
Abstract
Promoting patchy recruitment of shade tolerant tree species into the midstory is an important step in developing structural diversity in second-growth stands. Variable-density thinning (VDT) has been proposed as a strategy for accelerating structural diversity, as its combination of within-stand treatments (harvest gaps, thinning, and non-harvested skips) should create variable overstory and understory conditions. Here we report on western hemlock (Tsuga heterophylla (Raf.) Sarg.) seedling and sapling densities in five mixed-conifer stands and Sitka spruce (Picea sitchensis (Bong.) Carr.) seedling and sapling densities in two stands in western Washington at 3,7, 10, and 16–17 years after VDT. Additionally, we report on western hemlock advance regeneration growth and survival in two stands over 14 years. Western hemlock seedling density was highest in the thinned treatment but only significantly so in Year 10. In contrast, the gaps contained significantly more western hemlock saplings in Years 7 and 10 and significantly greater growth of western hemlock advance regeneration through Year 10. Skips embedded within the VDT did not differ significantly from unharvested reserves in terms of seedling or sapling densities of either species. Sitka spruce seedling density was highest in the gap and thinned treatments, but saplings were uncommon in all treatments. Collectively, these results indicate that our variant of VDT promoted patchy, midstory recruitment of western hemlock but failed to recruit Sitka spruce saplings in either stand where it established. Consequently, more intensive variants of VDT may be required to promote midstory recruitment of species less tolerant of shade than western hemlock.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Restoring structural complexity to second-growth stands has become increasingly important in the mixed-conifer forests of the Pacific Northwest (PNW) (Donato et al. 2012; Franklin and Johnson 2012). Interest in restoring structural diversity increased in response to the Northwest Forest Plan, which prioritized creating habitat for biodiversity conservation over timber production on federal lands (Thomas et al. 2006). Compared to old growth, second-growth stands are deficient in many of the structural characteristics that contribute to late-successional habitat (Carey and Johnson 1995; Carey 2003) including: coarse woody debris (CWD) (Spies et al. 1988; Tyrrell and Crow 1994), cavity trees and snags (Goodburn and Lorimer 1998; McGee et al. 1999), large diameter trees (Spies and Franklin 1991), and spatial heterogeneity (Spies 2004; Donato et al. 2012). Second-growth stands also feature greater canopy continuity and less microclimate diversity than old growth (Franklin and Van Pelt 2004), raising concerns over the loss of understory biodiversity and tree regeneration. Restoring structural attributes such as large diameter trees and CWD is a long-term process that could take several decades in second-growth stands (Harmon et al. 1986; Stone et al. 1998). In contrast, obtaining herbaceous species diversity and stimulating the recruitment of shade tolerant conifers to increase vertical and horizontal heterogeneity can be achieved more rapidly with silvicultural intervention (Puettmann et al. 2016). However, studies examining the effectiveness of silvicultural treatments beyond the first decade are needed to determine whether structural development is continuing at an accelerated pace.
Variable-density thinning (VDT) is one approach that has been applied to enhance structural diversity in even-aged stands (Ashton and Kelty 2018). VDT differs from traditional thinning in that its primary goal is to increase structural diversity through varying overstory density within a stand (Carey 2003; Comfort et al. 2010). Many variants of VDT utilize a combination of within-stand treatments including retaining unharvested small patches (hereafter referred to as skips), thinning at varying intensities, and creating canopy gaps to promote heterogenous conditions (Dodson et al. 2014; Berrill et al. 2018; Dagley et al. 2018). The varying microclimates produced across the VDT treatments have been shown to promote a range of responses in diameter and height growth (Davis et al. 2007; Roberts and Harrington 2008; Comfort et al. 2010; Sullivan and Sullivan 2016; Willis et al. 2018), crown length (Sullivan and Sullivan 2016; Seidel et al. 2016; Willis et al. 2018), and understory cover and richness (Bailey and Tappeiner 1998; Ares et al. 2009, 2010; Sullivan and Sullivan 2016; Cole et al. 2017; Curtis et al. 2017), thereby setting the stand on a trajectory towards greater structural complexity and species diversity.
Promoting the recruitment of shade tolerant conifers into the midstory is an important step in accelerating the development of late-successional habitat in second-growth stands (Spies and Franklin 1991; Carey 2003). Unlike variable-retention harvesting, which seeks to regenerate new seed sources in older stands while retaining biological legacies (i.e., is a variant of final or regeneration harvesting) (Nyland 2016), VDT seeks to create spatial heterogeneity and multi-layered canopies but retain much of the previous overstory for a deferred harvest (Franklin et al. 2018). Given the presence of advance regeneration, VDT may be capable of quickly achieving structural restoration objectives (Comfort et al. 2010). However, in stands lacking advance regeneration, developing structural diversity will require the recruitment of a new cohort.
Patchy regeneration may be promoted by VDT as increasing spatial variation in overstory density alters resources available to the midstory and understory. Canopy gaps offer several potential advantages for conifer regeneration and have been shown to benefit conifer recruitment (Gray and Spies 1996; Coates 2002). For example, canopy gaps have been shown to provide favorable environments for seedling germination and early survival (Wright et al. 1998; Coates 2002). Canopy gaps may also improve early seedling survival by reducing the risk of leaf smothering, which can be problematic for species such as western hemlock (Tsuga heterophylla (Raf.) Sarg.) and Sitka spruce (Picea sitchensis (Bong.) Carr.), which have small seeds and short germinants (Thornburgh 1969; Minore 1979; Packee 1990). Due to their higher understory light and moisture availability (Gray et al. 2002), canopy gaps also promote seedling growth (Gray and Spies 1996; Nabel et al. 2013; DeMontigny and Smith 2017) and may release advance regeneration into larger size classes (Dodson et al. 2014; Puettmann et al. 2016).
