
Abstract
Background
Tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) is an apoptosis inducer that exhibits an ideal therapeutic safety profile with less adverse effects than conventional chemotherapy. However, the occurrence of TRAIL resistance has been reported in various cancers including colorectal cancer (CRC). Substantial efforts have been channelled towards managing TRAIL resistance including identifying molecular targets. Interleukins (ILs) have been recently shown to play critical roles in modulating TRAIL sensitivity in cancer cells.
Methods and Results
This study investigated the roles of two ILs, IL-8 and IL⍺, in TRAIL resistance in CRC. TRAIL-resistant HT-29 and TRAIL-sensitive HCT 116 cells, were treated with human recombinant IL-8 and IL-1⍺. The results indicated that treatment with IL-8 (2.5 ng/mL) significantly protected TRAIL-sensitive HCT 116 cells from TRAIL-induced cell death (p < 0.05). However, IL-1⍺ did not play a role in modulating CRC cells’ responses to TRAIL. Data from RT-qPCR and Western blotting revealed the molecular regulations of IL-8 on TRAIL decoy receptor genes (OPG) and autophagy-related genes (BECN1 and LC3B) expression. The activation of the phosphoinositide 3-kinase (PI3K) pathway was shown to counteract TRAIL-induced cell death. By inhibiting its activation with wortmannin, the protective role of IL-8 against TRAIL treatment was reversed, suggesting the involvement of the PI3K pathway.
Conclusion
Collectively, findings from this study identified the role of IL-8 and PI3K in modulating CRC cells’ sensitivity to TRAIL. Further validation of these two potential molecular targets is warranted to overcome TRAIL resistance in CRC.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
According to the latest report from the Global Cancer Observatory (GLOBOCAN) in 2022, colorectal cancer (CRC) has become the second leading cause of mortality worldwide. Over 1.9 million new cases of CRC are expected every year with a projected number of at least 904,000 deaths [1]. Early detection of the disease has always been the priority for a good five-year survival prognosis in CRC patients. However, despite the emergence of new screening and diagnostic tools, 61% of cases were diagnosed at an advanced stage with distant metastases. Surgical resection along with adjuvant therapy remains the mainstay treatment for patients with advanced CRC [2]. When tumour lesions become metastatic, optimal treatment shifts from curative surgical control to systemic treatment [3].
Chemotherapy has been around as the mainstay therapy in managing metastatic CRC. However, the adverse effects of CRC chemotherapy systemic toxicity and low tumour-specific selectivity have led to the development of targeted therapies [4, 5]. Tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL), a potent inducer of apoptosis, has been identified to play an important role in immune surveillance and the elimination of cancer cells [6, 7]. Moreover, IL demonstrates a high specificity to cancer cells [6, 8]. This superior safety profile prompts the development of TRAIL-based therapies for cancer clinical applications. Promising results were obtained in both preclinical studies and early-phase clinical trials [9], However, these TRAIL-based therapies have yielded unsatisfactory outcomes in the subsequent randomised clinical trials [10]. TRAIL resistance has become one of the significant challenges in TRAIL-based therapy among a vast proportion of human cancers, including bladder, prostate, and colorectal cancer [11,12,13]. Understanding the mechanisms underlying TRAIL resistance in cancer is, therefore, crucial to overcoming this hurdle and improving the outcomes of TRAIL-based treatment.
Recently, the overexpression of interleukins has been found to promote TRAIL resistance in breast and pancreatic cancer [14, 15]. To date, in CRC, only interleukin (IL)-4 and − 6 have been recognised to play crucial roles in drug resistance [16,17,18]. Hence, IL-8 and IL-1 alpha (IL-1⍺) are the subjects of interest in this study. These two interleukins are vital immune system components and have been shown to regulate CRC cells’ drug sensitivity [19, 20]. IL-8 was first identified as a chemoattractant, responsible for neutrophil infiltration in acute inflammation [19]. On the other hand, IL-1⍺, a member of damage-associated molecular patterns (DAMPs), is involved in the initiation of inflammation and restoration of cellular homeostasis [21]. Interestingly, apart from their physiological roles, the elevated levels of IL-8 and IL-1⍺ found in several cancers, especially CRC, underscoring their potential correlation with the pathogenesis of this disease [22].
IL-8 activates several downstream signalling cascades, including the pro-survival phosphoinositide 3-kinase (PI3K) is crucial for cancer metastases, and drug resistance [19, 23, 24]. In the study by Wilson et al., IL-8 attenuated TRAIL-induced cell death in prostate cancer cells via upregulating the expression of cellular FLICE (FADD-like IL-1β-converting enzyme)-inhibitory protein (c-FLIP) [15]. However, the role of IL-8 in modulating CRC cells’ sensitivity to TRAIL treatment is yet to be investigated. Apart from investigating the anti-apoptotic protein expression, how the TRAIL decoy receptors (DcR); DcR1, DcR2 and osteoprotegerin (OPG) influence TRAIL sensitivity should also be explored in CRC cells. This is because the three decoy receptors are known to inhibit TRAIL-induced cell death and studies have found that the decoy receptor levels are elevated in both serum and tumour samples of CRC patients [25,26,27,28,29].
