
Abstract
The most common form of the disease caused by Toxoplasma gondii (T. gondii) is latent toxoplasmosis due to the formation of tissue cysts in various organs, such as the brain. Latent toxoplasmosis is probably a risk factor in the development of some neuropsychiatric disorders. Behavioral changes after infection are caused by the host immune response, manipulation by the parasite, central nervous system (CNS) inflammation, as well as changes in hormonal and neuromodulator relationships. The present review focused on the exact mechanisms of T. gondii effect on the alteration of behavior and neurotransmitter levels, their catabolites and metabolites, as well as the interaction between immune responses and this parasite in the etiopathogenesis of psychiatric disorders. The dysfunction of neurotransmitters in the neural transmission is associated with several neuropsychiatric disorders. However, further intensive studies are required to determine the effect of this parasite on altering the level of neurotransmitters and the role of neurotransmitters in the etiology of host behavioral changes.
Similar content being viewed by others

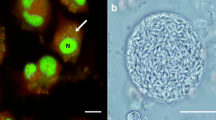
Avoid common mistakes on your manuscript.
Introduction
Mental and behavioral disorders are considered major public health problems. These disorders affect 10–16% of the world population every year (Fekadu et al. 2010; Wang et al. 2006). One of the major causes of disability and mortality around the world is neurological disorders. Population growth and aging population significantly increase the burden of neurological disorders in the past 25 years worldwide. Generally, the rate of mortalities and number of disability-adjusted life years (DALYs) resulted from neurological disorders increased by 36.7% and 7.4% within 1990 and 2015 (Feigin et al. 2017). The results of a systematic analysis on the global burden of mental diseases showed that neurological disorders were responsible for 10.2% of global DALYs (i.e., 250.7 million) and 16.8% of global mortalities (i.e., 9.4 million) (Feigin et al. 2017). The causes of mental illnesses are often unknown; however, biological, psychological, and environmental factors play a role in causing psychiatric disorders individually or in combination. Among the biological factors, genetic factors, traumatic brain injury, noninfectious diseases, such as epilepsy, birth trauma, and oxygen starvation are important. Likewise, the influence of gut flora on brain function has shown a two-way interaction between the gut and brain that affects an individual’s mood, motivation, decision-making, and cognitive abilities (Tyebji et al. 2019). Therefore, microorganisms that cause gastrointestinal diseases may affect mental health (Bolton and Robertson 2016). In addition, infectious diseases can be among the causes of mental disorders. Infectious diseases, such as malaria, syphilis, Legionnaire’s disease, diphtheria, typhoid, herpes, and human immunodeficiency virus (HIV), cause psychiatric symptoms (Bolton and Robertson 2016). One of these diseases is toxoplasmosis, which is associated with altered neurological function (Tedford and McConkey 2017).
Toxoplasma gondii (T. gondii) is an obligate intracellular protozoan infecting almost all warm-blooded animals, including mammalian and several species of birds (as intermediate hosts), and its definitive host is a cat family (i.e., Felidae) (Fuglewicz et al. 2017; McConkey et al. 2013). T. gondii possesses a complex life cycle, which is composed of the three infectious phases: a sexually replicative stage in definitive hosts, an asexual replicative stage in the intermediate hosts, and an exogenous stage (oocysts containing sporozoites) (Montoya and Liesenfeld 2004). During these stages, the parasite is observed in three infectious forms, including tachyzoite present in body fluids during acute infection, bradyzoite contained within tissue cysts during chronic infection, and oocyst (excreted from the feces of cats) (Tenter et al. 2000). The seroprevalence of T. gondii infection measured by specific anti-Toxoplasma immunoglobulin G (IgG) antibodies widely varies within the range of 1–100% worldwide. Variations in prevalence depend on the environmental and socioeconomic conditions, including climatic and geographic factors, hygiene standards, feeding and cultural habits, as well as professions (Dubey 1998; Dubey and Beattie 1988; Furtado et al. 2011; Tenter et al. 2000). The lowest seroprevalence (about 1%) was reported in some countries in the Far East, and the highest seroprevalence (less than 90%) was reported in some parts of European and South American countries (Flegr et al. 2014a). Different factors, such as the strain and virulence of parasite, number of ingested organisms, immunity status, gender, and genetic background of the host, are crucially important in the progress of infection (Montoya and Liesenfeld 2004). The most important routes of infection are the consumption of oocysts present in the environment, drinking contaminated water, ingestion of tissue cysts in raw or undercooked meat of intermediate hosts, organ transplantation, and transplacental transmission of tachyzoites (Carruthers and Suzuki 2007; da Silva and Langoni 2009; Flegr et al. 2014b; Tedford and McConkey 2017). It is believed that if most healthy individuals are infected with this parasite, they are generally asymptomatic or show a subclinical course with minor symptoms (Montoya and Liesenfeld 2004). Primary infection in pregnant women can be transmitted to the fetus and lead to spontaneous abortion or childbirth (Flegr et al. 2014a). Toxoplasmosis can cause a variety of life-threatening clinical signs in cases of congenital infection which can result in a wide diversity of manifestations in neonates, such as hydrocephalus, microcephaly, and retinochoroiditis, or immunocompromised patients, including HIV-positive individuals, cancer patients, and transplant recipients (Ahmadpour et al. 2014; Jones et al. 2003). In the aforementioned subjects, acute infection or reactivation of latent toxoplasmosis might develop severe clinical complications with poor prognoses, such as toxoplasmic encephalitis, brain abscess, pneumonitis, myocarditis, and chorioretinitis (Abedalthagafi et al. 2009; Ahmadpour et al. 2014; Sutterland et al. 2015). Some diseases, such as ocular toxoplasmosis, occur both in immunocompetent and immunocompromised individuals (Furtado et al. 2011; Klaren and Kijlstra 2002; Montoya and Liesenfeld 2004). Parasite through the Trojan horse mechanism can migrate to the immunoprivileged organs, eyes, testes, and especially brain (Jaroslav Flegr 2015). This parasite possibly enters the brain as early as 7 days after the infection (Konradt et al. 2016). Lesions in the brain by interfering with the functions of the brain in the region around the lesion through paracrine secretions or mass effects can appear as behavioral symptoms (Mentzer et al. 2012). The results of a study carried out on mice showed that the cyst cannot directly cause behavioral changes by disrupting the surrounding tissue (Gatkowska et al. 2012). The possible ability of the parasite to change human behavior can be influenced by the factors, such as one's genetic susceptibility, time of the person’s life at which the infection occurs, severity of the initial acute infection, ingestion stage of the parasite, as well as the exact number and location of parasitic cysts in the brain at the chronic stage of infection (Henriquez et al. 2009; McConkey et al. 2013).
