
Abstract
Cabbage head splitting can greatly affect both the quality and commercial value of cabbage (Brassica oleracea). To detect the genetic basis of head-splitting resistance, a genetic map was constructed using an F2 population derived by crossing “748” (head-splitting-resistant inbred line) and “747” (head-splitting-susceptible inbred line). The map spans 830.9 cM and comprises 270 markers distributed in nine linkage groups, which correspond to the nine chromosomes of B. oleracea. The average distance between adjacent markers was 3.6 cM. A total of six quantitative trait loci (QTLs) conferring resistance to head splitting were detected in chromosome 2, 4, and 6. Two QTLs, SPL-2-1 and SPL-4-1, on chromosomes 2 and 4, respectively, were detected in the experiments over 2 years, suggesting that these two potential loci were important for governing the head-splitting resistance trait. Markers BRPGM0676 and BRMS137, which were tightly linked with head-splitting resistance, were detected in the conserved QTL SPL-2-1 region using bulked segregant analysis. Synteny analysis showed that SPL-2-1 was anchored to a 3.18-Mb genomic region of the B. oleracea genome, homologous to crucifer ancestral karyotype E block in chromosome 1 of Arabidopsis thaliana. Moreover, using a field emission scanning electron microscope, significant differences were observed between the two parental lines in terms of cell structures. Line “747” had thinner cell wall, lower cell density, larger cell size, and anomalous cell wall structure compared with the resistant line “748.” The different cell structures can provide a cytological base for assessing cabbage head splitting.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cabbage (Brassica oleracea L. var. capitata) is an important vegetable crop that is cultivated and consumed worldwide. Cabbage heads are harvested as a leafy vegetable for human consumption. Splits decrease the attractiveness of cabbage heads, lower their quality, and open wounds for insect and disease infection, thereby resulting in economic loss. Splitting or cracking occurs in a wide range of vegetables such as tomato (Peet 1992), Chinese cabbage (Niu et al. 2010), and cabbage (Zhuang et al. 2009; Su et al. 2012), as well as fruit crops such as cherry, apple, and litchi (Christensen 1976; Proctor and Lougheed 1980; Wang et al. 2006). A number of factors involved in the environmental, cultural, and anatomical features, as well as cell structure, have been reported to be among the causes of fruit splitting and cracking (Andersen and Richardson 1982; Emmons and Scott 1997; Savvas et al. 2008). It is commonly believed that cracking or splitting occurs as a result of environmental factors such as rainfall before harvest, sudden increase in the soil moisture content, differentials between day and night humidity, temperature fluctuations, and light exposure. Among the cultural factors, irregular overhead watering is believed to be one of the causes of cracking and splitting. Peet and Willits showed that cracking could be reduced in greenhouse tomatoes by decreasing the amount of water applied (Peet and Willits 1995). Anatomical studies in cherry tomato indicated that the ability of fruit to resist cracking was highly related to the thickness of the cuticular membrane (CM) (Matas et al. 2004). Although many factors have been reported to be responsible for fruit cracking, no reliable cultural measures that prevent cracking have been found. Thus, it has been suggested that selection and breeding of cracking-/splitting-resistant cultivars may be the way to protect crops against fruit cracking.
Cultivar differences in cracking susceptibility are considerable, indicating the role of genetic control in fruit cracking. In tomato, two recessive loci (cr cr and r1 r1) that control radial crack resistance have been identified (Young 1959). Down-regulation of the expression of a beta-galactosidase gene (TBG6) was shown to change tomato fruit morphology and cuticle layer thickening, leading to increased fruit cracking (Moctezuma et al. 2003). Expansins, which are known to be involved in cell wall extension, are now being given more attention in fruit cracking studies. In apple, an expansin gene, MdEXPA3, which was found to be expressed in the pericarp, has been associated with reduced susceptibility to fruit cracking (Kasai et al. 2008). In litchi, two expansin genes, LcExp1 and LcExp2, have been identified, and their expression profiles have been compared between resistant and susceptible cultivars (Wang et al. 2006).
Recently, considerable attention has been paid to the morphological features, physiological characters, and cultivation measures related to head splitting in cabbage (Zeng et al. 2011). However, the genetic base and molecular mechanism involved in regulating cabbage head splitting are still poorly understood. Chiang was the first to determine the mode of inheritance, revealing at least three gene pairs involved in controlling head splitting that displayed mostly additive, but partial, dominance (Chiang 1972). Understanding the genetic basis and developing linked molecular markers for head-splitting resistance will help in selecting and breeding resistant cabbage cultivars.
