
Abstract
The apple (Malus x domestica) scab (Venturia inaequalis) resistance genes Rvi4 and Rvi15 were mapped to a similar region on the top of linkage group 2 and both resistance genes elicit the same type of resistance reaction, i.e., a hypersensitive response; hence, it is suspected that the two genes may be the same. As the two resistance genes Rvi4 and Rvi15 are currently used in apple breeding, it is important to clarify whether the two resistance genes are the same or not. Several approaches were used to make this determination. First, the pedigree of the genotype GMAL 2473, the source of Rvi15, was reconstructed. GMAL 2473 was found to be an F1 of ‘Russian seedling’, the genotype, which is known to also be the source of Rvi4. Next, it was further demonstrated that ‘Regia’, a cultivar known to carry Rvi4 (and Rvi2), carries the same gene (Vr2-C), which was demonstrated to be the gene inducing Rvi15 resistance. Finally, it was shown that transgenic lines carrying Vr2-C are compatible with race 4 apple scab isolates. Taken all together, these results definitively demonstrate that Rvi4 and Rvi15 are the same resistance gene. For future studies, we suggest referring to this resistance with the first name that was assigned to this gene, namely Rvi4.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Apple scab, caused by the ascomycete Venturia inaequalis (Cooke) G. Winter, is the most important fungal disease in apple production in regions with frequent rain during spring (Belete and Boyraz 2017). The control of the disease in conventional, integrated, and biological production is based on the frequent application of fungicides (Zaller et al. 2023). The use of scab resistant cultivars has the potential of reducing the dependence on these products. The large majority of apple scab resistant cultivars that are currently available carry the apple scab resistance gene Rvi6 (formerly Vf) originating from the wild apple Malus floribunda 821 (Khajuria et al. 2018). Unfortunately, this resistance has been overcome by the pathogen in Europe (Parisi et al. 2003, Parisi et al. 2004; Patocchi et al. 2020) and in the USA (Beckerman et al. 2009; Papp et al. 2020). The breakdown of Rvi6 intensified the efforts of breeders to use other resistance genes and to combine them to produce cultivars with more durable resistance.
Only a limited number of commercial cultivars carrying apple scab resistance genes other than Rvi6 are available and few cultivars carrying scab resistance gene pyramids have been developed so far. Examples are ‘Reka’, which carries Rvi2; ‘Regia’, ‘Realka’, ‘Rea Agatav’, and ‘Rea Bellina’, which carry the combination of Rvi2 and Rvi4; and ‘Rea Cadis’, which carries a combination of Rvi4 and Rvi6. To date, numerous scab resistance genes have been identified, 17 of which have been included in the Rvi nomenclature system (Bus et al. 2011). A summary of the first 10 years of the VINQUEST initiative (Patocchi et al. 2020) reported that most of these single genes were overcome. Exceptions were Rvi11 (formerly Vbj) from the wild apple genotype Malus baccata jackii (Gygax et al. 2004) and Rvi15 from GMAL 2473 (alias Vr2, Patocchi et al. 2004) that were reported as not overcome. Rvi15 was considered to be the most promising resistance gene that could be incorporated relatively quickly into a new cultivar in combination with other scab resistance gene(s) for durable resistance because the fruits of its source GMAL 2473 are of much better quality (e.g., fruit size of about 6 cm measured in August, Patocchi A. unpublished) than those of Malus baccata jackii (fruit size of ca. 1.5 cm measured in August).
Rvi15 is a resistance gene that induces pin-point pits (Galli et al. 2010a). This gene has been mapped on the top of linkage group (LG) 2 (Patocchi et al. 2004) and then identified via positional cloning. Galli et al. (2010b and c) found three putative resistance genes (Vr2-A, Vr2-B, and Vr2-C) characterized by a toll and mammalian interleukin-1 receptor protein nucleotide-binding site leucine-rich repeat structure (TIR_NBS_LRR) on the BAC clone spanning the resistance region of Rvi15. The three genes Vr2-A, Vr2-B, and Vr2-C had coding sequences of 3273 bp (1091 amino acids), 4032 bp (1344 amino acids), and 3296 bp (1099 amino acids), respectively, and showed differences in all five domains, the most evident being in the length of the leucine-reach repeat domain (Galli et al. 2010c). In a subsequent study, Schouten et al. (2014) functionally demonstrated that the candidate gene Vr2-C was the gene promoting Rvi15 resistance. Recently, several Rvi15 ‘Gala Galaxy’ trans- and cisgenic lines have been developed (Giovanni Broggini personal communication).
Rvi4, named Vh4 until the nomenclature of the apple scab resistance genes was revisited in Bus et al. (2011), is a resistance gene that also induces pin-point pits (Bus et al. 2005). Interestingly, Bus et al. (2005) mapped Rvi4, which is derived from R12740-7A (alias ‘Russian seedling’, Dayton et al. 1953), to the top end of the LG 2, very closely to Rvi15, but at a different map position. In fact, in Galli et al. (2010b), Rvi15 mapped to a position 2 cM above the SSR marker CH02c02c, while Rvi4 was mapped to 5 cM below the same marker in Bus et al. (2005). Yet, the mapping of Rvi4 conducted by Bus et al. (2005) was refined in the work of Jänsch et al. (2015). Checking the original molecular data of Bus et al. (2005), genotyping mistakes were found, and their correction led to Rvi4 being remapped to 0.4 cM above CH02c02c. With this new information, amplicons originally developed for Rvi15 were searched for SNPs associated with Rvi4. Fifty-three SNPs, all co-segregating with Rvi4, were found (Jänsch et al. 2015).
