
Abstract
Rheumatoid arthritis (RA) is an autoimmune disease characterized by chronic inflammation of synovial joints and often associated with chronic pain. Chronic joint inflammation is attributed to severe proliferation of synoviocytes and resident macrophages and infiltration of immune cells. These cells secrete pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α), interleukin 6 (IL-6) and IL-17 to overcome actions of anti-inflammatory cytokines, thereby maintaining chronic inflammation and pain. The imbalance between pro-inflammatory cytokines (produced by M1 macrophages) and anti-inflammatory cytokines (produced by M2 macrophages) is a feature of RA progression, but the switch time of M1/M2 polarization and which receptor regulates the switch remain unsolved. Here we used an established RA mouse model to demonstrate that TNF-α expression was responsible for the initial acute stage of inflammation and pain (1–4 weeks), IL-17 expression the transition stage (4–12 weeks), and IL-6 expression the later maintenance stage (> 12 weeks). The switch time of M1/M2 polarization occurred at 4–8 weeks. We also identified a potential compound, anthra[2,1-c][1,2,5] thiadiazole-6,11-dione (NSC745885), that specifically inhibited T-cell death-associated gene 8 (TDAG8) function and expression. NSC745885 decreased joint inflammation and destruction and attenuated pain by reducing cytokine production and regulating the M1/M2 polarization switch. TDAG8 may participate in regulating the M1/M2 polarization and temporal expression of distinct cytokines to control RA progression.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rheumatoid arthritis (RA), an autoimmune disease affecting 0.5% to 1% of the population worldwide, is associated with chronic inflammation and pain [15, 39]. RA is characterized by chronic inflammation of the synovium with hyperplasia of synovial lining cells, then leads to progressive and erosive destruction of bone and cartilage [10, 27].
In synovium, macrophage-like and fibroblast-like synoviocytes are activated and proliferate; they along with infiltrated macrophages secrete pro-inflammatory cytokines (e.g., tumor necrosis factor α [TNF-α], interleukin 1β [IL-1β], IL-6), and degradative enzymes that maintain inflammation, promote osteoclast maturation and activation, and finally destroy the articular structure [23]. Activated macrophages also produce anti-inflammatory cytokines (e.g., IL-10) to promote the resolution of inflammation and tissue repair, thus ameliorating the disease.
Pro- and anti-inflammatory cytokines are produced by M1 (classically activated) and M2 (alternatively activated) macrophages, respectively [14]. The imbalance between pro- and anti-inflammatory cytokines (M1/M2 macrophages) could be a key mechanism of rheumatic disease progression. RA patients have more of a M1 macrophage profile than do people with spondyloarthritis such as psoriatic arthritis [42] or osteoarthritis [40]. An acute hypoxia environment favors M2 macrophage polarization, but chronic hypoxia triggers M1 polarization [33]. Depletion of macrophages by using clodronate-containing liposomes reduced M1 macrophage number and arthritis score in mice [43]. Intraperitoneal injection of clodronate to remove macrophages also inhibited mechanical and thermal hyperalgesia [31]. In addition to macrophages, T cells also play important roles in RA progression. In the RA synovium, the bias is toward T helper 1 (Th1)-like cells [32], and the Th1 cytokine IL-17 contributes to RA progression by regulating IL-6 production [5].
TNF-α, IL-6 and IL-17 appear to play a pivotal role in inducing and maintaining RA disease progression, but they also contribute to RA-associated pain [36]. Neutralization of TNF-α attenuated hyperalgesia in an antigen-induced arthritis (AIA) model [3], and TNF-α blocker treatment reduced nociceptive responses in arthritis patients [16]. Injection of IL-6 into a normal knee caused long-lasting mechanical hyperalgesia [1], and deficiency of IL-6 receptor attenuated disease severity and mechanical hyperalgesia in an AIA model [8, 29]. Administration of IL-17 into a normal knee joint caused persistent pain on both innocuous and noxious rotation of the joint [34], and IL-17—deficient mice showed attenuated AIA-induced hyperalgesia [9]. In addition, T-cell death-associated gene 8 (TDAG8), recently found to regulate Th17 cells and IL-17 secretion [12], was found associated with regulating RA progression and pain [18].
Immune cells and immune cell- or synoviocyte-derived cytokines are important in RA disease severity and pain. However, how RA develops from the acute stage to the chronic state and whether different immune cells and cytokines regulate different stages of inflammation and pain remain unclear. In this study, we used an established RA mouse model [18] to address this question. We found that TNF-α was responsible for the initial acute stage of inflammation and pain (1–4 weeks), IL-17 was responsible for the transition stage (4–12 weeks); and IL-6 was responsible for the later maintenance stage (> 12 weeks). The ratio of M1/M2 macrophages was decreased in the transition stage (4–12 weeks), then increased after 12 weeks. We also identified a potential compound, anthra[2,1-c][1,2,5] thiadiazole-6,11-dione (NSC745885), that decreased pannus, reduced bone and cartilage damage, and attenuated pain by reducing the ratio of M1/M2 macrophages at 12 weeks and TNF-α, IL-17 and IL-6 production at 4, 8, 12 weeks. Given that NSC745885 specifically inhibited TDAG8 function and expression, TDAG8 likely participates in regulating M1/M2 polarization and the temporal expression of distinct cytokines, thus contributing to regulation of RA progression.
Materials and methods
Animals, agents and constructs
ICR mice (8–12 weeks old) were purchased from BioLASCO Taiwan (Taipei) and housed 3 to 5 per cage under a 12-h light/dark cycle (lights on at 7:00 am and off at 7:00 pm) with food and water ad libitum in a temperature- and humidity-controlled environment at the National Central University, Taiwan. Care and use of mice conformed to the Guide for the Use of Laboratory Animals (US National Research Council) and the experimental procedures were approved by the local animal use committee (IACUC, National Central University, Taiwan). All behavior testing was performed between 9:00 am and 5:00 pm. Efforts were made to minimize the number of animals used and their suffering.
