
Abstract
Sapium sebiferum (L.) Roxb. [S. sebiferum] is one of the most important woody oil trees and traditional herbal medicines in China. Diacylglycerol acyltransferases (DGATs) esterify sn-1,2-diacylglycerol with a long-chain fatty acyl-CoA acting as a key enzyme at the last and rate-limiting step of triacylglycerol (TAG) biosynthesis in eukaryotic organisms. Although at least 61 DGAT1 sequences from 48 organisms have been identified, until now there have been almost no reports on the DGAT1 gene in S. sebiferum (SsDGAT1). In an attempt to clarify SsDGAT1’s function, we cloned the CDS (coding sequence) of SsDGAT1 by RACE (rapid amplification of cDNA ends) technology. The full-length CDS of SsDGAT1 contains 1524 bp, encoding a protein of 507 amino acids. Even recombinant SsDGAT1 was able to restore TAG biosynthesis in the yeast strain S. cerevisiae H1246 TAG-deficient mutant. Moreover, when SsDGAT1 was placed under the control of 35S promoter in Brassica napus L. W10, the total oil content of transgenic rape plants showed an increase of 12.3–14.7 % compared with the wild type. In addition, the transgenic rapeseed with heterologous expression of SsDGAT1 showed an alteration in seed oil composition overall: a significant decrease in oleic acid levels but a tendency towards an increase in linolenic acid levels. The result confirmed that SsDGAT1 may be involved in flux control of oil biosynthesis and could be used specifically to manipulate and improve oil content and composition in plants. These experimental findings suggest that we might be able to develop a plant high in industrial oils by over-expression of SsDGAT1 in S. sebiferum.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
At present, as the conflict between ever-increasing consumption of fossil fuels and rapidly dwindling resources worsens every year, there is panic about the scarcity of fuel and therefore more and more calls to seek other alternative sources of fuel in the future. Triacylglycerols (TAGs) occur largely in oil-rich plants as oils, so the TAGs of oil-rich plants are being considered as a possible renewable source of fuels in the form of biodiesel (Cahoon et al. 2007; Durrett et al. 2008; Yang et al. 2015). The added advantage of biofuels is less emission of greenhouse gases (Yang et al. 2015). However, traditional TAG-oil-rich plants for biodiesel such as soybean, rapeseed and sunflower are also food oil crops (Yang et al. 2015). Owing to the importance and economics of food oil, the ideal crop source for biodiesel products should be non-edible and grown on waste-agricultural lands. Therefore, oil-rich plants such as Sapium sebiferum (L.) Roxb. (Euphorbiaceae) [S. sebiferum] seem to be a better source for biodiesel products.
S. sebiferum is one of the most important perennial woody trees. In China, S. sebiferum has been cultivated for over 1400 years for seed oil and traditional herbal medicines (Peng et al. 2008). Moreover, Yang et al. (2007) reported that the seeds of S. sebiferum contain many kinds of fatty acids, such as oleic acid (15.12 %), linoleic acid (31.60 %), caproleic acid (2.70 %), linolenic acid (44.19 %) and palmitic acid (6.38 %). The parameters of S. sebiferum oil such as the cetane number, kinematics of viscosity and exhaust emission meet the requirements of biodiesel production (Divi et al. 2016). A previous study showed that chemical component of caproleic acid is 2t,4c-decadienoic acid (10:2Δ2t, 4c, stillingic acid), which belongs to the conjugated fatty acids (Yang et al. 2015).
The terminal enzyme of the Kennedy pathway, diacylglycerol acyltransferase (DGAT; EC 3.2.1.20), is one of the rate-limiting steps in storage lipid accumulation in plants and plays a central role in diverting fatty acid flux towards the formation of TAGs (He et al. 2004; Jako et al. 2001; Lung and Weselake 2006; Yu et al. 2006, 2008). DGATs are generally localized in the endoplasmic reticulum, although a cytosolic form has been identified in peanuts (Saha et al. 2006). At least two different types of DGAT gene families (DGAT1 and DGAT2) that differ considerably in sequence have been reported, and play non-redundant roles in TAG biosynthesis (Kroon et al. 2006; Shockey et al. 2006). These two forms of the DGAT enzyme have species-specific preferences for TAG production and accumulation during seed development. He et al.’s results indicated that DGAT1 plays a dominant role in ricinoleic acid production in castor bean (2004), whereas the data of Kroon et al. (2006) suggested that DGAT2 is the major contributor to TAG formation in castor seed oil. On the other hand, Shockey et al.’s study (2006) indicated that DGAT2 rather than DGAT1 plays a key role in TAG accumulation in developing seeds of the tung tree. Moreover, DGAT3 was also identified in some plants and fungi, such as peanut (Arachis hypogaea) (Saha et al. 2006), Arabidopsis (Bouvier et al. 2000; Hernández et al. 2012), Vernicia fordii (Cao et al. 2013) and Phaeodactylum tricornutum (Cui et al. 2013). There was even a study suggesting that DGAT3 could play a role in an alternative pathway for TAG synthesis during pxa1 seedling establishment (Hernández et al. 2012). Analysis of publicly available Affymetrix data revealed that DGAT3 expressed throughout seed development and seedling establishment in Arabidopsis (Hernández et al. 2012).
Much previous research has indicated that manipulation of DGAT1 has a considerable effect on plant oil development and its composition. On the one hand, there was deficient DGAT activity, less oil accumulated and decreased TAG/DAG ratio in Arabidopsis lines lacking DGAT1 (Katavic et al. 1995; Routaboul et al. 1999; Zou et al. 1999), but decreased seed oil content and increased protein and carbohydrate in RNAi-mediated silencing of DGAT1 in tobacco (Zhang et al. 2005). On the other hand, the oil levels of Arabidopsis, tobacco and Brassica napus L. were increased in DGAT1 over-expression lines (Jako et al. 2001; Weselake et al. 2008; Andrianov et al. 2010).
PDAT1 (phospholipid:diacylglycerol acyltransferase) is another key enzyme which makes TAGs independent of the Kennedy pathway and is involved in TAG biosynthesis in plants (Zhang et al. 2009). Furthermore, PDAT1 may have overlapping functions with DGAT1 for normal viable pollen and seed development in Arabidopsis (Zhang et al. 2009). However, a more recent study suggested that DGAT2 and PDAT play critical roles in epoxy and hydroxy fatty acid accumulation in seed oils of source plants, while DGAT1 appears to be a key enzyme for oil accumulation in all plants that have been studied, especially in Arabidopsis and soybean (Li et al. 2010). At the same time, the accumulation of total hydroxy fatty acids was significantly enhanced up to 25 % in castor-like oil in transgenic Arabidopsis seeds heterologously expressing PDAT1-2 of castor bean (Kim et al. 2011).