Similar to IL-8, the understanding of another IL family member, IL-1⍺ and its role in modulating TRAIL sensitivity in CRC is limited. Recently, a study by Pimentel et al., utilising breast cancer cells demonstrated that the downregulation of IL-1⍺ via siRNA technique restored TRAIL sensitivity, as indicated by the increase in colony formation and cell growth [14]. In CRC cells, Cheng et al. showed that stimulation with IL-1⍺ significantly upregulates the expression of the TRAIL decoy receptor, OPG [20]. However, it remains unclear whether the IL-1⍺-induced OPG overexpression influences CRC cells’ TRAIL sensitivity.
Thus, this study aimed to investigate the role of IL-8 and IL-1⍺ in modulating CRC cell’s sensitivity to TRAIL treatment. Apart from identifying the regulators of TRAIL sensitivity, this study also focuses on elucidating potential mechanisms that could modulate TRAIL sensitivity in CRC cells. Therefore, this study hypothesises that these ILs modulate TRAIL sensitivity in CRC cells via the activation of the PI3K pathway. The findings of this study showed that the CRC cells’ responses to TRAIL treatment are dependent on IL-8 and PI3K pathways while independent of IL-1⍺. Further exploration of these two molecular targets is warranted to better understand their roles and potentially enhance the efficacy of TRAIL-based therapies in CRC.
Materials and methods
Cell culture
Human CRC cell lines HT-29 and HCT 116 were obtained from the American Type Culture Collection (ATCC, USA). The HT-29 and HCT 116 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) medium (Corning, USA) supplemented with glucose (4.5 g/L), l-glutamine (584 mg/mL) and sodium pyruvate (110 mg/mL) containing 10% foetal bovine serum (FBS), penicillin (100 U/mL) and streptomycin (100 µg/mL) in Cells were incubated in a humidified incubator at 37 °C with 5% of CO2.
Cell treatment
The IC50 of TRAIL was determined to evaluate the TRAIL sensitivity in both HT-29 and HCT 116 cells. Briefly, cells were seeded in 96-well plates and treated with recombinant human TRAIL (PeproTech, USA, 310-04) in a concentration-dependent manner (3.81–4000 ng/mL) for 24 h. To investigate the role of IL-1⍺ and IL-8 in modulating the sensitivity towards TRAIL treatment in both HT-29 and HCT 116, cells were pre-treated with the recombinant human interleukin IL-1⍺ (Sigma, USA, SRP6295) or IL-8 (Sigma, USA, 208-IL-010) before treating the cells with TRAIL (44.2 ng/mL). To study the role of PI3K in modulating TRAIL sensitivity, cells were pre-incubated with IL-8 (2.5 ng/mL) in the presence of PI3K inhibitors (wortmannin 10 µM, Abcam, USA) or the vehicle control (0.04% ethanol, Fisher Scientific, USA) for 72 h followed by TRAIL treatment.
3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide (MTT) assay
MTT assay was utilised to determine the viability of the cells. At the end of the cell treatment time point, the cell culture media was removed, and 100 µL of fresh media consisting of 1 mg/mL of MTT powder was added into each well. After 25–30 min of incubation time at 37 °C, the media was withdrawn. A hundred µL of dimethyl sulfoxide (DMSO) was added to each well, followed by gentle shaking until purple formazan crystals were fully dissolved. The absorbance indicating viable cells was measured at 570 nm using a multi-mode microplate reader (Spectramax).
RT-qPCR
Cells were treated with recombinant human IL-1⍺ (100 ng/mL) and IL-8 (2.5 ng/mL) for 24 and 72 h. A two-step RT-qPCR assay was utilised to assess the gene expressions of TRAIL receptors (DR4, DR5, DcR1, DcR2 and OPG), apoptosis gene (caspase 3, 8, 9, cFLIP and cytochrome c) and autophagy-related genes (beclin-1, LC3B, p62 and GABARAP). Total RNA extraction was performed using Monarch® Total RNA MiniPrep Kit (New England Biolabs, USA, T2010) according to the manufacturer’s guidelines. A Nanodrop Spectrophotometer (Thermo Scientific) was used to quantify and determine the purities of the extracted RNA samples. RNA samples were then reversed transcribed to complementary DNA (cDNA) using LunaScript RT SuperMix Kit (New England Biolabs, USA, E3010) and 96-well PCR Thermal Cycler (Veriti®, Applied Biosystems) according to the manufacturer’s instructions. The conditions were as follows: 25 ̊C for 2 min, 55 ̊C for 10 min and 95 ̊C for 1 min. Following that, qPCR was performed with the cDNA generated using Luna® Universal qPCR Master Mix (New England Biolabs, USA, M3003) and Real-Time PCR machine. The thermocycling conditions were as follows: initial denaturation at 95 ̊C for 1 min, followed by each step of denaturation at 95 ̊C for 15 s and annealing extension at 55 ̊C for 30 s, for 40 cycles. The glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was chosen to be the internal reference gene for the conversion of CT value to relative expression with the 2−ΔΔCt data analysis method. Details of the primer sequences are listed in Supplementary Table 1.