With Rvi4 and Rvi15 now mapping to the same region, both inducing a hypersensitivity response (HR), and both being of particular interest for the breeders, it was necessary to definitively prove whether or not the two apple scab resistance genes are indeed the same.
To answer this question, (1) the alleles of the molecular markers linked with/in physical proximity of Rvi4 and Rvi15 amplified from Rvi4 and Rvi15 genotypes were compared, (2) part of the pedigree of GMAL 2473 was reconstructed and the haplotypes of cultivars/selections with Rvi2, Rvi4, both resistances, and ‘Russian Seedling’ were compared, (3) amplicons spanning the whole Vr2-C coding region of ‘Regia’ (Rvi2 and Rvi4) were sequenced and compared with those of GMAL 2473 (Rvi15), and (4) Rvi4 and Rvi15 genotypes were inoculated with apple scab isolates known to overcome Rvi4 resistance.
Materials and methods
Plant material and DNA extraction
GMAL 2473 used in this study was the same as used in Patocchi et al. (2004), Galli et al. (2010a), Galli et al. (2010b), Galli et al. (2010c), and Jänsch et al. (2015). It originated from the Geneva National Germplasm Repository (PI 589835). This genotype is not available anymore at the US repository. Selections 04E1-84, 04E1-860, 04E1-595, 04E1-918, and 04E1-33 are progeny of a ‘Golden Delicious’ x GMAL 2473 cross previously used by Galli et al. (2010b). These seedlings, the only still available of this cross, were used for phasing the SNP markers alleles of GMAL 2473.
The leaf tissue of ‘Regia’, TSR33T239, and GMAL 2473 that was used for the molecular marker analysis derived from the collection of genotypes of Agroscope Wädenswil (Switzerland). The leaf tissue of ‘Regia’ and GMAL 2473 that was used for the sequencing of Vr2-C genomic DNA and cDNA derived from trees of the collection of JKI Dresden-Pillnitz (Germany). The seven transgenic lines Vr 2c-603/1, Vr 2c-604/1, Vr 2c-608/1, Vr 2c-610/1, and Vr 2c-612/1 used for the scab inoculation test were those developed by Schouten et al. (2014). Control genotypes of this latter experiment were ‘Regia’, ‘Gala’, and ‘Gala’ wild type (wt) used for the generation of the transgenic lines, TSR33T239 and GMAL 2473 of the collection of JKI Dresden-Pillnitz (Germany). All the not genetically modified genotypes were grown in the field, while the transgenic lines were grown in a greenhouse under standard conditions.
Genomic DNA of field- or greenhouse-grown plants was extracted from about 100 mg of fresh leaf tissue using the DNeasy® Plant Mini Kit (Qiagen, Hilden, Germany) according to manufacturer’s instructions.
Isolates for scab inoculation tests
Three different Venturia inaequalis isolates were used in this study: 104 (race 1), 1634 (race 4), and Regia2 (race 2, 4). Isolate 104 was isolated from ‘Golden Delicious’ (Guillaumès et al. 1995), whereas isolate 1634 was isolated from TSR33T239, a genotype carrying Rvi4 (Caffier et al. 2015). Regia2 was isolated from ‘Regia’, which carries both resistance genes Rvi2 and Rvi4. The latter isolate is therefore virulent to both Rvi2 and Rvi4 (Peil et al. 2018).
Molecular marker analysis
The curated SNP array data and the sources for each accession used are listed in supplementary tables 1 to 3 (online resource 1). SNP data from the 20K SNP genotyping array (Bianco et al. 2014) was generated for some individuals in this study, made available from previous studies, or shared for use in this study for pedigree reconstruction and haplotype tracing. Data for 04E1-84, 04E1-860, 04E1-595, 04E1-918, and 04E1-33 was generated by Agroscope. SNP array data for ‘Regia’, ‘Reka’, and their common parent PRI 388-14 (Andreas Peil and Eric van de Weg unpublished, also known as OR40T43) was made available from the FruitBreedomics project (Laurens et al. 2018). SNP array data for the three offspring of ‘Russian Seedling’ were made available from the Palouse wild cider apple breeding program (James et al. 2022) and used for phasing the SNP data of ‘Russian Seedling’. The SNP array data was curated following the methods described in Vanderzande et al. (2019) using the set of 10,321 robust SNPs used in Volk et al. (2022).
Pedigree reconstruction for GMAL 2473 and pedigree validation for ‘Regia’, ‘Reka’, and their pedigree ancestors were conducted using the methods described in Vanderzande et al. (2019) and Howard et al. (2021). Parent-offspring duo relationships were ordered using the parent-offspring order resolution test (Howard et al. 2022). Candidate pedigree ancestors of GMAL 2473 were considered from a panel of over 5000 cultivars and germplasm accessions from an ongoing, large-scale collaborative pedigree reconstruction project (Howard et al. 2018a). SNP array data was phased using FlexQTL (Bink et al. 2014). Phased data for ‘Russian Seedling’, GMAL 2473, PRI 388-14, ‘Regia’, and ‘Reka’ were compared to demonstrate identity by descent over the regions of chromosome 2 containing Rvi2 and Rvi4.