NSC745885 and its derivatives (NSC757963, NSC761886, NSC757967) were synthesized in a previous study [20]. Complete Freund’s adjuvant (CFA), 2-(N-morpholino) ethanesulfonic acid (MES), and 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) were from Sigma. Tofacitinib was from Selleckchem. Proton-sensing receptor genes including mTDAG8, mOGR1, mGPR4, mG2A, mASIC3, mASIC1a, and mTRPV1 were cloned into the vector pIRES-hrGFP-2a (pIRES-GFP) as described [6, 19].
Animal models and compound administration
The acid model was established by intraplantar injection with 25 µl acid buffer (10 mM MES in saline, pH 5.0). For compound treatments, various concentrations of NSC745885, NSC757963, NSC761886, or NSC757967 were intraplantarly pre-injected before acid buffer injection, followed by animal behavior experiments at 60 min after acid injection. For gene expression experiments, lumbar 4–6 (L4-6) dorsal root ganglia (DRG) ipsilateral to injected paws were isolated at 60 min after acid injection for RNA extraction.
In the RA model, mice were injected with 5 μg CFA in the right tibiotarsal joint once a week for 4 weeks as described [18]. NSC745885 (160 or 300 μg/kg) was intraperitoneally (i.p.) injected into RA mice, followed by animal behavior experiments at 90 min after NSC745885 injection. For long-term treatments, 160 μg/kg NSC745885 was ip injected weekly for 9 consecutive weeks from week 5 of CFA injection.
Arthritis score and histological analysis
In the RA model, the severity of arthritis was scored from 0 to 5 as described [18]. Each limb was graded and given a maximum possible score of 15, the maximum score for an animal was 60.
Histological staining was performed as described [18]. Briefly, at 0, 4, 8, and 12 weeks after CFA injection, tibiotarsal joints were excised, fixed, decalcified, embedded in paraffin and sectioned with use of a microtome, then stained with hematoxylin and eosin (by the Taiwan Mouse Clinic, Taipei). Images were observed by light microscopy (Lecia, ICC50HD).
Arthritic changes were scored as described [18] on a scale from 0 to 5:0 = normal; 3 = moderate pannus, bone erosion or cartilage damage; 5 = severe pannus, bone erosion or cartilage damage. From each joint, 6 areas of 2 sections were scored. Mean scores were the average of all section scores for each animal.
Some joint sections were immersed in 100% xylene twice, then in 100%, 95%, 90%, 80% and 70% ethanol sequentially to remove paraffin wax, then placed in Tris-buffer (20 mM Tris base, 150 mM NaCl, pH 7.6) containing 1% bovine serum albumin and stained with anti-CD80 (1:250, Biorbyt, UK) [37], CD68 (1:100, Biorbyt) [37], CD163 (1:100, Biorbyt) [41], inducible nitric oxide synthase (iNOS, 1:100, GeneTex, North America) [21, 41], or CD206 (1:200, GeneTex, North America) [21, 41] antibodies at 4 ℃ for 16 h. Sections were then stained with alkaline phosphatase-conjugated goat anti-rabbit IgG (1:5000, Jackson Immunoresearch) at room temperature for 1 h, and signals were developed by using nitro-blue tetrazolium chloride and 5-bromo-4-chloro-3′-Indolyphosphate p-toluidine (Millipore). For each joint, 9 regions of one section were scored. Total immune cells were the average of 9 regions.
Measurement of cytokine levels
At 0, 4, 8, 12 weeks after CFA injection, mice were sacrificed and blood was collected as described [18]. Briefly, blood was left to clot and centrifugated to obtain serum samples. Serum levels of TNF-α (#MTA00B), IL-17 (#M1700) or IL-6 (#M6000B) were measured with ELISA kits from R&D Systems (Minnesota, USA) as instructed.
Assessment of acid-induced pain and arthritic pain
Mice with or without treatments were tested for withdrawal threshold to mechanical stimuli (von Frey filaments, Touch-Test; North Coast Medical, Morgan Hill, CA) applied to the hindpaw. Mice were pre-trained for 1 to 2 h each day for 2 days before the test. Von Frey fibers were applied 5 times at 5-s intervals to the plantar surface of each hindpaw at various times after injections. The paw withdrawal threshold (PWT) was determined when paw withdrawal was obtained in more than 3 of 5 applications.
Quantitative RT-PCR
DRG RNA extraction was performed as described [19]. Each DRG pool contained at least 9 DRG from 3 mice. RNA was extracted by using the RNeasy kit (Qiagen, Valencia, CA). The gene primers (100 nM, sequences listed in Table 1, derived cDNA and master mix [SYBR green I and AmpliTaq Gold DNA polymerase (Applied Biosystems, Foster City, CA)] were mixed for PCR reactions and product detection by use of ABI Prism 7300. For each assay, preparations were run in triplicate. The thermal cycling conditions were 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, and 60 °C for 1 min. The threshold cycle (Ct values of both the targets and internal references (mGAPDH or β-actin) were measured from the same samples, and the expression of the target genes relative to that of mGAPDH or β-actin was calculated by the comparative Ct method.
Cell culture and transfection
Human embryonic kidney 293 cells (HEK293T, obtained from the Bioresource Collection and Research Center of Food Industry Research and Development Institute, Taiwan) were cultured and maintained in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen) supplemented with 10% fetal bovine serum (Invitrogen) and 0.1% gentamicin. For Ca2+ imaging experiments, HEK293T cells were seeded at 4 × 105 on 24-mm poly-d-lysine-coated coverslips, transfected with 1.5 µg plasmids, then underwent Ca2+ imaging. For cAMP assay, HEK293T cells were seeded at 1.6 × 105 per well (70–80% confluence) in 12-well cell plates, transfected with 0.6 µg plasmid, then underwent cAMP assay.