Nevertheless, several factors can limit midstory development within gaps. Although initially promoted in gaps, seedling and sapling growth and survival may decline once root and crown expansion from bordering trees limits resource availability (Pacala et al. 1994; Gray et al. 2002; Walters et al. 2006). Growth and survival may also be stunted by the presence of a dense shrub layer, which can limit sapling development through resource competition (Roberts et al. 2005; Harrington 2006; Maguire et al. 2009). Declining resources within gaps may also alter species composition by providing a competitive advantage to species such as western hemlock, which is adept at surviving in extreme shade (Minore 1979; Packee 1990). Finally, advance regeneration may be damaged during the harvesting operation (Harrington et al. 2005), potentially causing high rates of mortality (Tesch et al. 1986) or reducing future growth (Youngblood 2000; Newton and Cole 2006).
Fewer resources are available to conifer regeneration in the thinned and skip treatments compared to the gaps. Previous studies have found abundant seedling establishment following thinning (Bailey and Tappeiner 1998); however, sapling recruitment has been limited in all but the most intensive thinning treatments (Nabel et al. 2013; Dodson et al. 2014). Similarly, the low resource environments of unthinned areas have been shown to inhibit seedling establishment and survival (Gray and Spies 1996; Coates 2002). In a VDT, skips (unthinned areas) and gaps can be nested within a matrix of thinned areas, creating the potential for edge effects. Previous studies have shown that trees established near a gap edge or skid trail respond with increased diameter growth (Roberts and Harrington 2008; Gray et al. 2012). Thus, skips nested within VDT could provide a different environment for seedling establishment and growth than the larger unharvested areas reported in previous studies (Gray and Spies 1996; Coates 2002).
This study examined the establishment of western hemlock and Sitka spruce seedlings (5–130 cm in height) and recruitment of western hemlock saplings (> 130 cm height, 0–4.9 cm dbh) at intervals of 4, 7, 10, and 16–17 years after implementing VDT in five second-growth mixed-conifer stands in western Washington. In addition, we report on the 14-year growth and survival of western hemlock large advance regeneration (sapling size class: > 130 cm height, 0–4.9 cm dbh prior to project initiation) after implementation of VDT in two second-growth mixed conifer stands. Specifically we were interested in whether: (1) seedling and sapling densities of shade tolerant conifers in the skip treatment approximated that in unharvested reserves; (2) gaps promoted greater seedling and sapling densities of shade tolerant conifers than the skips or thinned treatments; (3) increasing shrub cover was associated with reduced seedling and sapling densities of shade tolerant conifers; (4) the effects of individual VDT treatments on seedling and sapling densities of shade tolerant conifers were sustained over time; (5) western hemlock and Sitka spruce responded differently to individual VDT treatments; and (6) gap creation hindered or promoted the survival and growth of large advance regeneration of shade tolerant conifers. The findings from this study will provide an extended evaluation of the effectiveness of VDT at promoting the recruitment of shades tolerant conifers into the midstory in second-growth stands and increase our general understanding of regeneration dynamics in the PNW.
Methods
Site description
Data were collected from five stands within the Olympic Habitat Development Study (OHDS) on the Olympic Peninsula in western Washington, USA (Fig. 1). Sites ranged in elevation from 150 m to nearly 600 m. Annual precipitation at our sites ranged from 1460 to 3362 mm and occurred primarily as winter rain. Temperature in Forks, WA ranged from an average high of 24.0 °C in August to an average low of 0.7 °C in December (Willis et al. 2018). The soil types at all sites were characterized as deep, well drained loams. Conditions at the sites were typical of second-growth mixed-conifer stands in the region (Table 1).

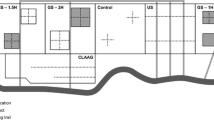
Area map illustrating the location of the five study stands on the Olympic National Forest in western Washington where measurements were collected
Treatments
Each stand was managed under a VDT approach consisting of three overstory treatments: thinning, small gaps, and uncut skips. VDT was implemented over 6–10 ha in each stand. In addition, a similarly sized area was left unharvested to serve as a control (hereafter referred to as the reserve) within each stand. Our VDT prescription designated 75% of the area to receive a general thinning, 10% to be in gaps and 15% in skips (Fig. 2). The thinning treatment was implemented as a thinning from below with 25% of the basal area removed from the lower crown classes. Gaps, or expanded existing openings, were approximately 0.04–0.05 ha in size and were created by cutting all stems of the dominant species. This range of gap sizes was selected to reduce potential windthrow risk. Retention preference in the thinned and gap treatments was given to hardwoods and less common conifer tree species, such as western redcedar, Pacific yew (Taxus brevifolia Nutt.), or Pacific silver fir (Abies amabilis Dougl. Ex Forbes), with the goal of increasing tree species diversity at the stand-level. Favoring hardwoods and less common conifer species resulted in a limited number of gaps retaining 1–3 small trees. Gaps were non-randomly located to avoid being in proximity to a skip (Fig. 2). The skips ranged in size from 0.1 to 0.3 ha and were non-randomly located with the intention of preserving existing late successional structures such as large-diameter snags or CWD. No harvesting or mechanized traffic occurred within the skips. The purpose of the reserve was to examine whether the skips were functioning like larger, unharvested areas in terms of their effects on seedling and sapling densities.