In this study, we aimed to: (1) identify quantitative trait loci (QTLs) that control resistance to head splitting in cabbage using a new developed genetic linkage map of a F2/3 population; (2) discuss the anatomical features that might be related to head splitting based on a cytological analysis; and (3) develop efficient molecular markers linked to head-splitting resistance for marker-assisted selection (MAS) in breeding programs.
Materials and methods
Plant materials and trait measurement
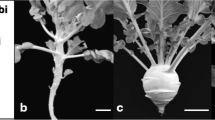
A total of 188 F2 cabbage plants derived from crossing two diverse cabbage inbred lines “747” (head-splitting susceptible) and “748” (head-splitting resistant) were used to construct a genetic linkage map. Parental line “747” is an early head-splitting, long head, and diamondback moth-susceptible type, while “748” is a round head, head-splitting, and diamondback moth-resistant type. The “748” shows no splitting even continuously growing for 3 weeks after maturity. The two parental lines were mid-late-maturing breeding materials. The growing period of “747” and “748” is approximately 100–110 and 110–120 days from germination to maturity, respectively. Each 600 F2 individuals which derived from two different F1 plants, together with each twenty parental lines and F1 plants were grown from 25 February to 20 June in 2011 for detecting the segregation ratios. For the phenotypic screening of the head-splitting resistance trait, 15 F3 plants derived by selfing each of the 188 F2 plants along with parental lines and F1 were grown in two replicates from 17 February to 12 June in 2012 and 24 February to 19 June in 2014. Forty head-splitting-susceptible and head-splitting-resistant plants each were selected from F2 and F2/3 progenies and, along with the two parental lines, were used to establish two bulked pools. All the plants were germinated and grown for 1 month in a glasshouse and then transferred to an open field of the Chungnam National University, Daejeon, South Korea. The spacing was 35 cm between plants and 40 cm between rows and the field was mulched with black plastic film. Spray irrigation system was used in this study to avoid irregular overhead watering. The head-splitting resistance phenotypes were scored as follows: 0, non-head splitting; 1, slight head splitting (head split width ≤20 % of head diameter); 2, medium head splitting (head split width >20 % and ≤40 % of head diameter); and 3, severe head splitting (head split width >40 % of head diameter) (Fig. 1).

Head-splitting phenotypes. a 0, non-head splitting; b 1, slightly head splitting (head split width ≤20 % of head diameter); c 2, medium head splitting (head split width >20 % and ≤40 % of head diameter); and d 3, severe head splitting (head split width >40 % of head diameter)
Scanning electron microscopy
A field emission scanning electron microscope (FESEM; Model: JSM-7000F; JEOL, Germany) was employed to observe the epidermis morphology and anatomical structure. Five individuals were collected at early heading period from each parental line, and the midrib from the same position of the first outer leaf of head was examined under FESEM. Fresh sample sections were fixed in 70 % ethanol before dehydrating in an ethanol series (80, 90, 95, and 100 %) in 30-min steps. The 100 % dry ethanol was changed twice, and the tissues were then dehydrated in an isoamyl acetate–ethanol series (1:2, 1:1, 2:1), with 1 h for each step. The tissues were then immersed in 100 % isoamyl acetate, which was changed once after 1 h, and stored overnight at room temperature before being critical point-dried (CPD; Model: CPD 030 critical point dryer, Bal-Tec, Germany) with CO2 the next day. The CPD specimens were mounted on metallic stubs using double-sided conductive carbon tape and sputter-coated with a thin layer of gold (Model: 208HR High Resolution Sputter Coater, Ted Pella, Inc., USA) before being examined under a FESEM at 10–20 kV.
DNA extraction and marker genotyping
Young leaves were collected from the F2 plants and parental lines for DNA isolation. DNA was extracted using the CTAB method with minor modifications (Li et al. 2010). DNA concentration was determined using a nucleic acid detector and diluted to 5 ng μl−1 for polymerase chain reaction (PCR). BAC-derived simple sequence repeats (SSRs) prefixed by “cnu,” “nia,” “BRPGM” (Li et al. 2010), “BrgMS,” and 272 intron polymorphic (IP) markers (Panjabi et al. 2008) and 332 other SSR markers (prefixed by BRAS, CB, MR, OI, Na, Ni, FITO, and BRMS) from B. napus, B. rapa, and B. oleracea (Uzunova and Ecke 1999; Lowe et al. 2002, 2004; Suwabe et al. 2002, 2006; Piquemal et al. 2005; Radoev et al. 2008; Iniguez-Luy et al. 2008, 2009; Smooker et al. 2011), which have already been mapped in the B. oleracea C-genome chromosomes, were used to screen for polymorphisms in the parental lines. The PCRs all had an initial denaturation of 94 °C for 4 min, followed by 35 cycles of 94 °C for 30 s, the appropriate annealing temperatures for each primer pair, 72 °C for 30 s, and a final step of 72 °C for 5 min. The PCR products were resolved by 8 % polyacrylamide gel electrophoresis, as described by Li et al. (2010).