Imaging of intracellular Ca2+ level ([Ca2+]i)
[Ca2+]i imaging was performed as described [6]. Briefly, transfected cells were pre-incubated at 37 °C with 2.5 µM, Fura-2 acetoxymethyl easter (Fura-2-AM, Molecular Probes) for 40 min, then washed 2 or 3 times with HEPES/MES buffer (125 mM NaCl, 1 mM KCl, 1 mM MgCl2, 5 mM CaCl2, 8 mM glucose, 10 mM HEPES, 15 mM MES, pH 7.6), supplemented with 300 µl HEPES/MES buffer (pH 7.6), then stimulated with pH 6.8 or 5.5 HEPES/MES buffer (600 µl). The pH-evoked Ca2+ transients were recorded by use of a Ca2+ imaging system equipped with a Leica DMI3000B fluorescence microscope and analyzed by use of MetaFluor software.
cAMP assay
The cAMP assay was performed as described [6]. Transfected cells were pre-incubated with serum-free DMEM containing 30 µM of the phosphodiesterase inhibitor RO201724 (Sigma), then stimulated with pH 6.8 HEPES/MES buffer containing 30 µM RO201724 for 30 min at 37 °C. After stimulation, cells were lysed in ethanol. The lysates were dried and cAMP level was quantified by use of a cAMP immunoassay kit (Assay Designs, MI) according to the manufacturer’s protocol.
Statistical analysis
All data are presented as mean ± SEM. One-way ANOVA with post-hoc Bonferroni test was used for calcium imaging and cAMP assay. Two-way ANOVA with post-hoc Bonferroni test was used for mechanical tests and cytokine assays. Mann–Whitney U test was used for analysis of immune cells, pannus, bone erosion and cartilage damage. p < 0.05 was considered statistically significant.
Results
Temporal expression of TNF-α, IL-6, and IL-17 levels in RA mice
Although TNF-α. IL-6, and IL-17 are important for RA development, the expression changes of these cytokines during RA development are still unclear, especially in RA animal models. Thus, we used an established RA animal model [18] to explore cytokine expression changes. In RA mice, joint swelling occurred during the first week and lasted for 12 weeks (Fig. 1a). The severity of joint swelling increased over time, peaked at 4 weeks (mean 36.3 ± 3.2) and lasted for 12 weeks (Fig. 1b). Bilateral hyperalgesia for mechanical stimuli also developed during the first week (mean ipsilateral PWT = 0.69 ± 0.31 and contralateral PWT = 0.81 ± 0.21) and remained for 12 weeks (mean ipsilateral PWT = 0.13 ± 0.03 and contralateral PWT = 0.42 ± 0.01) (Fig. 1c).

Intra-articular repeated administration of complete Freund’s adjuvant (CFA) induces long-term hyperalgesia and differential expression of cytokines. ICR mice (8–12 weeks old) were injected with 5 μl of 100% CFA (5 μg) in the right tibiotarsal joint (ipsilateral joint) 4 times at 1-week intervals, then underwent mechanical behavior tests. Black arrows are CFA treatments. a Images showing joint swelling at 0, 1, 2, 4, 8, 12 weeks. b Severity of arthritis presented as arthritis scores (n = 3). **p < 0.01, ***p < 0.001, 0 weeks vs other weeks by one-way ANOVA. c Paw withdrawal threshold (PWT) to mechanical stimuli measured at weekly intervals (n = 3). ***p < 0.001, 0 weeks vs other weeks by two-way ANOVA. d Levels of serum TNF-α, IL-6 and IL-17 (percentage of maximum levels) at 0, 4, 8 12 weeks
Serum levels of TNF-α, IL-17 and IL-6 were measured in RA mice at 0, 4, 8, 12 weeks (Fig. 1d). TNF-α level peaked at week 4, then declined at week 8, and returned slightly at week 12. IL-17 level did not increase until week 8 and decreased at week 12. IL-6 level gradually increased over time and reached a high level at week 12. These results suggest that TNF-α could be responsible for initial RA development, IL-17 for the transition from an initial RA stage to a chronic stage, and IL-6 for maintaining chronic RA.
RA progression favors polarization of M1 macrophages
The imbalance between pro-inflammatory (M1) and anti-inflammatory (M2) macrophages could be a key mechanism of rheumatic disease progression; however, the relationship between M1/M2 imbalance and RA progression is not clear in animal models. To address this question, we stained histological samples of joints from RA mice at 0, 4, 8, and 12 weeks with anti-CD68, anti-CD80 and anti-CD163 antibodies. The number of CD68+ cells was increased from week 4, peaked at week 8 and was maintained at high levels to week 12 (Fig. 2a, b). CD80+ cell number was greatly increased from week 8 and continued to increase at week 12 (Fig. 2a, c). The number of CD163+ cells was significantly increased from week 4 and peaked at week 8, but declined at week 12 (Fig. 2a, d). To confirm the CD80+ and CD163+ results, similar experiments were performed using iNOS as M1 marker and CD206 as M2 marker. iNOS-positive cell number was greatly increased from week 8 as CD80+ number (Fig. 3a, b) and CD206+ cell number was increased from week 4 and peaked at week 8 (Fig. 3a, c). The ratio of CD80 (M1) to CD163 (M2) was 89:11 at week 0 and was decreased at week 4 (32:68), then gradually returned at week 8 (73:27) and increased at week 12 (96:4) (Fig. 4a). Similar results were obtained from the ratio of iNOS to CD206 (Fig. 4b), the ratio of CD80 to CD206 (Fig. 4c), and the ratio of iNOS to CD163 (Fig. 4d). It suggests that the transition stage (4–8 weeks) favored M2 polarization and the later stage favored M1 polarization.