Although the function of DGAT1 has recently been well studied in some species, there has been no reported investigation of SsDGAT1 in S. sebiferum until now. In this paper, the annotation of gene structure for SsDGAT1 from S. sebiferum has been carried out based on clone data. In addition, gene function analysis of SsDGAT1 will provide an excellent reference for the molecular characterization of SsDGAT.
Materials and methods
Plant material
Developing seeds of S. sebiferum were taken from the tree grown in the fields of Central South University of Forestry and Technology and collected at ~15-day intervals from the day after pollination (DAP) to seed fully matured, and all stages of these seeds were used for RNA isolation. For investigating the expression of SsDGAT1, tissues such as the root, stem and leaf were harvested from 30-day-old seedlings and flower materials were acquired from adult plant. All tissues mentioned above were frozen in liquid nitrogen and stored at −80 °C until further experiment.
RNA preparation and reverse transcription
RNA was extracted from various samples using a RNA purificatiob kit (Invitrogen™, USA) according to the manufacturer’s protocol. To synthesize complementary DNA (cDNA) from isolated RNA, reverse transcription was performed using a reverse transcription kit (MBI, Lithuania) following the manufacturer’s instructions.
Isolation of the conserved region and full-length cDNA of SsDGAT1
For amplifying homologous region sequences of SsDGAT1, the second-step PCR amplification was performed using primers SsDGAT1-Fc [5′-AGTCATGC(A/G)GGTCT(C/G)TT(T/C) AA(C/T)TGTATA-3′, forward]/SsDGAT1-Rc [5′-GCC(C/T)GTTCA(C/T)A(G/A)TGG ATGGT (C/T)CGCGAT-3′, reverse], which were designed based on multiple amino acid sequence alignment of DGAT1 homologs from plants of interest (Fig. 1b), such as Vernicia fordii (GenBank accession No. DQ356680.1), Jatropha curcas (GenBank accession No. ABB84383.1), Ricinus communis (GenBank accession No. XP_002514132.1) and Camellia oleifera (GenBank accession No. JF264900). The PCR cycling parameters were 35 cycles with denaturation for 30 s at 94 °C, annealing for 30 s at 60 °C, and extension for 5 min at 72 °C. The PCR reaction mix comprised of 10× PCR buffer, first-strand cDNA, 10 μmol of each primer pair, 20 mM dNTP mix and Taq polymerase (Takara, Dalian, China) in a total volume of 20 μL. The PCR product was sub-cloned into pGEM®-T Easy vector (Promega, USA) and confirmed by DNA sequencing. In order to obtain 5′ and 3′ residue sequences of SsDGAT1 from S. sebiferum, the RACE clone was adopted with four pairs of degenerate primers (F1: 5′-ACTTCATGGTTGCTCCCACCCTATGT-3′, R1: 5′-CTAATA CGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3′, F2: 5′-CGAATGGATGCA GTGCGAGGATAAC-3′, R2: 5′-CTAATACGACTCACTATAGGGC-3′, F3: 5′-AGCCAAG TTATCCTCGCACTGCATCCA-3′, R3: 5′-ACTATAGGGCAAGCAGTGG TATCAACGCAGA GT-3′, F4: 5′-CTTCATGGTTGCTCCCACCCTATGTTA-3′, R4: 5′-CTAATACGACTCACTA TAGGGC-3′). Primers (F1, R1 and F2, R2) were used for the first and second step clones of the 5′ sequence of SsDGAT1, respectively. For the 3′ RACE clone, primers (F3, R3 and F4, R4) were respectively utilized in the first and second steps. Gene-specific primers F0 (5′-ATGGCGGTTCTTGACGC-3′) and R0 (5′-TCAACCTGATTCAGTTAGCCCTTTTCG-3′) were designed to obtain the complete open reading frame (ORF) by RT-PCR. This ORF fragment was cloned and sequenced.

Cloning of SsDGAT1 from S. sebiferum seed cDNA. a Amplification of the conserved region of SsDGAT1 by RT-PCR; b amplification of the 3′ terminus of SsDGAT1 by 3′ RACE; c amplification of the 5′ terminus of SsDGAT1 by 5′ RACE; d amplification of the full length of SsDGAT1 by RT-PCR; e amino acid sequence alignment of DGAT1 homologs from various plant species; f phylogenetic analysis of DGAT1s; g predicted transmembrane domain of SsDGAT1. The evolutionary relationships among the 64 DGAT1 from 49 organisms were subjected to phylogenetic analysis based on the neighbor-joining method of Tamura et al. (2007). The framed section indicates the SsDGAT1 protein studied here. Genes arere labeled with their GenBank accession numbers, followed in parentheses by the initials of the organism. The calculated distance values are indicated before the gene labels, and reflect the degree of divergence between all pairs of SsDGAT1 sequences analyzed. Abbreviations: Aa in XP_001658299, Aedes aegypti; Ac in EGC41804.1, Ajellomyces capsulatus H88; Ao in EEQ 31683.1, Arthroderma otae CBS 113480; At in NP_179535.1, Arabidopsis thaliana; Bb in AAZ22403.1, Bubalus bubalis; Bn in AAD45536.1, in AAD40881.1, Brassica napus; Bj in AAY40784.1, in AAY40785.1, Brassica juncea; Bt in NP_777118 2, Bos taurus; Ca in ABD59375.1, Capra hircus; Cf in XP_849176.1 and XP_858062.1, Canis familiaris; Cv in EFN50697.1, Chlorella variabilis; Dd in XP_645633.2, Dictyostelium discoideum AX4; Dm in NP_609813.1 and NP_995724.1, Drosophila melanogaster; Dr in NP_001002458.1 and NP_956024.1, Danio rerio; Ea in AAV31083.1, Euonymus alatus; Ep in ACO55635.1, Echium pitardii; Gm in AAS78662.1 and BAE93461.1, Glycine max; Hs in NP_036211.2, Homo sapiens; Jc in ABB84383.1, Jatropha curcas; Lj in AAW51456 1, Lotus japonicas; Ma in EFY86774.1, Metarhizium acridum CQMa 102; Ma in EFY97444.1, Metarhizium anisopliae ARSEF 23; Md in XP_001371565.1, Monodelphis domestica; Mm in XP_001090134.1, Macaca mulatta; Mm in NP_034176.1, Mus musculus; Mt in ABN09107.1, Medicago truncatula; Nt in AAF19345.1, Nicotiana tabacum; Oa in NP_001103634.1, Ovis aries; Oc in XP_002724427.1, Oryctolagus cuniculus; Oe in AAS01606.1, Olea europaea; Os in NP_001054869.2, Oryza sativa japonica; Pb in EEH17170.1, Paracoccidioides brasiliensis Pb03; Pf in AAG23696.1, Perilla frutescens; Pp in XP_001770929.1, Physcomitrella patens; Pp in EFA85004.1, Polysphondylium pallidum PN500; Pt in XP_520014.2, Pan troglodytes; Pt in XP_002177753 1, Phaeodactylum tricornutum CCAP 1055/1; Pt in XP_002308278.1 and XP_002330510.1, Populus trichocarpa; Rc in XP_002514132.1, Ricinus communis; Rn in NP_445889.1, Rattus norvegicus; Sb in XP_002439419.1 and XP_002437165.1, Sorghum bicolor; Sk in XP_002736160.1, Saccoglossus kowalevskii; Sm in XP_002964165.1, Selaginella moellendorffii; Ss in NP_999216.1, Sus scrofa; Tc in XP_975142.1, Tribolium castaneum; Tg in AAP94209.1, Toxoplasma gondii; Tm in AAM03340.2, Tropaeolum majus; Vf in DQ356680.1, Vernicia fordii; Vg in ABV21945.1, Vernonia galamensis; Vv in XP_002279345.1, Vitis vinifera; Zm in 2-EU039830, Zea mays