Linkage and QTL analysis
A genetic map was constructed using JoinMap version 4 (Stam 1993; Van Ooijen 2006). Logarithm of the odds (LOD) scores of 4.0–6.0 were used to assign the markers to linkage groups (LGs), and Kosambi’s mapping function was used to convert the recombination value into the map distance (Kosambi 1944). The threshold for goodness of fit was set to ≤5.0, with a recombination frequency of <0.4 and minimum LOD score of 3.0. QTL mapping of head-splitting resistance was performed using the composite interval mapping (CIM) function provided in Windows QTL Cartographer version 2.5_011 (Wang et al. 2012a). To declare the presence of a QTL, genome-wide threshold values (P = 0.05) were estimated from 1000 permutations of trait data across all genetic intervals. Tests for the presence of a QTL were performed at 2-cM intervals using a 5-cM window and five background cofactors, which were selected by forward regression analysis following (Li et al. 2013). QTL names were designated as SPL (for splitting), followed by chromosome number and QTL number.
Results
Construction of the B. oleracea linkage map
The genetic map of cabbage was constructed using 188 F2 lines, derived from a cross between a head-splitting-susceptible inbred line (747) and a head-splitting-resistant line (748). A polymorphism test between the parental lines detected 322 (8.65 %) clear-cut polymorphic markers from among the 3721 SSR and IP markers that were tested. Of the 322 markers, 52 markers could not be mapped to the chromosome due to highly distorted segregation or their inability to cluster with the LGs. The cabbage map that was constructed had a total of 270 marker loci distributed among nine LGs. The LGs were anchored as chromosomes 1–9 based on previously mapped markers (Uzunova and Ecke 1999; Lowe et al. 2002, 2004; Suwabe et al. 2002, 2006; Piquemal et al. 2005; Radoev et al. 2008; Iniguez-luy et al. 2008, 2009; Smooker et al. 2011). The marker sequences were also aligned with the B. oleracea genomic sequences (http://brassicadb.org/brad/) to confirm their physical position in the B. oleracea C-genome. The linkage map included 151 BAC-anchored SSR markers prefixed by “cnu,” “nia,” “BrgMS,” and “BRPGM” developed in B. rapa, eight Brassica juncea IP markers, 54 B. rapa EST-SSRs prefixed by “ACMP,” and 57 public SSR markers, which covered a total distance of 830.9 cM with an average distance between loci of 3.6 cM (Supplementary Table S1). Chromosome 3 was the longest LG covering 138.1 cM, and chromosome 5 was the shortest covering 55.6 cM (Fig. 2 and Supplementary Table S1).

Nine linkage groups (LGs) on the Brassica oleracea genetic map. “Chr” followed by a number indicates the B. oleracea C-genome chromosomes. Rectangular bars on the right side of each LG indicate head-splitting resistance QTLs
Cytological analysis using a scanning electron microscope
To investigate the possible reason for cabbage head splitting at the cytological level, the leaf epidermal and cell structures of the two parental lines were compared by FESEM. Five individuals were collected at early heading period from each parental line, and the midrib from the same position of the first outer leaf of head was examined under FESEM. The individual size of “747” is much smaller than “748” which also can be identified from the cross-section structure of the midrib under 20× FESEM in Fig. 3a, b. The head-related traits, including head weight, head length, and head width, were measured in F2/3 plants in year 2012 (Supplementary Table S2). Head splitting shown no correlation with head width and negative correlation with head weight and length. The cell size of “747” is bigger than “748” can be observed under 500× FESEM in Fig. 3c, d. Cell number in midrib cross-sections was counted under the FESEM. The total cell numbers on three pointed sections area of midrib from the two parental lines (resistant “748” to susceptible “747”) were 134–101, 138–89, and 179–94. The ratio of the average cell number (resistant “748” to susceptible “747”) was 1.58, suggesting lower cell density in the midrib from the susceptible line “747.” The line “747” had thinner cell wall than “748” that can be identified under 1000× FESEM in Fig. 3e, f. Interestingly, susceptible line “747” showed anomalous cell wall in which the cellulose microfibrils loosen and separate from each other under 20,000× FESEM (Fig. 3g); however, resistant line “748” showed that the cellulose microfibrils were anchored well (Fig. 3h). All the results were observed in multiple experiments. In brief, discovery of the anatomical characteristics of cells in the midrib of the head-splitting-susceptible line “747” compared with the head-splitting-resistant line “748” evidenced the thinner cell wall, lower cell density, larger cell size, and anomalous cell wall structure.