Western blotting
Western blotting was used to investigate the phosphorylation of the PI3K pathway in HCT 116 cells. Briefly, upon IL-8 exposure, the cells were lysed by adding 100 µL of RIPA Lysis Buffer System (Santa Cruz Biotechnology Inc, USA, sc-24948 A) in the presence of 1% phenylmethylsulphonyl fluoride (PMSF), 1% protease inhibitor cocktail and 1% sodium orthovanadate. Subsequently, cell lysates were collected and centrifuged at 10,000 g with a refrigerated centrifuge (TOMY, CAX 371) for 10 min to remove cell debris. Protein concentration was quantified using the Nanodrop Spectrophotometer (Thermo Scientific 2000). Sixty µg of protein was denatured in SDS-PAGE sample loading buffer, Laemmli 2x Concentrate (Sigma, USA, S3401) at 100 °C for 10 min and loaded into 10% TGX FastCast Acrylamide Gels (BioRad Laboratories, USA, 1610173). Proteins were separated at 60 V for 30 min and 175 for 45 min (BioRad Laboratories, USA, 1658033) in 1x SDS Running Buffer and transferred to a 0.2 μm polyvinylidene difluoride (PVDF) membrane at 115 V for 50 min in ice-cold 1x Transfer Buffer. The membrane was blocked overnight at 4 °C in 5% bovine serum albumin (BSA) (Capricorn Scientific, USA, BSA-1 S) to prevent non-specific binding. Subsequently, the membrane was incubated overnight at 4 °C with primary antibodies; phosphorylated and total PI3K, AKT and protein S6 diluted in 5% BSA. Details of antibodies and dilutions are listed in Supplementary Table 2. Next, the membrane was washed with 0.1% PBS/ Tween-20. The membrane was incubated with HRP-conjugated secondary antibodies (1:5000) at room temperature for 1.5 h. The bands of the proteins were detected using enhanced chemiluminescence (ECL) (BioRad Laboratories, USA) and visualised using the FluoChemR Imaging System. To ensure equal loading of protein samples, the membranes were re-probed with housekeeping antibody (b-actin). Densitometric analysis of protein bands was performed using ImageJ software (version 1.53).
Statistical analysis
Mean data were presented as absolute values ± standard error mean (SEM). Statistical analysis was performed using one-way with Dunnett’s post hoc test when comparing stimulants to the control group. One/two-way ANOVA with Bonferroni’s correction post-hoc test was used when there were intragroup comparisons (i.e. stimulation in the presence/absence of inhibitors).
Results
The IC50 of TRAIL in HT-29 and HCT 116 CRC cells
The HT-29 and HCT 116 CRC cells exhibited contrasting responses to TRAIL treatment. To assess their sensitivity to TRAIL, a cell viability assay using MTT was conducted. The cells were treated with increasing TRAIL concentrations (3.81–4000 ng/mL) to ascertain the half-maximal inhibitory concentration (IC50). This highest concentration was selected based on the findings by Toscano et al. [30]. The viability of HT-29 cells was not affected by the increasing concentration of TRAIL (Fig. 1a). In contrast, the viability of HCT 116 cells decreased in a concentration-dependent manner (Fig. 1b). The IC50 of TRAIL in HCT 116 cells was determined to be 44.2 ng/mL, while this could not be determined in HT-29 as the cells were irresponsive to TRAIL. To study the effect(s) of interleukins in modulating TRAIL sensitivity, the TRAIL-sensitive HCT 116 cells were utilised for the subsequent experiments.

HT-29 and HCT 116 cells exhibit differential responses upon TRAIL treatment. Cells were seeded in 96-well plates and treated with TRAIL in a concentration-dependent manner for 24 h. The graph above represents cell viability % against the log concentration of TRAIL treatment (3.91–4000 ng/mL). MTT assay was utilised to determine the cell viability in (a) HT-29 and (b) HCT 116 cells. The results were presented as a percentage of cell viability relative to control. Each data point represented the mean ± SEM of three independent experiments, the biological triplicates (n = 3). ND = Not determined.
