
Abstract
Ecological interactions, such as facilitation among plant species, has been identified as key for restoration actions. The stress-gradient hypothesis predicts that facilitation increases under harsh environments, but disturbance type and nurse shrub characteristics may modify this prediction. We aim at assessing the effect of two interacting factors (herbivory pressure and light availability) and palatability of the nurse shrub on the importance of nurse effect for active restoration of mixed evergreen forests of northern Patagonia (Argentina) for silvopastoral use. We planted Austrocedrus chilensis seedlings under two silvopastoral use intensities (higher and lower according to livestock seasonal movements) and under three micro-site treatments (palatable shrub, non-palatable shrub, inter-canopy), and evaluated survival and growth during three consecutive years. Under higher use intensity (increasing stress of light availability as desiccation risk and herbivory) we found contrasting effects of the nurse effect on seedling survival and growth, but the palatability of nurse shrub had no effect. Under higher use intensity, seedling survival was lower, while growth was higher. Higher values of seedling survival were found under low use intensity inter-canopy micro-site. Under high use, the presence of a nurse shrub is important for summer seedling survival, indicating its role in avoiding desiccation. The same occurred for winter seedling survival, where a nurse shrub may decrease mainly trampling risk. We highlight the importance of considering the interaction between disturbance factors for the net outcome of the nurse effect in active restoration actions, as it may allow for diverse actions to restoration.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Understanding the role of positive interactions among plant species has been identified as a key element for planning restoration actions (Gómez-Aparicio 2009; Barbosa and Asner 2017). Plants may compete for scarce resources but may also facilitate their neighbors through grazing protection and/or micro-climatic regulation such as shading. The stress-gradient hypothesis (SGH) predicts that the prevalence and strength of facilitative and competitive interactions varies inversely along a given stress gradient, with facilitation expected to be more common under higher than lower abiotic stress conditions (Bertness and Callaway 1994). For example, the nurse effect (facilitation of one species on the regeneration of another) has been found as a key mechanism for plant regeneration in harsh environments (Soliveres and Maestre 2014). Moreover, the use of the SGH frameworks is useful to predict potential interaction outcomes under two or more disturbances. In this sense, recent empirical research shows that the predictions of SGH may also depend on the type of stress (i.e. biotic or abiotic) and on the characteristics of the interacting species regarding the analyzed stress (i.e. palatable or non-palatable when grazing is considered as biotic stress) (Soliveres and Maestre 2014; Brooker et al. 2008; Maestre et al. 2009; Soliveres et al. 2011).
The consideration of more than one disturbance is a valuable aspect of the SGH, as environmental stresses are known to be complex, where sometimes biotic (i.e. herbivory) and abiotic (i.e. light availability) factors interact and the final balance of species interaction outcome will depend on the response of the interacting species (Smit et al. 2007). For example, for herbaceous species it has been proposed that if interacting species respond similarly to the abiotic stress, the negative effect of herbivory pressure may diminish with the increase of the abiotic stress (i.e. summer desiccation). In such cases, the importance of the nurse effect will be diminished (Smit et al. 2009). Moreover, the performance of a beneficiary in the presence of a benefactor (nurse) may also vary depending on certain characteristics of the beneficiary species (Maestre et al. 2009; Smit et al. 2009; Navarro-Cano et al. 2019). For example, under a stress factor of herbivory pressure, the nurse effect of non-palatable species on the regeneration of palatable species is well recognized, as it diminishes trampling and browsing risk (Vandenberghe et al. 2009; Peláez et al. 2019). Yet, the interaction between disturbance intensity and species characteristics in the extent of the nurse effect is less understood (Soliveres and Maestre 2014).
Considering how multiple stressors influence species interactions stands out as a key research need to understand the consequences of land-use changes on community structure and functioning (Beauchesne et al. 2021). Silvopastoral production, a common type of land-use in temperate forests (i.e. Patagonian forests), commonly consists of a complex interaction of two disturbances which are deliberately integrated: extensive cattle grazing and wood and timber extraction. When the regeneration of canopy species is analyzed, the nature of silvopastoral disturbance involves two different mechanisms: abiotic stress through a bottom-up mechanism where summer droughts increase desiccation risk, and biotic stress through a top-down mechanism driven by cattle browsing and trampling (Peri et al. 2016; Gordon et al. 2018). The complexity of this land-use stems from the fact that the nature and magnitude of stress factors on individuals and populations depend on species-specific responses (Beauchesne et al. 2021). For example, light availability may be a limiting resource for understory vegetation under closed canopy forests or a stress factor under high irradiation conditions in open canopy forests if species require nursing from others. In Patagonian temperate forests, the combined effect of both stress factors increases understory biodiversity and modifies community composition and ecosystem functioning (Chillo et al. 2018), but little is known about how these changes influence ecological interactions, an understudied consequence of land-use change. In particular, regeneration of Austrocedrus chilensis (D. Don) Pic. Serm. & Bizzarri (cordilleran cypress) naturally occurs in small gaps or even under closed canopy (Veblen et al. 1995; Dezzoti 1996; Amoroso and Larson 2010), and saplings are browsed by cattle (Blackhall et al. 2008).
In this sense, nurse effect has been recognized as crucial for the establishment and growth of A. chilensis seedlings, as it decreases water stress from evapotranspiration in dry summers (Kitzberger et al. 2000; Letourneau et al. 2004; Nuñez et al. 2009). But higher light availability may also represent higher resource availability. Thus, the combined effect of both stress factors on plant facilitation is still unclear. In Northwestern Patagonia (Argentina), livestock integrated into forest represents the greatest land-use (Amoroso et al. 2021). In situations where silvopastoral use has determined strong changes in forest structure, compromising the regeneration and future perpetuity of the resource, it is necessary to consider the implementation of active restoration practices to guarantee the establishment of regeneration (sensu Rey Benayas et al. 2009; Jones 2013). Although there are numerous experiences of active restoration of A. chilensis in pure and mixed forests after fires and high-grading (Oudkerk et al. 2003; Rovere 2008; Urretavizcaya et al. 2018, 2017, 2015; Urretavizcaya and Defossé 2019; Caselli et al. 2020), many aspects are unknown about forest active restoration under silvopastoral use.