Phylogenetic analysis
The phylogenetic analysis was performed in accordance with the method of Wang et al. (2015).
Putative transmembrane domains
We predicted the transmembrane domain of SsDGAT1 using http://www.cbs.dtu.dk/services/TMHMM/online software.
Semi-quantitative RT-PCR and quantitative real-time PCR (qPCR)
Semi-quantitative RT-PCR and/or qPCR were performed to detect the expression of SsDGAT1 in each organ and in seeds at different growth phases. Ss18S (GenBank accession No. DQ864377.1) fragment, encoding a 240-bp region of Triadica sebifera 18S ribosomal RNA, was used as internal reference. Primer sets used for semi-quantitative RT-PCR and qPCR were as follows: SsDGAT1-specific 5′ (5′-TTACCAGCCAAGTTATCCTCG-3′) and 3′ (5′-AGCAGTAGAACATGCAGAGCC-3′), internal control’s Ss18S-F (5′-ATGGGATAACATCATAGGATTTCGG-3′) and Ss18S-R (5′-TGGTCGGCATCGTTTATGGTT-3′). The thermal cycling program of semi-quantitative RT-PCR consisted of 30 cycles of 30 s at 94 °C, 30 s at 55 °C, and 30 s at 72 °C. qPCR was performed using a CFX-96 (Bio-Rad, USA) Real-Time system. Samples were run in triplicate, with starting quantity estimated from critical thresholds using the standard curve of amplification.
Yeast transformation and selection
The SsDGAT1 gene was introduced into the pYES2 vector by using XhoI and XbaI digestion sites and then the pYES2-SsDGAT1 construct was transferred into S. cerevisiae H1246 (W303; MATα are1-Δ::HIS3 are2-Δ::LEU2 dga1::KanMX4 lro1-Δ::TRP1 ADE2 met ura3) (Sandager et al. 2002) using the polyethylene glycol (PEG)/lithium acetate method (Gietz and Woods 1998). Yeast cells harboring the empty pYES2 vector were used as the negative control and S. cerevisiae W3013A was used as the wild-type (WT) yeast strain in this experiment. Selection of positive transformants was carried out as described by Wang et al. (2015).
Lipid extraction and analysis
Extraction and analysis of lipids from yeast cell was performed as described previously (Wang et al. 2015). TAGs were separated from total lipids by thin-layer chromatography (TLC) using a solvent system of hexane/ether/acetic acid (70/30/1, v/v/v) (Xu et al. 2013). Individual lipid spots were visualized by exposing the silica gel plates (Shanghai, China) to iodine vapor.
Analysis of seed oil, protein, glucosinolate and glucose content in B. napus
The oil, protein, glucosinolate and glucose content of mature B. napus seed were determined by the method of Lock et al. (2009).
Construction and identification of transgenic B. napus plants
The SsDGAT1 gene from S. sebiferum cDNA was amplified using the primers 5′-TCCCCCGGG ATGGCGGTTCTTGACGC-3′ and 5′-CGCGGATCC ACCTGATTCAGTTAGCCCTTTTCG-3′ (restriction sites Sma1 ‘CCCGGG’ and EcoR1 ‘GGATCC’ are shown in boldface, and the sequence corresponding to SsDGAT1 is underlined). The amplified product was sub-cloned into the SmaI–XhoI cloning site of the pGEM®-T Easy vector (Promega, USA). After the cloned intact SsDGAT1 was confirmed correct by sequencing, the SmaI–XhoI DNA fragments of SsDGAT1 were excised from pGEM®-T Easy vector by SmaI and XhoI digestion and inserted into the binary vector pEGAD (Fig. 4a). The construct 35S::GFP-SsDGAT1 was introduced into Agrobacterium tumefaciens strain GV3101 by electroporation using a Gene Pulser II system (Bio-Rad, Hercules, CA, USA) and the stabilized positive constructs selected with 100 mg L−1 rifampicin and 50 mg L−1 kanamycin were confirmed by PCR, and then B. napus plants were transformed by the Agrobacterium tumefaciens-mediated floral dip transformation method (Zhou et al. 2012). In detail, the grown healthy flora inflorescences at the top of B. napus plants which have many immature flower clusters and only few fertilized siliques were selected as transformation material, while the siliques, opened flowers found on the flora inflorescences and all other branches and flora inflorescences that could be immersed were removed with pruning scissors before gene transformation. The pruned inflorescences were swathed and immersed in the plastic valve bag with Agrobacterium containing 35S::GFP-SsDGAT1 inoculum (OD = 0.6) for 30 s with gentle agitation. The dipped inflorescences were removed from the solution and the treated inflorescences drained for 3–5 s. A film of liquid should be observed coating the inflorescences. The bacteria were distributed to all inflorescences including very young flower shoots by gently pressing the outside of the plastic bag with the hands, taking care not to break the B. napus plants and inflorescences. The transformation procedure was repeated twice every 3 days after the first transformation. The 35S::GFP-SsDGAT1 construct and its transgenic daughter lines were selected for further study. T1 generation seeds were selected using the herbicide Basta (v/v, 1:1000). T2 lines were used for seed oil analysis.