The physical positions for Rvi2 and Rvi4 were estimated by BLASTing sequences associated with them against the Malus x domestica HFTH1 Genome v1.0.a1 assembly (Zhang et al. 2019). For Rvi2, the sequences (KM104993 and KM105136, respectively) containing the SNP markers FBsnRvi2-5 and FBsnRvi2-6.1, which flank Rvi2 (Jänsch et al. 2015), were used. For Rvi4/Rvi15, the primer sequence Vr2Cfull_fw was used as the position of this sequence on HFTH1 was found to be close to those of the marker FBsnRvi4.1 co-segregating with Rvi4 in Jänsch et al. (2015).
The physical coordinates retrieved were compared to the 10,321 robust SNPs on the SNP array to identify flanking SNPs to use as a focal point for building haplotypes covering Rvi2 and Rvi4 to use for haplotype tracing as described in Howard et al. (2018b).
The Rvi4 and Rvi15 molecular markers (SSR and KASP™) listed in supplementary tables 4 and 5 (online resource 2) were tested on DNA of the genotypes GMAL 2473 (Rvi15), ‘Regia’ (Rvi2 and Rvi4) and ‘Gala’. ‘Gala’ was included to support the transfer of the SSR markers in other laboratories (correction of possible shift of the allele sizes).
SSR analyses were conducted at Ecogenics GmbH (Balgach, Switzerland), while SNP genotyping using the competitive allele specific PCR genotyping system (KASP™) was conducted at LGC Genomics Ltd. (Teddington, UK).
Sequencing of Vr2-C genomic DNA and cDNA from Rvi4 and Rvi15 genotypes
The primers Vr2C_full_fw and Vr2C_full_II_rev were used for full-length amplification of Vr2-C from ‘Regia’ and GMAL 2473. The PCR was performed in 1× Long Range PCR buffer containing 10 ng template DNA, 5 μl dNTP’s (0.2 mM of each), 2.5 μl of each primer (10 μm), 0.25 μl Long Range PCR Enzyme Mix (Thermo Fisher Scientific, Erlangen, Germany), and ddH2O in a total volume of 50 μl. After denaturation for 3 min at 94 °C, 10 PCR cycles were performed with 20 s at 94 °C, 30 s at 70 °C, and 6 min at 68 °C. Subsequently, 25 PCR cycles were performed with 20 s at 94 °C, 30 s at 70 °C, and 6 min at 68 °C with a time increment of 5 s per cycle. After final extension at 68 °C for 10 min, the reaction was stored at 10 °C.
Vr2-C cDNA from ‘Regia’ and GMAL 2473 was synthetized as follows: young leaf tissue was homogenized using the Retsch Mixer Mill MM400 (Retsch, Haan, Germany). The total RNA was extracted using the InviTrap Spin Plant RNA Mini Kit (Stratec Molecular GmbH, Berlin, Germany). Genomic DNA contaminations were removed with the DNA-Free Kit (Life Technologies GmbH, Darmstadt, Germany). For cDNA synthesis, the RevertAid™ First Strand cDNA Synthesis Kit (Thermo Scientific, Braunschweig, Germany) was used and 1 μg of total RNA as template. RT-PCR for detecting full-length Vr2C transcripts was performed using the primers Vr2C_full_fw and Vr2C_full_II_rev.
Amplified RT-PCR products were separated on a 0.8–1.0% agarose gels, isolated and ligated into the vector PCRTM2.1-TOPOTM (Invitrogen, Groningen, The Netherlands), and transferred into One ShotTM cells according to the manual. Plasmids for sequencing were isolated with the QIAGEN Plasmid Mini Kit (QIAGEN, Hilden, Germany). Sequencing was performed at Eurofins Genomics (Ebersberg, Germany) using the sequencing primers listed in Table 1.
Scab inoculation
Conidia for inoculation were propagated as described by Peil et al. (2018) with small modifications for isolates 104 and 1634. Isolates 104 and 1634 were additionally propagated on plates containing 0.75% malt extract. Conidia were washed from the cellophane membranes and diluted with water to a concentration of 1.5–2.5 × 105 conidia/ml. Up to five replicates per genotype were produced by grafting budwood on the M9 rootstock (online resource 4). Plants were grown in a ventilated greenhouse (18°C day/15 °C night) for about 9 weeks. The youngest leaves of actively growing shoots were spray inoculated as described in Peil et al. (2018). Three replicates of each genotype were inoculated with isolate 104. Five replicates of each transgenic line and GMAL2473 and one replicate of ‘Regia’ were inoculated with isolate 1634. Five replicates of each transgenic line and ‘Regia’ and three replicates of GMAL2473 were inoculated with Regia2. Symptoms were scored after 21 and 28 day post-inoculation (dpi) according to (Chevalier et al. 1991). Score values were as follows: 0—no symptoms, 1—pin point pits, 2—chlorosis, 3a—necrotic and some chlorotic lesions with very slight occasional sporulation, 3b—clearly sporulating chlorotic and necrotic lesions, and 4—sporulating lesions.
