
Abstract
Nutrient return to soil by applying forest industry residues help ameliorate soil nutrient exhaustion and promote forest management sustainability. This study evaluated the effects of forest industry residue application on soil attributes, on forest floor amounts, and on growth and nutrition of Pinus taeda L. (loblolly pine). Six rates (0, 14, 25, 49, and 60 T ha−1) of residue (boiler ash and cellulose sludge mixture) were applied to P. taeda stands on a low fertility soil. Seven years after application, trees were harvested for wood volume, wood biomass, canopy mass, and nutrient composition. Wood density was determined along the trunk length. Soil samples from 0–10 to 10–20 cm depths were collected for evaluating chemical attributes. Although residue application only resulted in small increases in soil Ca and P availability, high yield enhancements were observed with no compromise to wood density. Maximum growth response was obtained with the 49 T ha−1 residue application. This rate enhanced trunk volume from 117 to 250 m3 ha−1, total biomass from 76 to 127 T ha−1, and total C from 34.5 to 57.2 T ha−1. Residue application decreased Mn content in all biomass compartments, and values above 193.5 (needles) and 26.2 mg kg−1 (bark) defined low annual growth increases. Residue application also decreased Al, Fe, S, B, Ca, and P concentrations in branches or needles. Forest floor was enhanced by residue amendment, which increased C sequestration by 7 T/ha. Forest sustainability practices can be enhanced by applications of industrial residues while improving tree yield and nutrition.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pinus taeda L. (loblolly pine) is a major forest species under subtropical conditions worldwide, and the Brazilian forest industry has relied on this fast growing species (IBÁ 2019). Although P. taeda is well known for its high adaptability to very acidic low fertility soils (Alves et al. 2013; Motta et al. 2014), nutrient amendment to these soils can result in varied growth responses, especially in the first forest production cycle. Bellote et al. (2005) suggested that management interventions at forest sites can directly affect wood quality, plantation growth rates, and wood biomass accumulation in rotations. Others have shown that nutrient amendment with different sources can result in large productivity improvement (Rodrigues 2004; Rodriguez et al. 2018), suggesting that there are conditions where lack of nutrients limit productivity (Sass et al. 2020).
Soil exhaustion has become a great concern in the forest community due to many harvest cycles without nutrient replacement (Richter et al. 1994; Motta et al. 2014; Gatiboni et al. 2020). Significant portions of nutrients remove by harvests become industrial residue wastes that are disposed in landfills and subsequently removed from natural nutrient cycling (Matysik et al. 2001; Jacobson 2003). These residue sources contain many nutrients, and application of these residues often results in larger and more frequent pine growth response compared to mineral supplement applications (Motta et al. 2014). Rodriguez et al. (2018) reported a 127% increase in pine commercial volume with the application of 84 T ha−1 of pulp-mill sludge in southern Brazil. However, establishing rate recommendations can be difficult since these residues can vary widely in their nutrient composition and how they influence soil acidity (Guedes and Poggiani 2003; Rocha et al. 2004; Abreu et al. 2017). Changes in Pinus growth due to industrial residue application can also be associated with changes in one or more nutrients or toxic elements in tree tissues (Matysik et al. 2001; Jacobson 2003; Rodrigues 2004; Paim 2007; Sikstrom et al. 2010). However, Varnagiryte-Kabašinskiene et al. (2015) observed no changes in canopy tissue nutrient content despite growth response to ash application. Another problem is that residue can not be incorporated into soil after trees are planted; thus residue use is relegated to soil surface applications (Motta et al. 2014). In such situations, Batista et al. (2015) and Marschner and Wilczynski (1991) reported that significant portions of Ca and Mg applied as lime were found suspended in the forest floor litter and did not reach the mineral soil surface. Batista et al. (2015) also reported that the soil surface (upper few centimeters) was affected to a larger extent than subsurface soil.
Addition of nutrients to forest systems may change the quantity and quality of litterfall input and decomposition/humification rate which are major factors controlling the amount of forest floor necromass (Krishna and Mohan 2017). If nutrient amendment increases tree growth and subsequent litterfall input, it is plausible that there will be an increase in forest floor necromass (Consalter et al. 2020). In contrast, nutrient addition (or soil acidity correction) could improve litterfall composition which may enhance litter decomposition and/or promote microorganism activity thus leading to reduced forest floor necromass (Lundström et al. 2003). Such scenarios that affect forest floor necromass magnify difficulties in predicting the influence of surface applications of residues. Eleven years after cellulose residue application, Rabel et al. (2020) reported increases in forest floor necromass while others have reported decreases (Marschner and Wilczynski 1991; Lundström et al. 2003).
The goal of this study was to enhance tree growth on poor sites by applications of industrial forest residue. We hypothesized that high residue application rates would enhance properties of very low fertility soils resulting in better tree nutrition and growth without compromising wood quality and soil protection.
Materials and methods
Experimental area
The experimental study site was located in Piraí do Sul, Paraná state, southern Brazil; this area was 1102 m above sea level with geographical coordinates of 24\(^\circ\) 24′ 27.36′′ S and 49\(^\circ\) 58′ 34.70′′ W. According to Köppen classification, the climate was Cfb temperate oceanic (Alvares et al. 2013; Paraná 2018a, b). Average annual precipitation during the experimental period (2011–2018) varied from 1532 to 1983 mm yr−1 with an average of 90 mm for the driest months (July–August) and 190 mm for the wettest period (January) (Águas Paraná 2018a, b) (Table S1). The soil was classified as Neossolo Quartzarênico Órtico típico (Embrapa, 2013), equivalent to Arenosols (WRB, 2006), developed from sandstone parent material with undulated relief (Sass 2016). Prior to study initiation, two 12 year production cycles of P. taeda were cultivated using shallow cut management without soil amendments and fertilizers.
Before beginning the experiment, soil sampling (0–20 cm depth) was performed for chemical and granulometric characterization following methodology described by Embrapa (2009). The following results were obtained: 3.67 g dm−3 of organic carbon; 4.07 of pH (CaCl2—0.01 M); 0.2 and 0.01 mg dm−3 of P and Na, respectively (Mehlich-I extraction); 3.97 cmolc dm−3 of (H + Al); 0.04 cmolc dm−3 of Ca2+; 0.02 cmolc dm−3 of Mg2+; 0.03 cmolc dm−3 of K+; 0.69 cmolc dm−3 of Al+ 3; and base saturation of 2.5%. Granulometry showed 138 g kg−1 of clay, 75 g kg−1 of silt, and 787 g kg−1 of sand.