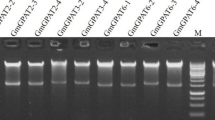
DNA of 7-day-old seedlings was extracted from the WT and Basta-resistant transgenic lines using the CTAB method (Murray and Thompson 1980). The Basta resistance bar gene, GFP gene and GFP-SsDGAT1 gene were determined by PCR analysis with primers (bar-F: 5′-CTACATCGAGACAAGCACGGT-3′ and bar-R: 5′-CTGAAGTCCAGCTGCCAGAA-3′, GFP-F: 5′-GACCCTGAAGTTCATCTGCACCACC-3′ and GFP-R: 5′-CCGATGGGGGTGTTCTGCTG-3′, G + S-F: 5′-TAAACGGCCACAAGTTCAGCG-3′, G + S-R: 5′-CGTCTACTGTTCTTGCATTCCACC-3′). PCR was performed at 94 °C for 5 min followed by 30 cycles of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 30 s, and then 72 °C for 5 min. PCR products were analyzed using 1.0 % agarose gel electrophoresis. Total RNA was extracted from 100 mg of seedlings of 7-day-old B. napus using Trizol reagent (Invitrogen, USA) and treated with RNase-free DNase I (Promega Biotech Co., USA). The DNAse-digested RNA samples were used for reverse transcription by Superscript III reverse transcriptase (Invitrogen). Samples serving as cDNA stocks for PCR analysis were stored at −80 °C. Gene expression was analyzed by semiquantitative RT-PCR using rTaq DNA polymerase (Takara, Dalian, China) and a programmable thermal cycler CFX-96 (Bio-Rad, USA) in a 20-µL solution containing 1 µL cDNA stock. The PCR amplification consisted of an initial denaturation step at 94 °C for 5 min followed by 22 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 20 s. The sense and antisense primers of SsDGAT1 were 5′-TAGTTCGCCATATCTACTTTCCA-3′ and 5′-AACCTGATTCAGTTAGCCCTTTT-3′, respectively. BnACTIN7 was used an internal control; its sense primer (5′-TGTTGCTATCCAGGCTGTTCT-3′) and anti-sense primer (5′-TTAATGTCACGGACGATTTCC-3′) were designed according to the Genbank Accession Number AF111812.
DNA gel blotting
Genomic DNA was isolated from leaves of individual seedlings using the Plant DNA extraction kit (Takara) according to the manufacturer’s instructions. After endonuclease digestion with EcoRI and electrophoresis on a 1 % agarose gel, DNA fragments were transferred to a positively charged nylon membrane (Roche). Probes were prepared using a PCR DIG probe synthesis kit (Roche). Hybridization was performed according to the DIG Application Manual (Roche). Hybridization was carried out at 42 °C and washing was performed under high-stringency conditions at 68 °C. GFP gene was used as probe in this study; the primers of GFP were GFP-F: 5′-CACAAGTTCAGCGTGTCCG-3′, GFP-R: 5′-AGTTCACCTTGATGCC GT TC-3′.
Immunoblot analysis
Total protein was isolated from B. napus seeds after flowering for 60 days according to the procedure described by Zhou et al. (2012). The enhanced chemiluminescence immunoblot analysis was performed following the method of Wang et al. (2015). In this study, the immunoblot was probed with appropriate primary antibodies (anti-GFP antibody) and secondary antibody conjugated with horseradish peroxidase (Beyotime Institute of Biotechnology, China). Histone1 was used to verify equal loading. All other chemicals were acquired from Sigma (St. Louis, MO, USA).
Oil analysis by GC–MS
The B. napus seed lipids were extracted from 10 mg dried seeds of control and transgenic lines (Li et al. 2006). The oil analysis was carried out by GC–MS according to the method of Misra et al. (2013).
Results
Cloning of the SsDGAT1 gene from S. sebiferum
An 837-bp conserved nucleotide sequence corresponding to the SsDGAT1 gene (Fig. 1a) was obtained by PCR amplification, homologous region cloning and sequencing, then a 3′ terminal fragment of the SsDGAT1 gene was successfully amplified based on this nucleotide sequence (Fig. 1b). Furthermore, a 5′ terminal fragment of the SsDGAT1 gene was produced by nested PCR after 5′ RACE (Fig. 1c). Sequencing results showed that the 3′ terminal fragment and 5′ terminal fragment were 783 and 812 bp in length, respectively. The SsDGAT1 full-length cDNA with 1524 bp (Fig. 1d) was determined by the overlapping sequences of the 5′ and 3′ ends and then amplified by RT-PCR, which encoded a protein with 507 amino acids (Fig. 1e). We have submitted the sequence to National Center for Biotechnology Information (NCBI); the BankIt No. is 1926735 and the accession number is KX372341. The transmembrane structure analysis result indicated that SsDGAT1 encoded a peptide consisting of nine putative transmembrane domains (Fig. 1g). A phylogenetic tree was generated from an alignment of the amino acid sequences between SsDGAT1 and four DGAT1 homologs from diverse species, and three highly conserved motifs, the Basic motif acyl-CoA binding signature, DAG-binding and putative ER retrieval motifs (ER-DIR), were contained in all the five DGAT1 proteins (Fig. 1e). The dendrogram revealed that SsDGAT1 was most closely related to RcDGAT1, JcDGAT1 and VfDGAT1, which together formed a separate cluster from other DGAT1 branches (Fig. 1f).