Our main goal is to assess the effect of a complex land-use (i.e- silvopastoral use) on the relative importance of the nurse effect for active restoration of A. chilensis timber resource in mixed A. chilensis and Nothofagus dombeyi (Mirb) Blume forests in northern Patagonia, Argentina. To do so, we evaluated the nurse effect of shrubs of different palatability on the survival and growth of A. chilensis seedlings planted under two silvopastoral use intensities. First, we hypothesize that different silvopastoral use intensities modify the balance of the facilitative interaction (nurse effect), being more important under higher disturbance intensity. But this relationship will vary based on the palatability of the benefactor species. Thus, we predict that A. chilensis seedlings will experience higher survival and growth beneath a nurse shrub in higher than in lower silvopastoral use. Second, we hypothesize that under increasing abiotic stress (desiccation risk), the biotic stress factor (herbivory pressure) will be less important. We predict that under higher silvopastoral use intensity, there will be no effect of the nurse shrub palatability on survival and growth of A. chilensis seedlings.
Methods
Study area
We studied understory plant communities in mixed forest of A. chilensis and N. dombeyi of north-western Patagonia, in El Manso river basin region (Río Negro, Argentina). These forests are characterized by a dense stratified evergreen canopy cover (> 90%) with trees up to 30 m high. The relative dominance of the main species changes based on landscape factors such as soil depths, slope and exposure (Amoroso et al. 2018). Climate is characterized by cold wet winters and mild dry summers, with mean annual temperature of 9.3 °C and mean annual precipitation of 900 mm. In this area, summer is considered a dry season, as evapotranspiration deficits occur all through summer (Madariaga 2019). Soils are Andisols with deep profiles containing volcanic ash. This soil type enables the occurrence of the forest, as it stores moisture from the rainy season (winter) and releases it throughout the dry season (summer).
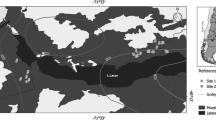
The traditional cattle production system in these forests consists of moving cattle seasonally through different elevations and forest types (Cardozo 2014). In particular, during summer cattle are moved to highlands, while during winter cattle returns and remains in middle and lowlands dominated by mixed evergreen forests. Within the forest, cattle graze mainly in open canopy areas (< 50% cover) generated by partial and selective wood and timber extraction. These open areas are known as high silvopastoral use intensity, hereafter “high use” sites (Fig. 1). These high use sites represent sites with the highest livestock and logging intensity use found within the forest. The canopy of the surrounding forest is partly closed (50–90% cover) as less timber extraction occurs. These areas are less used by cattle and are known as low silvopastoral use intensity, hereafter “low use” sites (Fig. 1). These low use sites could be considered as the lowest use within the mixed evergreen forests; lower use areas can be found but far away and in other forest types. During summer cattle are moved to deciduous highland forests and, thus, no herbivory pressure occurs in middle and low-land mixed evergreen forests during the growing season. Thus, the use intensity represents a complex land-use where two stress factors interact: herbivory pressure through a top-down mechanism, and light availability through a bottom-up mechanism (see Arias Sepúlveda and Chillo 2017; Amoroso et al. 2018 and Chillo et al. 2018 for further details of sites characteristics).

Schematic representation of the experimental design with two levels of analysis: use intensity and micro-site. Sites are mixed forests of A. chilensis (light grey canopy) and N. dombeyi (dark grey canopy) under low (1) and high (2) use intensity. Micro-sites designed to evaluate nurse effect consisted of three treatments: P—planted seedling (in black) under palatable nurse shrub, I—planted seedling at the inter-canopy (open-condition, no nurse shrub), and N—planted seedling under non-palatable nurse shrub
Experimental and sampling design
Within this complex land-use system, we established an experimental design with two treatments (Fig. 1): (A) Silvopastoral use intensity, to evaluate the effect of silvopastoral use intensity on the nurse effect of shrubs on A. chilensis seedlings growth and survival; (B) Nurse shrub identity, to evaluate the role of palatability on the nurse effect on A. chilensis seedlings growth and survival. The scale of analysis in this experimental design is the micro-site level, in order to assess the micro-site effect on A. chilensis seedling survival and growth.
Thus, within each silvopastoral use treatment (low and high use intensity), the experiment was conducted at different micro-sites representing three levels of nurse shrub identity treatment: below canopy of a palatable shrub (P), below canopy of a non-palatable shrub (N), and the inter-canopy (open condition) without nurse effect (I) (Fig. 1). To select these micro-sites we searched for shrubs at least 1 m tall, of the most common and abundant species in these forests (mainly Aristotelia chilensis, Schinus patagonicus and Lomatia hirsuta). The condition of palatable and non-palatable was determined based on the relative palatability of the species present at each site. Shrubs considered palatable were mainly Aristotelia chilensis, S. patagonicus and Myoschilos oblongum (Gowda et al. 2014). S. patagonicus is considered a moderately palatable species (Blackhall et al. 2015), and it was chosen only when Aristotelia chilensis was absent. Non-palatable shrubs were mainly L. hirsuta and Berberis sp., and sometimes Rosa rubiginosa (Gowda et al. 2014). R. rubiginosa is an exotic and highly invasive thorny shrub and it has been recognized as a key nurse shrub for A. chilensis seedlings in sites under high livestock pressure (Svriz et al. 2013).