Scanning electron microscope images of the midrib at early heading stage of cabbage. The midrib cross-section of parental line “747” (a) and “748” (b) under ×20 magnification. The midrib cross-section of parental line “747” (c) and “748” (d) under ×500 magnification. The midrib cell cross-section of parental line “747” (e) and “748” (f) under ×1000 magnification showing the thickness of cell wall. The midrib cell wall cross-section of parental line “747” (g) and “748” (h) under ×20,000 magnification. The anomalous cell wall structure was pointed out by black arrow in “747” (g)
QTL mapping of head-splitting trait
The parental lines were chosen for their contrasting characteristics for head-splitting resistance. To detect the segregation ratios, each 600 F2 individual was grown which derived from two different F1 plants. Head-splitting phenotype was investigated when the most of cabbage plants were matured. The segregation ratios of head-splitting resistance to head-splitting susceptible were shown 320–253 and 376–198 in two different F2 families, respectively. Chi-square tests were performed to test the goodness of fit of the observed segregation ratios among F2 plants. The Chi-square tests were not considered to the 1:3 segregation ratios at 0.05 level of probability. These results were values that the head-splitting resistance could be accessed for QTL identification. The F1 was partially resistant to head splitting, indicating the resistance was likely governed by partial dominance genes. In the F2:3 segregating population, a continuous distribution of head-splitting severity was observed in mature cabbage plants grown in the field, suggesting that multiple genes were involved in B. oleracea resistance to head splitting, as shown in the Fig. S1.
QTL mapping, for the head-splitting resistance trait, was conducted using the phenotypic data scored in years 2012 and 2014. To declare the presence of a QTL, genome-wide threshold values (P = 0.05) were estimated from 1000 permutations of trait data across all genetic intervals. The LOD thresholds were set as 3.8 and 3.6 for years 2012 and 2014, respectively. Six different QTLs associated with resistance to head splitting were detected in three chromosomes, chromosome 6, chromosome 2, and chromosome 4. Two of the QTLs (SPL-2-1 and SPL-2-2) were located on chromosome 2, one QTL (SPL-4-1) was on chromosome 4, and three QTLs (SPL-6-1, SPL-6-2 and SPL-6-3) were on chromosome 6. Two of the QTLs, SPL-2-1 and SPL-4-1, were detected in the two experiments conducted in years 2012 and 2014 (Fig. 2). Of the other four QTLs, SPL-6-1 and SPL-6-2 were detected only in 2012, and SPL-2-2 and SPL-6-3 were detected only in 2014 (Table 1). SPL-2-1 explained 10.47 and 14.95 %, and SPL-4-1 explained 7.84 and 8.93 % of the phenotypic variation in the data for the 2 years, respectively (Table 1). We considered these two QTLs as important loci that explain the comparatively high phenotypic variations. The LOD values for the six QTLs ranged from 4.18 to 6.16, and the percentage of phenotypic variance explained by each QTL ranged from 2.18 to 18.5 %.
Flanking marker sequences of the QTLs regions were aligned to the A. thaliana (The Arabidopsis Information Resource, http://arabidopsis.org) and B. oleracea genomes (Liu et al. 2014) to detect interesting B. oleracea genes and to identify corresponding homologous Arabidopsis genes (Supplementary Table S3). We detected four crucifer building blocks (Schranz et al. 2006) that were harboring four of the head-splitting-resistance QTLs. The QTL SPL-2-1 region was anchored to a 3.18-Mb region of the B. oleracea genome that harbors 222 genes that were homologous to A. thaliana E blocks. QTLs SPL-6-1 and SPL-6-2 showed synteny to the E block, and QTL SPL-6-3 showed synteny to C block. Genes located in the SPL-2-1 and SPL-6-1 regions were retrieved from the B. oleracea C-genome (Supplementary Table S4). The functions of most of the genes in these regions are still unknown according to the gene annotation (http://brassicadb.org/brad/).