Investigation of the expression pattern of SsDGAT1
To clarify the spatio-temporal expression pattern of SsDGAT1 in S. sebiferum, semi-quantitative RT-PCR and qPCR were carried out using cDNA extracted from various organs. As shown in semi-quantitative RT-PCR analysis, the SsDGAT1 gene had a high expression level in seed, leaf and stem, whereas a low transcript level of SsDGAT1 was detected in flower and root (Fig. 2a). This semi-quantitative RT-PCR result was coincident with qPCR (Fig. 2b). During developmental stages of seeds, the relative expression levels of SsDGAT1 peaked at 120 days after pollination (DAP) (Fig. 2c), which indicated that the oil might have formed at a point during this time. Overall, SsDGAT1 was observed at high levels of expression in the seed, leaf, and stem but low levels in the flower and root; during the entire development of seeds, the highest SsDGAT1 expression was found at 120 DAP.

Investigation of the expression pattern of SsDGAT1. a Semi-quantitative RT-PCR to investigate the expression level of SsDGAT1 in various organs; b qPCR to investigate the expression level of SsDGAT1 in various organs; c qPCR to investigate the expression level of SsDGAT1 at different developmental stages of S. sebiferum seeds. Samples were collected at 15-day intervals from the day after pollination (DAP) to seed fully matured
Evaluation of SsDGAT1 as a functional DGAT enzyme
To investigate the function of SsDGAT1, its complete coding sequence was expressed in the TAG-deficient S. cerevisiae strain H1246, which contains disruptions of four acyltransferase genes contributing to TAG synthesis: are1, are2, dga1 and lro1 (Sandager et al. 2002). In this series of assays, SsDGAT1 was driven by the galactose-inducible GAL1 promoter in pYES2 to generate the plasmid pYES2-SsDGAT1 and the wild-type S. cerevisiae strain W3013A was used as a positive control. TLC analysis of extracted lipids revealed marked accumulation of TAG in cells of H1246 with pYES2-SsDGAT1 construct as well as W3013A, but absent in cells of H1246 transformed with only the empty vector pYES2 (Fig. 3), which suggested that SsDGAT1 possessed sufficient DGAT activity to restore TAG synthesis in S. cerevisiae H1246.

TLC analysis of neutral lipids from yeast transformants. FFA free fatty acid
Molecular characterization of SsDGAT1 transgenic B. napus
An expression cassette fused the green fluorescent protein (GFP) gene, the SsDGAT1 gene and the herbicide Basta-resistant selectable marker gene was constructed, in which SsDGAT1 and GFP was driven by the CaMV35S promoter, and expressed in B. napus plants (Fig. 4a). Twenty-nine Basta-resistant independent B. napus lines were obtained after the first treatment with the Basta herbicide (Fig. 4b). Three Basta-resistant independent lines (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7 and 35S::GFP-SsDGAT1-10) with 3:1 segregation ratio of Basta-resistant:Basta-sensitive T1 seedlings were selected for further studies. The bar genes (Fig. 4c), GFP genes (Fig. 4d) and GFP-SsDGAT1 genes (Fig. 4e) were detected in these three Basta-resistant lines but not in WT plants, showing that the expression cassette was successfully integrated into the B. napus genomic DNA, importantly, which was as a single copy insert in transgenic lines genomic DNA by DNA gel blot using GFP gene as a probe (Fig. 4f). In addition, the high transcription signal of SsDGAT1 by semi-quantitative RT-PCR was clearly observed in these three Basta-resistant B. napus lines but was not detected in WT seedlings (Fig. 4g), which further confirmed that the SsDGAT1 gene was truly expressed in transgenic plants. Moreover, an immunoblot assay also showed that the GFP-SsDGAT1 fusion protein was accumulated in transgenic B. napus plants but not in the WT (Fig. 4h). Taken together, all the results demonstrated that SsDGAT1 was successfully over-expressed in B. napus.

Transformation and molecular characterization of transgenic B. napus plants. a Construct of SsDGAT1 expression cassette; b screening of representative Basta-resistant B. napus seedling; c PCR analysis for bar gene from 7-day-old transgenic seedlings (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7, 35S::GFP-SsDGAT1-10); d PCR analysis for GFP gene from 7-day-old transgenic seedlings (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7, 35S::GFP-SsDGAT1-10); e PCR analysis for GFP-SsDGAT1 gene from 7-day-old transgenic seedlings (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7, 35S::GFP-SsDGAT1-10); f DNA gel blot analysis using GFP gene as a probe for 7-day-old transgenic lines (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7, 35S::GFP-SsDGAT1-10); g semi-quantitative RT-PCR of 7-day-old transgenic lines (35S::GFP-SsDGAT1-4, 35S::GFP-SsDGAT1-7, 35S::GFP-SsDGAT1-10); the housekeeping gene Ss18S was used as a control; h immunoblots of protein from seeds of transgenic and WT plants. GFP-SsDGAT1 was detected using an anti-GFP antibody
Enhancement of seed oil content in over-expressing transgenic B. napus
To determine whether the increased SsDGAT1 expression was accompanied by a rise in oil content in matured seeds, we measured the latter in the over-expressing SsDGAT1 seeds as well as in WT seeds. Notably, over-expressing SsDGAT1 led to about 13 % higher oil content in transgenic seeds than WT seeds (Fig. 5a). Interestingly, other metabolite pools, such as glucosinolates, glucose and protein, all showed an trend of increase in transgenic seeds compared with WT seeds (Fig. 5b–d).

Analysis of the seed oil, protein, glucosinolate and glucose content, 1000-seed weight and fatty acid composition in WT and transgenic B. napus. Values were calculated on a dry weight basis. Bars indicate mean ± standard error. Statistical analysis was carried out by Student’s t test and Fisher’s least squares difference/P = 0.001 for a–f). * P < 0.05, ** P < 0.01. a Investigation of the seed oil content in WT and transgenic B. napus. b Investigation of the seed protein content in WT and transgenic B. napus. c Investigation of the seed glucosinolate content in WT and transgenic B. napus. d Investigation of the seed glucose content in WT and transgenic B. napus. e 1000-seed weight of seeds in WT and transgenic B. napus. f Fatty acid composition of seeds in WT and transgenic B. napus
Alteration of the seed weight and fatty acid composition in B. napus plants over-expressing SsDGAT1
To test if the increased SsDGAT1 expression was associated with the mass of seeds, we determined the 1000-seed weight. As shown in Fig. 5e, seeds of B. napus over-expressing SsDGAT1 showed 21 % greater 1000-seed weight than the WT seeds. In addition, the fatty acid composition was analyzed to examine the effect of increased SsDGAT1 expression on the proportion of individual fatty acids. Notably, plants over-expressing SsDGAT1 showed amounts of oleic acid (18:1) decreased by 13.3–18.3 %, whereas linoleic acid (18:2) and linolenic acid (18:3) content were observed to increase by 14.5–17.8 % and 7.4–11.2 % respectively, compared with the WT (Fig. 5f).