Treatments and experimental design
Planting of P. taeda seedlings occurred in January 2011 on a spacing of 2.5 m × 2.5 m. Six months after planting, industrial residue was manually applied to the soil surface (without incorporation) at the following rates: T1—control (0 T ha−1); T2—14 T ha−1; T3—25 T ha−1; T4—49 T ha−1; and T5—60 T ha−1. The experimental design was a randomized block with five replications for at total of 25 plots (plots were 156 m2 in size). Each plot consisted of five rows with five trees per row. The nine central trees were considered the useful experimental area.
Industrial residue characterization
The applied residue was a combination of cellulose sludge (70%) and boiler ash (30%), which had undergone a 12-month open-air composting process. Chemical characteristics of this industrial residue were previously described by Sass (2016) (see Table S2).
Soil sampling and chemical analysis
In July 2018, soil was collected from all plots at depths of 0–10 cm and 10–20 cm. The following parameters were assessed: pH in 0.01 M CaCl2 (1:2.5 soil: solution), Al3+, (H + Al), Ca2+, Mg2+, K+, P, Zn, Mn, Fe, Cu, and C (Silva et al.,1999). Extraction of Al3+, Ca2+, and Mg2+ was performed using 1 M KCl and Mehlich-I (0.025 M HCl and 0.0125 M H2SO4) was used to extract K+, P and micronutrients (Zn, Mn, Fe, and Cu). Carbon was determined after digestion with sodium dichromate (Na2Cr2O7 + H2SO4). Elements were quantified using the following techniques: Ca, Mg, Fe, Mn, Zn, and Cu—Atomic absorption spectrophotometry; C and P—Spectrophotometry/Photocolorimetry; K—Flame emission spectrometry; Al—Titration with sodium hydroxide using bromothymol blue indicator.
Dendrometric evaluation
Seven years after planting, dendrometric evaluations were performed by examining the nine central trees in each plot. Total heights were measured with a Haglöf digital clinometer, and diameters at breast height (DBH) were evaluated using a dendrometric tape. After data collection, basal area was calculated on a 1600 trees per ha basis.
Biomass assessment
From each experimental plot, the tree with DBH closest to the mean was selected for biomass assessment. Length and diameter of the crown was measured and separated into three parts (lower, middle, and upper third of crown). Branches were separated into live (with green needles) and dead (dry without needles). These fractions were weighed in the field, and needles from branches were separated and weighed to determine green mass. In the laboratory, samples were dried, weighted for dry mass, and processed for nutritional characterization.
Total trunk height and commercial height (total trunk minus portions less than 8 cm in diameter) were measured. Volume (with and without bark) was calculated using the methods of Hohendadl (1936). Diameters were measured at the trunk base and at points located at 10%, 30%, 50%, 70% and 90% of commercial height; using these measurements, tree volume (m3) was determined using the shape function with an adjustment value of 0.33.
Samples (5 cm thick discs) were taken at the same cut points as above, and the NBR 11,941:2003 immersion method was used for basic wood density evaluation (ABNT, 2003). Density of each segment was then extrapolated to the calculated volume (m3) of each segment.
Samples for nutrient determination in wood and bark were collected from 0% (base), 50% (middle), and 100% (commercial tip) of the trunk length. These results were used to calculate average nutrient content of trees.
Forest floor assessment
Forest floor litter was sampled at six points in each plot using a frame (0.25 × 0.25 m). The frame was placed on the forest floor and needles along with other residue with no decomposition signs (i.e., new forest floor = new FF) were cut, manually removed, and placed into bags. The remaining materials (collectively referred to as old FF) were characterized as fragmented and partially fragmented litter (fragmented litter—F) and totally decomposed portions (humified layer—H); these samples were placed into another bag. Each old FF litter sample was placed in a container of water, manually agitated to remove soil, and floating material was manually removed into bags. Some of the suspension was gently decanted into a fine sieve to remove any remaining soil; this process was repeated several times until all organic material was separated. All sample material was then placed into bags, dried, and weighed as done with the fresh litter. Both new FF and old FF samples were oven dried (65o C), weighed, and ground in a Wiley mill.
Total C was determined using a Perkin-Elmer CHNS/OPEZ 400 series Elemental Analyzer. In addition, color of new FF and old FF was determined according to the Munsell color system.
Tree nutrient assessment
Needle, live branch, dead branch, bark, and wood samples were dried at 60º C, ground in a Wiley mill, and subjected to dry digestion (Martins and Reissmann 2007) before determinations of P, K, Ca, Mg, S, Fe, Mn, B and Al using an Optical Emission Spectrometer with Inductively Coupled Plasma (ICP–OES Varian, 720–ES). Carbon and N analyses were done using a Perkin-Elmer CHNS/OPEZ 400 series Elemental Analyzer.
Masses (dead branches, live branches, needles, bark and wood) and corresponding average nutrient concentrations were used to calculate nutrient content in each tree part. Values were extrapolated to a hectare basis based on a population of 1,600 trees per ha.
Statistical analysis
Soil characteristics, litter, nutrient concentration, and nutrient content data were subjected to the Bartlett test followed by analysis of variance (ANOVA), and Tukey's test using a 95% confidence level (p < 0.05). Regression analyses were done using DBH and height or volume, and when significant, the Maximum Technical Efficiency rate (MTE) was calculated using the first derivative of the second-degree equation (X = -b1 / 2.b2). The annual average increment (AAI) was calculated by dividing final tree volume by the age of the P. taeda plantation. Regression trees were utilized to predict the annual average increment (AAI). Statistical analyses were performed using R software (version 3.5.3) and regression tree was performed by using the function ctree of “party” package (Hothorn et al., 2006).
Results
Soil proprieties
No major changes in soil chemical proprieties were observed seven years after residue application (Table 1). Soil pH varied by depth, where the 0–10 cm layer was slightly higher (average value of 4.08) compared to the 10–20 cm layer (average value of 3.90). The same was observed for exchangeable Al. At the 10–20 cm depth, the H + Al and Ca concentration had the highest value at the 49 T ha−1 residue rate. This was also true for P concentration at the same soil depth.
Phosphorus increased from 2.64 mg kg−1 (control) to 14.24 mg dm−3 (49 T ha−1) at the 10–20 cm soil depth, and a small increase from 1.72 mg dm−3 (control) to 4.42 mg dm−3 (49 T ha−1) was seen in the upper layer (0–10 cm). Residue application did not change soil C concentration, but average values at the 0–10 cm layer (18.9 g dm−3) were lower compared to those observed in the 10–20 cm layer (24.8 g dm−3) (Table 1).