An example arrangement of overstory treatments within a VDT. Yellow squares represent areas where harvest gaps (0.04–0.05 ha) were created, green squares depict the location of unharvested skips (0.1–0.3 ha), while white squares (each 10 m × 10 m) were thinned from below with 25% of the basal area removed. Gray squares represent natural clearings. Red circles represent circular plots, while the dashed black lines denote the boundary of the stem-mapped plot
Field measurements
We recorded seedling (5–130 cm height) and sapling (> 130 cm height, 0–4.9 cm dbh) densities at 3, 7, 10, and 16–17 years after VDT. Seedlings and saplings were recorded within 2-m fixed-radius circular plots (hereafter referred to as circular plots) located within the VDT and reserve at each stand. Stratified sampling included 24–30 circular plots in the VDT and in 10–12 circular plots in the reserve (Fig. 2). The higher sampling intensity within the VDT was needed to account for the increased spatial variability introduced by the gap and skip treatments. Percent cover of all shrubs was recorded on concentric 5.64-m radius fixed-area plots centered on the circular plots each measurement year. Total shrub cover was estimated to the nearest 5% [or nearest 1% for circular plots where shrubs were uncommon (0–5%) or dominant (95–100%)].
We also tracked tagged western hemlock large advance regeneration (sapling size class: > 130 cm height, 0–4.9 cm dbh prior to project initiation) in a stem-mapped plot (approximately 1.5 ha) established within the VDT (Fig. 2). The goal of this monitoring was to have a more precise measure of advance regeneration survival and recruitment into larger size classes than could be obtained from the count data collected in the circular plots. All western hemlock large advance regeneration were tagged and measured immediately prior to harvesting and remeasured for diameter growth and at ages 5, 10, and 14. Survival was assessed on all tagged western hemlock large advance regeneration at age 14. Large advance regeneration was not tagged in the reserve. Sitka spruce large advance regeneration was assessed but not analyzed due to low representation across treatments and stands.
Data analysis
Seedling and sapling densities
Preliminary analysis for seedling density indicated a strong species effect, prompting us to analyze western hemlock and Sitka spruce separately following the same protocol. Circular plots located within each stand (Rail, Snow White, Fresca, Clavicle, Bait) were included in the analysis for western hemlock, but low seedling density across all overstory treatments limited our analysis of Sitka spruce to data from Clavicle and Fresca.
We used a repeated-measures, mixed-effects analysis of variance (ANOVA) to model changes in seedling and sapling densities in the circular plots located within the VDT and reserve at each stand across years. In the analyses of western hemlock seedling and sapling densities, overstory treatment (gap, thin, skip, reserve), time (3, 7, 10, 16–17 years), and shrub cover were considered fixed-effects. Stand was considered a random-effect for both western hemlock analyses. The analysis of Sitka spruce seedling density modeled the same factors but considered stand a fixed effect due to Sitka spruce only occurring at two stands (Fox et al. 2015). Circular plots were considered random sub-samples in both species’ models. A spatial Gaussian structure was chosen to model serial correlation after comparing multiple models with Akaike’s Information Criterion (AIC). A log (x + 1) transformation was needed to satisfy the assumptions of normality and equal variance in all analyses.
Factors influencing seedling and sapling densities were identified through backwards selection. For both analyses, the full model contained the main effects of overstory treatment, time, shrub cover, and stand. Additionally, we examined two-way interactions between overstory treatment x time, overstory treatment x shrub cover, and a three-way interaction between overstory treatment, time, and shrub cover. Initial model runs producing an interaction-term exceeding the recommended threshold for pooling (P > 0.25) resulted in the elimination of the highest-order interaction with the highest P value (Bancroft 1964). We repeated this procedure iteratively until all interaction terms fell below the pooling threshold. Significant effects of stand, overstory treatment, time, or overstory treatment x time were further investigated with a Tukey’s adjusted multiple comparison test. A critical value of α = 0.05 was used to determine significance for main effects and interactions for all tests. All analyses were conducted in SAS (Version 9.4, Cary NC).
Advance regeneration dynamics
Survival of 1472 western hemlock large advance regeneration was modeled with logistic regression. Insufficient large advance regeneration (n < 20) at Bait, Clavicle, and Fresca limited our analysis to Rail and Snow White. Survival was assessed at 14 years post-treatment, and tested for stand, overstory treatment, and initial size effects. Factors influencing large advance regeneration survival were investigated through backwards selection following the protocol described for our analysis of seedling and sapling densities. The full model contained the main effects of overstory treatment (gap, thin, skip), stand (Rail and Snow White), initial diameter, and the interaction between overstory treatment and initial diameter. The full model was weighted to adjust for the higher density of western hemlock large advance regeneration in the thinned overstory treatment.
Mean annual diameter growth between Years 1–5, 6–10, and 11–14 of all surviving large advance regeneration was modeled with a repeated-measures, mixed-effects analysis of variance. Backwards selection was used to identify factors influencing diameter growth. The full model contained the fixed-effects of overstory treatment, stand, initial sapling size, time (5, 10, 14 years), and the two-way interactions between overstory treatment × time and overstory treatment × initial sapling size. A spatial Gaussian structure was chosen to model serial correlation after comparing multiple models with Akaike’s Information Criterion (AIC). Significant effects of stand, overstory treatment, time, and overstory treatment x time were further investigated with a Tukey’s adjusted multiple comparison test. A critical value of α = 0.05 was used to determine significance for main effects and interactions for all tests. In addition to our growth analysis, we examined the development of large advance regeneration by diameter size class within overstory treatments at Rail and Snow White using a Sankey diagram (https://bl.ocks.org/d3noob/5028304).
Results
Seedling and sapling densities
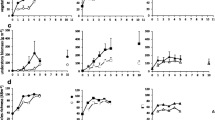
Overall seedling density averaged 159,900 seedlings per ha with western hemlock (142,550 seedlings per ha) as the dominant seedling species in all stands (Table 1). Overstory treatment effects on western hemlock seedling density varied significantly over time (P < 0.0001) (Table 2). After Year 3, in which no significant differences were detected, trends emerged among overstory treatments (Fig. 3). The thinned treatment consistently had the highest seedling density and significantly exceeded the other treatments in Year 10. Although generally higher across measurement years, seedling density in the skip treatment did not differ significantly from that in the reserve. Lastly, seedling density in the thinned treatment only significantly exceeded the gap treatment in Year 10. Shrub cover was highest in the gap treatment (29% ± 3) and lowest in the skip treatment (11% ± 3) but was not significantly correlated with western hemlock seedling density (P = 0.4582) (Table 2). No other investigated interactions significantly influenced western hemlock seedling density.