In spring (September) 2017 we planted 204 seedlings of A. chilensis. Seedlings were 1 year old and 5–10 cm tall, produced in a local nursery with seeds from Lago Puelo National Park (70 km from the study site). A total of 33 seedlings were assigned to two of the three nurse shrub identity treatments (P and N below canopy levels) at each silvopastoral use intensity (two levels). For the inter-canopy without nurse effect treatment (I) 36 seedlings were established only under the high use intensity treatment as high mortality was suspected; while under low use intensity 33 seedlings were established in the inter-canopy treatment. Seedlings were planted in groups of 3–6 individuals, at least 20 m apart from each other, based on the spatial variability and nurse shrub availability of each stand. Stands were separated at least 100 m from each other, or by a geographical feature (i.e. creek, steep slope, etc.). In both P and N treatments the orientation of the seedling in relation to the plant was the same (southwest), and seedlings were planted close to the main branch of the shrub (10 cm approx.), always beneath the canopy. Seedlings were revisited two weeks after planting to evaluate planting failure.
Seedling survival and growth were evaluated during three consecutive years. Seedling survival was evaluated at the end of the growing season (March) for the first 3 years (2018, 2019 and 2020) and at the end of the winter (September) for the first 2 years (2018–2019). Due to seasonal movements of cattle, middle and lowlands rest from herbivory pressure during summer. By measuring seedling survival after the growing season, we expect to assess survival to drought and desiccation, as summer corresponds to the dry season and cattle is not present. By measuring seedling survival at the end of the winter we expect to assess survival to herbivory pressure. Seedling growth in height (mm) and diameter at the root collar (RCD, mm) was measured at the end of the growing season (March). Measures were taken with a digital caliper (Apogee Instruments, USA). Growth by the end of the growing season was then calculated by subtracting the growth of each year to the value of the previous year.
To characterize micro-site conditions we measured light condition and soil moisture every time growth was measured. Light condition was registered with a 50 cm long ceptometer (Apogee Instruments, USA) containing five light sensors. A mean measure of PAR income of the 5 sensors is provided, allowing the inclusion of the high micro-site variability of the measure. Two readings of PAR income registered by the ceptometer were taken 10 cm apart from each other, at 30 cm from the soil, on top of each seedling, and the mean value of those readings was used. At each silvopastoral use intensity, light condition readings were taken at midday of the same day, on sunny days (i.e. cloudy days where avoid). Together with light condition readings, soil moisture at the seedling micro-site was registered with a Decagon ProCheck S2 sensor that provides instant humidity readings at 10 cm depth. The correlation between micro-site light (log transformed) and soil moisture was evaluated with the Pearson correlation coefficient. As both measures were correlated, we decided to use light conditions for further analyses (Fig. S1).
Statistical analyses
The effect of silvopastoral use intensity, nurse shrub identity (P, N, I), and micro-site light availability (only for summer) on seedling survival was evaluated for each season using generalized linear mixed models, considering a binomial distribution of the variables. All factors were considered as fixed effects, stand (group of planted seedlings) was considered as random effect factor and models with and without interactions were built. A multi-model inference was used considering the Akaike Information Criterion (AIC) and ANOVA analyses using chi-squared test. The best fit model was chosen based on the lower AIC and the significance of the difference between models. Further, when a best fit model showed interactions between categorical factors (i.e. intensity and micro-site), the difference between levels of the factors was analyzed based on a posteriori Tukey test. The effect of silvopastoral use intensity, nurse shrub identity (I, N, P) and micro-site light availability on seedlings growth rate (final RCD and height—initial RCD and height) was evaluated for each growing season using generalized linear models, considering the normal and gamma distribution of the variables, and stand as random factor. Analyses were performed using glmer() function in R software (The R Development Core Team 2017). FA multi-model inference was used considering the Akaike Information Criterion (AIC) and ANOVA analyses using chi-squared test. The best fit model was chosen based on the lower AIC and the significance of the difference between models. Relative importance values of all analyzed variables were calculated with the importance() function in the MuMin package, where the Akaike weights are summed between all models for each explanatory variable (Bartón 2019).
Results
At the micro-site level, light and soil moisture were correlated (first summer r = − 0.22 p = 0.004; second summer r = − 0.25 p = 0.003; third summer r = − 0.29 p = 0.009), thus we decided to keep micro-site light availability for further analysis because it showed higher variability between treatments (Figs. S2, S3). Overall, by the end of the experiment, more seedlings survived under low than high silvopastoral use intensity (Fig. 2). When comparing the nurse effect, the best fit model for survival success varied between seasons (winter and summer) and years since establishment. Silvopastoral use intensity was a significant explanatory variable at every season, followed by nurse shrub identity (I, N, P) and light availability (Table 1, Fig. 2). Seedling survival to the first and second summer, and second winter also showed a significant interaction between intensity and micro-site (Table 1). Specifically, for the first and second summer, seedlings planted in high use intensity inter-canopy micro-site showed significantly less survival than seedlings planted in high use under nurse shrub protection (both P and N) and in low use inter-canopy micro-site (Tables S1, S2, and Fig. 2). Third summer showed an interaction between intensity and light, but as light is a continuous factor we were not able to perform a posterior test. Finally, and after the second winter, seedlings planted in high use inter-canopy micro-site showed lower survival than seedlings planted in high use under nurse shrub protection (both P and N). Moreover, seedlings planted in high use palatable shrub micro-site showed lower survival than seedlings planted all three micro-sites (I, N, P) under low use intensity (Table S3, Fig. 2).