Tree growth
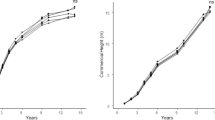
Diameter at breast height was positively influenced by residue treatments (p < 0.01) with no difference among rates (Fig. 1a). The highest value was observed at the 49 T ha−1 rate (attaining a diameter of 18.44 cm), which represented a gain of ~ 4 cm relative to the control (Fig. 1a). However, average commercial tree height was not affected by residue application (p = 0.22; Fig. 1b) despite some increases in total tree height at higher rates (Fig. 1b). Similarly, trunk volume was influenced by residue without difference among rates. Volume without bark (p < 0.01) increased from 96 to 211 m3 ha−1 and volume with bark (p < 0.01) increased from 117 to 250 m3 ha−1 (Fig. 1c), with the highest value observed at 49 T ha−1. Thus, the Annual Average Increment (AAI) for volume without bark was 13.7 m3 ha−1 year−1 in the control treatment and 30.2 m3 ha−1 year−1 for the ̴49 T Mg ha−1 rate (i.e., AAI increased more than two times).

Diameter at breast height—DBH (a), total and commercial height (b) commercial volume with bark and commercial volume without bark (c) of 7 year-old Pinus taeda at different rates of industrial residue (0, 14, 25, 49, and 60 T ha−1)
Tree crown length was influenced by residue use (p = 0.01) and reached an average of 10.4 m at the two highest rates, compared to 8.9 m for the control (Fig. 2a). For crown width, residue application did not result in differences; averages were 2.00 m for the lower (p = 0.49) and middle (p = 0.69) thirds, and 1.50 m for the upper third (p = 0.80) (Fig. 2b).

Crown length (a) and crown width (b) of 7 year-old Pinus taeda at different rates of industrial residue (0, 14, 25, 49, and 60 T ha−1). Column means with the same letters do not differ statistically at the 5% probability level (Tukey's test). ns = not significant
Total tree biomass increased from 75.71 (control) to 127.39 T ha−1 (49 T ha−1 rate), and wood biomass (trunk without bark) increased from 48.91 (control) to 84.12 and 77.30 T ha−1 (Table 2), for 49 and 60 T ha, respectively. There was no difference among tree biomass components (wood, bark, needles and live and dead branches) as a result of industrial residue amendment (Table 2). Regardless of treatment, the decreasing sequence for total tree biomass components was: wood (64.60–68.99%) > bark (10.28–14.40%) > live branches (8.85–11.65%) > needles (4.55–7.98) > dead branches (3.20–7.85%) (Table 2).
Wood quality
Wood density varied with trunk height (p < 0.01), decreasing from the base to the top of the tree (Table 3). This also decreased for the highest treatment (p = 0.02), but there was no interaction with height vs residue application (p = 0.83). However, weighted density (due to differences in volume of trunk segments) did not show significant variation with residue application, and density values were close to ideal for the pulp and paper industry (0.405 g cm−3).
Tree tissue elemental composition
There was no effect of residue application on concentrations of C, N, K, Mg and S in tree tissues (Table 4). Phosphorus, Ca, and S concentrations showed the highest values in needles of the control; this was also true for P, Mg, and S in live branches. Concentrations of Fe, Mn, B, and Al decreased with increasing residue rate (Table 4). For bark and wood, only Mn concentration was decreased by residue application (Table 5). Manganese was the only element that displayed decreases in all five evaluated tissues types with residue application (Tables 4 and 5). Regression trees confirm the importance of Mn nutrition in tree growth; among all elements analyzed, only Mn was a predictor of AAI. Manganese concentrations below 193.5 mg kg−1 in needles were associated with the highest AAI (Fig. 3a); similarly, bark concentrations lower than 26.17 mg kg−1 were also associated with the highest AAI (Fig. 3b).

Regression tree of annual average increment (AAI) explained by elemental composition of needles (a) and base bark (b)
Forest biomass nutrient content
The structural elements C and N increased in bark and wood fractions as productivity increased in response to residue application (Table S4). Potassium, Ca, Mg, and S content increased only in the wood fraction (Table S4). In contrast, micronutrient and Al decreases were observed only in crown compartments (i.e., needles and live and dead branches). In these three fractions, Mn and B content decreased with residue application, Al decreased in dead branches and needles, and Fe decreased in dead branches (Table S3).
Macronutrients, micronutrients, and Al content in aboveground tree biomass followed the sequence C > N > K > Ca > P > Mg > S for macronutrients, and Al > Mn > Fe > B for micronutrients and Al (Table S4). Macronutrients and Al amounts mirrored biomass production, and attained highest values with the rate (49 T ha−1) that produced maximum growth. There was a decrease in total Mn and B in aboveground parts, suggesting that large decreases in tissue concentration accompanied increased growth.
Forest floor
The residue treatment that resulted in maximum growth increased the amount of necromass in both old FF and total litter (Table 6). The relative increase was larger for old FF (23.8 to 46.0 T ha−1) than for new FF (5.1 to 6.3 T ha−1). In general, increased necromass was not associated with amendment rate.
Regardless of treatment, all new FF was Reddish brown after grinding, indicating no influence of residue application. However, ground old litter was observed to have changed color with residue application. The control had color between Dark reddish gray and Dark gray, while treatments receiving residue varied between Dark gray and Black (Table S5); color of old FF also varied among individual samples (Table S5). The authors believe that small charcoal fragments found in applied residue may have influenced color changes.
Concentrations of C in new FF (437–460 g kg−1) were higher than those observed in old FF (318 to 350 g kg−1). There was no effect of residue amendment on C concentration for new FF and old FF. Consequently, total carbon content in litter increased with residue application, with an increase of ~ 60% at 49 T ha−1 compared to the control.
Discussion
Soil proprieties
Despite amount of residue applied, enhancement was only observed for Ca and P at 10–20 cm depth. Although Ca availability was more than doubled, values were still considered very low (< 0.5 cmolc dm−3) or low (0.5–1.0 cmolc dm−3) (Pauletti and Motta 2018), while P availability increased from very low (< 2.0 mg dm−3) to low (2.0–3.0 mg dm−3) and medium (4.0–5 mg dm−3). Thus, observed improvements are probably attributable to P availability, which has been shown to be the element most limiting P. taeda growth (Consalter et al. 2020; Sikstrom et al. 2010).