The role of IL-8 and IL-1⍺ in modulating the expression of TRAIL receptors
The potential role of these interleukins in regulating the expression of TRAIL receptors was then investigated in the TRAIL-sensitive cell line, HCT 116. The results indicated that IL-8 significantly enhanced the expression of OPG approximately 2.5-fold (Fig. 2a). In comparison, the expressions of death receptors (DR4 and DR5) and the two other decoy receptors (DcR1 and DcR2) remained unchanged with IL-8. In a study conducted by Granci et al., the importance of the ratio of death receptors: decoy receptors was reported. A higher risk of disease progression was found in patients who concomitantly expressed low/medium levels of DR4 and high levels of DcR1 [31]. In this study, when the ratio of IL-8-induced OPG against both death receptors (OPG: DR4 and OPG: DR5) was assessed, there was an increase of more than two-fold. These results suggest that the presence of IL-8 may influence TRAIL sensitivity by modulating the expression of TRAIL decoy receptor OPG. In contrast, IL-1⍺ did not affect the expression of TRAIL death and decoy receptors (Fig. 2b).

The mRNA relative expression of TRAIL receptors in HCT 116 cells. Cells were treated with (a) IL-8 (2.5 ng/mL) for 72 h and (b) IL-1⍺ (100 ng/mL) for 24 h, followed by quantitative RT-qPCR to determine the relative mRNA expression levels of the TRAIL death (DR4 and DR5) and decoy receptors (DcR1, DcR2 and OPG). Bars represent mean data (± SEM). One-way ANOVA with Dunnett’s correction was used to determine statistical differences. A p-value ≤ 0.05 is considered statistically significant
The role of IL-8 and IL-1⍺ in apoptosis induction
TRAIL-mediated cell death occurs via the induction of apoptosis. To examine whether IL-8 and IL-1⍺ influence TRAIL sensitivity by regulating apoptosis, the expression levels of apoptosis effectors (caspase-8, -3 and − 9) and inhibitors (cytochrome c and cFLIP) were assessed upon treatment with IL-8 and IL-1⍺. Both IL-8 and IL-1⍺ treatment did not result in the expression change of the apoptosis effectors and inhibitor-related genes (Fig. 3a and b).

The mRNA expression of apoptosis regulators in HCT 116 cells. HCT 116 cells were treated with (a) IL-8 (2.5 ng/mL) for 72 h and (b) IL-1⍺ (100 ng/mL) for 24 h, followed by quantitative RT-qPCR to determine the relative mRNA expression of apoptosis effectors and inhibitors. Bars represent mean data (± SEM). One-way ANOVA with Dunnett’s correction was used to determine statistical differences. A p-value ≤ 0.05 is considered statistically significant
The role of IL-8 and IL-1⍺ in autophagy activation
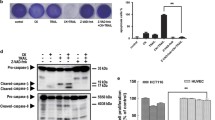
Autophagy and apoptosis are closely related mechanisms in which their regulators’ crosstalk influences TRAIL sensitivity. Hence, the expression levels of autophagy-related genes (beclin-1, LC3B, p62 and GABARAP) were investigated in HCT 116. Stimulation with IL-8 resulted in significant downregulation of BECN (Fig. 4a) and upregulation of the LC3B (Fig. 4b). IL-1⍺ did not regulate the mRNA expressions of autophagy-related genes in HCT 116 cells. This finding was validated by western blot analysis of the LC3B and p62 protein (Fig. 4c).

The impact of IL-8 and IL-1⍺ on autophagy-related genes and proteins. HCT 116 cells were treated with (a) IL-8 (2.5 ng/mL) for 72 h and (b) IL-1⍺ (100 ng/mL) for 24 h followed by quantitative RT-qPCR to determine the relative mRNA expression of autophagy-related genes. (c) cells were treated with IL-1⍺ (100 ng/mL) followed by Western blotting to determine the relative protein expression of autophagy-related proteins. Bars represent mean data (± SEM). One-way ANOVA with Dunnett’s correction was used to determine statistical differences. A p-value ≤ 0.05 is considered statistically significant
The role of IL-8 and IL-1⍺ in modulating HCT 116 cells’ TRAIL sensitivity
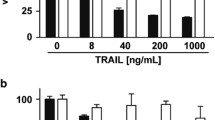
Cell viability assessment was conducted to elucidate the functional consequences of IL-8 and IL-1⍺ on TRAIL sensitivity in HCT 116 cells. The cells were treated with TRAIL in the presence and absence of IL-8 (Fig. 5a) or IL-1⍺ (Fig. 5b) pre-stimulation. Cell viability was unaffected by IL-1⍺ pre-stimulation, whereas IL-8 pre-stimulation resulted in an increased in cell viability. This suggests that IL-8 but not IL-1⍺ may have a direct role in modulating the sensitivity of HCT 116 cells to TRAIL treatment.