RA progression favors M1 macrophage polarization. Histology of joints from RA mice at weeks 0, 4, 8, 12 stained with anti-CD68 (b), CD80 (c), or CD163 (d) antibodies. Synovial sublining regions are shown in (a). Black arrows are labeled cells. Data are mean ± SEM cell density (cells/mm2) (b–d). *p < 0.05, ***p < 0.001, 0 week vs 4, 8, 12 weeks by Mann–Whitney U test

Histology of joints from RA mice at weeks 0, 4, 8, 12 stained with anti-iNOS (b) or CD206 (c) antibodies. Synovial sublining regions are shown in (a). Black arrows are labeled cells. Data are mean ± SEM cell density (cells/mm2) (b, c). *p < 0.05, ***p < 0.001, 0 week vs 4, 8, 12 weeks by Mann–Whitney U test

The ratio of M1 to M2 macrophages at weeks 0, 4, 8, 12. a M1:CD80; M2:CD163. b M1:iNOS; M2:CD206. c M1:CD80; M2:CD206. d M1:iNOS; M2:CD163
NSC745885 inhibits TDAG8-mediated signals and gene expression to relieve acid-induced pain
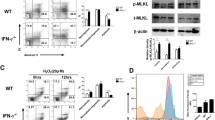
We previously found that TDAG8 gene deletion or suppression by shRNA reduced RA-associated pain [18]. However, how TDAG8 regulates RA pain is unknown. To further address this question, we synthesized a series of NSC745885 homologues [20] to screen potential TDAG8 antagonists or agonists. Because TDAG8 is a proton-sensing receptor, we used an acid-induced pain model as a primary model to screen compounds. Different concentrations of NSC745885 (Fig. 5a) homologues were intraplantarly injected into mice, followed by injection of pH 5.0 acid buffer. At 60 min after acid injection, NSC745885, NSC757963, and NSC757967 but not NSC761886 reversed the acid-induced pain (Fig. 5b). We isolated DRG from NSC745885-treated mice and found only the expression of TDAG8 among other possible genes inhibited either using GAPDH or β-actin as the internal control (Fig. 5c, d), which suggests that NSC745885 inhibited TDAG8 expression to block acid-induced pain.

NSC745885 inhibits TDAG8-mediated signals and gene expression to relieve acid-induced pain. a Structure of NSC745885. b Mice were intraplantarly pre-injected with different doses of NSC745885 (885, n = 6), NSC757963 (n = 3), NSC761886 (n = 3), or NSC757967 (n = 3), followed by pH 5.0 acid injection or non-injection control (Ctrl, n = 6). The PWT was measured at 60 min post-acid injection. ***p < 0.001, acid-injected vs non-injected groups; ###p < 0.001, compound-treated vs acid-injected by one-way ANOVA. c, d Mice (n = 3) were pre-injected with or without 0.25 nmol NSC745885 before pH 5.0 acid injection. At 60 min post-acid injection, quantitative RT-PCR was used to measure the expression of genes in the ipsilateral lumbar 4–6 DRG. The expression of each gene was normalized to the internal control mGAPDH (c) or β-actin (d). *p < 0.05, (+) 885 vs (−) 885 by one-way ANOVA. e, f, HEK293T cells transfected with mTDAG8-IRES or IRES vector were pretreated without or with different concentrations of NSC745885, followed by pH 6.8 HEPE/MES buffer for cAMP measurement (e) or pH 5.5 HEPE/MES buffer treatments for calcium imaging (f). *p < 0.05, ***p < 0.001, 885-treated (0.1, 1, 3, 10 μM) vs untreated (0 μM) by one-way ANOVA. g HEK293T cells transfected with different genes were pretreated without or with 1 μM NSC745885 before pH 6.8 (for OGR1) or pH 5.5 (for other genes) HEPE/MES buffer, followed by detection of calcium level. ***p < 0.001, (+) 885 vs (−) 885 (0 μM) by one-way ANOVA
Because TDAG8 activation induces cAMP accumulation and intracellular calcium increase [7], we then examined whether NSC745885 could inhibit TDAG8 function. HEK293T cells were transfected with TDAG8 or vector and pre-treated with different concentrations of NSC745885, then stimulated with pH 6.8 buffer. NSC745885 at 0.1 and 1 μM partially reduced cAMP level but at 3 μM completely blocked cAMP accumulation (Fig. 5e). NSC745885 at 1, 3, 10 μM completely inhibited calcium increase (Fig. 5f). We examined the NSC745885 inhibitory effect in other proton-sensing receptors and found only TDAG8-mediated calcium increase specifically inhibited (Fig. 5g). These results suggest that NSC745885 is able to inhibit TDAG8 gene expression and function.
NSC745885 attenuates RA-induced mechanical hyperalgesia and synovial inflammation
If NSC745885 inhibits TDAG8 function, it should reduce RA-associated pain. To address this question, after CFA injection for 4 weeks, mice were intraperitoneally (i.p.) injected once with NSC745885 at 160 and 300 µg/kg, then underwent a mechanical behavior test. Both 160 and 300 µg/kg NSC745885 attenuated the mechanical hyperalgesia at 90 min (Fig. 6a). NSC745885 300 µg/kg injected weekly for 9 consecutive weeks effectively reduced the mechanical hyperalgesia in both ipsilateral and contralateral sides, although arthritis scores remained unchanged (Fig. 6b–d). As compared with tofacitinib (commercial RA drug, 3 mg/kg), NSC745885 (300 µg/kg) had a greater analgesic effect from weeks 4 to 7 on the ipsilateral side (Fig. 6c).

NSC745885 reduces mechanical hyperalgesia in RA mice. Mice were treated as in Fig. 1. a After 4 weeks, mice were intraperitoneally injected with different doses of NSC745885 (160, 300 µg/kg), then underwent mechanical behavior tests with von-Frey filaments. **p < 0.01, ***p < 0.001, NSC745885 vs vehicle by one-way ANOVA. b Severity of arthritis is presented as arthritis scores. c, d For long-term treatments, vehicle (n = 6), 300 µg/kg NSC745885 (885, n = 5) or 3 mg/kg tofacitinib (Tofa, n = 3) was intraperitoneally administered weekly for 9 consecutive weeks. Mechanical tests were performed 90 min after compound injection every week from week 4. Data are mean ± SEM of total tested mice. *p < 0.5, **p < 0.01, ***p < 0.001, vehicle- vs NSC745885-treated at 90 min; #p < 0.5, ##p < 0.01, ###p < 0.001 vehicle- vs tofacitinib-treated at 90 min by two-way ANOVA. Black arrows are CFA treatments and white arrows are compound treatments
Histopathological examination of joints of mice treated with NSC745885, tofacitinib, or vehicle at 12 weeks revealed that both NSC745885 and tofacitinib reduced pannus, but only NSC745885 decreased bone erosion and cartilage damage induced by repeated CFA injection (Fig. 7a–d).