Results
Molecular marker analysis
SSR and SNP molecular markers previously reported to be associated with the resistance genes Rvi4 and Rvi15 have been tested on ‘Regia’ and GMAL 2473. The results for ‘Regia’ and GMAL 2473 were identical. In fact, both genotypes amplified with all markers the allele in coupling with the Rvi4 or Rvi15 resistance (Table 2).
Pedigree reconstruction of GMAL 2473
GMAL 2473 and ‘Russian Seedling’ shared at least one allele at every locus, indicating that they have a parent-offspring relationship. ‘Russian Seedling’ was confirmed as the parent of GMAL 2473 via the parent-offspring order resolution test using the five offspring of GMAL 2473 (04E1_33, 04E1_595, 04E1_84, 04E1_860, and 04E1_918) that were included in this study. Roughly 30% of the haplotypes from the unknown parent of GMAL 2473 are identical to those in ‘Duchess of Oldenburg’, thus indicating ‘Duchess of Oldenburg’ as a likely grandparent of the unknown parent of GMAL 2473. This result provided further confirmation that GMAL 2473 was an offspring rather than the parent of ‘Russian Seedling’ because there were not haplotypes of ‘Duchess of Oldenburg’ in ‘Russian Seedling’, which would have been expected if the opposite relationship were true.
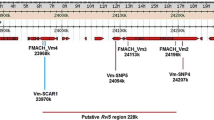
GMAL 2473 inherited the haplotype and the corresponding region harboring Rvi4/15 from ‘Russian Seedling’ but did not inherit Rvi2 due to a close recombination (Fig. 1). PRI 388-14 inherited both haplotypes containing the regions of Rvi2 and Rvi4 through its ungenotyped parent PRI 45-48, which matches its recorded pedigree of ‘Delicious’ x ‘Russian Seedling’. ‘Regia’ and ‘Reka’ inherited the haplotypes containing the region of Rvi2 from PRI 388-14, though only ‘Regia’ inherited the region containing Rvi4.

A Pedigree of descendants of ‘Russian Seedling’ and B depiction of chromosome 2 haplotypes of selected descendants of ‘Russian Seedling’, with the estimated locations of haplotypes containing the scab resistance genes Rvi2 (pink) and Rvi4 (green) highlighted
Sequencing of Vr2-C genomic DNA and cDNA of ‘Regia’
Full-length genomic DNA sequences of Vr2-C were amplified (amplicon of ~5.500 bp) from genomic DNA of GMAL 2473 and ‘Regia’ (amplicon of ~5.500 bp). The PCR product of ‘Regia’ was cloned and sequenced. The 5.594 bp long sequence (GeneBank: OR220343) was aligned to the sequence of the BAC clone 32A4 of GMAL 2473 (GenBank: KF055410.1). Both sequences were nearly identical. Differences were found at three different positions. Two nucleotides (AT) in a microsatellite repeat were missing (10 vs 11 AT repeats) in the sequence obtained from ‘Regia’ at position 2322–2323. Two nucleotides were missing in a region with a repetition of T (10 vs 12) at position 3706–3707, and one T was missing in a region with a repetition of T (7 vs 8) at position 4143 (Suppl. Fig. 1).
The amplicons of the full length coding sequence (CDS) of Vr2-C obtained from ‘Regia’ and GMAL 2473 had also the same size (data not shown). The PCR products were cloned, and eight clones of ‘Regia’ and six clones of GMAL 2473 were sequenced. Six clones of ‘Regia’ and four clones of GMAL 2473 were nearly identical. Two clones per genotype were slightly different. These clones appeared to be other splice variants. The sequences of the six ‘Regia’ clones and the four GMAL 2473 clones were aligned separately and consensus sequences for both genotypes were generated. The consensus CDS sequences (GeneBank: OR220341 and OR220342) were both 3.366 bp long and 100% identical. Subsequently, these CDS sequences were aligned to the GMAL 2473 BAC 32A4 sequence (GenBank: KF055410.1), the genomic Vr2-C sequence of ‘Regia’ and the predicted Vr2-C CDS sequence published by Galli et al. (2010a). Both CDS sequences (‘Regia’ and GMAL 2473) matched perfectly to the BAC 32A4 sequence. However, some differences were found to the predicted Vr2-C CDS of Galli et al. (2010b). The predicted Vr2-C CDS has a length of 3.288 bp and consists of eight exons and seven introns. The CDS sequences of ‘Regia’ and GMAL 2473 obtained in this study have a length of 3.366 bp and consist of six exons and five introns (Suppl. Fig. 1).