Discussion
In oil-rich seeds, the most important form of energy storage is triacylglycerols (TAGs), especially in woody oil tree seeds. DGAT is believed to be a key enzyme synthesizing TAGs from DAGs in the major flux control step in oil biosynthesis. Previous studies indicated that TAGs are synthesized by four independent family genes, DGAT1, DGAT2, DGAT3 and PDAT, each encoding unique proteins (Shockey et al. 2006; Zhang et al. 2009; Cao et al. 2013). In plants, a member of the DGAT1 family plays a major role in TAG accumulation in several tissues including seeds while members of the DGAT2 and PDAT families appear to be active mainly in seeds, especially in the accumulation of unusual fatty acids (FAs) in seed oils such as castor (Kroon et al. 2006; Kim et al. 2011) and tung tree (Shockey et al. 2006). At the same time, previous research suggested that DGAT3 is expressed throughout seed development and seedling establishment in Arabidopsis, and could play a role in an alternative pathway for TAG synthesis during pxa1 seedling establishment (Hernández et al. 2012). In our study, we characterized a homolog of DGAT1 from S. sebiferum, a seed-oil-rich woody tree, showed its expression pattern and assessed its role in B. napus for the first time, which led to an increase in oil accumulation and alteration of FA composition in transgenic rapeseed oil.
The SsDGAT1 gene encoded a peptide consisting of nine putative transmembrane domains and a relatively hydrophilic N-terminal segment, like other DGAT1 proteins reported previously (Lung and Weselake 2006; Misra et al. 2013), besides an YYHDL motif at the C-terminus that has been reported to be required for ER localization (Shockey et al. 2006). Transcript accumulation of the SsDGAT1 mRNA in seeds revealed a specific increase during the later stages of seed development with a maximum level at 120 days after pollination, which matched the results reported by Xu et al. (2011) and showed a pattern similar to other oil-seed crops such as B. napus, Ricinus communis, Euphorbia lagascae and Vernonia galamensis (Li et al. 2010). Of course, the relative expression of SsDGAT1 was detected in other tissues, particularly high in the stem but still much lower than in seeds.
In addition to transcriptional characterization of SsDGAT1, functional validation of SsDGAT1 was demonstrated by the effect of its expression on a yeast mutant and B. napus. In the complementation analysis, when SsDGAT1 was transformed into yeast mutant cell H1246, a quadruple-deficient strain in lipid biosynthesis, a compensating TAG accumulation was able to be regenerated. Importantly, expression of SsDGAT1 under the CaMV35S promoter in B. napus did indeed improve rapeseed oil biosynthesis, increasing oil content by 12.3–14.7 % over that of the control. Considering that the DAG pool in seeds is small, the expression of SsDGAT1 brought about a considerable increase in seed oil biosynthesis, which indicated that it must in some way influence the biosynthesis and/or the flux of FAs that enter the TAG pathway and their subsequent incorporation in seed TAGs. Indeed, changes in expression of DGAT in Arabidopsis, B. napus and Nicotiana tabacum have been shown to affect not only oil content but also seed size and shape and a large number of developmental events (Jako et al. 2001; Weselake et al. 2007; Sharma et al. 2008; Misra et al. 2013). Additionally, many genes were found to be differentially regulated in DGAT over-expressing Arabidopsis lines and hormone levels, such as those of auxin, cytokinins and abscisic acid, were unusually influenced by DGAT expression in plants (Sharma et al. 2008). This may partly explain the changes that go beyond utilization of the unused DAG pool.
Interestingly, the oil content of over-expressed seeds increased by 13.0 % and the level of linolenic acid was higher at 11.2 % compared with control. This study suggested that over-expressing heterologous DGAT1 in plants can not only increase oil content but also alter oil composition, which was mirrored similarly by the result of Wang et al. (2014), who reported that over-expression of Sesamum indicum L. DGAT1 (SiDGAT1) in Arabidopsis thaliana ‘Col 0’ resulted in an increase in oil content and seed weight, and altered the oil composition. As well-known, there is a double bond in linolenic acid which can store up more energy when the carbon skeleton is unchanged. Thus, increased unsaturated fatty linolenic acid content, meaning an increase in double bonds, can meet the requirements of biodiesel production. Nevertheless, there was no change in the levels of saturated FAs such as palmitic acid and stearic acid. It has been speculated that variety in oil composition between different plants may be partly due to the specificities of DGATs for various fatty acyl-CoAs (saturated 16:0, 18:0 or mono- and polyunsaturated FAs), especially in seeds such as those from tung, cocoa, rapeseed and castor where there is often a higher incorporation of many unusual FAs in seeds (Burgal et al. 2008; Griffiths and Harwood, 1991; Kroon et al. 2006; Shockey et al. 2006). Other studies suggested that the differences in oil composition reflected differences in acyl-CoA pools and not necessarily FA specificities of DGAT: e.g. the manipulation of the tobacco FA pool by expression of an epoxygenase gene from E. lagascae (Cahoon et al. 2002), and higher incorporation of the modified FAs into the oil, although with no manipulation of the DGAT but only by the co-expression of a △12-epoxygenase and a △12-desaturase from C. palaestina in Arabidopsis (Zhou et al. 2006). In our results, a visible increase in linolenic acid and a specific decrease in oleic acid from SsDGAT1 expression would suggest a higher affinity for linolenic acids and a lower one for oleic acid.
Our study shows that the seed oil was markedly increased and glucosinolates and glucose were significantly decreased by over-expression of SsDGAT1 in B. napus. Further studies indicated that the increased seed oil content was largely compensated by decreases in other metabolite pools, such as glucosinolates and glucose, but most predominantly protein. Indeed, this relationship has been negatively demonstrated in tobacco, where both protein and carbohydrate increased as seed oil decreased in DGAT1-silenced plants (Zhang et al. 2005); at the same time, this relationship has been negatively demonstrated in B. napus (Lock et al. 2009).