Histology of ipsilateral side joints in compound-treated RA mice. Mice were treated as in Fig. 6c. Histology samples of joints from vehicle-, tofacitinib- or NSC745885-treated mice at week 12 stained with hematoxylin and eosin are shown in (a). Data are mean ± SEM severity score for pannus (b), bone erosion (c), and cartilage damage (d). ***p < 0.001, vehicle vs compound-treated; ###p < 0.001, tofacitinib- vs NSC745885-treated by one-way ANOVA
NSC745885 alters M1/M2 macrophage polarization and inhibits the increase in cytokine levels
To understand whether NSC745885 modulates M1 or M2 macrophages to affect hyperalgesia and inflammation, we stained histological samples of joints from NSC745885-, tofacitinib-, or vehicle-treated mice at week 12 with anti-CD68, -CD80 or -CD163 antibodies. Both NSC745885 and tofacitinib significantly reduced CD68+ cell number (Fig. 8a, b), which was consistent with reduced pannus (Fig. 7b). Both NSC745885 and tofacitinib decreased CD80+ cell number, but only NSC745885 increased CD163+ cell number (Fig. 8a, c, d). CD163+ macrophages have roles in anti-inflammation and tissue repair, which could explain why NSC745885 treatment decreased bone erosion and cartilage damage. Although both tofacitinib and NSC745885 decreased the total number of immune cells, only NSC745885 treatment decreased the ratio of M1 to M2 macrophages (70:30 vs 96:4 with vehicle treatment and 95:5 with tofacitinib treatment; Fig. 8e). These results suggest that NSC745885 regulates the switch of M1/M2 polarization. The switch of M1/M2 polarization could explain in part the attenuated mechanical hyperalgesia.

Histology of ipsilateral side joints stained with macrophage markers in compound-treated RA mice. Mice were treated as in Fig. 6c. Histology samples of joints from control (ctrl, without CFA injection), vehicle-, tofacitinib (Tofa)- or NSC745885 (885)-treated mice at week 12 stained with anti-CD68 (b), -CD80 (c), or -CD163 (d) antibodies. Synovial sublining regions are shown in (a). Data are mean ± SEM cell density (cells/mm2) (b–d). * p < 0.05, **p < 0.01, ***p < 0.001, ctrl vs treated group; #p < 0.05, ##p < 0.01, ###p < 0.001, compound- vs vehicle-treated group by Mann–Whitney U test. Black arrows are labeled cells. e Ratio of M1 to M2 macrophages at week 12 for vehicle-, tofacitinib- or NSC745885-treated mice
Serum levels of TNF-α, IL-17 and IL-6 were examined at 0, 4, 8, 12 weeks in NSC745885- or vehicle-treated mice. NSC745885 treatment inhibited the increase in TNF-α level at week 4, IL-17 level at week 8, and IL-6 level at week 12 (Fig. 9a–c), which suggests that NSC745885 inhibited cytokine increase at different times, contributing to attenuated mechanical hyperalgesia.

Serum levels of TNF-α, IL-6 and IL-17 in NSC745885-treated or untreated RA mice. Mice were treated as in Fig. 6c. Blood samples were taken from NSC745885-treated or untreated RA mice at week 0, 4, 8, 12 to measure serum levels of TNF-α (a), IL-17 (b), and IL-6 (c). *p < 0.05, **p < 0.01, ***p < 0.001, 0 vs 4, 8, 12 weeks; #p < 0.05, ##p < 0.01, 885(+) vs 885(−) by two-way ANOVA

A putative model of RA progression modulating by temporal expression of distinct cytokines and M1/M2 macrophage polarization. a In the initial acute stage (1–4 weeks) of RA, synovial macrophages and intimal fibroblast-like synoviocytes (CD68+ cells) proliferate and together with infiltrated pro-inflammatory (M1) macrophages secrete TNF-α, triggering inflammation and pain. The ratio of M1 to M2 macrophages is high. In the transition stage (4–12 weeks), anti-inflammatory and repair macrophages (CD163+ cells) are largely increased to promote the resolution of inflammation and ameliorate the disease severity. The ratio of M1 to M2 macrophages is decreased. IL-17 produced by Th-17 cells mediates the transition to the chronic state. With RA progression (> 12 weeks), the number of anti-inflammatory (M2) macrophages is decreased and pro-inflammatory (M1) macrophages (CD80+ cells) increased to secrete IL-6, resulting in chronic inflammation and pain. b NSC745885 inhibits TDAG8 gene expression and function; inhibits proinflammatory cytokines TNF-α, IL-17, and IL-6; reduces M1 macrophage number and increases M2 macrophage number to reduce pannus and joint destruction and attenuating pain
Discussion
In this study, we first demonstrated that RA progression in the mouse model required temporal expression of distinct cytokines and a switch of M1/M2 polarization. TNF-α was responsible for the initial acute stage of inflammation and pain (1–4 weeks), IL-17 the transition stage (4–12 weeks) and IL-6 the later maintenance stage (> 12 weeks). In the initial stage, the number of synovial macrophages and intimal fibroblast-like synoviocytes (CD68+ cells) was increased to trigger inflammation and pain, then in the transition stage, the number of anti-inflammatory and repair macrophages (CD163+ cells) was greatly increased to reduce inflammation and joint damage. With RA progression, the number of CD163+ cells (M2 macrophages) was decreased, but that of CD80+ cells (M1 macrophages) was increased, thereby resulting in chronic inflammation and pain (Fig. 10a). Here we identified a potential compound, NSC745885, that inhibited TDAG8 function and expression, which reduced synovial inflammation, joint damage, and associated pain. Such effects were probably attributed to reduced cytokine levels (TNF-α, IL-17 and IL-6) and the switch of M1/M2 polarization (Fig. 10b), which implies that TDAG8 regulates RA disease severity and pain by modulating M1/M2 polarization and cytokine levels at different stages.
RA is a systemic inflammatory disease. How and when the acute inflammation becomes systemic, chronic inflammation is important to dissect the RA pathogenesis. Resolution of inflammation is an active process, and the inability to resolve acute inflammation may lead to a chronic inflammatory state. The role of macrophages in RA is increasingly appreciated. Consistent with a previous study finding a correlation of synovial macrophage number with disease activity [38], we found that synovial CD68+ cell number was greatly increased from week 4, peaked at week 8, then was maintained until week 12.