Identification of markers associated with head-splitting resistance
Bulked segregant analysis (BSA) was performed to identity SSR molecular markers linked to the head-splitting-resistance QTLs. Forty head-splitting-susceptible and 40 head-splitting-resistant selected F2 and F2/3 progenies which were different from the F2 mapping population, and along with the two parental lines, were used to establish two bulked pools. All of the 270 markers that were mapped on the LGs were used to screen for head-splitting resistance. Twenty-six of the markers were selected to screen for tightly linked molecular markers in 80 randomly selected F2 and F2/3 plants that were resistant (40 plants) and susceptible (40 plants) to head splitting. Two of the 26 SSR markers (BRMS137 and BRPGM0676) were found to segregate exactly between the resistant bulks and susceptible bulks, suggesting they might be tightly linked with head-splitting resistance (Fig. 4). The BRPGM0676 and BRMS137 markers were mapped in the conserved SPL-2-1 QTL region (Fig. 2).

Genotyping of SSR markers BRMS137 and BRPGM0676 showing their tightly linked with the head-splitting QTL SPL-2-1 on chromosome 2. Eighty randomly selected F2 and F3 segregating progenies were used. 747 indicates the head-splitting susceptible parent “747” and 748 indicates the head splitting-resistant parent “748”
Discussion
Genetic linkage maps of B. oleracea have been developed using RFLP, AFLP, RAPD, SSR, and SNP markers from F2, doubled haploid (DH) and backcross (BC1) cabbage populations (Slocum et al. 1990; Bohuon et al. 1996; Farinho et al. 2004; Wang et al. 2012b). In the present study, the cabbage genetic map was constructed mainly to identify genomic regions that may control the cabbage head-splitting resistance trait. The genetic map was constructed using mostly B. rapa BAC-derived and EST-derived SSR markers. Marker sequences were aligned with the B. oleracea genome (http://brassicadb.org/brad/), to confirm their physical position in the B. oleracea genome. The B. rapa and B. oleracea genomes have both been sequenced (Liu et al. 2014; Wang et al. 2011) and Arabidopsis genes for plant growing and development, disease resistance, and metabolic related have been annotated. This information could be used to compare and identify homologous genes in the Brassica genomes for different interesting traits.
Inheritance of cabbage head splitting was first reported by Chiang (1972) who showed that more than three gene pairs were involved in controlling the head-splitting trait. Two moderate inbred lines of early-splitting and late-splitting cabbages and their F1, F2 and backcross progenies were used in this study (Chiang 1972). Su et al. (2012) reported that head splitting was controlled by two additive-dominant-epistasis major genes and detected an interaction effect between these two genes. The parental lines used in the present study showed different head-splitting characters. Line “747” was an early head-splitting type, and line “748” showed no head splitting during the whole growing period, while the F1 was partially resistant to head splitting, indicating that the resistance is likely governed by partial dominance genes. The continuous distribution of head-splitting severity was observed in the F2:3 segregating population, suggesting that multiple genes were involved in B. oleracea resistance to head splitting, which is consistent with the findings of a previous study (Su et al. 2012). Many types of fruit cracking have been found in tomato and other fruits, including longitudinal, burst, concentric, star, and radial cracking (Peet 1992). In this study, we observed two types of head splitting in the cabbage population: horizontal and radial splittings (additional data are given in Fig. S2). Most of the heads started splitting from the midrib of head outer leaves and then progressed to the inner leaves. Previous studies in tomato have found that different types of cracking were probably controlled by different sets of genes (Reynard 1960; Walter 1967). In this study, we were concerned only with the splitting degree, not the splitting types. For a better understanding of the genetics of the different splitting types, new populations could be developed using plants that displayed different splitting types.
To date, no unified standard for cabbage head-splitting assessment is available. In a preliminary study, a splitting ratio was used as an identification index (Su et al. 2012). We classified the splitting into four degrees, and consensus QTLs were detected successfully using the average values of the phenotypes, indicating the feasibility of this evaluation method. We also used a splitting index which has been used widely in disease resistance analysis (Piao et al. 2004), but no reasonable QTL was detected. QTLs SPL-2-1 and SPL-4-1 were identified in both 2 years as important loci for conferring head-splitting resistance. Another four loci were only detected in 1-year experiment, which indicated that these loci are affected by environmental factors and cultivation methods. Using BSA combined with SSR marker analysis helped in identifying two SSR markers (BRMS137 and BRPGM0676) that were tightly linked with head-splitting resistance. These markers were located in the central QTL region of SPL-2-1, confirming that SPL-2-1 was an active region that was likely to harbor genes for head-splitting control. These two identified SSR markers could be useful for cabbage head-splitting marker-assisted selection (MAS) in breeding programs. The synteny analysis of the head-splitting resistance QTLs between B. oleracea and A. thaliana, identified QTLs SPL-2-1 (chromosome 2) and SPL-6-1 and SPL-6-2 (chromosome 6) as belonging to the crucifer E block, indicating that common ancestral genomic regions were possibly involved in the evolution of head-splitting resistance genes in B. oleracea by genomic duplication or triplication events (Tang et al. 2012), as reported previously for other traits such as morphological (Li et al. 2013) and clubroot disease traits (Suwabe et al. 2006).