The effect of IL-8 and IL-1⍺ on the cell viability of HCT 116 cells. Cells were pre-treated with (a) IL-8 (2.5 ng/mL) for 72 h and (b) IL-1⍺ (100 ng/mL) for 24 h followed by TRAIL treatment for 24 h. MTT assay was utilised to determine the cell viability. The results were presented as a percentage of cell viability relative to untreated control and TRAIL alone treatment. Statistical analysis was performed using one-way ANOVA with Dunnett’s correction ± SEM (N at least 3). A p-value ≤ 0.05 is considered statistically significant
The role of PI3K pathway activation in modulating TRAIL sensitivity
In cancer, IL-8-induced phosphorylation of ribosomal protein S6, a downstream target of the PI3K pathway has been implicated in the resistance to targeted therapies including the TRAIL therapy [32]. Hence, Western blot analysis was conducted to explore the effect of IL-8 on the phosphorylation of protein S6. Time course experiments showed that exposure to IL-8 did not consistently induce phosphorylation of protein S6 (Supplementary Fig. 1). In addition, the phosphorylation of upstream proteins in the PI3K pathway (PI3K and AKT) was also examined. Despite the total proteins were detectable and not regulated, their phosphorylated proteins were undetectable (Supplementary Fig. 2). To confirm the role of PI3K, further investigation using pharmacological inhibitors should be conducted in response to TRAIL treatment. Wortmannin, an inhibitor of PI3K was used to elucidate the role of PI3K in the functional consequences of IL-8 on TRAIL sensitivity. Cells were pre-incubated with IL-8 in the presence or absence of wortmannin. Subsequently, cells were exposed to TRAIL treatment and an MTT assay was conducted. Consistent with the previous findings, IL-8 significantly increased the cell viability to TRAIL treatment reducing TRAIL sensitivity (Fig. 6). In addition, the presence of wortmannin significantly reversed the increase in cell viability induced by IL-8. Contradicting with the previous western blot data, this MTT data sheds light on the potential involvement of the PI3K pathway in modulating TRAIL sensitivity in CRC.

The modulatory role of IL-8 in CRC cells is potentially dependent on the activation of the PI3K pathway. Cells were treated with TRAIL in the presence or absence of IL-8 (2.5 ng/mL) and PI3K inhibitor (wortmannin). Vehicle control was to ensure results observed were due to the inhibition of PI3K and not ethanol used for reconstitution. The results were presented as a percentage of cell viability relative to the untreated control. Bars represent mean data (± SEM). One-way ANOVA with Bonferroni’s correction was used to determine statistical differences. A p-value ≤ 0.05 is considered statistically significant. * = p < 0.05 ; **= p < 0.01
Discussion
In this study, the response of CRC cells to TRAIL and the role of IL-8 and IL-1⍺ in modulating TRAIL sensitivity were investigated. The results showed that HCT 116 cells were susceptible to TRAIL-induced cell death, whereas HT-29 cells were resistant. Distinct regulatory roles of interleukins were observed. IL-8 protected TRAIL-sensitive HCT 116 cells from TRAIL-induced cell death while IL-1⍺ did not have any modulatory effect. Additionally, IL-8 regulated the gene expression of TRAIL decoy receptor (OPG) and autophagy-related markers; beclin-1 and LC3B in HCT 116 cells, while IL-1⍺ had no effect. The role of the PI3K pathway in modulating TRAIL sensitivity was investigated by using a pharmacological PI3K inhibitor, wortmannin. In the presence of wortmannin, the protection offered by IL-8 from TRAIL-induced cell death was reversed suggesting that IL-8 mediates its effect by a PI3K-dependent pathway.
The differences in cellular viability pattern between the HT-29 and HCT 116 with a gradually increasing concentration of TRAIL demonstrate the distinctive sensitivities against TRAIL treatment. Several studies have also obtained similar results to the findings observed in this study [33,34,35]. Recently, this distinctive responsiveness to TRAIL was shown to be associated with the microsatellite stability of these cells [33]. In the study by Fohlen et al., HCT 116 was categorised as a cell line with microsatellite instability (MSI) whereas HT-29 was microsatellite stable (MSS) [36]. In a clinical study involving CRC patients with known MSI/MSS status, it was reported that patients with MSI status have better prognosis when compared to patients with MSS status [34]. However, in CRC cell lines (HCT 116 and HT-29) the association between the MSI status and TRAIL sensitivity is yet to be established. Comprehensive analysis of the gene expression profiles between MSI and MSS CRC cell lines could provide further insights into the mechanisms underlying differential response observed to TRAIL therapy.
In addition, the mutation in the tumour suppressor gene (TP53) is also one of the factors that can influence TRAIL-induced cell death in CRC [30, 35]. Willms et al. Showed that cells with wild-type (WT) p53 were reported to be more sensitive to TRAIL treatment than p53-mutated cells [35]. Furthermore, p53 also regulates the expression of TRAIL decoy receptors impacting TRAIL sensitivity [30]. A previous study documented the status of p53 as WT in HCT 116 cells while mutated in the HT-29 cells [36]. This current study demonstrated that HCT 116 (WT p53) are susceptible to TRAIL, while HT-29 (mutated p53) is resistant, supporting the notion that p53 status influences TRAIL sensitivity between CRC cell lines. Detail investigations into the contrasting characteristics of these two CRC cell lines will help identify potential predictive markers for selecting patients who would benefit most from TRAIL-based therapy.