Flegr et al. in 2014 studied the seroprevalence of latent toxoplasmosis and its relationship with mortality and disability rates of specific diseases in 88 countries around the world. The analyses revealed that DALYs associated with 23 of the 128 analyzed diseases correlated with the prevalence of toxoplasmosis (18 positive and 5 negative), and the other 12 cases showed a positive trend (Flegr et al. 2014a). In addition, the probable association between T. gondii infection and mental disorders has been investigated in several systematic reviews and meta-analyses. The most common of these disorders are major depression disorder (MDD) (Nayeri Chegeni, et al. 2019a; Sutterland et al. 2015), schizophrenia (SZ) (Sutterland et al. 2015; E. F. Torrey et al. 2007), bipolar disorder (BD) (de Barros et al. 2017; Sutterland et al. 2015), addiction (Sutterland et al. 2015), obsessive–compulsive disorder (OCD) (Nayeri Chegeni et al. 2019b), Alzheimer's disease (AD) (Bayani et al. 2019; Nayeri Chegeni, et al. 2019c), Parkinson’s disease (PD) (Bayani et al. 2019), attention-deficit hyperactivity disorder (ADHD) (Nayeri et al. 2020a), epilepsy (Ngoungou et al. 2015; Sadeghi et al. 2019), suicide attempts (Sutterland et al. 2019; Amouei et al. 2020), traffic accidents (Gohardehi et al. 2018; Sutterland et al. 2019), autism (Nayeri et al. 2020b), and headache (Nayeri et al. 2020c). The results of these studies showed that chronic T. gondii infection is a factor associated with SZ, BD, OCD, AD, addiction, epilepsy, suicide attempts, autism, headache and traffic accidents but not with MDD, ADHD, and PD. T. gondii infection may also be associated with some other disorders, but further studies are needed to confirm these relationships. For example, a case–control study showed that chronic toxoplasmosis was not associated with sleepiness and sleep apnea syndrome severity in obese patients suspected of sleep apnea syndrome (Dard et al. 2020).
There are several mechanisms used by the parasite to cause the disease. The first mechanism is the alteration of different neurotransmitters pathways indicating that T. gondii has genes encoding proteins homologous to aromatic amino acid tyrosine hydroxylase (TH) and dopamine receptor D2 (D2R). The parasite also increases dopamine synthesis and tryptophan degradation and decreases serotonin synthesis (Henriquez et al. 2009). The second mechanism used by the parasite is the induction of the immune responses (Henriquez et al. 2009), and the third is the induction of endocannabinoids (Melamede 2009). Accordingly, different changes, such as anatomical, immunological, and pathological, as well as changes in neurotransmitter levels and gene expressions, may occur during the brain infection with T. gondii. Moreover, some of these changes may result in various neurological diseases (Abdoli et al. 2014). The ability of the parasite to influence neurotransmitter pathways is one of the most important mechanisms. Firstly, in the 1970s, Stibbs (Stibbs 1985) described changes in different neurotransmitter concentrations with T. gondii infection and reported a decrease in norepinephrine and serotonin synthesis, as well as increasing dopamine synthesis in the brain of infected mice with chronic infection. According to the above-mentioned content and importance of the role of neurotransmitters in causing mental disorders, the present study aimed to review the current state of knowledge regarding the role of neurotransmitters and increasing or decreasing their levels in the development of mental disorders. In addition, this study aimed to identify several gaps that can improve the diagnosis and treatment of mental health problems.
Neurotransmitter systems
The nervous system includes billions of specialized cells called neurons. Neurons are cells that establish chemical communication in the brain. A neuron has two ends (i.e., one axon and one dendrite). The normal functioning of the central and peripheral nervous system requires efficient communication between neurons (Patri 2019). Neurotransmitters are a diverse group of chemical compounds in the brain that serves as the mediator for the transmission of nerve impulses from the presynaptic site of one neuron to the postsynaptic site of another neuron via synapses (Leite et al. 2017). These neurotransmitters are stored in small packets called synaptic vesicles in axons (or presynaptic neurons) (Ayano 2016). The arrival of a nerve impulse or an action potential at the nerve terminal leads to the release of neurotransmitters (Patri 2019). The decreased release of neurotransmitters results from the loss of neurons and synapses leading to learning and memory problems (Parlog et al. 2015). The nuclear factor kappa light-chain enhancer of activated B cells (NF-κB) pathway is an intracerebral inflammatory pathway activated in the brain of mice chronically infected with T. gondii. The overexpression of cytokines, such as tumor necrosis factor (TNF), interleukin 1 beta (IL-1β), and Toll-like receptors, activated the NF-κB pathway (Courtois and Gilmore 2006; Wang et al. 2019), and the activation of this pathway may enhance the inflammatory response to the CNS injury. Excessive immune responses can induce neuronal apoptosis and lead to decreased synaptic density in the brain (Zhang et al. 2014). Substances that act as neurotransmitters can be classified into different groups as follows:
-
1.
Amino acids: Glycine, glutamate, aspartic acid, and gamma-aminobutyric acid (GABA).
-
2.
Amino acid-derived amines: Norepinephrine (noradrenaline), epinephrine (adrenaline), dopamine, serotonin (5-HT), and histamine.
-
3.
Gasotransmitters: Nitric oxide (NO), carbon monoxide (CO), and hydrogen sulfide (H2S).
-
4.
Peptides: Substance P and endorphins.
-
5.
Purines: Adenosine triphosphate.
-
6.
Others: Acetylcholine (Abg Abd Wahab et al. 2019; Mustafa et al. 2009; Patri 2019).
