inflammatory pain hypersensitivity. J Neurosci 22(2):478–485
Liu Y, Obata K, Yamanaka H, Dai Y, Fukuoka T, Tokunaga A, Noguchi K (2004) Activation of extracellular signal-regulated protein kinase in dorsal horn neurons in the rat neuropathic intermittent claudication model. Pain 109(1–2):64–72. doi:10.1016/j.pain.2004.01.010
Kawasaki Y, Kohno T, Zhuang ZY, Brenner GJ, Wang H, Van Der Meer C, Befort K, Woolf CJ, Ji RR (2004) Ionotropic and metabotropic receptors, protein kinase A, protein kinase C, and Src contribute to C-fiber-induced ERK activation and cAMP response element-binding protein phosphorylation in dorsal horn neurons, leading to central sensitization. J Neurosci 24(38):8310–8321. doi:10.1523/jneurosci.2396-04.2004
Wang H, Dai Y, Fukuoka T, Yamanaka H, Obata K, Tokunaga A, Noguchi K (2004) Enhancement of stimulation-induced ERK activation in the spinal dorsal horn and gracile nucleus neurons in rats with peripheral nerve injury. Eur J Neurosci 19(4):884–890
Sugimoto T, Hara T, Shirai H, Abe T, Ichikawa H, Sato T (1994) c-Fos induction in the subnucleus caudalis following noxious mechanical stimulation of the oral mucous membrane. Exp Neurol 129(2):251–256
Acknowledgments
This study was supported by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (24592764).
Conflict of interest
The authors do not have any conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Terayama, R., Kishimoto, N., Yamamoto, Y. et al. Convergent Nociceptive Input to Spinal Dorsal Horn Neurons After Peripheral Nerve Injury. Neurochem Res 40, 438–445 (2015). https://doi.org/10.1007/s11064-014-1484-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-014-1484-y