Pioneering studies on tomato have reported some anatomical characteristics of fruit crack susceptible cultivars (Peet 1992), such as large fruit size, low skin tensile strength, and thin skin. Whether head splitting is associated with other morphological traits and whether consensus genomic regions with QTL clusters can be identified will be one of focuses of our future studies. Besides showing differences for head-splitting resistance, the parental lines exhibited contrasting heading habits, head maturity, and leaf morphology characters. This would facilitate the association of splitting resistance to other heading traits by QTL mapping analysis. The leaf samples for the cytological investigation were collected at the early heading stage for head-splitting sign identification. The cytological analysis by FESEM showed significant differences in the microconfiguration and cell structure of the two parental lines. Peet (1992) reported that the large fruit size was easier to crack in tomato. In our study, susceptible line “747” had smaller individual size; however, the single cell size was larger than resistant line “748.” The head splitting showed negative correlation with head weight and length (Supplementary Table S2). It suggested that the large cell size and thin cell wall were more related with head splitting than head size in this study. Cell wall of susceptible line appears thinner and anomalous with the cellulose microfibrils loosening and separating from each other (Fig. 3g), thereby inducing splitting. Another possible reason for splitting is the water absorption capacity of the cells. The cell sizes were larger in the susceptible line, which may have increased their capacity for water absorption. Consequently, the water content in the leaves would increase sharply, leading to splitting. Therefore, controlling irrigation during maturity periods is an important cultural practice to prevent splitting. In a fruit cracking anatomical study, the thickness of the cuticular layer was found to be associated with cracking (Huang et al. 2004; Demirsoy and Demirsoy 2004). However, in our study, no differences in the thickness of the cuticular layer were observed in the parental lines.
Recently, expansins involved in cell wall extension have been reported to be closely related with fruit cracking in litchi, apple, and fruits (Wang et al. 2006; Kasai et al. 2008). In our study, the cell wall of the susceptible line depicted anomalous structure with the cellulose microfibrils loosening and separating from each other (Fig. 3g) which may be associated with expansins genes. We analyzed the genes that fell into the two important QTL regions, and more than two hundred genes were obtained, due to the relatively large QTL region detected by primary mapping population. Even though one cabbage gene Bol028570 lies within the QTL region of chromosome 2 which was orthologous to an Arabidopsis expansin gene (At1g69530) (Supplementary Table S4), whether this expansin gene or other candidate genes do indeed contribute to the genetic control of head splitting or change the cell wall structure still needs to be elucidated. Further studies are required to: (1) isolate candidate genes from the two parental lines to compare their gene structures and to design gene-specific SNP or Indel markers linked with head splitting; (2) conduct genomic and transcriptomic sequencing of the two parental lines to detect other novel genes, as well as the signal transduction pathway that may be involved in head splitting; and (3) fine map and clone the candidate head-splitting resistance genes in the two major QTL genomic regions. Studies of this kind will provide a fundamental understanding of the mechanisms of head-splitting resistance and help in the breeding of head-splitting resistance cultivars in cabbage.