Average seedling and sapling density per hectare for western hemlock and seedling density per hectare for Sitka spruce within each overstory treatment (gap, thin, skip, reserve) at each measurement year (3, 7, 10, 16–17). At each year, overstory treatments with differing letters were found to be significantly different (Tukey’s HSD post hoc comparisons, α = 0.05). Error bars represent 1 SE
Sitka spruce seedling density across Clavicle and Fresca averaged 44,500 seedlings per ha. Seedling density did not significantly vary between Clavicle (49,750 ± 14,200) and Fresca (41,500 ± 14,600) (P = 0.6469) (Table 2). Like western hemlock, overstory treatment effects varied significantly over time (P = 0.0003) (Table 2). After Year 3, Sitka spruce seedling density was generally higher in the gap and thinned treatments compared to the skips or reserve (Fig. 3). In Year 7, Sitka spruce seedling density in the gap treatment significantly exceeded that in the other treatments. Finally, seedling density in the skip treatment did not differ significantly from the reserve in any measurement year. Neither the main effects of shrub density (P = 0.1250) (Table 2) nor any other investigated interactions significantly influenced Sitka spruce seedling density.
Overall, western hemlock sapling density averaged 17,550 saplings per ha. The effect of overstory treatment on western hemlock sapling density also varied significantly with time (P = 0.0002) (Table 2). Western hemlock sapling density was significantly higher in the gap treatment than in the other overstory treatments in Years 7 and 10 (Fig. 3). However, the gap and thinned treatments did not differ in sapling density in Year 17. No significant differences in sapling density were detected in any measurement year between the skip treatment and the reserve. Neither the main effects of shrub cover (P = 0.6308) (Table 2) nor any other investigated interactions significantly affected western hemlock sapling density. Sitka spruce saplings were only present at Clavicle; where they averaged 1750 saplings per ha and were present on only 20% of the circular plots.
Advance regeneration
After 14 years, survival of western hemlock large advance regeneration averaged 90%. Initial sapling size had a significant positive association with survival (P = 0.0002), but this relationship was extremely weak (R2 < 0.01) (Table 3). Survival of western hemlock advance large regeneration was high in both stands but survival at Rail (95% ± 2) significantly exceeded that at Snow White (88% ± 1) (P < 0.0001) (Table 3). Among overstory treatments, survival of western hemlock large advance regeneration was generally high and did not statistically differ (P = 0.0812) (Table 3). In addition, the effect of overstory treatment on western hemlock large advance regeneration survival was not significantly influenced by initial size (P = 0.1995) (Table 3).
Annual diameter growth of western hemlock large advance regeneration averaged 0.23 cm year−1. Growth was significantly affected by stand (P < 0.0001), initial diameter (P < 0.0001), time (P < 0.0001) overstory treatment (P < 0.0001), and the interaction between overstory treatment and time (P < 0.0001) (Table 3). The annual growth rate at Rail (0.31 cm year−1 ± < 0.01) significantly exceeded growth at Snow White (0.19 cm year−1 ± < 0.01). Like survival, diameter growth was positively associated with initial size of the advance regeneration, but this relationship was also extremely weak (R2 = 0.01). The gap treatment supported higher diameter growth over the first decade but did not significantly exceed growth in the thinned treatment after Year 10 (Fig. 4). Diameter growth of western hemlock large advance regeneration declined in all treatments between Years 11 and 14.

The effect of overstory treatments (gap, thin, skip) on mean annual diameter growth (cm) of western hemlock large advance regeneration (sapling size class: > 130 cm height, 0–4.9 cm dbh prior to project initiation) over 14 years. At each year, overstory treatments with differing letters were found to be significantly different (Tukey’s HSD post hoc comparisons, α = 0.05). Error bars represent 1 SE
Variable-density thinning had a similar effect on size class development of western hemlock large advance regeneration at Snow White and Rail. At both stands, the gap treatment promoted the highest annual progression of western hemlock large advance regeneration into the largest size class (> 8.1 cm) (0.9% at Snow White and 2.4% at Rail), while the skip treatment supported the lowest (0.1% at Snow White and 0.2% at Rail) (Fig. 5). Although Snow White initially supported a higher density of western hemlock large advance regeneration in the 2.6–5.0 size class at Year 0, Rail supported a higher total percentage in the largest size class by Year 14. The percentage of western hemlock large advance regeneration growing into the next diameter class was similar at both stands between Years 0–5, but a greater percentage advanced at Rail during the two subsequent measurement periods.

Development of western hemlock large advance regeneration (sapling size class: > 130 cm height, 0–4.9 cm dbh prior to project initiation) within overstory treatments (gap, thin, skip) at Snow White and Rail over 14 years. Each color represents a different diameter size class (or mortality). The numbers within the bars represent the number of seedlings in each size class
Discussion
Seedling density
Promoting patchy recruitment of shade tolerant conifers into the midstory in second-growth stands is an important step in restoring late successional habitat (Spies and Franklin 1991; Bauhus et al. 2009). Stands with existing advance regeneration will likely require less time to achieve structural restoration objectives if treated with VDT than stands where saplings must be developed from a new cohort (Comfort et al. 2010). However, as evident by a sub-set of stands included in this study, advance regeneration may not be available for recruitment in all stands.