Neurotransmitters are divided into three groups of excitatory, inhibitory, and modulatory based on their effects on signal transmission (Tyebji et al. 2019). Amino acid neurotransmitters and amino acid-derived amines are inhibitory and excitatory (Patri 2019). Some neurotransmitters (e.g., glutamate, serotonin, acetylcholine, epinephrine, and norepinephrine) are excitatory; however, other neurotransmitters (e.g., glycine and GABA) are inhibitory (Abg Abd Wahab et al. 2019; Patri 2019). Unlike other neurotransmitters, dopamine can act as both inhibitory and excitatory depending on its position in the brain and which receptor it connects to (Nakamura et al. 2014). Amino acid neurotransmitters play a role in fast synaptic transmission; nevertheless, amino acid-derived amines are involved in slow synaptic transmission (Patri 2019). Out of the aforementioned groups of neurotransmitters, amino acids (i.e., glutamate and GABA), amino acid-derived amines (i.e., norepinephrine, epinephrine, dopamine, and serotonin), and others (i.e., acetylcholine and NO) are involved in the CNS infection by T. gondii. Therefore, this article reviewed the possible roles and mechanisms of these neurotransmitters in mental disorders caused by T. gondii infection (Fig. 1).

Potential mechanisms utilized by Toxoplasma gondii to alter the level of neurotransmitters and intermediate host behavior
Dopamine
Dopamine (4-[2-aminoethyl] benzene-1,2-diol) is one of the most important catecholamine neurotransmitters in the mammalian CNS (Fig. 2). Dopamine is a chemical material released by one neuron to send signals to other neurons. Furthermore, dopamine is a chemical messenger and plays a key role in the functioning of the kidney, as well as hormonal and cardiovascular systems (van Staden and van Staden 2012). In 1910, dopamine was firstly synthesized by George Barger and James Ewens (van Staden and van Staden 2012). In addition, the function of dopamine as a neurotransmitter was initially recognized in 1958 (van Staden and van Staden 2012). The cell bodies of the dopaminergic neurons are concentrated in several areas of the brain, including the ventral tegmental area, substantia nigra, and retrorubral field, and projected to the basal ganglia, limbic regions, olfactory bulbs, hippocampus, and cortex. The prefrontal cortex involved in planning, decision-making, problem-solving, and coordinating performance in humans is rich in dopamine (Diamond et al. 2004). Dopaminergic neurons appear in humans and rats early within 6–8 weeks of pregnancy and 10–15 days of gestation, respectively. The appearance of these neurons occurs later in males than that reported for females (Herlenius and Lagercrantz 2004). There are five known dopamine receptors (DR) (i.e., D1, D2, D3, D4, and D5); however, D1R and D2R are two main types used by the brain (Herlenius and Lagercrantz 2004). The D1R mediates excitatory signaling pathways; nonetheless, D2R mediates inhibitory signaling pathways (Nakamura et al. 2014). Dopamine is involved in many brain circuits in the nervous systems associated with mediating motor control, emotional and cognitive aspects, arousal, reward, and feeding behavior (Abg Abd Wahab et al. 2019). The presence of dopamine neurotransmitter is necessary for the long-term regulation of brain functions (Tyebji et al. 2019). Dopamine is an important neurotransmitter that plays a role in the etiology of several human disorders, such as AD, PD, SZ, and depression (Davis et al. 1991). An increase in dopamine levels in the brains of the mice infected with T. gondii was described in 1985 (Stibbs 1985). Dopamine levels decrease in some psychiatric disorders and increase in others; for example, decreased dopamine levels are often observed in patients with SZ (Davis et al. 1991); however, the levels of this neurotransmitter increase in patients with MDD, AD, and PD (Wolfe et al. 1990). The concentration of dopamine increased in the acute phase of T. gondii infection in mice (Gatkowska et al. 2013). In addition, this increase in concentration was observed in the chronic stage, as well as the tissue and their surroundings (Prandovszky et al. 2011). On the other hand, since dopamine increases the number of T. gondii tachyzoites in cultured fibroblasts, it can be said that neurotransmitters may manipulate the parasite (Stibbs 1985). The antipsychotic dopamine-blocking agent haloperidol (i.e., a typical antipsychotic known dopamine D2 antagonist) used in the treatment of SZ and other psychiatric disorders can inhibit the in vitro replication of T. gondii (Webster et al. 2006). The results of in vitro studies indicated that dopamine may play a variety of roles in the brain, including reproduction, chemotherapy, infection efficiency, and conversion of cyst stage to tachyzoite (Strobl et al. 2012). Tissue cysts have been identified throughout the brain, although there are a greater number of cysts in the amygdala and nucleus accumbens (Gonzalez et al. 2007; Vyas 2015). These dopamine-containing regions play important functions, including basal ganglia in the control of movements, nucleus accumbens and hippocampus in learning, pleasure, memory, consciousness, dependence, information processing, and language (Prandovszky et al. 2011; Taupin 2007). In addition, the amygdala is associated with a range of cognitive functions, namely motivation, emotion, learning, cognition, memory, attention, perception, and fear (Baxter and Murray 2002; Prandovszky et al. 2011). Most of the effects of T. gondii infection are reported in the hippocampus and amygdala (Dalimi and Abdoli 2012). The T. gondii genome includes two aromatic acid hydroxylase genes (i.e., AAH1 and AAH2) that encode tyrosine and phenylalanine hydroxylases (Gaskell et al. 2009). These enzymes catalyze phenylalanine to tyrosine and tyrosine to 3,4-dihydroxyphenylalanine (L-Dopa), as the direct precursor of dopamine, that could directly affect dopamine and/or serotonin biosynthesis. These enzymes result in the alteration of the host behavior (Elsheikha and Zhu 2016). In addition, it has been reported that TH identified within T. gondii cysts is encoded by the parasite itself (Prandovszky et al. 2011). The expression of the AAH2 gene increased in the dormant bradyzoite stage (Webster et al. 2006); however, the results of a study showed that the disruption of the AAH2 gene had no effect on the growth and differentiation of the parasite, as well as the global dopamine levels of the brain (Z. T. Wang et al. 2015). Another study reported that the AAH gene plays a role in the enhancement of the ability to transmit infection in cats (Z. T. Wang et al. 2017). This section addresses several mechanisms by which the parasite may alter dopamine levels.