Weeks 1 to 4 may represent an acute stage for RA development. At this stage, severe proliferation of synoviocytes and macrophages causes hypoxia, which prolongs macrophage survival [35] and affects macrophage polarization. An acute hypoxia environment favors M2 macrophage polarization, but chronic hypoxia triggers M1 polarization [33]. In agreement, we found the number of M2 macrophages (CD163+) increased from week 4, peaked at week 8, and then decreased at week 12. In contrast, the number of M1 macrophages (CD80+) started to increase from week 8 and especially at week 12. The initial inflammatory stage (1–4 weeks) may cause acute hypoxia to trigger M2 polarization from week 4. M2 macrophages induce anti-inflammatory/pro-resolving and tissue-repair actions to resolve inflammation (4–8 weeks). Indeed, in our mouse model, pannus and bone erosion were slightly decreased at week 8 [18], probably due to the action of M2 macrophages. If inflammation is not resolved, then prolonged hypoxia (> 8 weeks) favors M1 polarization and survival to lead to chronic inflammatory disease (> 12 weeks). The ratio of M1 to M2 macrophages was higher in the beginning of RA, then declined at week 4, gradually returned at week 8, and was maintained at a high level at week 12. The changes in M1/M2 polarization indicated the transition from the acute to chronic state of inflammation.
Corresponding with macrophage actions, TNF-α is a master cytokine released by monocytes/macrophages at an acute state to induce other pro-inflammatory cytokines (IL-1β, IL-6) [4, 28]. We found TNF-α as the first cytokine to regulate RA progression in our RA mice, and the action of IL-6 appeared after TNF-α. In a previous study of patients with pre-RA (early RA) and established RA [24], TNF-α levels were slightly higher in pre-RA patients, but IL-6 levels were significantly increased in established RA patients. Consistent with Kokkonen et al. [24], we found high TNF-α levels at week 4, which then declined. IL-6 levels were gradually increased from week 4 after TNF-α levels peaked. IL-6 expression became dominant at week 12 to maintain the chronic state. IL-6 production was consistent with previous suggestions that IL-6 acts on both acute and chronic phases of inflammation [11]. A previous study found high serum IL-6 level only in RA patients but not in other inflammatory arthritic patients [17]. Therefore, serum IL-6 level is a good indicator of chronic inflammation. In our RA mouse model, serum IL-6 level was high at 12 weeks, which suggests that RA mice were in a chronic inflammatory state at 12 weeks.
TNF-α and IL-6 are also associated with the development and expansion of Th17 cells that produce IL-17; IL-17 also facilitates TNF-α and IL-6 secretion [5, 25]. As expected, we found the highest IL-17 levels at 8 weeks after TNF-α level peaked and IL-6 expression was induced, then IL-17 level declined at 12 weeks. These data agree with Kokkonen et al. [24] of high IL-17 level in pre-RA but not established RA patients. In the Kokkonen et al. study, RA progression was divided into only two stages: early and established RA. Both TNF-α and IL-17 levels were high in early RA, but IL-6 levels were high in established RA. According to this model, 1- to 12-week RA mice are in the early RA stage, but > 12-week RA mice are in established RA. In the RA mouse model, we could further divide the early RA stage into the acute stage and transition stage, with established RA the maintenance stage (Fig. 10). TNF-α is the master cytokine regulating the acute state, IL-7 controls the transition state, and IL-6 is responsible for the maintenance state. During the transition, the ratio of M1 to M2 macrophages was low, reflecting their functions in the resolution of inflammation and chronic development of inflammation.
TNF-α, IL-17 and IL-6 all directly or indirectly regulate nociceptors, thereby contributing to pain [1, 3, 8, 9, 16, 29, 34]. Pro-inflammatory macrophages are also involved in pain via cytokine actions. The use of leukapheresis therapy in severe RA to remove active monocytes lowers cytokine production [13]. Arthritis in animals is ameliorated by depletion of CCR2+ monocytes by using anti-CCR2 monoclonal antibodies or macrophages by using clodronate-containing liposomes [2, 43]. Intraperitoneal injection of clodronate to remove macrophages also inhibits mechanical and thermal hyperalgesia [31]. Thus, macrophage and continuous cytokine actions may explain why long-lasting pain accompanies RA. In agreement with this hypothesis, NSC745885 treatment attenuated the mechanical hyperalgesia due to inhibition of increased cytokine levels at different stages and the decrease in ratio of M1 to M2 macrophages. Pannus, bone erosion, and cartilage damage were also reduced with NSC745885 treatment.
NSC745885 specifically inhibited TDAG8 expression and TDAG8-mediated signaling. TDAG8 deletion or suppression attenuates RA progression and pain [18], so the NSC745885-mediated inhibitory effect could be due to blocking TDAG8 expression and function. Given that TDAG8 is expressed in nociceptors, macrophages, T cells, and microglia [19, 22, 26, 30], blocking TDAG8 expression and function may have direct effects on nociceptors to inhibit pain signals and indirect effects via immune cells to reduce pain signals. Blocking TDAG8 expression and function continuously inhibited different cytokines, thereby resulting in long-lasting inhibition of mechanical hyperalgesia. TDAG8 could be a potential drug target for future therapeutic development.
Conclusions
This study demonstrated that TNF-α, IL-17 and IL-6 regulated different stages of RA progression in the animal model. TNF-α (peak expression at 4 weeks) was the master cytokine for the acute stage, IL-17 (peak expression at 8 weeks) was responsible for the transition stage, and IL-6 (peak expression at 12 weeks) maintained the chronic state. In the acute stage, initial inflammation and severe proliferation of synoviocytes and macrophages caused the acute hypoxia that favored M2 macrophage polarization in the transition stage and reduced the ratio of M1 to M2 macrophages. In the chronic state, the long-lasting hypoxia favored M1 macrophage polarization and increased the ratio of M1 to M2 macrophages, thereby contributing to RA progression. NSC745885 continuously blocked different cytokines and reduced the ratio of M1 to M2 macrophages, attenuating long-lasting mechanical hyperalgesia. NSC745885 action was via blocking TDAG8 expression and function. This is the first study to provide evidence that TDAG8 regulates different pro-inflammatory cytokines and M1/M2 macrophage polarization to control RA progression and pain. TDAG8 could be a potential drug target for the development of therapeutic treatments.