In general, when there is no incorporation of residue and lime into soil, effects are concentrated in the first few centimeters of the upper soil profile. Eleven years after broadcasting 30 T ha−1 of lime over the forest floor, large acidity variations found in forest floor litter and in the 5 cm soil depth prompted Meiwes (1995) to state that decades may be needed to change acidity in deeper soil layers. Conversely, Marschner and Wilczynski (1991) applied 6.1 T ha−1 of lime to a 40 year-old forest and found soil changes to a depth of 10 cm after 3 years. These varied findings highlight the necessity for more studies examining the influence of residue on deep soil layers.
Batista et al. (2015) and Rabel et al. (2020) found that thick forest floor litter can be a barrier to industrial residue and lime applications. However, residue application in our study occurred in the absence of significant litter amounts to compromise soil contact and reaction with soil. In addition, the low adsorption capacity and small buffer capacity due to low clay concentration (13%) were excellent soil characteristics for observing changes in soil properties. Thus, the lack of large changes in soil properties demonstrates the low potential of our residue for correcting soil acidity and supplying nutrients.
A previous evaluation of soil fertility in this study area found no change in pH, Ca, and available P for three consecutive years following residue application (Sass et al. 2020). Thus, changes observed in soil 7 years after application indicated a slow release of nutrients from applied residue. Rabel et al. (2020) evaluated the effects of using alkaline residues (from recycled paper) after 1, 5, and 10 years and found that the greatest change occurred 5 years after application, which also suggested slow reaction. Unlike lime sludge (Corrêa et al. 2007), residue used in our study had no acidity correction capacity, and unlike sewage sludge (Ibrahim et al., 2019), our residue was nutrient poor.
Tree growth
In contrast to minimal changes observed in soil, tree growth response was notable; volume increased more than 100% in the 49 T ha−1 treatment. It is important to highlight that residue application only increased Ca and P; concentrations of both nutrients were more than doubled. This beneficial effect was not unexpected since this area suffered from a combination of naturally low soil fertility and nutrient exhaustion from successive Pinus cultivation without fertilization or liming (Gatiboni et al. 2020). These responses to residue confirm the high growth potential and the lack of limiting factors (water availability and rooting soil depth) in this area.
Large yield enhancements have been observed under similar soil conditions and management (Rodrigues 2004; Rodriguez et al., 2018). Rodrigues (2004) also observed gains of 80% in height, DBH, and volume in response to application of industrial residue (20 T ha−1) on low fertility sandy soils. Likewise, working with P. taeda on a sandy soil, Rodriguez et al. (2018) found that higher rates of composted cellulose sludge (84 T ha−1) increased tree diameter, height, and volume by 24, 37, and 127%, respectively. Sikstrom et al. (2010) found a sixfold volume improvement in Pinus sylvestris for a drained peat area amended with ash. Others reported increases close to 15% with residue application on clay-loam (Rabel et al. 2020) and clayey soils (Paim 2007). Results of Rodriguez et al. (2018) confirm the potential of applying cellulose sludge as a fertilizer to improve wood productivity of forest plantations established on low fertility soils. In general, growth improvements have been observed in P. taeda plantations under conditions without climatic and depth limitations in nutrient poor soils receiving fertilizer (Samuelson et al. 2004; Albaugh et al. 2020).
Biomass partitioning
Unlike our study where biomass abundance followed the sequence wood > branches (live and dead) > bark > needles, Corrêa and Bellote (2011) found wood > bark > branches > needles for sites with 10.5 to 12 year-old Pinus caribaea var. hondurensis. Albaugh et al. (2004) also reported differences for P. taeda where 8 year-old trees followed the sequence stem > branch > needles > bark and 16 year-old trees had a sequence of wood > branch > bark > needles. They also reported a sharp decrease for needle (20–5%) and increase for stem (49–58%) contributions to total aboveground biomass in these respective age classes. Thus, our 7 year-old P. taeda system displayed similarities to 16 year-old trees evaluated by Albaugh et al. (2004).
We noted no differences in crown tree partitioning as a result of residue amendment. This is contrary to the general statement that fertilization or better fertility conditions increases stem partitioning to tree crown biomass. Corrêa and Bellote (2011) found 57, 22, 12, and 8% for poor growth sites compared to 67, 18, 8, and 8% for good growth sites for stem, bark, branches, and needles of Pinus caribaea var. hondurensis, respectively. Albaugh et al. (2004) reported changes in stem partitioning from 58 to 63% in tree crown as result of fertilization in a 16 year-old P. taeda forest. However, Samuelson et al. (2004) found no differences in tree crowns (foliage, live branches and stem) of six year-old P. taeda. However, our higher study rate promoted a decrease in tree growth (likely due to some nutritional imbalance) similar to that observed by Rodrigues (2004).
Wood quality
Although increased growth of P. taeda in response to residue application resulted in decreased average wood density, weighted density was unaffected. This variation may be associated with tree growth rate, where higher growth rate results in lower density (Pretzsch et al. 2018; Antony et al. 2009). However, Castelo et al. (2008) observed the opposite for 14, 16, and 18 year-old P. taeda (i.e., higher growth rate resulted in higher density), while Albaugh et al. (2004) found no variation in wood density due to fertilization. It is important to note that our 7 year-old trees had not reached normal harvest age (typically 15 years in Brazil) and that older trees have more adult wood which tends to have higher density (Latorraca and Albuquerque 2000). Since wood density remained very close to and sometimes surpassed values of the pulp industry (0.405 g cm−3), the double increase in growth did not decrease tree quality.
Tissue elemental composition
Residue application caused Mn to vary more than other elements in plant tissues; this element decreased in needles, dead branches, live branches, bark, and wood. Rodriguez et al. (2018) found decreased Mn concentration in P. taeda woody tissue. Toxic effects of Mn have been observed in a gymnosperm native to southern Brazil (Araucaria angustifolia Bert. O. Ktze.), with an inverse relationship being observed between needle Mn concentration and growth (Hoogh 1981). This was also true in the present study based on regression tree results. Motta et al. (2014) indicated that low growth sites on acidic soils were characterized by high Mn and low Fe in leaf tissue. Given high soil acidity, Gomes et al. (2019) suggested that Pinus species are more sensitive to Mn than Al indicating possible Mn toxicity or imbalance. Analyzing Pinus oocarpa and Pinus caribaea var hondurensis, Gomes et al. (2019) found a fivefold increase in needle Mn concentration when soil pH was reduced from 5 to 4. Sikstrom et al. (2010) reported increased Mn in Pinus sylvestris leaf tissue due to use of alkaline ash. Working with P. taeda seedlings, South et al. (2017) did not observe changes in leaf Mn concentration with pH variation due to addition of acidifiers and acidity correctors. In the current study, despite no change in soil pH, residue addition was associated with less Mn absorption which could partially explain increased Ca availability in soil. However, the lack of Ca change in aboveground tissues indicates the need for more research to elucidate the effects of residue application on Mn and Ca dynamics in P. taeda plantations.