Structure of dopamine
Role of cytokines and enzymes
Macrophages and dendritic cells (DCs) secrete interleukin-12 (IL-12) during the stimulation of T. gondii antigen. Toll-like receptors (TLR)-11/TLR-12 dimer is important for DC response and IL-12 production (Koblansky et al. 2013; Andrade et al. 2013). In fact, mice lacking TLR-11 produce lower levels of IL-12 (Yarovinsky et al. 2005; Sher et al. 2017), and a lack of TLR-12 results in a loss of resistance to toxoplasmosis as well as a deficiency in the MyD88 encoding gene (Yarovinsky et al. 2005; Sher et al. 2017). Mice deficient in the MyD88 adapter molecule are highly sensitive to toxoplasmosis (Koblansky et al., 2013). The IL-12 maintains a long-lasting immune response in the chronic stage of infection. The IL-12 also activates the production of interferon-gamma (IFN-γ) by natural killer (NK) cells and T cells (Hunter et al. 1995; Yap et al. 2000). The IFN-γ is the most important cytokine regarding resistance to T. gondii that can eliminate the parasite and inhibit its proliferation through specific mechanisms (Dupont et al. 2012). One of these mechanisms is releasing indoleamine 2,3-dioxygenase (IDO) (F. Wang and Lidow 1997). This enzyme induces the degradation of tryptophan (Pfefferkorn 1984), and kynurenic acid (KA) and quinolinic acid (QA) are the products of tryptophan catabolism in the brain that increase oxidative stress, damaging cells, and apoptosis (Schwarcz et al. 2012). In human patients, high levels of QA and KA are associated with many neurological disorders, such as SZ and depression (Notarangelo et al. 2014; Schwarcz and Pellicciari 2002). In addition, in baseline conditions, kynurenine is mainly metabolized to KA, and the accumulation of this catabolite in the CNS due to its N-methyl-D-aspartate (NMDA) antagonistic properties can alter dopamine metabolism (Hilmas et al. 2001; Mortensen et al. 2007). Also, KA is an antagonist to alpha 7 nicotinic acid receptor and increases the release of amphetamine-stimulated dopamine in the striatum (Elsheikha and Zhu 2016). A seven-fold increase in KA has been reported in the brains of animals infected with T. gondii (Tedford and McConkey 2017). Medications known as dopamine-releasing agents, such as amphetamine, can exacerbate psychotic symptoms in patients and cause psychiatry in normal people (Laruelle et al. 1996). Chronic T. gondii infection can produce an abnormal response to dopamine stimulant, and behavioral deficits in response to amphetamine have been observed in mice infected with T. gondii (Xiao et al. 2016). Another enzyme that may affect dopamine levels is catechol-O-methyltransferase. This enzyme as a candidate gene for some psychiatric disorders, such as suicidal behaviors, is the major dopamine-degrading enzyme in the prefrontal cortex (Cáceda 2014). However, the expression of this gene was not significantly different in Toxoplasma-infected mice, compared to that reported for the controls (Xiao et al. 2014). Monoamine oxidase A (MAOA) is an enzyme that plays an important role in the metabolism of biogenic amines such as dopamine; in rodents, the infection with T. gondii results in the decrease of this enzyme (Cáceda 2014; Xiao et al. 2014). The decreased activity of this enzyme is associated with personality features, such as impulsivity and aggression. In addition, the increased activity of this enzyme leads to a decrease in dopamine levels that is associated with mood disorders and aggressive behaviors (Cáceda 2014).
Another point is that IFN-γ causes tissue damage via mechanisms, such as the production of NO and the destruction of dopamine-producing neurons (Kusbeci et al., 2011). This cytokine and other pro-inflammatory cytokines (e.g., IFN-α and IFN-β) activate guanosine triphosphate cyclohydrolase 1 (GTP-CH1) that leads to the increased production of neoprene [increasing the concentration of NO via the creation of tetrahydrobiopterin (BH4)] and decreased levels of amino acids phenylalanine and tyrosine. The two above-mentioned amino acids are precursors for the biosynthesis of dopamine. The BH4 plays a role in the synthesis of dopamine (Neurauter et al. 2008). Cytokines can decrease dopamine accessibility in related brain areas by reducing the concentrations of BH4 (Kitagami et al. 2003). Moreover, cytokines and inflammatory mediators show that Toxoplasma-induced encephalitis may affect neurotransmitters through the activation of mitogen-activated protein kinases (MAPK) (Zhu et al. 2005; Zhu et al. 2010). The activation of MAPK pathway can have a dual effect on dopamine. Also, murine TLR-11 and 12 of DCs, recognize a Toxoplasma pathogen-associated molecular pattern (Profilin) to primarily signal through the adaptor protein MyD88, leading to the activation of MAPK (Koblansky et al., 2013; Yarovinsky et al. 2005; Sanecka and Frickel 2012; Arnold-Schrauf et al. 2015). Profilin is essential for the invasion and active egress of T. gondii from the cells (Egan et al. 2009; Kucera et al. 2010; Plattner et al. 2008). Imiquimod (an efficient immunomodulatory drug against certain viral and parasitic infections) upregulates the expression of TLRs 7, 11, and 12, following interconversion from bradyzoites to tachyzoites. As a result, the MyD88 pathway is activated, leading to the induction of an immune response to control Toxoplasma foci (Hamie et al., 2021).
The MAPK signaling results in reduced dopamine recycling and overstimulation of its receptors (Zhu et al. 2005, 2010). The decreased synthesis of dopamine is associated with the disruption of sleep, fatigue, as well as gastrointestinal and musculoskeletal disorders (Hsu et al. 2014). Another point about dopamine is the presence of a relationship between dopamine and sex hormones, such as testosterone, both of which increase during latent toxoplasmosis (Hull et al. 2004). Testosterone increases the synthesis of NO in the medial preoptic area (Hull et al. 2004). The mediator of the connection between testosterone and dopamine is NO because NO and other inflammatory cytokines, such as IL-2 and IL-6, increase dopamine levels (Hull et al. 2004; Prast and Philippu 2001; West et al. 2002). Moreover, increasing testosterone enhances dopamine levels (Hull et al. 1995). Another mechanism by T. gondii for impaired dopamine signaling is the in vitro upregulation of microRNA-132 (Tyebji et al. 2019). MicroRNAs are a class of small noncoding ribonucleic acids involved in regulating gene expression (Lamadrid-Romero et al. 2018; Lim et al. 2005). Changes in host miRNAs following T. gondii infection are mainly related to immunity and apoptosis processes (Xu et al. 2013). The miR-132 regulates neuronal and immune functions and is involved in the differentiation of dopamine neurons (Yang et al. 2012; Wanet et al. 2012). Also, miR-132 upregulation modulates a set of important pathways that have a critical role in vital brain functions, translational initiation and regulation, meiosis, mitosis, carcinogenesis, and cell growth, differentiation, survival, and proliferation (Xiao et al. 2014). The dysregulation of microRNA-132 expression can be associated with various behavioral and neuropsychiatric disease models, such as SZ, AD, and PD, in humans and animals (Hinze-Selch 2015; Miller et al. 2012; Wanet et al. 2012). The microRNA-132 expression was down-regulated during chronic infection (Li et al. 2015; Remenyi et al. 2013; Yang et al. 2012); nevertheless, acute T. gondii infection had an opposite effect (Li et al. 2015). The parasite probably needs these changes at the stage chronic in its life cycle to have complete control over the host cell (Abo-Al-Ela 2019). In addition to these mechanisms, a substance may play a role in the development of mental disorders; one study investigated the effects of exposure to T. gondii alone and methylmercury (MeHg), along with T. gondii in vivo. The obtained results showed that MeHg and T. gondii had synergistic effects and significantly increased the number of cysts in the brains of mice (King et al. 2003). Thimerosal, an organic-mercury compound used as a preservative in vaccines and serum to block the growth of contaminants in biological materials (Fernandes Azevedo et al. 2012), inhibits dopamine-stimulated methylation reactions in the body. These reactions are important for growth factors and promotion of normal neuronal growth (Waly et al. 2004).