References
Andersen PC, Richardson DG (1982) A rapid method to estimate fruit water status with special reference to rain cracking of sweet cherries. J Am Soc Hortic Sci 107:441–444
Bohuon E, Keith D, Parkin I, Sharpe A, Lydiate D (1996) Alignment of the conserved C genomes of Brassica oleracea and Brassica napus. Theor Appl Genet 93(5):833–839
Chiang MS (1972) Inheritance of head splitting in cabbage (Brassica oleracea L. var. Capitata L.). Euphytica 21:507–509
Christensen JV (1976) Cracking in cherries. Dan J Plant Soil Sci 80:289–324
Demirsoy L, Demirsoy H (2004) The epidermal characteristics of fruit skin of some sweet cherry cultivars in relation to fruit cracking. Pak J Bot 36(4):725–731
Emmons CLW, Scott JW (1997) Environmental and physiological effects on cuticle cracking in Tomato. J Am Soc Hortic Sci 122:797–801
Farinho M, Coelho P, Carlier J, Svetleva D, Monteiro A, Leitao J (2004) Mapping of a locus for adult plant resistance to downy mildew in broccoli (Brassica oleracea var. Italica). Theor Appl Genet 109(7):1392–1398
Huang XM, Wang HC, Li JG et al (2004) Pericarp structure in relation to fruit cracking resistance in litchi (Litchi chinensis Sonn.). Acta Hortic 632:131–137
Iniguez-Luy FL, Voort AV, Osborn TC (2008) Development of a set of public SSR markers derived from geomic sequence of a rapid cycling Brassica oleracea L. genotype. Theor Appl Genet 117:977–985
Iniguez-luy FL, Lukens L, Farnham MW, Amasion RM, Osborn TC (2009) Development of public immortal mapping populations, molecular markers and linkage maps for rapid cycling Brassica rapa and B. oleracea. Theor Appl Genet 120:31–43
Kasai S, Hayama H, Kashimura Y, Kudo S, Osanai Y (2008) Relationship between fruit cracking and expression of the expansin gene MdEXPA3 in ‘Fuji’ apples (Malusdomestica Borkh.). Sci Hortic 116:194–198
Kosambi DD (1944) The estimation of map distances from recombination values. Ann Eugen 12:172–175
Li X, Ramchiary N, Choi SR, Nguyen VD, Hossain MJ, Yang HK, Lim YP (2010) Development of high density integrated reference genetic linkage map for multinational Brassica rapa genome sequencing project. Genome 53:939–947
Li X, Ramchiary N, Dhandapani V, Choi SR, Yang HK, Nou IS, Yoon MK, Lim YP (2013) Quantitative trait loci mapping in Brassica rapa revealed the structural and functional conservation of genetic loci governing morphological and yield component traits in the A, B, and C subgenomes of Brassica species. DNA Res 20:1–16
Liu S, Liu Y, Tong C et al (2014) The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat Commun 5:3930. doi:10.1038/ncomms4930
Lowe AJ, Jones AE, Raybould AF, Trick M, Moule CJ, Edwards KJ (2002) Transferability and genome specificity of a new set of microsatellite primers among Brassica species of the U triangle. Mol Ecol Notes 2:7–11
Lowe AJ, Moule C, Trick M, Edwards KJ (2004) Efficient large-scale development of microsatellites for marker and mapping applications in Brassica crop species. Theor Appl Genet 108:1103–1112
Matas AJ, Cobb ED, Paolillo DJ, Niklas KJ (2004) Crack resistance in cherry tomato fruit correlates with cuticular membrane thickness. HortScience 39:1354–1358
Moctezuma E, Smith DL, Gross KC (2003) Antisense suppression of a beta-galactosidase gene (TBG6) in tomato increased fruit cracking. J Exp Bot 54:2025–2033
Niu N, Zheng C, Zhang L, Xu Y, Zhang L, Fu W (2010) Inheritance and RAPD marker of head-splitting trait in Chinese cabbage. China Veg 14:44–48
Panjabi P, Jagannath A, Bisht NC, Padmaja KL, Sharma S, Gupta V, Pradhan AK, Pental D (2008) Comparative mapping of Brassica juncea and Arabidopsis thaliana using intron polymorphism (IP) markers: homoeologous relationships, diversification and evolution of the A, B and C Brassica genomes. BMC Genom 9:113
Peet MM (1992) Fruit cracking in tomato. HortTechnol 2:216–223
Peet MM, Willits DH (1995) Role of excess water in tomato fruit cracking. HortScience 30:65–68
Piao ZY, Deng YQ, Choi SR, Park YJ, Lim YP (2004) SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage (Brassica rapa ssp. pekinensis). Theor Appl Genet 108:1458–1465
Piquemal J, Cinquin E, Couton F, Rondeau C, Seignoret E, Perret D, Villeger MJ, Vincourt P, Blanchard P (2005) Construction of an oilseed rape (Brassica napus L.) genetic map with SSR markers. Theor Appl Genet 111:1514–1523
Proctor JTA, Lougheed EC (1980) Cracking of Golden Russet apples. Can Plant Dis Surv 60:55–58