The significance of IL-8 in modulating TRAIL sensitivity via the PI3K pathway was demonstrated in this study. In the present study, the inhibition of PI3K with wortmannin sensitised the response of IL-8-treated HCT 116 cells to TRAIL, indicating the importance of the PI3K pathway in TRAIL resistance. This finding is in line with a previous study where the PI3K pathway activation has been shown to counteract TRAIL-induced apoptosis in pancreatic cancer [15]. The role of PI3K in modulating drug sensitivity has been implicated in various studies where the inhibition of the PI3K pathway successfully re-sensitised cancer cells to the anti-cancer therapy [24, 37]. This suggests that targeting the PI3K pathway could be a viable strategy to overcome drug resistance including TRAIL resistance and enhance the effectiveness of anti-cancer treatment.
IL-8 modulated the expression of TRAIL decoy receptors and autophagy markers such as beclin-1 and LC3B but did not impact significantly apoptosis. This suggests that IL-8 may enhance TRAIL resistance through regulations of autophagy activation and decoy receptor overexpression, shedding light on its mechanism of action. The interplay between autophagy and TRAIL-induced apoptosis has been documented in the tumorigenesis and progression of CRC [38]. Autophagy activation in colon cancer cells offers protection against TRAIL-induced apoptosis [39]. Although it is tempting to speculate that IL-8-mediated TRAIL-resistance occurs via autophagy activation, however, in this current study IL-8 only upregulates LC3 while downregulates BECN1. As both LC3B and beclin-1 are important regulators of autophagy and were hypothesised to be upregulated by IL-8, further analysis on other markers of autophagy using other techniques such as immunofluorescence with live microscopy is required before confirming the mechanism of IL-8.
IL-1⍺ did not alter TRAIL-induced cell death in CRC cells, consistent with its molecular analysis where expression of TRAIL receptor genes, autophagy-related genes and apoptosis genes were not regulated. It is worth noting that a previous study conducted by Cheng et al. documented that IL-1⍺ upregulated the protein expression TRAIL decoy receptor OPG in HT-29 cells [20]. However, the association between OPG and TRAIL sensitivity was not explored in this study.
Conclusion
Findings from this study may unlock potential clues in combating the development of TRAIL resistance in CRC. While this study focuses on in vitro analysis, however, further validation in an in vivo model of CRC and clinical trials will be important to translate these findings into effective treatments for patients. An in vivo model of CRC would demonstrate a more complex tumour interaction and network involving IL-8, PI3K and TRAIL sensitivity. Additionally, in the clinical context, several antibodies and drugs against IL-8 have been developed and brought to clinical trials for efficacy testing. AMY109, a long-acting recycling anti-IL-8 antibody has demonstrated clinical potency by lowering the level of IL-8 [40]. Phase I clinical trials of HuMax-IL8 also show its ability to make tumour cells less resistant to treatment [41]. Alpelisib is a PI3K inhibitor that has been approved by the Food and Drug Administration (FDA) for the treatment of metastatic breast cancer [42]. However, none of the drugs have been used to tackle TRAIL resistance in CRC. Therefore, research on additional potential targets implicated in IL-8-induced TRAIL resistance should be highly promoted. This effort is crucial for maximizing the effectiveness of TRAIL-based treatments in CRC.
The clinical significance of this study lies in its potential to improve treatment outcomes for patients with CRC by elucidating the roles of IL-8 and the PI3K pathway in mediating susceptibility to TRAIL-induced cell death. The current research highlights novel targets for therapeutic intervention. Understanding these molecular mechanisms allows the development of strategies to enhance the effectiveness of TRAIL-based therapies, and bring TRAIL to the clinic. Furthermore, the study suggests that biomarkers like IL-8 and components of the PI3K pathway could be used to identify patients who are more likely to respond to TRAIL therapy, leading to more personalized and effective treatment plans. Ultimately, these insights could contribute to reducing CRC mortality and enhancing quality of life through individualising therapeutic approaches.
Data availability
No datasets were generated or analysed during the current study.