Serotonin
Serotonin is a neurotransmitter involved in the pathogenesis of some psychiatric disorders, such as depression, which vary with T. gondii infection (Fig. 3) (Mahmoud et al. 2016; Oxenkrug 2013; Tyebji et al. 2019). Moreover, serotonin (5-hydroxytryptamine, 5-HT) is a neurotransmitter released from serotonergic neurons. The neurons that produce serotonin are located in the midbrain, hypothalamus, substantia nigra, pineal gland, and raphe nuclei of the brain stem (Herlenius and Lagercrantz 2004). In humans, serotonergic neurons appear in 5–12 weeks of pregnancy (Herlenius and Lagercrantz 2004). Serotonergic cells send axons to the anterior part of the brain, and these cells could play an important role in the differentiation of neural precursors (Gaspar et al. 2003). Serotonin receptors are divided into 7 families (i.e., 5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6, and 5-HT7) and 14 subtypes (i.e., 5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E, 5-HT1F, 5-HT2A, 5-HT2B, 5-HT2C, 5-HT3, 5-HT4, 5-HT5A, 5-HT5B, 5-HT6, and 5-HT7). Most of the 5-HT receptors belong to the G-protein receptor family, and only 5-HT3 receptors are ligand-gated ion channels (Abg Abd Wahab et al. 2019). Serotonin plays an important role in innate and acquired immune responses and affects the secretion of cytokines by stimulating lymphocytes and monocytes (Durk et al. 2005). This neurotransmitter acts as a significant mediator in inflammatory processes, as well as endocrine and autonomic system (Ohno 2011). Serotonin modulates the inflammation of the nervous tissue, cell division, neuronal migration, cell differentiation, synaptogenesis, and motor behavior (Abg Abd Wahab et al. 2019; Gaspar et al. 2003). It also plays an important role in the regulation of important physiological processes, such as motor activity, body temperature, pain, sleep, cognitive function, and emotional behaviors (Ohno 2011). The tachyzoite phase of T. gondii requires tryptophan to replication (Fabiani et al. 2013). Tryptophan is catabolized by two pathways, including one through the kynurenine pathway and the other through tryptophan 5-hydroxylase decarboxylation, to form serotonin (Hsu et al. 2014). The biosynthesis of serotonin may be directly affected by two genes, namely AAH1 and AAH2, in the genome of T. gondii (Fuglewicz et al. 2017). Furthermore, serotonin may be involved in innate fear behavior (e.g., rodent avoidance of cat odors), and T. gondii-stimulated IFN-γ can potentially affect the attraction and loss of fear for predator odor in rodents (Graeff et al. 1997). These behavioral changes associated with chronic T. gondii infection may have an effect on neural and glial cysts, disruption of neurotransmitters, such as serotonin, and local inflammatory responses in the brain (Hurley and Taber 2012). The impaired metabolism of neurotransmitters, transporters, and their receptors has been associated with various types of pathological manifestations (e.g., the role of serotonin in depression) (Hamon and Blier 2013; Parlog et al. 2015). In addition, different neurotransmitters interact with each other in regulating the excitation and inhibition of motor neurons (Wagner et al. 2017). For example, serotonin plays a role in the reduction or enhancement of glutamate-mediated transmission and GABA-mediated transmission in motor structures (Ciranna 2006). Factors that alter serotonin levels are discussed in the following sections.

Structure of serotonin
Role of indoleamine 2, 3-dioxygenase
The infection with T. gondii induces a strong type 1 T helper (Th1) immune response in which IFN-γ, IL-12, and killer T cells play a major role (Munoz et al. 2011).The INF-γ and some cytokines, such as IL-2, induce the release of IDO, catabolizing tryptophan (Muller and Schwarz 2006; Tan et al. 2012). Tryptophan is an essential amino acid and a precursor to serotonin biosynthesis; therefore, when the concentration of IDO increases during T. gondii infection, it may decrease the synthesis of this neurotransmitter (Schwarcz et al. 2012). Tryptophan metabolism induces the kynurenine pathway in which the tryptophan is catabolized and its products are the two neuroactive metabolites, namely QA and KA (Schwarcz and Pellicciari 2002). Increased KA concentration is a characteristic of SZ (Sophie Erhardt et al. 2001a, b); however, QA upregulation has been implicated in the etiology of many mental disorders, including AD (Gulaj et al. 2010), Huntington’s disease (AL-Hadad et al. 2019; Guidetti et al. 2006), depression (Steiner et al. 2011), and autism spectrum disorder (Zimmerman et al. 2005), as well as in suicide attempts (Erhardt et al. 2013). The activation of the kynurenine pathway was reported in the brain of T. gondii-infected mice (Murakami et al. 2012).
Role of monoamine oxidase A
Serotonin is one of the biogenic amines that MAOA, a mitochondrial membrane enzyme, plays an important role in its metabolism (Courtet et al. 2005). It was reported that the infection with T. gondii reduced the in vivo expression of the MAOA gene (Sugden et al. 2016). Increasing the activity of this enzyme reduces the levels of serotonin (Courtet et al. 2005). The functional polymorphism of the MAOA gene has been connected with anxiety, depression, autism, alcoholism, aggression, suicidality, and impulsiveness (Caspi et al. 2002; Caspi et al. 2003; Lau et al. 2009; Lesch et al. 1996; Nilsson et al. 2006; Nilsson et al. 2008; Sjoberg et al. 2006).