Scab inoculation results
All three tested scab isolates proved to be pathogenic as they infected ‘Regia’ and/or the apple genotype they were isolated from (Table 3, Suppl. Table 6, online resource 4). Isolate 104 (race 1), originating from ‘Golden delicious’ showed an incompatible reaction and induced a HR in GMAL 2473 and in all the Vr2-C transgenic lines. A compatible interaction (i.e., strong sporulation) was observed between isolate 1634 (race 4) and TSR33T239, as well as with all the Vr2-C transgenic lines (Fig. 2). Only ‘Regia’ and GMAL 2473 were incompatible with isolate 1634. Isolate Regia2 (race 2, 4) was compatible with all the tested genotypes with the exception of GMAL 2473, which showed chlorotic lesions. ‘Regia’

Inoculation results of the Vr2-C transgenic line 612/1, TSR33T239, and GMAL 2473 with the scab isolate 1634 28 dpi. Vr2-C-612/1 and TSR33T239 show strong apple scab sporulation while GMAL 2473 did not show any symptoms
Discussion
All the results produced in this work indicate that Rvi4 and Rvi15 are the same gene, the definitive evidence being the CDS of Vr2-C found in ‘Regia’, the Vr2-C transgenic lines losing their resistance when inoculated with isolates overcoming Rvi4 resistance, the identification of ‘Russian Seedling’ (source of Rvi4) in the pedigree of GMAL 2473, and the demonstrated inheritance of haplotypes containing Rvi4 in both ‘Russian Seedling’ and GMAL 2473.
Since the first mapping of Rvi4 and Rvi15 to similar positions on the top of LG2, and because both genes induced the same resistance reaction type (a rapid hypersensitive response), it had been suspected that the two genes might be the same. This presumption found first confirmations when Jänsch et al. (2015) discovered that both genes mapped in the same region of LG2, and when performing marker-assisted selection (MAS) for both genes, it was discovered that Rvi4 and Rvi15 genotypes amplified the same alleles in coupling with the two resistance genes (Table 2). These findings motivated us to definitively clarify whether the two genes were actually the same.
The reconstruction of the pedigree of GMAL 2473 and those of ‘Regia’ and ‘Reka’ showed that both Rvi4 and Rvi15 originated from the same source, i.e., ‘Russian seedling’, and that Rvi2 and Rvi4 are each located on the two different homolog chromosomes 2 in ‘Russian Seedling’. PRI 45-48, a direct offspring of ‘Russian Seedling’ and grandparent of ‘Regia’, inherited a recombinant chromosome 2 from ‘Russian Seedling’, resulting in PRI 45-48 having both resistances, i.e., Rvi2 and 4 (Fig. 1A). PRI 45-48 transmitted most of this homolog, including both resistances, to PRI 388-14, which it then transmitted in full to its offspring ‘Regia’. However, since the two genes are relatively far apart (about 40 cM, estimated from the work of Bus et al. (2005) and about 51 cM in the iGL map used in this study for haplotype tracing), it is not surprising that ‘Reka’ inherited a recombinant homolog from PRI 388-14 that only contained Rvi2.
While the comparison of the full length CDS of Vr2-C of ‘Regia’ and GMAL 2473 showed no differences between each other and the genomic sequence of the BAC 32A4, differences of few nucleotides in three regions were observed between this sequence and the CDS predicted by Galli et al. (2010c). Differences between computer-calculated and experimentally studied CDSs occasionally occur (Vogt et al. 2013), and it is known that sequencing of transcripts usually improves the prediction (Minoche et al. 2015). Sequence differences were also observed while comparing the sequence of Vr2-C obtained from the BAC clone 32A4 and the consensus sequence of the cloned amplicons of Vr2-C from ‘Regia’. The three observed differences were all found in sequences known to be prone to sequences mistakes, i.e., microsatellites and relative long sequences of the same nucleotide. Moreover, these differences were observed in intronic regions, and therefore, they would not affect the function of the resistance gene. On the other hand, if real, these differences may be the results of mutations starting to accumulate over the generations.
The results of the scab inoculations done with race 1 (104) and race 4 (1634, Regia2) isolates clearly showed that all the genotypes carrying Rvi4 and Rvi15 were resistant (incompatible) to the race 1 isolate and susceptible (compatible) to race 4 isolates, which definitively confirmed that Rvi4 and Rvi15 are the same gene. One exception is GMAL 2473, that is resistant also to the race 4 isolates. This latter result is in line with the observations made in the context of the monitoring of the apple scab resistance gene breakdown of the VINQUEST initiative (Patocchi et al. 2020, http://www.vinquest.ch). In fact, so far, no compatible interaction on GMAL 2473 was observed over all the observation test sites, while for Rvi4 compatible interactions have been observed in six test sites (in general, only few scab lesions found, Patocchi et al. 2020). Also, a study testing the differential hosts of VINQUEST with a set of different isolates (Caffier et al. 2015) reported that the differential host h(15) (GMAL 2473) was incompatible with all the isolates tested, while the differential host h(4) (TSR33T239) was compatible to three isolates, including 1634. This difference could be due to a second (yet unknown) resistance locus possibly present in GMAL 2473. The presence of this second resistance locus, possibly inducing partial resistance (QTL), may explain why Patocchi et al. (2004) could not define whether the 337 progeny of a cross of “Idared” x GMAL 2473 segregate for one or two resistance genes. In contrast, Galli et al. (2010b), studying the segregation between susceptible and resistant progeny plants of a ‘Golden Delicious’ x GMAL 2473 cross, clearly found a 1:1 ratio. One hypothesis explaining these contrasting results could be the different inoculum used; those of the second study being able to overcome the second unknown resistance factor, while this was not the case for the inoculum used for the first study.