Number of surviving planted seedlings of A. chilensis through a three consecutive summers (2017–2020) experiment. Plantation: represents plantation survival, as it shows the number of surviving seedlings two weeks after plantation. HI: high silvopastoral use intensity, LO: low silvopastoral use intensity, I: seedling planted in the inter-canopy micro-site, N: seedling planted under a non-palatable nurse shrub micro-site, P: seedling planted under palatable nurse shrub micro-site. Significant differences are highlighted with a symbol above bars: *means differences between use intensities, #means differences between use intensities and micro-site
Seedling growth in RCD and height was mainly determined by silvopastoral use intensity during the first 2 years. Best fit model for growth in RCD in the first summer was a null model, with low importance of explanatory variables, while growth in height was higher under high use intensity (Fig. 3, Table 2). After the second summer, both growth in RCD and height were higher under high silvopastoral use intensity (Fig. 3).

Growth in diameter at collar (RCD) and height of planted seedlings of A. chilensis after the first and second summer since seedling plantation
Micro-site light availability showed an importance value of 0.45 and 0.51 for growth in RCD and height respectively, although it was not selected as the best fit model (Table 2). In the third summer, the best fit model for growth in RCD included silvopastoral use intensity, while growth in height was explained by silvopastoral use intensity, nurse shrub identity (I, N, P), micro-site light availability and the interaction among them (Fig. 4, and Table 2).

Growth in diameter at collar (RCD) and height of planted seedlings of A. chilensis after the third summer since seedling plantation. I: seedlings planted at the inter-canopy (open condition), N: seedlings planted under a non-palatable shrub, P: seedlings planted under a palatable shrub
Discussion
The nurse effect has been recognized as a key factor for restoration actions in harsh environments (Gómez-Aparicio 2009). Specifically, several studies have shown its importance for the survival of A. chilensis seedlings in active restoration actions in forest subject to extreme logging, fire and forest decline (Urretavizcaya and Defossé 2019, 2013). The SGH hypothesis predicts that facilitation effect increases under harsh environments, but the nature of the disturbance and the palatability of the nurse shrub may modify this prediction. Here we show how the complex nature of silvopastoral land-use can modify the effectiveness of the nurse effect on the survival and growth of planted seedlings after three growing seasons. Under increasing stress of both disturbances (desiccation risk and herbivory pressure), we found higher survival but lower growth rate of planted seedlings under nurse shrub, while being palatable or non-palatable had no significant effect.
Here we evaluated a complex disturbance of an abiotic stress driven by light availability, which can be seen as a stressful factor for plant growth and survival in both extremes of the gradient, from low photosynthetic radiation up to desiccation due to high radiation. For example, the regeneration of certain shade-intolerant canopy species and depend on the development of forest gaps for establishment, while for other species seedling survival increases as radiation decreases (Veblen 1989; Sack 2004; Rodríguez-García et al. 2011; Bannister et al. 2013; Caselli et al. 2021). To add complexity to this bottom-up mechanism, light requirements and drought tolerance of tree species can be different through the ontogeny (Niinemets 2010). Moreover, this disturbance interacts with a biotic stress due to herbivore pressure, which is low in closed canopy stands and higher in open canopy stands, as well as understory vegetation cover (La Manna et al. 2008; Chillo et al. 2018). Our contrasting results for seedling survival and growth due to this complex disturbance interaction do not allow us to fully accept our first hypothesis. We predicted that A. chilensis seedlings will experience higher survival and growth beneath a nurse shrub in high than in low silvopastoral use. We found that under high use intensity (lower canopy cover), the presence of a nurse shrub is always important for summer seedling survival, probably indicating the importance of the nurse shrub to avoid desiccation (Gómez-Aparicio et al. 2004). The same occurred for winter seedling survival under high use intensity, where the presence of a nurse shrub may decrease trampling risk. But contrary to our prediction, height and diameter growth was higher at the inter-canopy (open condition) than under a nurse shrub, and higher under high than low use intensity, probably due to higher light availability.
When considering A. chilensis seedling growth, we found a positive effect of increasing light availability due to canopy opening as found in other local studies (Gobbi and Schlichter 1998; Losada Palenzuela et al. 2018; Caselli et al. 2021). This increase in light availability is also known to favor understory species richness and abundance, especially of colonizing and fast-growing herbs (Arias Sepúlveda and Chillo 2017). Interestingly, our results showed that this increase in understory vegetation cover does not diminish A. chilensis planted seedlings growth. Similar results were recently found by Caselli et al. (2021), when neighboring herbaceous vegetation was considered. Moreover, in our study site lower canopy cover also implied higher herbivory pressure. Silvopastoral use is a land-use which generates open canopy stands where livestock browsing negatively affects sapling density and architecture (Veblen et al. 1992; Relva and Veblen 1998; Blackhall et al. 2008). But these open canopy stands also present higher light availability, a key limiting factor for plant’s growth in dense canopy forests (Peri et al. 2016). For the natural regeneration of A. chilensis under silvopastoral use, the output of the interaction of these contrasting stress factors is not straightforward, mainly because cattle is moved to other areas during summer, allowing a growing season without high herbivory pressure (Amoroso et al. 2018). This seasonal livestock movement that characterizes silvopastoral use in the study area (Chillo et al. 2021) may be a key issue that prevents failure of active restoration practices under chronic impacts (Ashton and Kelty 2018).