References
Bray F et al (2024) Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a Cancer Journal for clinicians. 74(3):229–263. https://doi.org/10.3322/caac.21834
Brown KGM et al (2019) Management of colorectal cancer. BMJ 366:l4561. https://doi.org/10.1136/bmj.l4561
Van Cutsem E et al (2014) Metastatic colorectal cancer: ESMO Clinical Practice guidelines for diagnosis, treatment and follow-up. Ann Oncol iii1–9 25 Suppl 3. https://doi.org/10.1093/annonc/mdu260
Hammond WA, Swaika A, Mody K (2016) Pharmacologic resistance in colorectal cancer: a review. Ther Adv Med Oncol 8(1):57–84. https://doi.org/10.1177/1758834015614530
Xie YH, Chen YX, Fang JY (2020) Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct Target Ther 5(1):22. https://doi.org/10.1038/s41392-020-0116-z
Kretz AL et al (2019) TRAILblazing strategies for Cancer Treatment. Cancers (Basel) 11(4). https://doi.org/10.3390/cancers11040456
Ndebele K et al (2008) Tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) induced mitochondrial pathway to apoptosis and caspase activation is potentiated by phospholipid scramblase-3. Apoptosis 13(7):845–856. https://doi.org/10.1007/s10495-008-0219-4
French LE, Tschopp J (1999) The TRAIL to selective tumor death. Nat Med 5(2):146–147. https://doi.org/10.1038/5505
Snajdauf M et al (2021) The TRAIL in the treatment of Human Cancer: an update on clinical trials. Front Mol Biosci 8:628332. https://doi.org/10.3389/fmolb.2021.628332
Ouyang X et al (2018) Phase III study of dulanermin (recombinant human tumor necrosis factor-related apoptosis-inducing ligand/Apo2 ligand) combined with vinorelbine and cisplatin in patients with advanced non-small-cell lung cancer. Invest New Drugs 36(2):315–322. https://doi.org/10.1007/s10637-017-0536-y
Metwalli AR et al (2010) Smac mimetic reverses resistance to TRAIL and chemotherapy in human urothelial cancer cells. Cancer Biol Ther 10(9):885–892. https://doi.org/10.4161/cbt.10.9.13237
Voelkel-Johnson C, King DL, Norris JS (2002) Resistance of prostate cancer cells to soluble TNF-related apoptosis-inducing ligand (TRAIL/Apo2L) can be overcome by doxorubicin or adenoviral delivery of full-length TRAIL. Cancer Gene Ther 9(2):164–172. https://doi.org/10.1038/sj.cgt.7700420
Jin Z et al (2004) Deficient tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) death receptor transport to the cell surface in human colon cancer cells selected for resistance to TRAIL-induced apoptosis. J Biol Chem 279(34):35829–35839. https://doi.org/10.1074/jbc.M405538200
Pimentel JM et al (2023) The role of the immune response and inflammatory pathways in TNF-related apoptosis-inducing ligand (TRAIL) resistance in triple-negative breast cancer cells. Am J Cancer Res 13(10):4678–4692
Wilson C et al (2008) Interleukin-8 signaling attenuates TRAIL- and chemotherapy-induced apoptosis through transcriptional regulation of c-FLIP in prostate cancer cells. Mol Cancer Ther 7(9):2649–2661. https://doi.org/10.1158/1535-7163.Mct-08-0148
Todaro M et al (2008) Apoptosis resistance in epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ 15(4):762–772. https://doi.org/10.1038/sj.cdd.4402305
Todaro M et al (2008) IL-4-mediated drug resistance in colon cancer stem cells. Cell Cycle 7(3):309–313. https://doi.org/10.4161/cc.7.3.5389
Hu F et al (2021) IL-6 regulates autophagy and chemotherapy resistance by promoting BECN1 phosphorylation. Nat Commun 12(1):3651. https://doi.org/10.1038/s41467-021-23923-1
David JM et al (2016) The IL-8/IL-8R Axis: a double Agent in Tumor Immune Resistance. Vaccines (Basel) 4(3). https://doi.org/10.3390/vaccines4030022
Cheng KJ et al (2022) Interleukin-1 alpha and high mobility group box-1 secretion in polyinosinic:polycytidylic-induced colorectal cancer cells occur via RIPK1-dependent mechanism and participate in tumourigenesis. J Cell Commun Signal. https://doi.org/10.1007/s12079-022-00681-3
Han H et al (2020) Danger signals in liver injury and restoration of homeostasis. J Hepatol 73(4):933–951. https://doi.org/10.1016/j.jhep.2020.04.033
Xie K (2001) Interleukin-8 and human cancer biology. Cytokine Growth Factor Rev 12(4):375–391. https://doi.org/10.1016/s1359-6101(01)00016-8
Maharati A, Moghbeli M (2023) PI3K/AKT signaling pathway as a critical regulator of epithelial-mesenchymal transition in colorectal tumor cells. Cell Communication Signal 21(1):201. https://doi.org/10.1186/s12964-023-01225-x
Dong C et al (2021) Activation of PI3K/AKT/mTOR pathway causes drug resistance in breast Cancer. Front Pharmacol 12:628690. https://doi.org/10.3389/fphar.2021.628690
Büneker C, Mohr A, Zwacka RM (2009) The TRAIL-receptor-1: TRAIL-receptor-3 and – 4 ratio is a predictor for TRAIL sensitivity of cancer cells. Oncol Rep 21(5):1289–1295. https://doi.org/10.3892/or_00000353