Activation of mitogen-activated protein kinases
During toxoplasmic encephalitis, several cytokines and inflammatory mediators are produced exerting their effect on neurotransmitters through the activation of the MAPK pathway (Haroon et al. 2012). This pathway controls various cellular processes, such as growth, differentiation, and stress responses. The stimulation of this pathway can enhance the function of the serotonin transporter and reduce serotonin at the level of the synapse (Zhu et al. 2005, 2010).
Glutamate
Glutamate is a nonessential amino acid whose metabolism and signaling are altered by T. gondii infection (Fig. 4) (Lang et al. 2018; Patri 2019). Since the 1950s, the excitatory action of glutamate has been described in the mammalian brain and spinal cord (Curtis and Watkins 1960; Hayashi 1952). In the late 1970s, glutamate was identified as the main excitatory transmitter in the vertebrate nervous system (Meldrum 2000). This neurotransmitter is synthesized from glucose and other precursors and cannot cross the blood–brain barrier (Patri 2019). The overproduction of glutamate occurs in the early postnatal period (e.g., after 1–2 weeks in the rat cortex and following 1–2 years in the human cortex) (Benítez-Diaz et al. 2003; Herlenius and Lagercrantz 2004). Given that glutamate is involved in different metabolic pathways, it is critical for maintaining normal CNS function (Abg Abd Wahab et al. 2019; Bak et al. 2006). Five types of glutamate transporters are known, including excitatory amino acid transporter 1, excitatory amino acid transporter 2, excitatory amino acid transporter 3, excitatory amino acid transporter 4, and excitatory amino acid transporter 5 (Y. Zhang et al. 2016). Glutamate receptors are located on neuronal and nonneuronal cells and involved in the regulation of many cognitive processes (Daikhin and Yudkoff 2000; Dauvermann et al. 2017). Alterations in the function or expression of the glutamate transporters (i.e., excitatory amino acid transporters [EAATs]) are shown in the pathogenesis of many psychiatric disorders, such as AD and PD (Benarroch 2010). The EAATs are involved in the prevention of glutamate dysregulation by controlling its release and reuptake. These transports can also play a role in neuronal development, memory, excitotoxicity, learning, pain perception, and motor behavior regulation (Patri 2019; Zhang et al. 2016). In addition, glutamate interacts with other neurotransmitters. In this regard, serotonin plays a role in glutamate-mediated transmission in the control structures of movement (Ciranna 2006; Wagner et al. 2017). The chronic infection of T. gondii results in a two-fold reduction in the expression of the glutamate transporter 1 (GLT-1) in glial cells and increase extracellular glutamate (David et al. 2016).

Structure of glutamate
Activation of N-methyl-D-aspartate receptor
The activity of IDO is mostly stimulated by IFN-γ, and the release of this enzyme results in the destruction of tryptophan (Dalimi and Abdoli 2012; Tedford and McConkey 2017). Tryptophan is an essential amino acid required for the growth of T. gondii; therefore, the growth of parasite is inhibited by decreasing the concentration of this amino acid (Cerávolo et al. 1999). Kynurenine is mainly metabolized to KA; however, the major product of kynurenine metabolism is QA in inflammatory conditions (Hilmas et al. 2001; Saito et al. 1992). The QA, an NMDA agonist produced by microglia, binds to glutamate N-methyl-D-aspartate receptors (NMDARs) (Guillemin et al. 2005; Schwarcz and Pellicciari 2002). The NMDAR plays an important role in the pathophysiology of several mental disorders, such as SZ (Javitt 2014; Moghaddam and Javitt 2012; Papouin et al. 2017), and it has been suggested as the mechanistic link between T. gondii infection and onset of SZ (Lucchese 2017; Tao Wang et al. 2013). Kannan et al. showed that exposure to T. gondii leads to the production of serum IgG antibodies to NMDARs. This change may provide the condition for a reduction in the function of glutamate receptors, and related disorders have been observed in SZ (Geetha Kannan et al. 2016; Kannan et al. 2017). Furthermore, the evaluation of T. gondii seronegative SZ patients and seropositive non-SZ controls showed lower NMDA IgG titers, compared to those reported for seropositive individuals with SZ (G Kannan et al. 2017). The activation of NMDAR stimulates excitotoxicity and oxidative stress in the brain (Elsheikha and Zhu 2016). On the other hand, KA, an antagonist of NMDAR produced in astrocytes, reduces glutamatergic neurotransmission and can lead to cognitive deficits (Goff 2015; Guillemin et al. 2005; Kegel et al. 2014; Schwarcz and Pellicciari 2002). In addition, KA has been implicated in the inhibition of alpha 7 nicotinic acetylcholine (α7nACh) receptors and modulation of glutamatergic neurotransmission (Beggiato et al. 2013). Glutamate converts to glutamine by glutamine synthetase, and changes in glutamine increase calcium entry into neurons which may play a role in excitotoxicity and altered neurotransmission (Elsheikha and Zhu 2016; Shen 2013). Some of the mechanisms affecting glutamate levels are discussed in the following sections.
Interaction of glutamate and gamma-aminobutyric acid
The GABA is an inhibitory neurotransmitter that may play a key role during T. gondii infection. This neurotransmitter is able to regulate glutamate signaling. The glutamic acid decarboxylase 67 (GAD67) converts glutamate to GABA. In infected animals, general changes are observed in the localization of GAD67; nevertheless, the GABAergic pathway remains intact (Brooks et al. 2015). In mice infected with T. gondii, the number of tissue cysts was low; however, the location of GAD67 was impaired throughout the brain. Finally, impaired GABAergic signaling and/or increased glutamate can cause neurophysiological outcomes, such as seizures, associated with T. gondii infection (Treiman 2001).
Decreased expression of glutamate transporter 1
A significant reduction in GLT-1 and a significant increase in CNS extracellular levels of glutamate were observed during T. gondii infection (David et al. 2016). In pathological conditions and dysregulation of the glutamate transporter expression and/or function, excess extracellular glutamate causes the death of excitotoxic cells in surrounding postsynaptic neurons by the overactivation of ionotropic glutamate receptors (Choi 1994). Several studies have shown the antitoxoplasmic effect of antipsychotic drugs on the inhibition of in vitro parasite proliferation; nonetheless, other studies have not demonstrated an in vivo effect. Ceftriaxone is a beta-lactam antibiotic that increases GLT-1 expression (Rothstein et al. 2005). Ceftriaxone reduced infection-induced extracellular glutamate concentrations and was neuroprotective in mice. However, it had no effect on the immune response to the infection and behavioral deficits in mice (David et al. 2016).