Concentrations of Mn in needles (190 mg kg−1) and bark (26 mg kg−1) may be related to increased average annual increment in P. taeda. This information indicates that bark could be used as an alternate tissue for nutrition diagnosis, which makes tissue collection easier. Other alternatives may be useful for other nutrient assessments; for example, root tissue has been proposed for P evaluations since collecting needles can be difficult, especially in older trees. High Mn values in needles tissue is not unusual (Consalter et al. 2020; Sass et al. 2020). In addition, K and Mg concentrations were close or slightly below critical levels (3.5 g kg−1 and 0.6 g kg−1, respectively) recognized for needle tissue (Pauletti and Motta 2018; Albaugh et al., 2008). High Mn levels could be related to nutritional imbalance rather than soil acidity based on evidence observed in the field, in control plot trees, and in areas adjacent to the experimental site.
In addition to Mn, decreases in Fe, Al, and B were observed in live branch and needle tissues. Decrease in P, Mg, and S observed in live branches (along with decreased P and Ca in needles) were in contrast to increases observed for soil P and Ca. This may have occurred due to increased tree growth without changing nutrient amount, consequently reducing nutrient concentrations (dilution effect). Likewise, Sass et al. (2020) reported P dilution in needles from the same forest site 3 years after residue application. It is import to note that dilution can result in P concentrations well below the critical level of 1.2 g kg−1 (Albaugh et al. 2010).
Therefore, the decrease in Mn and Al levels may have resolved a possible Mn toxicity or antagonistic effect interfering with absorption of other nutrients, thus increasing growth. Many studies have reported that high levels of exchangeable soil Al did not impact growth of Eucalyptus and Pinus species, indicating that both species are tolerant to this element (Gomes et al. 2019; Bassaco et al. 2018). The high values of macronutrients seen with the addition of 49 T ha−1 of residue were a result of better development and general tree growth with this rate of residue application.
Forest biomass nutrient content
Carbon stored in wood and bark was interesting since the concentration of this element in these fractions did not change (Table 5). Our work showed an increase of C in wood and bark of ~ 20 T ha−1 with the application of industrial residue (Table S4). Conditions favorable to improved soil nutrition can increase tree canopy growth and consequent accumulation of C (Samuelson et al. 2004; Tumushime et al. 2019). While Jokela et al. (2010) suggested that growth gains and C accumulation are greater in low fertility areas, Tumushime et al. (2019) reported that responses to fertilizer were lower in more productive areas.
Other nutrients usually tend to follow the accumulation pattern of biomass and C (Vogel et al., 2011). In our work, only N, K, Ca, Mg, and S in wood were affected by residue application (Table S4). This change was expected due to increases in the stand (Angel et al. 2019), however changes in other tissues and nutrients were also expected (Zhao et al. 2014).
Relating aerial biomass to nutrient content, work with 16 year-old (Sixel et al. 2015) and 25 year-old (Tumushime et al. 2019) P. taeda found that the needle fraction was the second most accumulator of aerial biomass behind wood tissue, which was different from our work that found an order of wood, bark, and live branches before needles. Although older forests tend to have higher amounts of nutrients than immature forests (Albaugh et al. 2004), P in wood shows little variation over time. We found 12.8 kg ha−1 of P (7 year-old), while Tumushime et al. (2019) found an average of 13.5 kg ha−1 (25 year-old), and Moro (2008) estimated 16–11 kg ha−1for 8, 12, 16 and 20 year-old forests. Greater P variations are found in needles due to age (Sixel et al. 2015; Tumushime et al. 2019). Thus, adequate diagnosis and nutritional management of sites may require nutrient monitoring in different forest compartments over time.
Litter
Total necromass found in this study (from 28.9 to 52.0 T ha−1) was smaller than observations by Trevisan et al. (1987) under similar conditions of soil and climate; their values ranged from 43.0 to 83.9 T ha−1 for 17 year-old P. taeda on three growth sites (poor, medium, and high). Differences between studies could be related to the age of our system at sampling (i.e., 7 year-old trees). In another Brazilian study, Rabel et al. (2020) found variation between 35.8 to 42.1 T ha−1 for 13 year-old P. taeda that received cellulose residue applications.
Results from our study confirms a high potential for litter accumulation in this area. For P. taeda systems, high potential for necromass accumulation on forest floors has been related to poor soil fertility and poor growth sites (Reissmann and Wisniewski 2015). Although results from the control indicate necromass accumulation potential, these results are inconsistent with increases in both forest floor necromass and tree growth as was observed in our study. Thus, more work is need to further evaluate the effect of fertility and fertilization on necromass accumulation on forest floors.
The observed trend for new FF accumulation as a result of residue application suggests some enhancement of litterfall deposition. Increases in old FF could be related to tree growth as well as possible addition of charcoal from residue application. Charcoal has been shown to have a long life span and has been observed in Amazonian dark earths (Terra Preta de Índio—TPI) on pre-Columbian Amerindian sites and near charcoal kiln production sites in Europe (Mastrolonardo et al. 2019; Hardy et al. 2017). Despite low change in soil properties, residue applications possibly influenced decomposition processes while adding recalcitrant C in the form of charcoal. Enhanced humification rate as proposed by Rabel et al (2020) may help explain FF increases from residue application. Our results confirmed higher level of necromass maintenance with addition of nutrients in the form of residue (Rabel et al., 2020) and lime/fertilizer (Consalter 2018). In contrast, Marschner and Wilczynski (1991) and Lundström et al. (2003) reported a large reduction in FF necromass from applying alkaline amendments (lime and ash), while Jandl et al. (2003) observed similar effects from a fertilizer/lime combination in European forests.
The variation in C for new FF (437 to 460 g kg−1) was very close to the range observed in needles (436 to 468 g kg−1—Table 4) which may reflect new litterfall input. Our C variation was larger than values reported by Marschner and Wilczynski (1991) for FF from limed (167 g kg−1) and control (267 g kg−1) forest plots. Prior to accounting for contaminants, these authors had corresponding values of 513 and 484 g kg−1 in these respective treatments. This illustrates that our new FF appears to represent the FF fraction with minimum disturbance and mineral contamination.