Zhang L, Fang B (2005) Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther 12(3):228–237. https://doi.org/10.1038/sj.cgt.7700792
Sanlioglu AD et al (2005) Surface TRAIL decoy receptor-4 expression is correlated with TRAIL resistance in MCF7 breast cancer cells. BMC Cancer 5:54. https://doi.org/10.1186/1471-2407-5-54
Lipton A et al (2002) Serum osteoprotegerin levels in healthy controls and cancer patients. Clin Cancer Res 8(7):2306–2310
Sheikh MS et al (1999) The antiapoptotic decoy receptor TRID/TRAIL-R3 is a p53-regulated DNA damage-inducible gene that is overexpressed in primary tumors of the gastrointestinal tract. Oncogene 18(28):4153–4159. https://doi.org/10.1038/sj.onc.1202763
Toscano F et al (2008) p53-Mediated upregulation of DcR1 impairs oxaliplatin/TRAIL-induced synergistic anti-tumour potential in colon cancer cells. Oncogene 27(30):4161–4171. https://doi.org/10.1038/onc.2008.52
Granci V et al (2008) Prognostic significance of TRAIL-R1 and TRAIL-R3 expression in metastatic colorectal carcinomas. Eur J Cancer 44(15):2312–2318. https://doi.org/10.1016/j.ejca.2008.06.042
Jeon YJ et al (2008) Ribosomal protein S6 is a selective mediator of TRAIL-apoptotic signaling. Oncogene 27(31):4344–4352. https://doi.org/10.1038/onc.2008.73
Duldulao MP et al (2012) Gene expression variations in microsatellite stable and unstable colon cancer cells. J Surg Res 174(1):1–6. https://doi.org/10.1016/j.jss.2011.06.016
Zeinalian M et al (2018) Clinical aspects of microsatellite instability testing in Colorectal Cancer. Adv Biomed Res 7:28. https://doi.org/10.4103/abr.abr_185_16
Willms A et al (2019) Impact of p53 status on TRAIL-mediated apoptotic and non-apoptotic signaling in cancer cells. PLoS ONE 14(4):e0214847. https://doi.org/10.1371/journal.pone.0214847
Fohlen A et al (2021) Anticancer drugs for intra-arterial treatment of Colorectal Cancer Liver metastases: In-Vitro Screening after short exposure time. Pharmaceuticals (Basel) 14(7). https://doi.org/10.3390/ph14070639
Liu R et al (2020) PI3K/AKT pathway as a key link modulates the multidrug resistance of cancers. Cell Death Dis 11(9):797. https://doi.org/10.1038/s41419-020-02998-6
Qian HR et al (2017) Interplay between apoptosis and autophagy in colorectal cancer. Oncotarget 8(37):62759–62768. https://doi.org/10.18632/oncotarget.18663
Kim SW et al (2016) Niacin alleviates TRAIL-mediated colon cancer cell death via autophagy flux activation. Oncotarget 7(4):4356–4368. https://doi.org/10.18632/oncotarget.5374
Nishimoto-Kakiuchi A et al (2023) A long-acting anti-IL-8 antibody improves inflammation and fibrosis in endometriosis. Sci Transl Med 15(684):eabq5858. https://doi.org/10.1126/scitranslmed.abq5858
Bilusic M et al (2019) Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. J Immunother Cancer 7(1):240. https://doi.org/10.1186/s40425-019-0706-x
Zhu M, Jin Q, Xin Y (2021) Recent clinical advances in PI3K inhibitors on colorectal cancer. Pharmazie 76(12):568–573. https://doi.org/10.1691/ph.2021.1820
Acknowledgements
This research is funded by the Ministry of Higher Education under the Fundamental Research Grant Scheme (FRGS) FRGS/1/2020/SKK0/UM/02/36 (FP108-2020) and MAKNA Cancer Research Award 2020 (Grant Number: PV001-2021). The student was sponsored by the Faculty of Medicine Postgraduate Scholarship Scheme (FOMPSS).
Funding
This research is funded by the Ministry of Higher Education under the Fundamental Research Grant Scheme (FRGS) FRGS/1/2020/SKK0/UM/02/36 (FP108-2020) and MAKNA Cancer Research Award 2020 (Grant Number: PV001-2021). The student was sponsored by the Faculty of Medicine Postgraduate Scholarship Scheme (FOMPSS).
Author information
Authors and Affiliations
Contributions
KXJJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing, Visualization. EHMM: Supervision, Writing – review & editing. SEF: Supervision, Writing – review & editing. FNF: Writing – review & editing. KMV: Writing – review & editing. KI: Writing – review & editing. *ZAI: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Resources, Writing – original draft, Writing - Review & Editing, Visualization, Supervision, Project administration, Funding acquisition. * KXJJ and ZAI contributed equally to this work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jong, K.X.J., Mohamed, E.H.M., Syafruddin, S.E. et al. IL-8 and PI3K pathway influence the susceptibility of TRAIL-sensitive colorectal cancer cells to TRAIL-induced cell death. Mol Biol Rep 51, 978 (2024). https://doi.org/10.1007/s11033-024-09895-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11033-024-09895-7