We found that, when considering the nurse effect for active restoration, complex interactions between disturbances with different mechanisms are reflected in different ways for seedling survival than for growth. For example, in Mediterranean climates it has been found that nurse shrubs favors seedling survival and growth of less-drought tolerant species. But in harsh environments, when the disturbance level was higher, competition for resources between nurse shrubs and seedlings led to a positive effect on survival but not on growth (Gaviet et al. 2016). Also, in temperate forest with dry summers, when the interaction of two disturbances was analyzed (drought stress and herbivory), a negative effect of nurse shrub was found when abiotic conditions were increased (i.e. drought), while the benefit for herbivory protection was subject to the type of herbivory and the abiotic condition (Guignabert et al. 2020). Accordingly, our results show how seedlings can experience lower survival and higher growth rates under higher silvopastoral use intensity, and although the nurse effect increases survival, it also negatively affects growth. The positive effect of nursing on A. chilensis regeneration had been recognized to vary based on the stress factor. For example, in the arid extremes of its distribution, the nurse effect is important to diminish water stress and its positive effect is higher than the potential competition of the nurse on saplings (Kitzberger et al. 2000; Nuñez et al. 2009). In more mesic conditions, alternatively, there is a negative effect of the nurse shrub on A. chilensis seedlings due to higher damage by arthropod´s herbivory (Caccia et al. 2009). In this sense we highlight how the complexity of the stress gradient is a key factor for nurse effect in restoration actions, besides the ontogeny of the beneficiary and the characteristics of the benefactor (Smit et al. 2007; Nuñez et al. 2009; Svirz et al. 2013; Anthelme et al. 2014).
Regarding the effect of the palatability of the nurse shrub, we hypothesized that under increasing abiotic stress, the biotic stress factor will be less important, and our results support our hypothesis. Although the presence of the nurse shrub was a key factor under high use intensity, whether the shrub was palatable or non-palatable did not have an effect on seedling survival. When considering the abiotic stress factor (light availability), nurse shrub identity has been recognized as important for seedling survival to summer desiccation in the more arid extreme of the A. chilensis distribution (Kitzberger et al. 2000). Also, when considering herbivory pressure, nurse shrub characteristics may modify the frequency and intensity of cattle browsing (Svriz et al. 2013). We found that the presence of a nurse shrub increases seedling survival during the first two summers after implantation, and negatively affects growth in diameter at the collar and in height during the third year after implantation. But these effects were not significantly different between palatable and non-palatable shrubs. These results may show that, in our study sites, disturbance due to cattle herbivory pressure is not as important as light availability pressure under both silvopastoral use intensities. But also, these results go in agreement with recent findings that when complex disturbance interactions are analyzed, an increase in the abiotic stress factor may determine facilitation outcome due to competition for limiting resources; and the effect of the biotic stress may mainly depend on the type of herbivory and the plant ontogeny (Niinemets 2010; Gavinet et al. 2016; Guignabert et al. 2020).
Finally, our results highlight the importance of considering the ecological mechanism behind the analyzed disturbance when planning active restoration actions, as it determines the effect of disturbance on forest regeneration. For example, in tropical forest cattle can be used for forest restoration as it diminishes seedling competition with grasses. Also, nurse shrubs are recognized to diminish grasses growth due to shading (Posadas et al. 2001; Ibanez and Hart 2020). In this case, a top-down mechanism of herbivory pressure on dominant grasses was determining restoration results. In our case study at temperate forest, with the same disturbance effect (cattle grazing and browsing) a bottom-up mechanism of light availability (favoring seedling growth but also increasing desiccation risk) seems to determine the results of active restoration for silvopastoral use.
Management implications
Our results have important restoration implications, and more importantly, allow for diverse actions to forest managers. Although the elimination of the disturbance would be ideal for restoration, the exclusion of livestock in a typical silvopastoral system of north-west Patagonia is not a viable solution in the short term. Thus, here we evaluated the outcome of active restoration actions such as planting without disturbance elimination, but rather considering alternatives for enchaining restoration success. If planting in the inter-canopy (open condition) would be preferred given the higher (almost double) growth rates, planting density in these areas must be higher to compensate for lower seedling survival. Yet, given the spatial variability in these areas, planting under all micro-site conditions is suggested. Under a scenario of chronic disturbance (i.e. continuity of timber extraction and livestock seasonal movements), efforts must be put to guaranteeing initial seedling survival by planting, primarily, under the nurse effect of shrubs independently of their characteristics (i.e. palatable or non-palatable). Additional actions would be required after 3 years in order to promote high growth rates by performing release cuttings of the seedlings by the manual removal of the nurse shrubs. If this last practice cannot be carried out, managers might consider increasing the number of seedlings and prioritize nurse-shrub conditions rather than the open condition even when future growth will be restricted.