Despite our efforts towards extracting mineral contaminants, the reduction in C concentration from new FF to old FF clearly indicated a dilution from mineral contaminants as reported by Marschner and Wilczynski (1991). This dilution effect could explain the wide variation of C concentration found by Koschke et al. (2011); these authors evaluated 38 sites in Germany for the influence of black carbon and reported ranges of 230 to 484 g kg−1 and 66 to 462 g kg−1 for partially and highly decomposed FF, respectively. Mineral contamination of FF seems to be greatly influenced by litter age since Rabel et al. (2020) reported ash percentages ranging from 2.5% for fresh litter to more than 50% for the fine humidified FF fraction.
Increased litter due to residue application can be important for soil conservation due to the undulating landscape and sandy soil texture of the study region. Additionally, more than 7 T ha−1 of C was accumulated in litter at the residue rate that provided maximum growth (49 T ha−1) relative to the control. Drum et al. (2019) confirmed increased C sequestration in aboveground tissues and litter compartments as result of improved Pinus forest management.
Conclusions
Regardless of rate, P. taeda timber volume was greatly increased by industrial residue application, which suggests a high potential for yield enhancement. Since tree height was not influenced by residue application, increases were related to diameter improvements. Despite these improvements, there was no difference in the proportion of tree biomass components (wood, bark, needles, and live and dead branches). A possible direct relationship between enhanced tree growth and total litter mass suggests increased litterfall input. Fragmented and humified forest floor litter fractions became darker with increased residue rate, indicating that these fractions trap charcoal fragments associated with industrial residues. In contrast to tree growth, only a few soil property and tissue nutrient parameters showed slight changes, illustrating the difficulty of associating Pinus growth with these factors. Changes in soil properties were restricted to increased P and Ca availability, however decreases in these nutrients were observed in tree tissues, likely due to a dilution effect. Although no change in soil pH was observed in our study, the noted decreases in needle Mn, Fe, B, and Al would normally be related to increased soil pH. While excess Mn could compromise tree growth under acidic conditions (due to toxicity or imbalance with other nutrients), residue application was observed to decrease Mn in all evaluated tissues. Wood quality needed for production processes was not impacted since wood density remained within industry standards regardless of applied residue rate. Application of industrial residues to low fertility soils planted with P. taeda is recommended for accelerated growth and stand production.
References
Abreu AHM, Marzola LB, Melo LA, Leles PS, Abel ELS, Alonso JM (2017) Urban solid waste in the production of Lafoensia pacari seedlings. Revista Brasileira De Engenharia Agrícola e Ambiental 21(2):83–87. https://doi.org/10.1590/1807-1929/agriambi.v21n2p83-87
Águas Paraná - Instituto das Águas do Paraná (2018) Dados de precipitação diária do município de Piraí do Sul—Paraná
Albaugh TJ, Maier CA, Campoe OC, Yáñez MA, Carbaugh ED, Carter DR, Cook RL, Rubilar RA, Fox TR (2020) Crown architecture, crown leaf area distribution, and individual tree growth efficiency vary across site, genetic entry, and planting density. Trees 34:73–88. https://doi.org/10.1007/s00468-019-01898-3
Albaugh JM, Blevins L, Allen HL, Albaugh TJ, Fox TR, Stape JL, Rubilar RA (2010) Characterization of foliar macro- and micronutrient concentrations and ratios in loblolly pine plantations in the southeastern United States. South J Appl for 34:53–64. https://doi.org/10.1093/sjaf/34.2.53
Albaugh TJ, Allen HL, Fox TR (2008) Nutrient use and uptake in Pinus taeda. Tree Physiol 28:1083–1098. https://doi.org/10.1093/treephys/28.7.1083
Albaugh TJ, Allen HL, Dougherty PM, Johnsen KH (2004) Long term growth responses of loblolly pine to optimal nutrient and water resource availability. For Ecol Manag 192(1):3–19
Alvares CA, Stape JL, Sentelhas PC, Gonçalvez JLM, Sparovek G (2013) Köppen’s Climate Classification Map for Brazil. Meteorol Z 22(6):711–728
Alves MJF, Melo VF, Reissmann CB, Kaseker JF (2013) Reserva mineral de potássio em latossolo cultivado com Pinus taeda L. Revista Brasileira de Ciência do Solo 37:1599–1610. https://doi.org/10.1590/S0100-06832013000600016
Angel HZ, Priest JS, Stovall JP, Oswald BP, Weng Y, Williams HM (2019) Individual tree and stand-level carbon and nutrient contents across one rotation of loblolly pine plantations on a reclaimed surface mine. New for 50:733–753. https://doi.org/10.1007/s11056-018-09696-4
Antony F, Jordan L, Schimleck LR, Daniels RF, Clark A III (2009) The effect of mid-rotation fertilization on the wood properties of loblolly pine (Pinus taeda). IAWA J 30(1):49–58
Associação Brasileira de Normas Técnicas ABNT (2003) NBR 11941:2003: Madeira—Determinação da densidade básica - ABNT/CB-029 Celulose e Papel, Rio de Janeiro, p 6
Bassaco MVM, Motta ACV, Pauletti V, Prior AS, Nisgoski S, Ferreira CF (2018) Nitrogen, phosphorus, and potassium requirements for Eucalyptus urograndis plantations in Southern Brazil. New for 49:681–697. https://doi.org/10.1007/s11056-018-9658-0