Concluding, this study demonstrated that Rvi4 and Rvi15 are the same apple scab resistance gene. The two names Rvi4 and Rvi15 now refer to same resistance gene, but a single name should be used in the future. We propose to use Rvi4, as this name was the first assigned to this resistance and because it is probably more well known in apple breeding than Rvi15.
Data availability
All the data supporting the results have been deposited in GeneBank (OR220341, OR220342 and OR220343) or can be found in the Supplementary information.
References
Beckerman J, Chatfield J, Draper E (2009) A 33-year evaluation of resistance and pathogenicity in the apple scab–crabapples pathosystem. HortScience 44(3):599–608. https://doi.org/10.21273/HORTSCI.44.3.599
Belete T, Boyraz N (2017) Critical review on apple scab (Venturia inaequalis) biology, epidemiology, economic importance, management and defense mechanisms to the causal agent. J Plant Physiol Pathol 5(2):2. https://doi.org/10.4172/2329-955X.1000166
Bianco L, Cestaro A, Sargent DJ, Banchi E, Derdak S, Di Guardo M, Salvi S, Jansen J, Viola R, Gut I (2014) Development and validation of a 20K single nucleotide polymorphism (SNP) whole genome genotyping array for apple (Malus ×domestica Borkh). PloS One 9(10):e110377. https://doi.org/10.1371/journal.pone.0110377
Bink MCAM, Jansen J, Madduri M, Voorrips RE, Durel CE, Kouassi AB, Laurens F, Mathis F, Gessler C, Gobbin D, Rezzonico F, Patocchi A, Kellerhals M, Boudichevskaia A, Dunemann F, Peil A, Nowicka A, Lata B, Stankiewicz-Kosyl M et al (2014) Bayesian QTL analyses using pedigreed families of an outcrossing species, with application to fruit firmness in apple. Theor Appl Genet 127(5):1073–1090. https://doi.org/10.1007/s00122-014-2281-3
Bus VGM, Rikkerink EHA, Caffier V, Durel CE, Plummers KM (2011) Revision of nomenclature of the differential host-pathogen interactions of Venturia inaequalis and Malus. Annu Rev Phytopathol 49:391–413. https://doi.org/10.1146/annurev-phyto-072910-095339
Bus VGM, Rikkerink EHA, van de Weg WE, Rusholme RL, Gardiner SE, Bassett HCM, Kodde LP, Parisi L, Laurens FND, Meulenbroek EJ, Plummer KM (2005) The Vh2 and Vh4 scab resistance genes in two differential hosts derived from Russian apple R12740-7A map to the same linkage group of apple. Mol Breed 15(1):103–116. https://doi.org/10.1007/s11032-004-3609-5
Caffier V, Patocchi A, Expert P, Bellanger MN, Durel CE, Hilber-Bodmer M, Broggini GAL, Groenwold R, Bus VGM (2015) Virulence characterization of Venturia inaequlis reference isolates on the differential set of Malus hosts. Plant Dis 99(3):370–375. https://doi.org/10.1094/Pdis-07-14-0708-Re
Chevalier M, Lespinasse Y, Renaudin S (1991) A microscopic study of the different classes of symptoms coded by the Vf gene in apple for resistance to scab (Venturia inaequalis). Plant Pathol 40(2):249–256. https://doi.org/10.1111/j.1365-3059.1991.tb02374.x
Dayton DF, Shay JR, Hough LF (1953) Apple scab resistance from R12740-7a, a Russian apple. Proc Am Soc Hortic Sci 62:334–340
Galli P, Broggini GAL, Gessler C, Patocchi A (2010a) Phenotypic characterization of the Rvi15 (Vr2) apple scab resistance. J Plant Pathol 92(1):219–226 https://www.jstor.org/stable/41998787
Galli P, Broggini GAL, Kellerhals M, Gessler C, Patocchi A (2010b) High-resolution genetic map of the Rvi15 (Vr2) apple scab resistance locus. Mol Breed 26(4):561–572. https://doi.org/10.1007/s11032-010-9391-7
Galli P, Patocchi A, Broggini GAL, Gessler C (2010c) The Rvi15 (Vr2) apple scab resistance locus contains three TIR-NBS-LRR genes. Mol Plant Microbe Interact 23(5):608–617. https://doi.org/10.1094/Mpmi-23-5-0608
Guillaumès J, Chevalier M, Parisi L (1995) Studies on the relationship between Venturia inaequalis and Malus x domestica in vitro plants. Can J Plant Pathol-Revue Canadienne de Phytopathologie 17(4):305–311
Gygax M, Gianfranceschi L, Liebhard R, Kellerhals M, Gessler C, Patocchi A (2004) Molecular markers linked to the apple scab resistance gene Vbj derived from Malus baccata jackii. Theor Appl Genet 109(8):1702–1709. https://doi.org/10.1007/s00122-004-1803-9
Howard NP, Albach DC, Luby JJ (2018a) The identification of apple pedigree information on a large set of apple germplasm and its application in apple breeding using new genetic tools. In: ecofruit, pp 88–91