References
Amoroso MM, Larson BC (2010) Can a natural experiment be used as a tool to design partial cutting regimes? The decline of Austrocedrus chilensis forests, an example. J for Res 15:38–45. https://doi.org/10.1007/s10310-009-0139-3
Amoroso MM, Chillo MV, Alcalá V, Rezzano CA, Arpigiani DF, Villacide EM (2018) ¿Cómo modifica el uso silvopastoril la estructura y la dinámica poblacional de los bosques mixtos de ciprés de la cordillera y coihue? Ecosistemas 27:33–40. https://doi.org/10.7818/ECOS.1502
Amoroso MM, Peri PL, Lencinas MV, Soler Esteban R, Rovere A, Gonzalez Peñalba M, Chauchard L, Urretavizcaya MF, Logercio G, Mundo I, Cellini JM, Dezzotti A, Attis Beltran H, Bahamonde H, Ladio A, Gowda J, Gallo L, Quinteros P, Sola G and Martinez Pastur G (2021) Capítulo 11: Región Patagónica (Bosques Andino-Patagónicos). In: Peri PL, Martínez Pastur G, Schlichter T (eds) Uso sostenible del bosque: Aportes desde la Silvicultura Argentina. Ciudad Autónoma de Buenos Aires, 889 pp
Anthelme F, Gómez-Aparicio L, Montúfar R (2014) Nurse-based restoration of degraded tropical forests with tussock grasses: experimental support from the Andean cloud forest. J App Ecol 51:1534–1543. https://doi.org/10.1111/1365-2664.12311
Arias Sepúlveda JE, Chillo V (2017) Cambios en la diversidad funcional del sotobosque y la tasa de descomposición frente a diferentes intensidades de uso silvopastoril en el noroeste de la Patagonia Argentina. Ecol Austral 27:29–38. https://doi.org/10.25260/ea.17.27.1.0.297
Ashton MS, Kelty MJ (2018) The practice of silviculture: applied forest ecology. Wiley, Hoboken, p 776
Bannister JR, Coopman R, Donoso P, Bauhus J (2013) The importance of microtopography and nurse canopy for successful restoration plant-ing of the slow-growing conifer Pilgerodendron uviferum. Forests 4:85–103
Barbosa JM, Asner GP (2017) Prioritizing landscapes for restoration based on spatial patterns of ecosystem controls and plant–plant interactions. J App Ecol 54:1459–1468. https://doi.org/10.1111/1365-2664.12857
Bartón K (2019) MuMIn: multi-model inference. R Packag. version 1.43.6. https://CRAN.R-project.org/package=MuMIn. Accessed 22 Mar 2021
Beauchesne D, Cazelles K, Archambault P, Dee LE and Gravel D (2021) On the sensitivity of food webs to multiple stressors. Ecol Letters 24: 2219–2237
Bertness MD, Callaway R (1994) Positive interactions in communities. Trends Ecol Evol 9:191–193. https://doi.org/10.1016/0169-5347(94)90088-4
Blackhall M, Raffaele E, Veblen TT (2008) Cattle affect early post-fire regeneration in a Nothofagus dombeyi-Austrocedrus chilensis mixed forest in northern Patagonia, Argentina. Biol Cons 141:2251–2261. https://doi.org/10.1016/j.biocon.2008.06.016
Blackhall M, Veblen TT, Raffaele E (2015) Recent fire and cattle herbivory enhance plant-level fuel flammability in shrublands. J Veg Sci 26:123–133. https://doi.org/10.1111/jvs.12216
Brooker RW, Maestre FT, Callaway RM, Lortie CL, Cavieres LA, Kunstler G, Liancourt P, Tielbörger K, Travis JMJ, Anthelme F, Armas C, Coll L, Corcket E, Delzon S, Forey E, Kikvidze Z, Olofsson J, Pugnaire F, Quiroz CL, Saccone P, Schiffers K, Seifan M, Touzard B, Michalet R (2008) Facilitation in plant communities: the past, the present, and the future. J Ecol 96:18–34. https://doi.org/10.1111/j.1365-2745.2007.01295.x
Caccia FD, Chaneton EJ, Kitzberger T (2009) Direct and indirect effects of understorey bamboo shape tree regeneration niches in a mixed temperate forest. Oecologia 161:771–780. https://doi.org/10.1007/s00442-009-1412-z
Cardozo AG (2014) Estrategias socio-productivas de establecimientos ganaderos del sudoeste de la provincia de Río Negro, Argentina. Tesis de Maestría, Universdiad de Buenos Aires, Argentina. 183 pp, Available from http://ri.agro.uba.ar/files/download/tesis/maestria/2014cardozoandreagabriela.pdf. Accessed 22 Mar 2021
Caselli M, Loguercio GÁ, Urretavizcaya MF, Defossé GE (2020) Developing silvicultural tools for managing mixed forest structures in Patagonia. For Sci 66:119–129. https://doi.org/10.1093/forsci/fxz052
Caselli M, Urretavizcaya MF, Logercio GA, Contardi L, Gianolini S, Defossé GE (2021) Effects of canopy cover and neighboring vegetation on the early development of planted Austrocedrus chilensis and Nothofagus dombeyi in north Patagonian degraded forests. For Ecol Manage 479:118543. https://doi.org/10.1016/j.foreco.2020.118543
Chillo V, Vázquez DP, Amoroso MM, Bennett EM (2018) Land-use intensity indirectly affects ecosystem services mainly through plant functional identity in a temperate forest. Funct Ecol 32:1390–1399. https://doi.org/10.1111/1365-2435.13064
Chillo V, Ladio AH, Salinas Sanhueza J, Arpigiani DF, Soler R, Cardozo A, Peri P, Amoroso MM (2021) Silvopastoral systems in Northern Argentine Chilean andean Patagonia: ecosystem services provision in a complex territory. In: MartínezPastur G et al (eds) Ecosystem services in Patagonia: a multi-criteria approach for an integrated assessment. Springer, Cham, pp 115–138
Dezzoti A (1996) Austrocedrus chilensis and Nothofagus dombeyi stand development during secondary succession, in Northwestern Patagonia, Argentina. For Ecol Manag 89:125–137. https://doi.org/10.1016/S0378-1127(96)03860-1
Gavinet J, Prévosto B, Fernández C (2016) Do shrubs facilitate oak seedling establishment in Mediterranean pine forest understory? For Ecol Manage 318:289–296. https://doi.org/10.1016/j.foreco.2016.09.045