Radoev M, Becker HC, Ecke W (2008) Genetic analysis of heterosis for yield and yield components in rapeseed (Brassica napus L.) by quantitative trait locus mapping. Genetics 179:1547–1558
Reynard GB (1960) Breeding tomatoes for resistance to fruit cracking. Proc Plant Sci FESEMinar. Campbell Soup Co, Camden, pp 19–36
Savvas D, Ntatsi G, Passam HC (2008) Plant nutrition and physiological disorders in greenhouse grown tomato, pepper and eggplant. Eur J Plant Sci Biotechnol 2:45–61
Schranz ME, Lysak MA, Mitchell-Olds T (2006) The ABC’s of comparative genomics in the Brassicaceae: building blocks of crucifer genomes. Trends Plant Sci 11:535–542
Slocum M, Figdore S, Kennard W, Suzuki J, Osborn T (1990) Linkage arrangement of restriction fragment length polymorphism loci in Brassica oleracea. Theor Appl Genet 80(1):57–64
Smooker AM, Wells R, Morgan C, Beaudoin K, Fraser CF, Bancroft I (2011) The identification and mapping of candidate genes and QTL involved in the fatty acid desaturation pathway in Brassica napus. Theor Appl Genet 122:1075–1090
Stam P (1993) Construction of integrated genetic linkage maps by means of a new computer package: JoinMap. Plant J 3:739–744
Su YB, Liu YM, Fang ZY, Yang LM, Zhuang M, Zhang YY, Zhang XL, Sun PT (2012) Genetic analysis of head-splitting resistance traits in cabbage. Acta Hortic Sin 39(8):1482–1490
Suwabe K, Iketani H, Nunome T, Kage T, Hirai M (2002) Isolation and characterization of microsatellites in Brassica rapa L. Theor Appl Genet 104:1092–1098
Suwabe K, Tsukazaki H, Iketani H, Hatakeyama K, Kondo M, Fujimura M, Nunome T, Fukuoka H, Hirai M, Matsumoto S (2006) Simple sequence repeat-based comparative genomics between Brassica rapa and Arabidopsis thaliana: the genetic origin of clubroot resistance. Genetics 173:309–319
Tang H, Woodhouse MR, Cheng F, Schnable JC, Pedersen BS, Conant G, Wang X, Freeling M, Pires JC (2012) Altered patterns of fractionation and exon deletions in Brassica rapa support a two-step model of paleohexaploidy. Genetics 190:1563–1574
Uzunova MI, Ecke W (1999) Abundance, polymorphism and genetic mapping of microsatellites in oilseed rape (Brassica napus L.). Plant Breed 118:323–326
Van Ooijen JW (2006) Joinmap 4, software for the calculation of genetic linkage maps in experimental populations. Kyazma BV, Wageningen
Walter JC (1967) Hereditary resistance to disease in tomato. Annu Rev Phytopathology 5:131–162
Wang Y, Lu W, Li J, Jiang Y (2006) Differential expression of two expansin genes in developing fruit of cracking-susceptible and -resistant litchi cultivars. J Am Soc Hortic Sci 131:118–121
Wang X, Wang H, Wang J et al (2011) The genome of the mesopolyploid crop species Brassica rapa. Nat Genet 43:1035–1039
Wang S, Basten CJ, Zeng Z-B (2012a) Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC. http://statgen.ncsu.edu/qtlcart/WQTLCart.htm
Wang W, Huang S, Liu Y et al (2012b) Construction and analysis of a high density genetic linkage map in cabbage (Brassica oleracea L. var.capitata). BMC Genom 13:523
Young HW (1959) Inheritance of radial fruit cracking in a tomato cross. Proc Fla State Hort Soc 72:207–210
Zeng A, Liu Y, Fang Z, Yan J (2011) Research progress of head splitting on cabbage (Brassica oleracea L. var. capitata L.). J Plant Genet Resour 2:307–310
Zhuang M, Zhang Y, Fang Z, Liu Y, Yang L, Sun P (2009) Studies on combining ability and heritability of splitting resistance characteristic in cabbage (Brassica oleracea var. capitata L.). China Veg 1:12–15
Acknowledgments
This work was supported by the Golden Seed Projet (No. 213002-04-1-SB110), Ministry of Agriculture, Food and Rural Affairs (MAFRA), Ministry of Oceans and Fisheries (MOF), Rural Development Administration (RDA), and Korea Forest Service (KFS).
Author information
Authors and Affiliations
Corresponding author
Additional information
Wenxing Pang and Xiaonan Li have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Fig. S1
Continuous distribution of head-splitting severity displayed in the F2:3 segregating population in year 2012 (A) and 2014 (B) experiment, respectively. (PPTX 72 kb)
Rights and permissions
About this article

Cite this article
Pang, W., Li, X., Choi, S.R. et al. Mapping QTLs of resistance to head splitting in cabbage (Brassica oleracea L.var. capitata L.). Mol Breeding 35, 126 (2015). https://doi.org/10.1007/s11032-015-0318-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-015-0318-1