Gamma-aminobutyric acid
The GABA is another neurotransmitter that appears to play a role in the mechanisms of anxiety and depression in the mammalian CNS (Fig. 5) (Kalueff and Nutt 2007), and its levels vary with T. gondii infection (Lang et al. 2018). The GABA, the most ubiquitous inhibitory neurotransmitter in the CNS, was identified in 1883. The main role of GABA is to modulate the flow and timing of excitatory neurotransmitters, such as glutamate, thereby preventing seizures (Bazemore and Florey 1956; Treiman 2001). Its inhibitory role was firstly introduced in the late 1950s by Bazemore and Florey (Bazemore and Florey 1956). The GABA is considered an important transmitter for the human infant, and the elimination of vitamin B6 from the human infant formula can lead to deadly consequences. The GABA deficiency is the major cause of these consequences and results in fatal seizures (Frimpter et al. 1969). The infection with T. gondii causes GABA release from the DCs (Fuks et al. 2012), and this protozoan uses this neurotransmitter as a carbon source (MacRae et al. 2012). The GABA is not a product of intermediary metabolism and is not used in protein synthesis. Moreover, it is produced in the neuronal tissue of glutamate by alpha decarboxylation of glutamic acid in the presence of GAD (Patri 2019). Probably 25–40% of nerve terminals contain GABA (Marín and Rubenstein 2001). About 95% of GABAergic cells are located in the striatum. The GABA activity can also be observed elsewhere, such as the cerebral cortex, lateral vestibular nucleus, and spinal cord (Patri 2019). There are two types of rapid and slow inhibitory receptors (i.e., ionotropic GABAA and metabotropic GABAB, respectively) (Herlenius and Lagercrantz 2004; Patri 2019). Ionotropic receptors increase Cl− ion conductance; however, metabotropic receptors indirectly change membrane ion permeability and neuronal excitability (Stein and Nicoll 2003). Given that different neurotransmitters interact, any disturbance in glutamate levels and signaling during T. gondii infection can exacerbate changes in GABA signaling in the brain (Ngô et al. 2017; Ngoungou et al. 2015; Palmer 2007; Tyebji et al. 2019). The neurophysiological outcomes are the result of the disruption of GABAergic signaling and elevated glutamate levels (Treiman 2001). The GABA activates GABAA receptors on Toxoplasma-infected DCs leading to a hypermigratory phenotype in those cells. This process was assumed to be a tool for parasite propagation (Brooks et al. 2015). Moreover, in order to regulate motor neurons, various neurotransmitters, such as serotonin and GABA, interact (Wagner et al. 2017). For example, GABA-mediated translocation occurs in structures controlling the movement affected by serotonin (Ciranna 2006). The following factors alter GABA levels.

Structure of GABA
Role of interferon-gamma
The IFN-γ production is the first line of defense against T. gondii infection. The IFN-γ is produced by NK and T cells (Hunter et al. 1996). The chemokines, such as chemokine (C–C motif) ligand 5 (CCL5), chemokine (C-X-C motif) ligand 9 (CXCL9), and chemokine (C-X-C motif) ligand 10 (CXCL10), are expressed by astrocytes and microglia during T. gondii infection in the brain. These compounds attract the produced IFN-γ to T. gondii antigens and lead to parasite release (Yasuhiro Suzuki et al. 2011). Macrophages are among the IFN-γ-activated cells that express C–C chemokine receptor type 5 in these cells. Activated macrophage prevents the proliferation of T. gondii tachyzoites (Denkers and Gazzinelli 1998). The combined expression of the three chemokines (i.e., CCL5, CXCL9, and CXCL10) plays an important role in maintaining the Th1 response and facilitating macrophage migration to the brain in order to control the parasite (Imtiaz A Khan et al. 2000). The IFN-γ plays an important role in the regulation of the behaviors, such as anxiety, depression, as well as cognitive memory, and T. gondii infection can disrupt these behaviors (Gonzalez et al. 2007; Mahmoud et al. 2017; Mahmoud et al. 2016). Moreover, IFN-γ increases tonic GABAergic inhibitory current and latency of seizure onset and reduces the severity of the seizure. A study reported that IFN-γ is involved in the regulation of GABAergic signaling and social behaviors in mice. Social deficits in IFN-γ knockout mice are rescued by the use of diazepam, a GABA activator (Tyebji et al. 2019). Diazepam increases the affinity of GABA to the GABAA receptor (Lavoie and Twyman 1996). The infection with T. gondii tissue cysts leads to changes in the distribution of GABA in the brain, which could disrupt the social activities that IFN-γ regulates in infected mice (Brooks et al. 2015; Tyebji et al. 2019). In IFN-γ knockout mice, disorders, such as depression and anxiety, increase and neurogenesis decreases in the hippocampus (Campos et al. 2014). Under the basal condition, IFN-γ knockout mice displayed impaired spatial recognition memory; however, in chronic stressful conditions, such as T. gondii infection, IFN-γ knockout mice exhibited enhanced memory function. This finding suggests that IFN-γ signaling during infection may worsen memory impairment (Litteljohn et al. 2014; Tyebji et al. 2019).
Dysregulation of glutamic acid decarboxylase 67
The major part of GABA (90%) in the CNS is produced by GAD67 as a structurally active enzyme; nevertheless, GAD65 is temporarily activated. The GAD65 plays a role in the rapid modulation of inhibitory neurotransmission (Asada et al. 1997; Condie et al. 1997). The GAD67 is the main enzyme for the neuronal biosynthesis of GABA and converts glutamate to GABA. The GABA is involved in the regulation of glutamate signaling (Brooks et al. 2015). The effect of T. gondii infection on GAD65 levels has not been studied (Tyebji et al. 2019); nevertheless, the infection with T. gondii causes spontaneous seizures and in vivo redistribution of GAD67 (Brooks et al. 2015). The cause of the seizures in mice with toxoplasmic encephalitis is certain changes in synaptic localization of GAD67 and impairment of GABAergic cells that can lead to extracellular accumulation of glutamate. One of the symptoms of MDD is GABAergic dysfunction in the prefrontal cortex (Croarkin et al. 2011; Karolewicz et al. 2010; Levinson et al. 2010; Tedford and McConkey 2017). This phenotype was also observed in the animal models of T. gondii infection (Mahmoud et al. 2017, 2016). T. gondii is directly involved in inhibitory neurotransmission and provides a molecular basis for the occurrence of various neurological consequences in patients with toxoplasmic encephalitis (Brooks et al. 2015).