Gobbi M, Schlichter T (1998) Survival of Austrocedrus chilensis seedlings in relation to microsite conditions and forest thinning. For Ecol Manage 111(1998):137–146
Gómez-Aparicio L (2009) The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and ecosystems. J Ecol 97:1202–1214. https://doi.org/10.1111/j.1365-2745.2009.01573.x
Gómez-Aparicio L, Zamora R, Gómez JM, Hódar JA, Castro J, Baraza E (2004) Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecol Appl 14:1128–1138
Gordon AM, Newman SM, Coleman BRW, Thevathasan NV (2018) Temperate agroforestry: an overview. In: Gordon AM, Newman SM, Coleman BRW (eds) Temperate agroforestry systems. CAB International, Wallingford, pp 1–6
Gowda JJH, Garibaldi A, Pirk G, Blackhall M, Chaneton EJ, de Paz M, Díaz S, Galende G, Mazía N, Paritsis J, Raffaele E, Relva MA, Sasal Y (2014) Herbívoros: actores clave. In: Raffaele E et al (eds) Ecología e historia natural de la Patagonia Andina, un cuarto de siglo de investigación en biogeografía, ecología y conservación. Fundación de Historia Natural Félix de Azara, Ciudad Autónoma de Buenos Aires, pp 91–112
Guignabert A, Augusuto L, Gonzalez M, Chioeaux C, Delerue F (2020) Complex biotic interactions mediated by shrubs: revisiting the stress-gradient hypothesis and consequences for tree seedling survival. J Appl Ecol 57:1341–1350. https://doi.org/10.1111/1365-2664.13641
Ibanez T, Hart PJ (2020) Spatial patterns of tree recruitment in a montane Hawaiian wet forest after cattle removal and pig population control. Appl Veg Sci 23:197–209. https://doi.org/10.1111/avsc.12478
Jones HP (2013) Impact of ecological restoration on ecosystem services. In: Levin S (ed) Encyclopedia of biodiversity, 2nd edn. Springer, Cham, pp 199–208
Kitzberger T, Steinaker DF, Veblen TT (2000) Effects of climatic variability on facilitation of tree establishment in Northern Patagonia. Ecology 81:1914–1924. https://doi.org/10.1890/0012-9658(2000)081[1914:EOCVOF]2.0.CO;2
La Manna L, Collantes C, Bava J, Rajchenberg M (2008) Seedling recruitment of Austrocedrus chilensis in relation to cattle use, microsite environment and forest disease. Ecologia Austral 18:27–41
Letourneau FJ, Andenmatten E, Schlichter T (2004) Effect of climatic conditions and tree size on Austrocedrus chilensis–shrub interactions in Northern Patagonia. For Ecol Manag 191:29–38. https://doi.org/10.1016/j.foreco.2003.11.002
Losada Palenzuela S, Amoroso MM, Bogino SM (2018) Regeneration dynamics of Austrocedrus chilensis and Nothofagus dombeyi in declining forests. Bosque 39:333–345
Madariaga MC (2019) El valle del río Manso inferior y su funcionamiento como sistema. Comunicación técnica Inta EEA S.C: Bariloche, Área Desarrollo Rural, N°250. Ediciones INTA. http://hdl.handle.net/20.500.12123/6359. Accessed 22 Mar 2021
Maestre FT, Callaway RM, Valladares F, Lortie CJ (2009) Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J Ecol 97:199–205. https://doi.org/10.1111/j.1365-2745.2008.01476.x
Navarro-Cano JA, Goberna M, Verdú M (2019) Using plant functional distances to select species for restoration of mining sites. J App Ecol 56:2353–2362. https://doi.org/10.1111/1365-2664.13453
Niinemets U (2010) Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: past stress history stress interactions, tolerance and acclimation. For Ecol Manag 260:1623–1639. https://doi.org/10.1016/j.foreco.2010.07.054
Nuñez CI, Raffaele E, Nuñez MA, Cuassolo F (2009) When do nurse plants stop nursing? Temporal changes in water stress levels in Austrocedrus chilensis growing within and outside shrubs. J Veg Sci 20:1064–1071. https://doi.org/10.1111/j.1654-1103.2009.01107.x
Oudkerk L, Pastorino M, Gallo L (2003) Siete años de experiencia en la restauración postincendio de un bosque de Ciprés de la Cordillera. Patagonia Forestal 9:4–7
Peláez M, Dirzo R, Fernandes GW, Perea R (2019) Nurse plant size and biotic stress determine quantity and quality of plant facilitation in oak savannas. For Ecol Manag 437:435–442. https://doi.org/10.1016/j.foreco.2019.02.010
Peri PL, Dube F, Varella AC (2016) Silvopastoral systems in the subtropical and temperate zones of South America: an overview. In: Peri PL, Dube F, Varella A (eds) Silvopastoral systems in Southern South America. Springer, Amsterdam, pp 1–8
Posadas JM, Aide MT, Cavelier J (2001) Cattle and weedy shrubs as restoration tools of tropical montane rainforest. Restor Ecol 8:370–379. https://doi.org/10.1046/j.1526-100x.2000.80052.x
Relva MA, Veblen TT (1998) Impacts of introduced large herbivores on Austrocedrus chilensis forests in Northern Patagonia, Argentina. For Ecol Manag 108:27–40. https://doi.org/10.1016/S0378-1127(97)00313-
Rey Benayas JM, Newton AC, Díaz A, Bullock JM (2009) Enhancement of biodiversity and ecological services by ecological restoration: a meta-analysis. Science 352:1121. https://doi.org/10.1126/science.1172460
Rodríguez-García E, Bravo F, Spies TA (2011) Effects of overstorey canopy, plant–plant interactions and soil properties on Mediterranean maritime pine seedling dynamics. For Ecol Manage 262:244–251. https://doi.org/10.1016/j.foreco.2011.03.029
Rovere AE (2008) Ensayo de restauración con Austrocedrus chilensis (Cupressaceae) en Patagonia, Argentina. In: González Espinosa M, Rey Benayas JM, Ramírez Marcial N (eds) Restauración de bosques en América Latina. Mundi-Prensa, México City, pp 6–21