Achieving patchy seedling establishment is not a late successional habitat restoration objective (Spies and Franklin 1991; Bauhus et al. 2009). Nevertheless, patterns in seedling establishment may indicate if VDT is having its desired effect on stand dynamics. Our variant of VDT created variation in seedling density among overstory treatments for western hemlock and Sitka spruce within the first decade following treatment. However, western hemlock and Sitka spruce differed in their response to VDT over time. Consistent with previous studies of natural regeneration, Sitka spruce seedling density was highest within the gap treatment in Year 7, while western hemlock seedling density was generally highest after Year 3 in the thinned treatment (Gray and Spies 1996; LePage et al. 2000; Quine 2001; Kuehne and Puettmann 2008; Nabel et al. 2013). Differences in seedling density among overstory treatments also developed more rapidly with Sitka spruce than western hemlock. Although seed rain was not measured directly, it is unlikely that differences in seed availability strongly influenced our results, as both species were well distributed within stands, and the distance from edge trees to gap center was well within the seed dispersal capabilities of both species (LePage et al. 2000; Beach and Halpern 2001). Western hemlock and Sitka spruce also preferentially establish on coarse woody debris and mineral soil compared to other substrates (Harmon and Franklin 1989; Gray and Spies 1996; LePage et al. 2000), suggesting potential differences in substrate availability among overstory treatments would not strongly affect either species. One plausible explanation for our results lies in the varying tolerance of western hemlock and Sitka spruce seedlings to resource availability within the overstory treatments. Due to its sensitivity to drought, shallow root system, and ability to germinate in shade, western hemlock seedling establishment has been shown to benefit from partial shade (Minore 1979; Gray and Spies 1996; Bennett et al. 2002; Gavin and Hu 2006). Given the greater shade tolerance of western hemlock (Minore 1979; Packee 1990; Bianchi et al. 2018), its seedlings are also more likely to persist in deep and partial shade than Sitka spruce (Mason et al. 2004). Thus, even though both species maintained large seedling banks in the gap and thinned treatments, the differing patterns in average seedling density are consistent with the silvics of each species (Harris 1990; Packee 1990). We acknowledge, however, that our analyses of seedling density were based on count data, which did not differentiate between changes in density caused by mortality or advancement to the sapling size class.
Sapling density and development of advance regeneration
Establishing seedlings of shade tolerant species has not been historically challenging in temperate ecosystems (Neuendorff et al. 2007; Kuehne and Puettmann 2008; Kern et al. 2017). More problematic has been creating conditions that allow for development beyond the seedling stage and out of range of many of the factors that threaten seedling survival on the forest floor (Webster et al. 2018). The lack of recorded saplings across all stands, treatments, and years indicates that none of the overstory treatments promoted Sitka spruce sapling development. It is likely that the thinning intensity used in this study was too low to stimulate Sitka spruce development, as previous studies have found that Sitka spruce regeneration increases with harvesting intensity (Deal and Tappeiner 2002). Moreover, this result corresponds with previous research suggesting that Sitka spruce requires gaps larger than 400 m2 to persist (Taylor 1990; Quine 2001), but contrasts with the findings of Page and Cameron (2006) who reported Sitka spruce persisting within gaps approximately 400 m2 in size. Although resource availability was not measured within the gaps, intense asymmetric competition for resources from edge trees may have constrained Sitka spruce sapling density (Gray et al. 2002; Walters et al. 2006), as the average height of residual trees in our stands (43–49 m at Fresca and Clavicle) far exceeded those reported by Page and Cameron (2006). The effect of larger edge trees on light availability in the gaps may also have been magnified by the low sun angle at the northerly latitude (approximately 47 °N) of our stands (Canham et al. 1990). Consequently, larger harvest gaps may be needed to recruit Sitka spruce saplings into the midstory in 60-to-90-year-old second-growth stands in the PNW.
In contrast to Sitka spruce, sapling density of western hemlock was consistently highest in the gaps. Given its ability to respond to increases in light availability (Coates and Burton 1999), it is not surprising that western hemlock sapling density was maximized within the gap treatment. This finding supports previous reports documenting successful recruitment of western hemlock within canopy gaps (Coates 2002; Nabel et al. 2013).
Limiting sapling recruitment in the skip treatment contributes to the overall development of patchy midstory structure within a VDT. Due to their relatively small size (0.1–0.3 ha) and nested location within the thinned matrix, edge effects from bordering overstory treatments could alter the function of VDT skips (Roberts and Harrington 2008; Willis et al. 2018); however, to our knowledge no previous study has compared sapling recruitment in VDT skip to an unharvested control to see if this occurs. Our results indicate that VDT skips functioned similarly to unharvested controls, as western hemlock sapling density in the skip treatment never exceeded that in the reserve. It should be recognized, however, that sapling density was highly variable in the skip and reserve treatments, and that sapling density of western hemlock in the skip treatment was never statistically lower than that in the thinned treatment. Thus, factors beyond harvesting intensity and skip size are contributing to sapling density, which may lead to differing responses to our variant of VDT if it is applied in other locations.
Advance regeneration is often stimulated by overstory harvesting and can contribute to midstory diversity (Shatford et al. 2009; Comfort et al. 2010). Compared to the other overstory treatments, western hemlock large advance regeneration in the gaps survived at a relatively high rate (86%) and grew at the fastest rate through the first decade. This finding is somewhat surprising considering that the odds of damage were likely highest in the gap treatment, where harvesting traffic was most intense. Moreover, harvesting damage has been shown to reduce survival and growth of advance regeneration of several shade tolerant conifer species (Newton and Cole 2006). The rapid initial growth response observed within the gaps suggests that damage to western hemlock large advance regeneration was minimal. Thus, our results are consistent with the suggestion of Puettmann et al. (2016) that the positive benefits of entering a stand to increase resource availability outweigh the potential risk of damage caused by stand entry.