References
Brenn D, Richter F, Schaible HG (2007) Sensitization of unmyelinated sensory fibers of the joint nerve to mechanical stimuli by interleukin-6 in the rat: an inflammatory mechanism of joint pain. Arthritis Rheum 56:351–359
Brühl H, Cihak J, Plachy J, Kunz-Schughart L, Niedermeier M, Denzel A, Gomez MR, Talke Y, Luckow B, Stangassinger M, Mack M (2007) Targeting of Gr-1+, CCR2+ monocytes in collagen-induced arthritis. Arthritis Rheum 56:2975–2985
Boettger MK, Hensellek S, Richter F, Gajda M, Stockigt R, Banchet GSV, Brauer R, Schaible HG (2008) Antinociceptive effects of tumor necrosis factor alpha neutralization in a rat model of antigen-induced arthritis: evidence of a neuronal target. Arthritis Rheum 58:2368–2378
Butler DM, Maini RN, Feldmann M, Brennan FM (1995) Modulation of proinflammatory cytokine release in rheumatoid synovial membrane cell cultures. Comparison of monoclonal anti TNF-alpha antibody with the interleukin-1 receptor antagonist. Eur Cytokine Netw 6(4):225–230
Chabaud M, Fossiez F, Taupin JL, Miossec P (1998) Enhancing effect of IL-17 on IL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritis synoviocytes and its regulation by Th2 cytokines. J Immunol 161(1):409–414
Chen YJ, Huang CW, Lin CS, Chang WH, Sun WH (2009) Expression and function of proton-sensing G-protein-coupled receptors in inflammatory pain. Mol Pain 5:19
Dai SP, Huang YH, Chang CJ, Huang YF, Hsieh WS, Tabata Y, Ishii S, Sun WH (2017) TDAG8 involved in initiating inflammatory hyperalgesia and establishing hyperalgesic priming in mice. Sci Rep 7:41415
Ebbinghaus M, Segond von Banchet G, Massier J, Gajda M, Bräuer R, Kress M, Schaible HG (2015) Interleukin-6-dependent influence of nociceptive sensory neurons on antigen-induced arthritis. Arthritis Res Ther 17:334
Ebbinghaus M, Natura G, Segond von Banchet G, Hensellek S, Böttcher M, Hoffmann B, Salah FS, Gajda M, Kamradt T, Schaible HG (2017) Interleukin-17A is involved in mechanical hyperalgesia but not in the severity of murine antigen-induced arthritis. Sci Rep 7(1):10334
Firestein GS (2003) Evolving concepts of rheumatoid arthritis. Nature 423:356–361
Gabay C (2006) Interleukin-6 and chronic inflammation. Arthritis Res Ther 8(suppl 2):S3
Gaublomme JT, Yosef N, Lee Y, Gertner RS, Yang LV, Wu C, Pandolfi PP, Mak T, Satija R, Shalek AK, Kuchroo VK, Park H, Regev A (2015) Single-cell genomics unveils critical regulators of Th17 cell pathogenicity. Cell 163:1400–1412
Hahn G, Stuhlmuller B, Hain N, Kalden JR, Pfizenmaier K, Burmester GR (1993) Modulation of monocyte activation in patients with rheumatoid arthritis by leukapheresis therapy. J Clin Invest 91:862–870
Hamilton Tak JAPP (2009) Analysis of the synovial cell infiltrate in early rheumatoid synovial tissue in relation to local disease activity. Arthritis Rheum 60:1210–1221
Heiberg T, Finset A, Uhlig T, Kvien TK (2005) Seven year changes in health status and priorities for improvement of health in patients with rheumatoid arthritis. Ann Rheum Dis 64:191–195
Hess A, Axmann R, Rech J, Finzel S, Heindl C, Kreitz S, Sergeeva M, Saake M, Garcia M, Kollias G, Straub RH, Sporns O, Doerfler A, Brune K, Schett G (2011) Blockade of TNF-α rapidly inhibits pain responses in the central nervous system. PNAS 108:3731–3736
Houssiau FA, Devogelaer JP, Van Damme J, de Deuxchaisnes CN, Van Snick J (1988) Interleukin-6 in synovial fluid and serum of patients with rheumatoid arthritis and other inflammatory arthritides. Arthritis Rheum 31(6):784–788
Hsieh WS, Kung CC, Huang SL, Lin SC, Sun WH (2017) TDAG8, TRPV1, and ASIC3 involved in establishing hyperalgesic priming in experimental rheumatoid arthritis. Sci Rep 7:8870
Huang CW, Tzeng JN, Chen YJ, Tsai WF, Chen CC, Sun WH (2007) Nociceptors of dorsal root ganglion express proton-sensing G protein-coupled receptors. Mol Cell Neurosci 36:195–210
Huang HS, Chen TC, Chen RH, Huang KF, Huang FC, Jhan JR, Chen CL, Lee CC, Lo Y, Lin JJ (2009) Synthesis, cytotoxicity and human telomerase inhibition activities of a series of 1,2-heteroannelated anthraquinones and anthra [1,2-d] imidazole-6,11-dione homologues. Bioorg Med Chem 17:7418–7428
Ishikawa J, Takahashi N, Matsumoto T, Yoshioka Y, Yamamoto N, Nishikawa M, Hibi H, Ishigro N, Ueda M, Furukawa K, Yamamoto A (2016) Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental rheumatoid arthritis. Bone 83:210–219
Jin Y, Sato K, Tobo A, Mogi C, Tobo M, Murata N, Ishii S, Im DS, Okajima F (2014) Inhibition of interleukin-1β production by extracellular acidification through the TDAG8/cAMP pathway in mouse microglia. J Neurochem 129:683–695
Kennedy A, Fearon U, Veale DJ, Godson C (2011) Macrophages in synovial inflammation. Front Immunol 2:52
Kokkonen H, Söderström I, Rocklöv J, Hallmans G, Lejon K, Rantapää DS (2010) Up-regulation of cytokines and chemokines predates the onset of rheumatoid arthritis. Arthritis Rheum 62(2):383–391
Kolls JK, Lindén A (2004) Interleukin-17 family members and inflammation. Immunity 21(4):467–476
Mogi C, Tobo M, Tomura H, Murata N, He XD, Sato K, Kimura T, Ishizuka T, Sasaki T, Sato T, Kihara Y, Ishii S, Harada A, Okajima F (2009) Involvement of proton-sensing TDAG8 in extracellular acidification-induced inhibition of proinflammatory cytokine production in peritoneal macrophages. J Immunol 182:3243–3251