Sack L (2004) Responses of temperate woody seedlings to shade and drought: do trade-offs limit potential niche differentiation? Oikos 107:110–127
Smit C, Vandenberghe C, Den Ouden J, Müller-Schärer H (2007) Nurse plants, tree saplings and grazing pressure: changes in facilitation along a biotic environmental gradient. Oecologia 152:265–273. https://doi.org/10.1007/s00442-006-0650-6
Smit C, Rietkerk M, Wassen MJ (2009) Inclusion of biotic stress (consumer pressure) alters predictions from the stress gradient hypothesis. J Ecol 97:1215–1219. https://doi.org/10.1111/j.1365-2745.2009.01555.x
Soliveres S, Maestre FT (2014) Plant-plant interactions, environmental gradients and plant diversity: a global synthesis of community-level studies. Persp Plant Ecol Evol Syst 16:154–163. https://doi.org/10.1016/j.ppees.2014.04.001
Soliveres S, Eldridge DJ, Maestre FT, Bowker MA, Tighe M, Escudero A (2011) Microhabitat amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: towards a unifying framework. Persp Plant Ecol Evol Syst 13:247–258. https://doi.org/10.1016/j.ppees.2011.06.001
Svriz M, Damascos MA, Zimmermann H, Hensen I (2013) The exotic shrub Rosa rubiginosa as a nurse plant. Implications for the restoration of disturbed temperate forests in Patagonia, Argentina. For Ecol Manag 289:234–242. https://doi.org/10.1016/j.foreco.2012.09.037
The R Development Core Team (2017) R: a language and environment for statistical computing. ISBN 3-900051-07-0
Urretavizcaya MF, Defossé GE (2013) Effects of nurse shrubs and tree shelters on the survival and growth of two Austrocedrus chilensis seedling types in a forest restoration trial in semiarid Patagonia, Argentina. Ann for Sci 70:21–30. https://doi.org/10.1007/s13595-012-0234-z
Urretavizcaya MF, Defossé GE (2019) Restoration of burned and post-fire logged Austrocedrus chilensis stands in Patagonia: effects of competition and environmental conditions on seedling survival and growth. Int J Wildlife Fire 28:365–376. https://doi.org/10.1071/WF18154
Urretavizcaya MF, Pastorino M, Mondino V, Contardi L (2015) La plantación con árboles nativos. In: Chauchard L, Frugoni MC, Nowak C (eds) Manual de buenas prácticas para el manejo de plantaciones forestales en el Noroeste de la Patagonia. Ministerio de Agroindustria, Buenos Aires, pp 335–368
Urretavizcaya MF, Gonda HE, Defossé GE (2017) Effects of post-fire plant cover in the performance of two cordilleran cypress (Austrocedrus chilensis) seedling stocktypes planted in burned forests of Northeastern Patagonia, Argentina. Environ Manag 59:419–430. https://doi.org/10.1007/s00267-016-0793-0
Urretavizcaya MF, Caselli M, Contardi L, Loguercio GA, and Defossé GE (2018) Enriquecimiento de bosques degradados de cirpés de la coordillera con especies nativas de alto valor forestal. Informe Final Proyecto PIA 14067. UCAR-CIEFAP, Argentina
Vandenberghe C, Smit C, Pohl M, Buttler A, Freléchoux F (2009) Does the strength of facilitation by nurse shrubs depend on grazing resistance of tree saplings? Basic Appl Ecol 10:427–436. https://doi.org/10.1016/j.baae.2008.08.009
Veblen TT (1989) Nothofagus regeneration in treefall gaps in Northern Patagonia. Can J for Res 19:365–371
Veblen TT, Mermoz M, Martin C, Kitzberger T (1992) Ecological impacts of introduced animals in Nahuel Huapi National Park. Argentina Cons Biol 6:71–83. https://doi.org/10.1046/j.1523-1739.1992.610071.x
Veblen TT, Burns BR, Kitzberger T, Lara A, Villalba R (1995) The ecology of the conifers of southern South America. In: Enright NJ, Hill RS (eds) Ecology of the Southern Conifers. Melbourne University Press, Carlton, pp 120–155
Acknowledgements
Mariana Campo, Brisa Serena Güenuleo, M. Noel Szudruk, Liliana Barbosa, Facundo Gómez and Manuel de Paz helped with field work. We are grateful to Lisandro and Oscar Lanfre for allowing us to install the experiment in their ranch; and to Ubuntu’s and R’s developers and contributors for making science available to everyone. This work was supported by CONICET, Agencia Nacional de Promoción Científica y Tecnológica [Grant No. PICT 2015-1692] and Universidad Nacional de Río Negro [Grant No. PI 40-B-478].
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
11056_2022_9914_MOESM2_ESM.pdf
Supplementary file2 (PDF 103 kb) Fig. S1 Correlation between light income readings (log transformed) and soil moisture readings at each micro-site, during three consecutive summers.
11056_2022_9914_MOESM3_ESM.pdf
Supplementary file3 (PDF 6 kb) Fig. S2 Low silvopastoral use intensity. Light and soil moisture conditions at each micro-site treatment (inter-canopy, non-palatable and palatable shrub) during three consecutive years.
11056_2022_9914_MOESM4_ESM.pdf
Supplementary file4 (PDF 6 kb) Fig. S3 High silvopastoral use intensity. Light and soil moisture conditions at each micro-site treatment (inter-canopy, non-palatable and palatable shrub) during three consecutive years.
Rights and permissions
About this article

Cite this article
Chillo, V., Amoroso, M., Arpigiani, D. et al. Forest active restoration for silvopastoral use in Northwestern Patagonia: relative importance of the nurse effect. New Forests 54, 201–215 (2023). https://doi.org/10.1007/s11056-022-09914-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-022-09914-0