Batista AH, Motta ACV, Reissmann CB, Schneider T, Martins IL, Hashimoto M (2015) Liming and fertilisation in Pinus taeda plantations with severe nutrient deficiency in savanna soils. Acta Sci Agron 37:117–125. https://doi.org/10.4025/actasciagron.v37i1.18061
Bellote AFJ, Tomazello FM, Dedecek RA (2005) Influência dos Nutrientes Minerais e do Solo sobre a Produtividade, a Estrutura Anatômica e a Densidade da Madeira de Pinus taeda. Documentos/embrapa Florestas, Colombo 58:1–94
Castelo PAR, Matos JLM, Dedecek RA, Lavoranti OJ (2008) Influência de diferentes sítios de crescimento sobre a qualidade da madeira de Pinus taeda. Floresta 38(3):495–506. https://doi.org/10.5380/rf.v38i3.12416
Consalter R (2018) Crescimento e nutrição de Pinus spp. em solos de baixa fertilidade no estado do Paraná. Dissertation, Universidade Federal do Paraná, Brazil
Consalter R, Barbosa JZ, Prior SA, Vezzani FM, Bassaco MVM, Pedreira GQ, Motta ACV (2020) Mid-rotation fertilization and liming effects on nutrient dynamics of Pinus taeda L. in subtropical Brazil. Eur J for Res 1:1. https://doi.org/10.1007/s10342-020-01305-4
Corrêa RS, Bellote AFJ (2011) Soil attributes and biomass yield from Pinus caribaea var hondurensis. Cerne 17(2):181–187. https://doi.org/10.1590/S0104-77602011000200004
Corrêa JC, Büll LT, Crusciol CAC, Marcelino R, Mauad M (2007) Correção da acidez e mobilidade de íons em Latossolo com aplicação superficial de escória, lama cal, lodos de esgoto e calcário. Pesq Agrop Brasileira 42(9):1307–1317. https://doi.org/10.1590/1807-1929/agriambi.v21n2p83-87
Drum CG, Vogel JG, Gezan SA, Jokela EJ (2019) Belowground processes for two loblolly pine (Pinus taeda L.) families respond differently to the intensity of plantation management. For Ecol Manage 441:293–301. https://doi.org/10.1016/j.foreco.2019.03.041
Empresa Brasileira de Pesquisa Agropecuária – EMBRAPA (2013) Sistema brasileiro de classificação de solos. 3ª edição. Brasília, DF: Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA), p 353
Empresa Brasileira de Pesquisa Agropecuária - EMBRAPA (2009) Manual de análises químicas de solos, plantas e fertilizantes. 2 ed. Brasília: Informação Tecnológica, p 628
Gatiboni LC, Da Silva WC, Mumbach GL, Schmitt DE, Iochims DA, Stahl J, Vargas CO (2020) Use of exchangeable and nonexchangeable forms of calcium, magnesium, and potassium in soils without fertilization after successive cultivations with Pinus taeda in southern Brazil. J Soils Sediments 20(2):665–674. https://doi.org/10.1007/s11368-019-02460-x
Gomes SS, Gonçalves JLM, Rocha JHT, Menegale MLC (2019) Tolerance of Eucalyptus and Pinus seedlings to exchangeable aluminium. Scientia Agricola 76(6):494–500. https://doi.org/10.1590/1678-992x-2018-0011
Guedes MC, Poggiani F (2003) Variação dos teores de nutrientes foliares em eucalipto fertilizado com biossólido. Scientia Forestalis 63:188–201
Hardy B, Cornelis JT, Houben D, Leifeld J, Lambert R, Dufey JE (2017) Evaluation of the long- term effect of biochar on properties of temperate agricultural soil at pre-industrial charcoal kiln sites in Wallonia, Belgium. Eur J Soil Sci 68(1):80–89. https://doi.org/10.1111/ejss.12395
Hohendadl W (1936) Der aufbau der Baumschafte. Forstwiss Forstwissenschaftliches Centralblatt 46:460–470
Hoogh R (1981) Site-nutrition- growth relationships of Araucaria angustifólia (Bert) O. Ktze in southern Brazil. 1981. Dissertation Doktorwurde – Der Forstwissenschaftlichen Fakultat der Albert- Ludwigs- Universitat zu Freiburg im Breisgau
Hothorn T, Hornik K, Zeileis A (2006) Unbiased recursive partitioning: a conditional inference framework. J Comput Graph Stat 15(3):651–674
IBÁ (Indústria Brasileira de Árvores) (2019) Dados e Estatísticas. IBA, Brasília: 2019, p 80
Ibrahim JFON, da Silva IV, Junior BFC, Paez DRM, Nascentes AL, da Silva LDB (2019) Utilização do lodo de esgoto na produção de mudas e no cultivo do eucalipto (Eucalyptus spp). Brazil J Anim Environ Res 2(1):564–579
Jacobson S (2003) Addition of stabilized wood ashes to Swedish coniferous stands on mineral soils—effects on stem growth and needle nutrient concentrations. Silva Fennica 37:437–450
Jandl R, Kopeszki H, Bruckner A, Hager H (2003) Forest soil chemistry and mesofauna 20 years after an amelioration fertilization. Restor Ecol 11(2):239–246. https://doi.org/10.1046/j.1526-100X.2003.00179.x
Jokela EJ, Martin TA, Vogel JG (2010) Twenty-five years of intensive forest management with southern pines: Important lessons learned. J Forest 108:338–347. https://doi.org/10.1093/jof/108.7.338
Koschke L, Lorz C, Fürst C, Makeschin F (2011) Black carbon in fly-ash influenced soils of the Dübener Heide Region, Central Germany. Water Air Soil Pollut 214:119–132. https://doi.org/10.1007/s11270-010-0409-x
Krishna MP, Mohan M (2017) Litter decomposition in forest ecosystems: a review. Energy Ecol Environ 2:236–249. https://doi.org/10.1007/s40974-017-0064-9
Latorraca JVF, Albuquerque CEC (2000) Efeito do rápido crescimento sobre as propriedades da madeira. Floresta e Ambiente 7(1):279–291
Lundström US, Bain DC, Taylor AFS, Van Hees PAW (2003) Effects of acidification and its mitigation with lime and wood ash on forest soil processes: a review. Water Air Soil Pollut 3(4):5–28
Marschner B, Wilczynski AW (1991) The effect of liming on quantity and chemical composition of soil organic matter in a pine forest in Berlin, Germany. Plant Soil 137:229–236. https://doi.org/10.1007/BF00011201
Martins, APL, Reissmann CB (2007) Determinação de nutrientes em material vegetal e extratos aquosos –manual operacional. UFPR, 2007.