Shrub competition
Competition from shrubs represents another potential challenge in developing midstory structural diversity (Royo and Carson 2006). Previous studies in the PNW have reported negative impacts of shrubs on growth of western hemlock seedlings in clear-cut stands (Mitchell et al. 2004; Harrington 2006). Consistent with other studies from thinned stands (Devine and Harrington 2008; Dodson et al. 2014), increasing shrub cover was not associated with declining seedling density of either species or sapling density of western hemlock in our study. Moreover, our results generally support the notion that shade tolerant conifers are relatively insensitive to understory competition (Beach and Halpern 2001). Thus, the lack of shrub competition observed in this study may have resulted from the combination of shrub suppression by the residual overstory and the shade tolerance of western hemlock and Sitka spruce (Minore 1979). Previous studies examining understory competition in thinned stands or small canopy gaps have also reported limited interference by shrubs (Gray and Spies 1997; Miller and Emmingham 2001; Kuehne and Puettmann 2008).
Conclusions
The primary objective of the Olympic Habitat Development Study was to determine whether VDT could accelerate the development of forest structure associated with late successional stands in much younger stands. We observed that sapling density of western hemlock was high in all overstory treatments, suggesting that some recruitment of western hemlock saplings into the midstory will occur regardless of silvicultural treatment. However, in terms of accelerating structural development, the gap treatment generally supported the highest western hemlock sapling density, large advance regeneration growth, and recruitment into larger size classes (> 8.1 cm), demonstrating that our variant of VDT at least partially succeeded in developing additional structural diversity in the midstory within the first decade after treatment. Our results also confirm that embedded skips (0.1–0.3 ha) did not differ from unharvested controls in seedling density of either species or sapling density for western hemlock; thus, the skips were effective in retaining some of the characteristics associated with untreated stands and contributed to the spatial diversity created by this version of VDT. In contrast to our results with western hemlock, our variant of VDT failed to recruit Sitka spruce saplings. Although our results for Sitka spruce are limited to the two stands in which it occurred, the overall dearth of saplings suggests that larger harvest gaps or a more intensive thinning prescription may be needed to promote midstory recruitment for species less tolerant of shade than western hemlock.
References
Ares A, Berryman SD, Puettmann KJ (2009) Understory vegetation response to thinning disturbance of varying complexity in coniferous stands. Appl Veg Sci 12:472–487
Ares A, Neill AR, Puettmann KJ (2010) Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. For Ecol Manag 260:1104–1113
Ashton MS, Kelty MJ (2018) The practice of silviculture: applied forest ecology. Wiley, Hoboken
Bailey JD, Tappeiner JC (1998) Effects of thinning on structural development in 40-to 100-year-old Douglas-fir stands in western oregon1. For Ecol Manag 108:99–113
Bancroft TA (1964) Analysis and inference for incompletely specified models involving the use of preliminary tests of significance. Biometrics 20:427–442
Bauhus J, Puettmann K, Messier C (2009) Silviculture for old-growth attributes. For Ecol Manag 258:525–537
Beach EW, Halpern CB (2001) Controls on conifer regeneration in managed riparian forests: effects of seed source, substrate, and vegetation. Can J For Res 31:471–482
Bennett JN, Andrew B, Prescott CE (2002) Vertical fine root distributions of western redcedar, western hemlock, and salal in old-growth cedar hemlock forests on northern Vancouver Island. Can J For Res 32:1208–1216
Berrill JP, Dagley CM, Gorman AJ, Obeidy CS, Powell HK, Wright JC (2018) Variable-density retention promotes spatial heterogeneity and structural complexity in a Douglas-fir Tanoak stand. Curr Trends For Res 1:1–9
Bianchi S, Hale S, Cahalan C, Arcangeli C, Gibbons J (2018) Light-growth responses of Sitka spruce, Douglas fir, and western hemlock regeneration under continuous cover forestry. For Ecol Manag 422:241–252
Canham CD, Denslow JS, Platt WJ, Runkle JR, Spies TA, White PS (1990) Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can J For Res 20:620–631
Carey AB (2003) Bio-complexity and restoration of biodiversity in temperate coniferous forest: inducing spatial heterogeneity with variable-density thinning. Forestry 76:127–136
Carey AB, Johnson ML (1995) Small mammals in managed, naturally young, and old-growth forests. Ecol Appl 5:336–352
Coates KD (2002) Tree recruitment in gaps of various size, clearcuts and undisturbed mixed forest of interior British Columbia, Canada. For Ecol Manag 155:387–398
Coates KD, Burton PJ (1999) Growth of planted tree seedlings in response to ambient light levels in northwestern interior cedar-hemlock forests of British Columbia. Can J For Res 29:1374–1382
Cole E, Newton M, Bailey JD (2017) Understory vegetation dynamics 15 years post-thinning in 50-year-old Douglas-fir and Douglas-fir/western hemlock stands in western Oregon, USA. For Ecol Manag 384:358–370
Comfort EJ, Roberts SD, Harrington CA (2010) Mid-canopy growth following thinning in young-growth conifer forests on the Olympic Peninsula western Washington. For Ecol Manag 259:1606–1614
Curtis RO, Harrington CA, Brodie LC (2017) Stand development 18 years after gap creation in a uniform Douglas-fir plantation. Res. Pap. PNW-RP-610. Portland, OR: US Department of Agriculture, Forest Service, Pacific Northwest Research Station. 28 p., 610