Conclusions
In this research, S. sebiferum SsDGAT1 transcription showed enhancement during seed development and that SsDGAT1 heterologous expression in yeast H1246 TAG-deficient mutant and in B. napus can lead to increased TAG biosynthesis. Furthermore, our results showed not only increased oil content but also alterations of oil composition: an increase in linolenic acids levels, a decrease in oleic acid levels and no changes in saturated fatty acid levels in SsDGAT1 over-expressing B. napus. Thus, SsDGAT1 could be considered as a candidate gene for increasing oil content and improving oil quality.
Abbreviations
- qPCR:
-
Quantitative real-time polymerase chain reaction
- RT-PCR:
-
Reverse-transcription polymerase chain reaction
- RACE:
-
Rapid amplification of cDNA ends
- GFP:
-
Green fluorescent protein
- cDNA:
-
Complementary DNA
- CTAB:
-
Hexadecyltrimethylammonium bromide
- TLC:
-
Thin-layer chromatography
- GC–MS:
-
Gas chromatography–mass spectrometry
- FA:
-
Fatty acids
References
Andrianov V, Borisjuk N, Pogrebnyak N, Brinker A, Dixon J, Spitsin S, Flynn J, Matyszczuk P, Andryszak K, Laurelli M, Golovkin M, Koprowski H (2010) Tobacco as a production platform for biofuel: overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotechnol J 8(3):277–287
Bouvier-Navé P, Benveniste P, Oelkers P, Sturley SL, Schaller H (2000) Expression in yeast and tobacco of plant cDNAs encoding acyl CoA:diacylglycerol acyltransferase. Eur J Biochem 267(1):85–96
Burgal J, Shockey J, Lu CF, Dyer J, Larson T, Graham I, Browse J (2008) Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol J 6:819–831
Cahoon EB, Ripp KG, Hall SE, McGonigle B (2002) Transgenic production of epoxy fatty acids by expression of a cytochrome P450 enzyme from Euphorbia lagascae seed. Plant Physiol 128:615–624
Cahoon EB, Shockey JM, Dietrich CR, Gidda SK, Mullen RT, Dyer JM (2007) Engineering oilseeds for sustainable production of industrial and nutritional feedstocks: solving bottlenecks in fatty acid flux. Curr Opin Plant Biol 10:236–244
Cao H, Shockey JM, Klasson KT, Chapital DC, Mason CB, Scheffler BE (2013) Developmental regulation of diacylglycerol acyltransferase family gene expression in tung tree tissues. PLoS One 8(10):e76946
Cui YL, Zheng GT, Li XQ, Lin HZ, Jiang P, Qin S (2013) Cloning and characterization of a novel diacylglycerol acyltransferase from the diatom Phaeodactylum tricornutum. J Appl Phycol 25:1509–1512
Divi UK, Zhou XR, Wang PH, Butlin J, Zhang DM, Liu Q, Vanhercke T, Petrie JR, Talbot M, White RG, Taylor JM, Larkin P, Singh SP (2016) Deep sequencing of the fruit transcriptome and lipid accumulation in a non-seed tissue of chinese tallow, a potential biofuel crop. Plant Cell Physiol 57(1):125–137
Durrett TP, Benning C, Ohlrogge J (2008) Plant triacylglycerols as feedstocks for the production of biofuels. Plant J 54:593–607
Gietz RD, Woods RA (1998) Transformation of yeast by the lithium acetate/single-stranded carrier DNA/PEG method. Methods Enzymol 26:53–66
Griffiths G, Harwood JL (1991) The regulation of triacylglycerol biosynthesis in cocoa (Theobroma cacoa) L. Planta 184:279–284
He X, Turner C, Chen G, Lin JT, Mckeon T (2004) Cloning and characterization of a cDNA encoding diacylglycerol acyltransferase from castor bean. Lipids 39:311–318
Hernández ML, Whitehead L, He ZS, Gazda V, Gilday A, Kozhevnikova E, Vaistij FE, Larson TR, Graham IA (2012) A cytosolic acyltransferase contributes to triacylglycerol synthesis in sucrose-rescued arabidopsis seed oil catabolism mutants. Plant Physiol 160:215–225
Jako C, Kumar A, Wei Y, Zou J, Barton DL, Giblin EM, Covello PS, Taylor DC (2001) Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight. Plant Physiol 126:861–874
Katavic V, Reed DW, Taylor DC, Giblin EM, Barton DL, Zou J, Mackenzie SL, Covello PS, Kunst L (1995) Alteration of seed fatty acid composition by an ethyl methane sulfonate-induced mutation in Arabidopsis thaliana affecting diacylglycerol acyltransferase activity. Plant Physiol 108:399–409
Kim HU, Lee KR, Go YS, Jung JH, Suh MC, Kim JB (2011) Endoplasmic reticulum-located PDAT1-2 from castor bean enhances hydroxy fatty acid accumulation in transgenic plants. Plant Cell Physiol 52(6):983–993
Kroon JTM, Wei WX, Simon WJ, Slabas AR (2006) Identification and functional expression of a type 2 acyl-CoA: diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds. Phytochemistry 67(23):2541–2549
Li Y, Beisson F, Pollard M, Ohlrogge J (2006) Oil content of Arabidopsis seeds: the influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 67:904–915
Li RZ, Yu KS, Hildebrand DF (2010) DGAT1, DGAT2 and PDAT expression in seeds and other tissues of epoxy and hydroxy fatty acid accumulating plants. Lipids 45:145–157
Lock YY, Snyderb CL, Zhu WM, Siloto RMP, Weselakeb RJ, Shaha S (2009) Antisense suppression of type 1 diacylglycerol acyltransferase adversely affects plant development in Brassica napus. Physiol Plant 137:61–71
Lung SC, Weselake R (2006) Diacylglycerol acyltransferase: a key mediator of plant triacylglycerol synthesis. Lipids 41:1073–1088
Misra A, Khan K, Niranjan A, Nath P, Sane VA (2013) Over-expression of JcDGAT1 from Jatropha curcas increases seed oil levels and alters oil quality in transgenic Arabidopsis thaliana. Phytochemistry 96:37–45
Murray MG, Thompson WF (1980) Rapid isolation of high molecular weight plant DNA. Nucleic acids Res 8(19):4321–4326
Peng X, Yi N, Cheng T (2008) Research advances in chemical constituents and pharmacological effects of Sapium sebiferum. Chin Wild Plant Resour 27(3):16–19