Howard NP, Albach DC, Luby JJ (2018) The identification of apple pedigree information on a large diverse set of apple germplasm and its application in apple breeding using new genetic tools. In: Proc. 18th International Conference on Organic Fruit Growing. Foerdergemeinschaft Oekologischer Obstbau e. V.(FOEKO), Hohenheim, Germany, pp 88–91. www.ecofruit.net/wp-content/uploads/2020/04/2_Howard_88-91.pdf
Howard NP, Van De Weg E, Luby JJ (2022) A new method to reconstruct the direction of parent-offspring duo relationships using SNP array data and its demonstration on ancient and modern cultivars in the outcrossing species Malus× domestica. Hortic Res 9. https://doi.org/10.1093/hr/uhab069
Howard NP, van de Weg E, Tillman J, Tong CBS, Silverstein KAT, Luby JJ (2018b) QTL characterized for soft scald and soggy breakdown in apple (Malus × domestica) through pedigree-based analysis of a large population of interconnected families. Tree Genet Genomes 14(2):1–16. https://doi.org/10.1007/s11295-017-1216-y
Jänsch M, Broggini GA, Weger J, Bus VG, Gardiner SE, Bassett H, Patocchi A (2015) Identification of SNPs linked to eight apple disease resistance loci. Mol Breed 35(1):45. https://doi.org/10.1007/s11032-015-0242-4
James T, Johnson A, Schaller A, Vanderzande S, Luo F, Sandefur P, Ru S, Peace C (2022) As it stands: the palouse wild cider apple breeding program. Plants 11:517–534. https://doi.org/10.3390/plants11040517
Khajuria YP, Kaul S, Wani AA, Dhar MK (2018) Genetics of resistance in apple against Venturia inaequalis (Wint.) Cke. Tree Genet Genomes 14(2):16. https://doi.org/10.1007/s11295-018-1226-4
Laurens F, Aranzana MJ, Arus P, Bassi D, Bink M, Bonany J, Caprera A, Corelli-Grappadelli L, Costes E, Durel C-E (2018) An integrated approach for increasing breeding efficiency in apple and peach in Europe. Hortic Res 5(1):11. https://doi.org/10.1038/s41438-018-0016-3
Liebhard R, Gianfranceschi L, Koller B, Ryder CD, Tarchini R, Van de Weg E, Gessler C (2002) Development and characterisation of 140 new microsatellites in apple (Malus x domestica Borkh.). Mol Breed 10(4):217–241. https://doi.org/10.1023/A:1020525906332
Minoche AE, Dohm JC, Schneider J, Holtgräwe D, Viehöver P, Montfort M, Sörensen TR, Weisshaar B, Himmelbauer H (2015) Exploiting single-molecule transcript sequencing for eukaryotic gene prediction. Genome Biol 16:184. https://doi.org/10.1186/s13059-015-0729-7
Papp D, Singh J, Gadoury D, Khan A (2020) New North American isolates of Venturia inaequalis can overcome apple scab resistance of Malus floribunda 821. Plant Dis 104(3):649–655. https://doi.org/10.1094/PDIS-10-19-2082-RE
Parisi L, Fouillet V, Schouten H, Groenwold R, Laurens F, Didelot F, Evans K, Fischer C, Gennari F, Kemp H (2004) Variability of the pathogenicity of Venturia inaequalis in Europe. Acta Hortic 663(1):107–113. https://doi.org/10.17660/ActaHortic.2004.663.13
Parisi L, Lespinasse Y, Guillaumes J, Kruger J (1993) A new race of Venturia inaequalis virulent to apples with resistance due to the Vf gene. Phytopathology 83(5):533–537
Patocchi A, Bigler B, Koller B, Kellerhals M, Gessler C (2004) Vr2: a new apple scab resistance gene. Theor Appl Genet 109(5):1087–1092. https://doi.org/10.1007/s00122-004-1723-8
Patocchi A, Wehrli A, Dubuis P-H, Auwerkerken A, Leida C, Cipriani G, Passey T, Staples M, Didelot F, Philion V, Peil A, Laszakovits H, Ruehmer T, Boeck K, Baniulis D, Strasset K, Vávra R, Guerra W, Masny S et al (2020) Ten years of VINQUEST: first insight for breeding new apple cultivars with durable apple scab resistance. Plant Dis 104(8):2074–2081. https://doi.org/10.1094/PDIS-11-19-2473-SR
Peil A, Patocchi A, Hanke M-V, Bus VG (2018) Apple cultivar Regia possessing both Rvi2 and Rvi4 resistance genes is the source of a new race of Venturia inaequalis. Eur J Plant Pathol 151:533–539. https://doi.org/10.1007/s10658-017-1383-6
Schouten HJ, Brinkhuis J, van der Burgh A, Schaart JG, Groenwold R, Giovanni A, Broggini L, Gessler C (2014) Cloning and functional characterization of the Rvi15 (Vr2) gene for apple scab resistance. Tree Genet Genomes 10(2):251. https://doi.org/10.1007/s11295-013-0678-9
Silfverberg-Dilworth E, Matasci CL, Van de Weg E, Van Kaauwen MPW, Walser M, Kodde LP, Gianfranceschi L, Durel C-E, Costa F, Yamamoto T, Koller B, Gessler C, Patocchi A (2006) Microsatellite markers spanning the apple (Malus x domestica Borkh) genome. Tree Genet Genomes 2:202–224. https://doi.org/10.1007/s11295-006-0045-1