Dagley CM, Berrill JP, Leonard LP, Kim YG (2018) Restoration thinning enhances growth and diversity in mixed redwood/Douglas-fir stands in northern California, USA. Restor Ecol 6:1170–1179
Davis LR, Puettmann KJ, Tucker GF (2007) Overstory response to alternative thinning treatments in young Douglas-fir forests of western Oregon. Northwest Sci 81:1–14
Deal RL, Tappeiner JC (2002) The effects of partial cutting on stand structure and growth of western-hemlock Sitka spruce stands in southeast Alaska. For Ecol Manag 159:173–186
DeMontigny LE, Smith NJ (2017) The effects of gap size in a group selection silvicultural system on the growth response of young, planted Douglas-fir: a sector plot analysis. For Int J For Res 90:426–435
Devine WD, Harrington TB (2008) Belowground competition influences growth of natural regeneration in thinned Douglas-fir stands. Can J For Res 38:3085–3097
Dodson EK, Burton JI, Puettmann KJ (2014) Multiscale controls on natural regeneration dynamics after partial overstory removal in Douglas-fir Forests in western Oregon, USA. For Sci 60:953–961
Donato DC, Campbell JL, Franklin JF (2012) Multiple successional pathways and precocity in forest development: can some forests be born complex? J Veg Sci 23:576–584
Fox GA, Negrete-Yankelevich S, Sosa VJ (eds) (2015) Ecological statistics: contemporary theory and application. Oxford University Press, Oxford
Franklin JF, Johnson KN (2012) A restoration framework for federal forests in the Pacific Northwest. J For 110:429–439
Franklin JF, Van Pelt R (2004) Spatial aspects of structural complexity in old-growth forests. J For 102:22–28
Franklin JF, Johnson KN, Johnson DL (2018) Ecological forest management. Waveland Press, Illinois
Gavin DG, Hu FS (2006) Spatial variation of climatic and non-climatic controls on species distribution: the range limit of Tsuga heterophylla. J Biogeogr 33:1384–1396
Goodburn JM, Lorimer CG (1998) Cavity trees and coarse woody debris in old-growth and managed northern hardwood forests in Wisconsin and Michigan. Can J For Res 28:427–438
Gray AN, Spies TA (1996) Gap size, within-gap position and canopy structure effects on conifer seedling establishment. J Ecol 84:635–645
Gray AN, Spies TA (1997) Microsite controls on tree seedling establishment in conifer forest canopy gaps. Ecology 78:2458–2473
Gray AN, Spies TA, Easter MJ (2002) Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests. Can J For Res 32:332–343
Gray AN, Spies TA, Pabst RJ (2012) Canopy gaps affect long-term patterns of tree growth and mortality in mature and old-growth forests in the Pacific Northwest. For Ecol Manag 281:111–120
Harmon ME, Franklin JF (1989) Tree seedlings on logs in Picea-Tsuga forests of Oregon and Washington. Ecology 70:48–59
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW (1986) Ecology of coarse woody debris in temperate ecosystems. Adv Ecol Res 15:133–302
Harrington TB (2006) Five-year growth responses of Douglas-fir, western hemlock, and western redcedar seedlings to manipulated levels of overstory and understory competition. Can J For Res 36:2439–2453
Harrington CA, Roberts SD, Brodie LC (2005) Tree and understory responses to variable-density thinning in western Washington. Gen. Tech. Rep. PNW-GTR-635. Portland, OR: US Department of Agriculture, Forest Service, Pacific Northwest Research Station, pp 97–106
Harris AS (1990) Picea sitchensis (Bong.) Carr. Sitka spruce. In: Burns RM, Honkala BH (eds) Silvics of North America, vol. 1: Conifers. US Department of Agriculture Forest Service Agriculture Handbook No 654, pp 260–267
Kern CC, Burton JI, Raymond P, D’Amato AW, Keeton WS, Royo AA, Walters MB, Webster CR, Willis JL (2017) Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. For Int J For Res 90:4–17
King JE (1966) Site index curves for Douglas-fir in the Pacific Northwest. Weyerhaeuser Company. Weyerhaeuser Forestry Research Center, Centralia, WA. Weyerhaeuser. For 8:1–49
Kuehne C, Puettmann KJ (2008) Natural regeneration in thinned Douglas-fir stands in western Oregon. J Sustain For 27:246–274
LePage PT, Canham CD, Coates KD, Bartemucci P (2000) Seed abundance versus substrate limitation of seedling recruitment in northern temperate forests of British Columbia. Can J For Res 30:415–427
Maguire DA, Mainwaring DB, Rose R, Garber SM, Dinger EJ (2009) Response of coastal Douglas-fir and competing vegetation to repeated and delayed weed control treatments during early plantation development. Can J For Res 39:1208–1219
Mason WL, Edwards C, Hale SE (2004) Survival and early seedling growth of conifers with different shade tolerance in a Sitka spruce spacing trial and relationship to understorey light climate. Silva Fenn 38:357–370
McGee GG, Leopold DJ, Nyland RD (1999) Structural characteristics of old-growth, maturing, and partially cut northern hardwood forests. Ecol Appl 9:1316–1329
Miller M, Emmingham B (2001) Can selection thinning convert even-age Douglas-fir stands to uneven-age structures? West J Appl For 16:35–43
Minore D (1979) Comparative autecological characteristics of northwestern tree species—a literature review. Gen. Tech. Rep. PNW-GTR-087. Portland, OR: US Department of Agriculture, Forest Service, Pacific Northwest Research Station: 1–72, 87
Mitchell AK, Dunsworth BG, Arnott JT, Koppenaal R, Benton R, Goodmanson G, Brown T, Sandford J (2004) Growth limitations of planted conifers regenerating under montane alternative silviculture systems (MASS): seven-year results. For Chron 80:241–250
Nabel MR, Newton M, Cole EC (2013) Abundance of natural regeneration and growth comparisons with planted seedlings 10–13 years after commercial thinning in 50-year-old Douglas-fir, Douglas-fir/western hemlock, Oregon Coast Range. For Ecol Manag 292:96–110