Routaboul JM, Benning C, Bechtold N, Caboche M, Lepiniec L (1999) The TAG1 locus of Arabidopsis encodes for a diacylglycerol acyltransferase. Plant Physiol Biochem 37:831–840
Saha S, Enugutti B, Rajakumari S, Rajasekharan R (2006) Cytosolic triacylglycerol biosynthetic pathway in oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. Plant Physiol 141:1533–1543
Sandager L, Gustavsson MH, Stahl U, Dahlqvist A, Wiberg E, Banas A, Lenman M, Ronne H, Stymne S (2002) Storage lipid synthesis is non-essential in yeast. J Biol Chem 277(8):6478–6482
Sharma N, Maureen A, Arvind K, Zhang Y, Giblin EM, Abrams SR, Zaharia LI, Pierre DT, Fobert R (2008) Transgenic increases in seed oil content are associated with the differential expression of novel Brassica-specific transcripts. BMC Genomics 9:619. doi:10.1186/1471-2164-9-619
Shockey JM, Gidda SK, Chapital DC, Kuan JC, Dhanoa P, Bland JM, Rothstein SJ, Mullen RT, Dyer JM (2006) Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell 18:2294–2313
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA 4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evolut 24:1596–1599
Wang ZK, Huang WJ, Chang JM, Sebastian A, YoG Li, Li HY, Wu XX, Zhang BB, Meng FL, Li WB (2014) Overexpression of SiDGAT1, a gene encoding acyl-CoA:diacylglycerol acyltransferase from Sesamum indicum L. increases oil content in transgenic Arabidopsis and soybean. Plant Cell Tissue Organ Cult 119:399–410
Wang YQ, Peng D, Zhang L, Tan XF, Yuan DY, Liu XM, Zhou B (2016) Overexpression of SsDGAT2 from Sapium sebiferum (L.) Roxb increases seed oleic acid level in Arabidopsis. Plant Mol Biol Rep 34:638–648
Weselake RJ, Shah S, Taylor DC, Zou J, Laroche A, Moloney M, Rakow G, Raney P, Harwood J (2007) Transformation of Brassica napus with diacylglycerol acyltransferase-1 results in increased seed oil content. In: Benning C, Ohrogge J (eds) Current advances in the biochemistry and cell biology of plant lipids. Aardvark Global Publishing, Salt Lake City, pp 232–234
Weselake RJ, Shah S, Tang M, Quant PA, Snyder CL, Furukawa TL, Zhu WM, Taylor DC, Zou JT, Kumar A, Hall L, Laroche A, Rakow G, Raney P, Moloney MM, Harwood JL (2008) Metabolic control analysis is helpful for informed genetic manipulation of oilseed rape (Brassica napus) to increase seed oil content. J Exp Bot 59:3543–3549
Xu R, Wang R, Liu A (2011) Expression profiles of genes involved in fatty acid and triacylglycerol synthesis in developing seeds of Jatropha (Jatropha curcas L.). Biomass Bioenergy 35:1683–1692
Xu J, Kazachkov M, Jia Y, Zheng Z, Zou J (2013) Expression of a type 2 diacylglycerol acyltransferase from Thalassiosira pseudonana in yeast leads to incorporation of docosahexaenoic acid β-oxidation intermediates into triacylglycerol. FEBS J 280:6162–6172
Yang Z, Qi Y, Wang X, Yang L, Lie A, Sun YP, Hu XH (2007) Preliminary report on research of Sapium sebiferum Roxb. seed producing biodiesel. Hubei For Sci Technol 6:32–34 (In Chinese)
Yang ML, Wu Y, Jin S, Hou JY, Mao YJ, Liu WB, Shen YC, Wu LF (2015) Flower bud transcriptome analysis of Sapium sebiferum (Linn.) roxb. and primary investigation of drought induced flowering: pathway construction and g-quadruplex prediction based on transcriptome. PLoS One. doi:10.1371/journal.pone.0118479
Yu K, McCracken JCT, Li R, Hildebrand DF (2006) Diacylglycerol acyltransferases from Vernonia and Stokesia prefer substrates with vernolic acid. Lipids 41:557–566
Yu K, Li R, Hatanaka T, Hildebrand D (2008) Cloning and functional analysis of two type 1 diacylglycerol acyltransferases from Vernonia galamensis. Phytochemistry 69:1119–1127
Zhang FY, Yang MF, Xu YN (2005) Silencing of DGAT1 in tobacco causes a reduction in seed oil content. Plant Sci 169:689–694
Zhang M, Fan J, Taylor DC, Ohlrogge JB (2009) DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 21(12):3885–3901
Zhou XR, Singh S, Liu Q, Green A (2006) Combined transgenic expression of Δ12-desaturase and D12-epoxygenase in high linoleic substrate seed oil leads to increased accumulation of vernolic acid. Funct Plant Biol 33:585–592
Zhou B, Lin JZ, Peng WS, Peng D, Zuo YH, Zhu DF, Huang XQ, Tang DY, Guo M, He RQ, Zhang JH, Li XS, Zhao XY, Liu XM (2012) Dwarfism in Brassica napus L. induced by the over-expression of a gibberellin 2-oxidase gene from Arabidopsis thaliana. Mol Breed 29:115–127
Zou J, Wei Y, Jako C, Kumar A, Selvaraj G, Taylor DC (1999) The Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol acyltransferase gene. Plant J 19:645–653
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31200516), the Outstanding Youth Scientific Research Foundation held by the Hunan Education Department (13B150), The Hunan Province Outstanding Youth Teacher Cultivation Project (2014gg04), the Youth Scientific Research Foundation of Central South University of Forestry & Technology (QJ2011046B), and the Foundation of the Project for Talents Introduction of the Central South University of Forestry & Technology (104-0182). We also thank Professor Aizhong Liu (Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences) for providing the yeast strain H1246 and the corresponding parental line for this study.
Author contributions
Idea and study design: DP and BZ; data collection and analysis: DP, BZ, LZ, DY and TX with the support of XL; writing of the manuscript: DP and BZ.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Peng, D., Zhang, L., Tan, X. et al. Increasing seed oil content and altering oil quality of Brassica napus L. by over-expression of diacylglycerol acyltransferase 1 (SsDGAT1) from Sapium sebiferum (L.) Roxb.. Mol Breeding 36, 136 (2016). https://doi.org/10.1007/s11032-016-0543-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-016-0543-2