Mastrolonardo G, Calderaro C, Cocozza C, Hardy B, Dufey J, Cornelis JT (2019) Long-term effect of charcoal accumulation in hearth soils on tree growth and nutrient cycling. Front Environ Sci 7:51. https://doi.org/10.3389/fenvs.2019.00051
Matysik MA, Gilmore DW, Mozaffari M, Halbach TR (2001) Application of wood ash, biosolids, and papermill residuals to forest soils—a review of the literature. Staff Paper Series no. 1531–23. St Paul: Department of Forest Resources, College of Natural Resources and Minnesota Agricultural Experiment Station, University of Minnesota
Meiwes KJ (1995) Application of lime and wood ash to decrease acidification of forest soils. Water Air Soil Pollut 85:143–152
Moro L (2008) Exportação de nutrientes em povoamentos de Pinus taeda L. baseada em volume estimado pelo sistema sispinus. Floresta 38:465–477
Motta ACV, Barbosa JZ, Consalter R, Reismann CB (2014) Nutrição e adubação da cultura do pinus. In: R.M. Prado, P.G.S, Wadt. Nutrição e adubação de espécies florestais e palmeiras. Gráfica e Editora Santa Terezinha, Jaboticabal, pp 383–425
Paim RM (2007) Efeito da aplicação de lama de cal e cloreto de potássio sobre as varáveis químicas e biológicas do solo, estado nutricional e crescimento do Pinus taeda L. sobre Latossolo em Três Barras, SC. Master Thesis, Universidade Federal do Paraná, Brazil
Paraná (2018) Instituto Agronômico do Paraná - IAPAR (Ed.). Cartas climáticas do Paraná, Brazil
Pauletti V, Motta ACV (2018). Manual de adubação e calagem para o estado do Paraná. Curitiba: Sociedade Brasileira de Ciência do Solo. Núcleo Estadual Paraná, p 482
Pretzsch H, Biber P, Schütze G, Kemmerer J, Uhl E (2018) Wood density reduced while wood volume growth accelerated in Central European forests since 1870. For Ecol Manage 429:589–616. https://doi.org/10.1016/j.foreco.2018.07.045
Rabel DDO, Maeda S, Araujo EM, Gomes JB, Bognolla IA, Prior SA, Magri E, Frigo C, Brasileiro BP, Santos MC dos P, Pedreira GQ, Motta ACV (2020). Recycled alkaline paper waste influenced growth and structure of Pinus taeda L. forest. New Forests https://doi.org/10.1007/s11056-020-09791-5.
Reissmann CB, Wisniewski C (2015) Aspectos nutricionais de plantios de pinus. In: Gonçalves JLM, Benedetti V (eds). Nutrição e fertilização florestal, Piracicaba: IPEF, pp 135–66
Richter DD, Markewitz D, Wells CG, Allen HL, April R, Heine PR, Urrego B (1994) Soil chemical change during three decades in an old-field loblolly pine (Pinus taeda L.) ecosystem. Ecology 75(5):1463–1473
Rocha GN, Goncalves JLM, Moura IM (2004) Mudanças da fertilidade do solo e crescimento de um povoamento de Eucalyptus grandis fertilizado com biossólido. Rev Bras Ciênc Solo 28(4):623–639
Rodrigues CM (2004) Efeito da aplicação de resíduo da indústria de papel e celulose nos atributos químicos, físicos e biológicos do solo, na nutrição e biomassa do Pinus taeda L. Master Thesis, Universidade Federal do Paraná
Rodriguez DRO, Andrade GC, Bellote AFJ, Tomazello FM (2018) Effect of pulp and paper mill sludge on the development of 17-year-old loblolly pine (Pinus taeda L.) trees in Southern Brazil. For Ecol Manag 422:179–189. https://doi.org/10.1016/j.foreco.2018.04.016
Samuelson LJ, Johnsen K, Stokes T (2004) Production, allocation, and stemwood growth efficiency of Pinus taeda L. stands in response to 6 years of intensive management. For Ecol Manag 192:59–70. https://doi.org/10.1016/j.foreco.2004.01.005
Sass AL (2016) Resíduo industrial celulósico no solo para melhorar o crescimento e a nutrição do Pinus taeda. Master Thesis, Universidade Federal do Paraná
Sass AL, Bassaco MVM, Motta ACV, Maeda S, Barbosa JZ, Bognola IA, Bosco JVG, Goularte GD, Prior SA (2020) Cellulosic industrial waste to enhance Pinus taeda nutrition and growth: a study in subtropical Brazil. Scientia Florestalis 48(126):3165. https://doi.org/10.18671/scifor.v48n126.13
Sikstrom U, Almqvist C, Jansson G (2010) Growth of Pinus sylvestris after the application of wood ash or P and K fertilizer to a peatland in southern Sweden. Silva Fennica 44:411–425
Silva VE, Silva PRT, Montanari R, Lisboa SDS, Batello ERB, Aguilar JV, Lisboa LAM, Albertini MM (2019) Produtividade de Pinus caribaea var. hondurensis e suas relações com atributos químicos dos solos em região de Cerrado brasileiro. Ciência Florestal 29(1):292–306
Sixel RMM, Arthur Junior JC, Gonçalves JLM, Alvares CA, Andrade GRP, Azevedo AC, Stahl J, Moreira AM (2015) Sustainability of wood productivity of Pinus taeda based on nutrient export and stocks in the biomass and in the soil. Rev Bras Ciênc Solo 39:1416–1427. https://doi.org/10.1590/01000683rbcs20140297
South DB, Nadel RL, Enebak SA, Bickerstaff G (2017) Sulfur and lime affect soil pH and nutrients in a sandy Pinus taeda nursery. Reforesta 4:12–20. https://doi.org/10.21750/REFOR.4.02.41
Trevisan E, Reissmann CB, Koehler JM, Lima JC (1987) Morfologia de horizontes orgânicos acumulados sob povoamentos de Pinus taeda L. em três sítios distintos. Revista Do Setor De Ciências Agrárias 9:59–62
Tumushime I, Vogel JG, Minor MN, Jokela EJ (2019) Effects of fertilization and competition control on tree growth and C, N, and P dynamics in a loblolly pine plantation in north central Florida. Soil Sci Soc Am J 83:242–251. https://doi.org/10.2136/sssaj2018.08.0289
Varnagiryte-kabašinskiene I, Stakenas V, Mikšys V, Kabašinskas A (2015) Vertical position of dry mass and elemental concentrations in Pinus sylvestris L. canopy under the different ash-nitrogen treatments. iForest - Biogeosciences and Forestry 8:838–845. https://doi.org/10.3832/ifor1342-007
Vogel JG, Suau LJ, Martin TA, Jokela EJ (2011) Long term effects of weed control and fertilization on the carbon and nitrogen pools of a slash and loblolly pine forest in north central Florida. Can J for Res 41:552–567. https://doi.org/10.1139/X10-234
World Reference Base for Soil Resources - WRB: A framework for international classification, correlation and communication. Food and Agriculture Organization of the United Nations. Rome, IUSS/ISRIC/FAO, 2006. 145p. (World Soil Resources Reports, 103)
Zhao D, Kane M, Teskey R, Markewitz D, Greene D, Borders B (2014) Impact of management on nutrients, carbon, and energy in aboveground biomass components of mid-rotation loblolly pine (Pinus taeda L.) plantations. Ann for Sci 71:843–851. https://doi.org/10.1007/s13595-014-0384-2
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pereira, M., Bassaco, M.V.M., Motta, A.C.V. et al. Influence of industrial forest residue applications on Pinus taeda: soil, litter, growth, nutrition, and wood quality characteristics. New Forests 54, 83–106 (2023). https://doi.org/10.1007/s11056-021-09902-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-021-09902-w