Morris VH, Cruwys SC, Kidd BL (1997) Characterization of capsacin-induced mechanical hyperalgesia as a marker for altered nociceptive processing in patients with rheumatoid arthritis. Pain 71:179–186
Nawroth PP, Bank I, Handley D, Cassimeris J, Chess L, Stern D (1986) Tumor necrosis factor/cachectin interacts with endothelial cell receptors to induce release of interleukin 1. J Exp Med 163:1363–1375
Nowell MA, Richards PJ, Horiuchi S, Yamamoto N, Rose JS, Topley N, Williams AS, Jones SA (2003) Soluble IL-6 receptor governs IL-6 activity in experimental arthritis: blockage of arthritis severity by soluble glycoprotein 130. J Immunol 171:3202–3209
Onozawa Y, Fujita Y, Kuwabara H, Nagasaki M, Komai T, Oda T (2012) Activation of T cell death-associated gene 8 regulates the cytokine production of T cells and macrophages in vitro. Eur J Pharmacol 683:325–331
Peng J, Gu N, Zhou L, Eyo UB, Murugan M, Gan WB, Wu LL (2016) Microglia and monocytes synergistically promote the transition from acute to chronic pain after nerve injury. Nat Commun 7:12029
Qin S, Rottman JB, Myers P, Kassam N, Weinblatt M, Loetscher M, Koch AE, Moser B, Mackay CR (1998) The chemokine receptors CXCR3 and CCR5 mark subsets of T cells associated with certain inflammatory reactions. J Clin Invest 101(4):746–754
Raggi F, Pelassa S, Pierobon D, Penco F, Gattorno M, Novelli F, Eva A, Varesio L, Giovarelli M, Bosco MC (2017) Regulation of human macrophage M1–M2 polarization balance by hypoxia and the triggering receptor expressed on myeloid cells-1. Front Immunol 8:1097
Richter F, Natura G, Ebbinghaus M, von Banchet GS, Hensellek S, König C, Bräuer R, Schaible HG (2012) Interleukin-17 sensitizes joint nociceptors to mechanical stimuli and contributes to arthritic pain through neuronal interleukin-17 receptors in rodents. Arthritis Rheum 64:4125–4134
Roiniotis J, Dinh H, Masendycz P, Turner A, Elsegood CL, Scholz GM, Hamilton JA (2009) Hypoxia prolongs monocyte/macrophage survival and enhanced glycolysis is associated with their maturation under aerobic conditions. J Immunol 82(12):7974–7981
Schaible HG (2014) Nociceptive neurons detect cytokines in arthritis. Arthritis Res Ther 16:470
Schrijver IA, Melief MJ, Tak PP, Hazenberg MP, Laman JD (2000) Antigen-presenting cells containing bacterial peptidoglycan in synovial tissues of rheumatoid arthritis patients coexpress costimulatory molecules and cytokines. Arthritis Rheum 43:2160–2168
Tak PP, Smeets TJ, Daha MR, Kluin PM, Meijers KA, Brand R, Meinders AE, Breedveld FC (1997) Analysis of the synovial cell infiltrate in early rheumatoid synovial tissue in relation to local disease activity. Arthritis Rheum 40(2):217–225
ten Klooster PM, Veehof MM, Taal E, Riel P, Laar M (2007) Changes in priorities for improvement in patients with rheumatoid arthritis during 1 year of anti-tumour necrosis factor treatment. Ann Rheum Dis 66:1485–1490
Tsuneyoshi Y, Tanaka M, Nagai T, Sunahara N, Matsuda T, Sonoda T, Ijiri K, Komiya S, Matsuyama T (2012) Functional folate receptor beta-expressing macrophages in osteoarthritis synovium and their M1/M2 expression profiles. Scand J Rheumatol 41(2):132–140
Udalova IA, Mantovani A, Feldmann M (2016) Macrophage heterogeneity in the context of rheumatoid arthritis. Nat Rev Rheumatol 12(8):472–485
Vandooren B, Noordenbos T, Ambarus C, Krausz S, Cantaert T, Yeremenko N, Boumans M, Lutter R, Tak PP, Baeten D (2009) Absence of a classically activated macrophage cytokine signature in peripheral spondylarthritis, including psoriatic arthritis. Arthritis Rheum 60:966–975
Zhang Q, Yuan R, Li C, Wei W, Shen W, Cui Y, Yuan X (2019) Macrophages depletion with clodronate-containing liposomes affects the incidence and development of rheumatoid arthritis. Z Rheimatol 78:996–1003
Acknowledgements
We thank the Taiwan Mouse Clinic (MOST 107-2319-B-001-002), which is funded by the National Research Program for Biopharmaceuticals (NRPB) at the Ministry of Science and Technology (MOST) of Taiwan, for technical support in H&E staining.
Funding
This work was supported by the funds from the Ministry of Science and Technology, Taiwan (Grant No. MOST 106-2320-B-008-004-MY3).
Author information
Authors and Affiliations
Contributions
CCK participated in the animal model, data analysis and manuscript writing. SPD contributed to in vivo and in vitro tests of compounds, data analysis and manuscript writing. HC participated in animal experiments, immunostaining, and cytokine analysis, and manuscript writing. HSH contributed to the compound design, synthesis, and testing. WHS initiated the project, conceived the study, and participated in experimental design, coordination and data interpretation, and manuscript writing.
Corresponding author
Ethics declarations
Conflicts of interest
None of the authors have any non-financial conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Kung, CC., Dai, SP., Chiang, H. et al. Temporal expression patterns of distinct cytokines and M1/M2 macrophage polarization regulate rheumatoid arthritis progression. Mol Biol Rep 47, 3423–3437 (2020). https://doi.org/10.1007/s11033-020-05422-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-020-05422-6