Vanderzande S, Howard NP, Cai L, Linge CDS, Antanaviciute L, Bink MC, Kruisselbrink JW, Bassil N, Gasic K, Iezzoni A (2019) High-quality, genome-wide SNP genotypic data for pedigreed germplasm of the diploid outbreeding species apple, peach, and sweet cherry through a common workflow. PloS One 14(6). https://doi.org/10.1371/journal.pone.0210928
Vogt J, Schiller D, Ulrich D, Schwab W, Dunemann F (2013) Identification of lipoxygenase (LOX) genes putatively involved in fruit flavour formation in apple (Malus × domestica). Tree Genet Genomes 9:1493–1511. https://doi.org/10.1007/s11295-013-0653-5
Volk GM, Peace CP, Henk AD, Howard NP (2022) DNA profiling with the 20K apple SNP array reveals Malus domestica hybridization and admixture in M. sieversii, M. orientalis, and M. sylvestris genebank accessions. Frontiers in. Plant Sci 13. https://doi.org/10.3389/fpls.2022.1015658
Zaller JG, Oswald A, Wildenberg M, Burtscher-Schaden H, Nadeem I, Formayer H, Paredes D (2023) Potential to reduce pesticides in intensive apple production through management practices could be challenged by climatic extremes. Sci Total Environ 872:162237. https://doi.org/10.1016/j.scitotenv.2023.162237
Zhang L, Hu J, Han X, Li J, Gao Y, Richards CM, Zhang C, Tian Y, Liu G, Gul H, Wang D, Tian Y, Yang C, Meng M, Yuan G, Kang G, Wu Y, Wang K, Zhang H et al (2019) A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour. Nat Commun 10(1):1494. https://doi.org/10.1038/s41467-019-09518-x
Acknowledgements
We thank Gisela Schulz and Ines Polster for taking care of the experimental plants in the greenhouse, and Cameron Peace and the Palouse wild cider apple breeding program for sharing data used for phasing ‘Russian Seedling’.
Funding
Open access funding provided by Agroscope
Author information
Authors and Affiliations
Contributions
Conceptualization: all authors; methodology: all authors; formal analysis and investigation: APeil, NPH., SB-S, IH, and HF; writing—original draft preparation: APatocchi, HFH, APeil; writing—review and editing: all authors; funding acquisition: nobody; resources: nobody; supervision: APatocchi, HFH, APeil, NPH.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
Supplementary Table 1 Apple genotypic profiles names (Analysis_Name), representative accessions (Accession_name) and associated accession IDs (Accession_ID) genotyped on the Illumina apple Infinium® 20K SNP array with the indicated genotypic profiles, the source of the SNP data (Data_source), and the collection site of the accessions sampled that were used for haplotype analysis and phasing in this study. Online Resource 1. Supplementary Table 2. Curated unphased SNP array data for genotypic profiles listed in Suppl. Table 1. Columns with numbers are the SNP index numbers from the Illumina apple Infinium® 20K SNP array. Online Resource 1. Supplementary Table 3. Curated phased SNP array data for genotypic profiles listed in Suppl. Table 1. Columns with numbers are the SNP index numbers from the Illumina apple Infinium® 20K SNP array. The column "VAR" denotes whether the haplotype data comes from the first (hap1) or second (hap2) parent. Online Resource 1. (XLSX 1926 kb)
ESM 2
Supplementary Table 4. Primers of SSR markers used for MAS for the genes Rvi4 and Rvi15. Online Resource 2. Supplementary Table 5. Primers of the KASP assays used for MAS for the genes Rvi4 and Rvi15. Online Resource 2. (PDF 163 kb)
ESM 3
Supplementary Table 6. Results of the artificial scab inoculation of the single replicates of the genotypes previously reported to carry Rvi4 or Rvi15 or a combination of both genes. 'Gala' was used as susceptible control. Online Resource 4. (PDF 9635 kb)
ESM 4
Supplementary figure 1. Sequence alignment of genomic and CDS sequences of Rvi15 of 'Regia' and GMAL 2473. Rvi15_CDS_GMAL and Rvi15_CDS_Regia, sequences amplified from cDNA of 'Regia' and GMAL 2473 in this study, Rvi15_CDS_GMAL_Galli, CDS of Rvi15 predicted by Galli et al. (2010c), Rvi15_genomic_Regia, genomic Rvi15 DNA sequence of 'Regia' amplified and sequenced in this study, Rvi15_genomic_BAC32A4, and genomic DNA sequence of Rvi15 obtained from the sequence of BAC32A4 (Galli et al. 2010a). Online Resource 3. (XLSX 14 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peil, A., Howard, N.P., Bühlmann-Schütz, S. et al. Rvi4 and Rvi15 are the same apple scab resistance genes. Mol Breeding 43, 74 (2023). https://doi.org/10.1007/s11032-023-01421-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-023-01421-0