Neuendorff JK, Nagel LM, Webster CR, Janowiak MK (2007) Stand structure and composition in a northern hardwood forest after 40 years of single-tree selection. North J Appl For 24:197–202
Newton M, Cole EC (2006) Harvesting impacts on understory regeneration in two-storied Douglas-fir stands. West J Appl For 21:14–18
Nyland RD (2016) Silviculture: concepts and applications. Waveland Press, Illinois
Pacala SW, Canham CD, Silander JA Jr, Kobe RK (1994) Sapling growth as a function of resources in a north temperate forest. Can J For Res 24:2172–2183
Packee EC (1990) Tsuga heterophylla (Raf.) Sarg., Western hemlock. In: Burns RM, Honkala BH (eds) Silvics of North America, vol. 1: Conifers. US Department of Agriculture Forest Service Agriculture Handbook No. 654, pp 613–622
Page LM, Cameron AD (2006) Regeneration dynamics of Sitka spruce in artificially created forest gaps. For Ecol Manag 22:260–266
Puettmann K, Ares A, Burton J, Dodson E (2016) Forest restoration using variable density thinning: lessons from Douglas-fir stands in Western Oregon. Forests 7:310
Quine CP (2001) A preliminary survey of regeneration of Sitka spruce in wind-formed gaps in British planted forests. For Ecol Manag 151:37–42
Roberts SD, Harrington CA (2008) Individual tree growth response to variable-density thinning in coastal Pacific Northwest forests. For Ecol Manag 255:2771–2781
Roberts SD, Harrington CA, Terry TA (2005) Harvest residue and competing vegetation affect soil moisture, soil temperature, N availability, and Douglas-fir seedling growth. For Ecol Manag 205:333–350
Royo AA, Carson WP (2006) On the formation of dense understory layers in forests worldwide: consequences and implications for forest dynamics, biodiversity, and succession. Can J For Res 36:1345–1362
Seidel D, Ruzicka KJ, Puettmann KJ (2016) Canopy gaps affect the shape of Douglas-fir crowns in the western Cascades, Oregon. For Ecol Manag 363:31–38
Shatford JP, Bailey JD, Tappeiner JC (2009) Understory tree development with repeated stand density treatments in coastal Douglas-fir forests of Oregon. West J Appl For 24:11–16
Spies TA (2004) Ecological concepts and diversity of old-growth forests. J For 102:14–20
Spies TA, Franklin JF (1991) The structure of natural young, mature, and old-growth Douglas-fir forests in Oregon and Washington. In: Ruggiero LF, Aubry B, Carey AB, Huff MH, et al. (eds) Wildlife and vegetation of unmanaged Douglas-fir forests. USDA Forest Service General Technical Report PNW-GTR-285, pp 91–109
Spies TA, Franklin JF, Thomas TB (1988) Coarse woody debris in Douglas-fir forests of western Oregon and Washington. Ecology 69:1689–1702
Stone JN, MacKinnon A, Parminter JV, Lertzman KP (1998) Coarse woody debris decomposition documented over 65 years on southern Vancouver Island. Can J For Res 28:788–793
Sullivan TP, Sullivan DS (2016) Acceleration of old-growth structural attributes in lodgepole pine forest: tree growth and stand structure 25 years after thinning. For Ecol Manag 365:96–106
Taylor AH (1990) Disturbance and persistence of Sitka spruce (Picea sitchensis (Bong) Carr.) in coastal forests of the Pacific Northwest, North America. J Biogeogr 17:47–58
Tesch SD, Lysne DH, Mann JW, Helgerson OT (1986) Mortality of regeneration during skyline logging of a shelterwood overstory. J For 6:49–50
Thomas JW, Franklin JF, Gordon J, Johnson KN (2006) The Northwest Forest Plan: origins, components, implementation experience, and suggestions for change. Conserv Biol 20:277–287
Thornburgh DA (1969) Dynamics of the true fir-hemlock forests of the west slope of the Washington Cascade range. Dissertation. University of Washington, Seattle, Washington, USA
Tyrrell LE, Crow TR (1994) Structural characteristics of old-growth hemlock-hardwood forests in relation to age. Ecology 75:370–386
Walters MB, Lajzerowicz CC, Coates KD (2006) Soil resources and the growth and nutrition of tree seedlings near harvest gap–forest edges in interior cedar-hemlock forests of British Columbia. Can J For Res 36:62–76
Webster CR, Dickinson YL, Burton JI, Frelich LE, Jenkins MA, Kern C, Raymond P, Saunders MR, Walters MB, Willis JL (2018) Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For Ecol Manag 421:98–108
Wiley KN (1978) Site index tables for western hemlock in the Pacific Northwest. Weyerhaeuser Forestry Paper 17
Willis JL, Roberts SD, Harrington CA (2018) Variable density thinning promotes variable structural responses 14 years after treatment in the Pacific Northwest. For Ecol Manag 410:114–125
Wright EF, Coates KD, Bartemucci P (1998) Regeneration from seed of six tree species in the interior cedar-hemlock forests of British Columbia as affected by substrate and canopy gap position. Can J For Res 28:1352–1364
Youngblood A (2000) Damage to residual trees and advance regeneration from skyline and forwarder yarding in mixed-conifer stands of northeastern Oregon. West J Appl For 15:101–107
Acknowledgements
We thank the USDA Forest Service Olympic National Forest for implementing the operational treatments, the USDA Forest Service Pacific Northwest Region and Pacific Northwest Research Station for funding, and our many coworkers for their efforts in plot installation and measurement. We would also like to thank Nancy Koerth for her statistical advice.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Willis, J.L., Harrington, C.A., Brodie, L.C. et al. Variable-density thinning promotes differential recruitment and development of shade tolerant conifer species after 17 years. New Forests 52, 329–348 (2021). https://doi.org/10.1007/s11056-020-09797-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-020-09797-z