Norepinephrine (Noradrenaline)
Norepinephrine is another neurotransmitter the infection of which with T. gondii affects its levels (Fig. 6) (E Fuller Torrey and Yolken 2003). Alteration in the levels of this neurotransmitter has been observed in various mental disorders, such as SZ, completed suicide, and suicide attempts (Arango et al. 1996; Ordway et al. 1994; E Fuller Torrey and Yolken 2003). The appearance of noradrenergic neurons occurs in the early stages of pregnancy (i.e., 12–14 days in mice and 5–6 weeks in humans) (Sundström et al. 1993). Norepinephrine is primarily produced in the brainstem neurons of the locus coeruleus and subcoeruleus, as well as several other nuclei (Xu et al. 2000). The effects of noradrenaline are mediated by adrenergic receptors. These receptors are divided into three main types (i.e., alpha-1, alpha-2, and beta), and each type of receptors is classified into three subtypes. These receptors are widely expressed throughout the CNS and in almost all peripheral tissues (Ahlquist 1948; Bylund et al. 1994). Among the adrenergic sites, alpha-2 adrenergic receptors are abundant, and these receptors spread throughout all the regions of the transient embryonic cortical (F. Wang and Lidow 1997). Norepinephrine is involved in regulating a number of physiological processes, including mood, sleep, arousal, aggression, memory, alertness, blood flow, and metabolism, as well as the endocrine and autonomic nervous systems (Axelrod and Kopin 1969; Feldman et al. 1997; Wang et al. 1999). The reduction in the concentration of this neurotransmitter has been reported in a variety of motor disorders in humans that may explain the possible relationship between toxoplasmosis and motor disorders (Nath et al. 1993; Rommelfanger et al. 2007). Ihara et al. in 2016 reported a reduction in norepinephrine levels in the cortex and amygdala in mice infected with T. gondii (Ihara et al. 2016). The results of another study showed that norepinephrine levels in mice infected with acute infection decreased by 28% (Stibbs 1985). In addition, latent toxoplasmosis can reduce the aversion of rodents to cat odors (Berdoy et al. 2000). These behavioral changes associated with latent T. gondii infection may impair the norepinephrine neurotransmitter (Hurley and Taber 2012), and the cortical noradrenergic system may play a role in fear avoidance (Andrews et al. 1993). The data on the changes in the levels of neurotransmitters in rats with high avoidance of cat odor demonstrated an increased cortical noradrenaline level after the exposure to cat odor (Andrews et al. 1993). There are several mechanisms involved in rodent aversion to cats. For example, the cat odor activates the wolframin gene in the amygdaloid region. Activation of this gene occurs in parallel with the activation of carboxypeptidase E, suggesting a possible role for wolframine in neuropeptide synthesis (Kõks et al. 2002). Mutations of the wolfram syndrome 1 (WFS1) gene are responsible for wolfram syndrome that is characterized by further neurological and endocrinological abnormalities. The WFS1 gene encodes wolframin. The function of the wolframin protein, a multispanning membrane glycoprotein of the endoplasmic reticulum, is not yet fully perceived (Hofmann et al. 2003). Despite the fact that its function is unknown, there is some evidence that wolframin mutations may predispose people to affective disorders. Swift et al. reported that 60% of wolfram syndrome patients showed psychiatric symptoms (Swift et al. 1998). Moreover, there is a node of the extra-hypothalamic vasopressinergic system in the poster-dorsal medial amygdala (MePD) of rats that contains many arginine vasopressin (AVP) neurons. T. gondii causes hypomethylation of the AVP promoter, resulting in increased expression of this peptide in the MePD of infected male rats. Infected animals exposed to cat odor show activation of vasopressinergic neurons and the accompanying loss can be reversed by the systemic hypermethylation of the MePD-AVP (Hari and Vyas 2014). These behavioral changes may be due to altered neurochemical pathways, including norepinephrine, as well as the presence of cysts in the brain (Klein 2003). There have been also several studies with contradictory results. For example, a systematic review examining the role of hormones in T. gondii infection reported a lack of association between noradrenaline and T. gondii (Galvan-Ramirez Mde et al. 2014). The following sections discuss several mechanisms involved in norepinephrine levels.

Structure of norepinephrine
Activation of guanosine triphosphate cyclohydrolase 1
The stimulation of IFN-γ and other proinflammatory cytokines results in GTP-CH1 activity. The activity of this enzyme leads to a decrease in phenylalanine and tyrosine. Since phenylalanine is a precursor of norepinephrine, lowering its levels results in the reduced synthesis of this neurotransmitter (Capuron et al. 2011; Sperner-Unterweger et al. 2014). In addition, since several cells metabolize dopamine to norepinephrine, the reduction of phenylalanine and tyrosine amino acids as the precursors of dopamine also affects the levels of norepinephrine produced (Capuron et al. 2011; Prandovszky et al. 2011).
Decreased expression of monoamine oxidase A
The MAOA and monoamine oxidase B (MAOB) play an important role in the metabolism of neurotransmitters and dietary amines (Grimsby et al. 1991). The MAOA preferentially oxidizes biogenic amines, such as norepinephrine, and is inactivated by acetylenic inhibitor clorgyline (Grimsby et al. 1991). The reduction of MAOA expression was demonstrated in mice with T. gondii infection. Five days after the infection, a 55% reduction in the expression of this enzyme was reported in the striatum of mice brain, compared to that reported for the controls (Xiao et al. 2014). The abnormal levels of monoamine oxidase (MAO) activity have been reported in some psychotic disorders, such as SZ (Wyatt and Murphy 1976), depression (White et al. 1980), and alcoholism (Sullivan et al. 1979). In addition, MAO inhibitors have been used as antidepressants (Murphy et al. 1984).
Epinephrine (Adrenaline)
Epinephrine is another neurotransmitter whose levels may be affected by T. gondii infection (Fig. 7). It has been shown that depression and suicidal behaviors are often associated with an imbalanced immune response and metabolism of neurotransmitters, such as epinephrine (Capuron and Miller 2011; Haroon et al. 2012; Sperner-Unterweger et al. 2014). Epinephrine was the first hormone to be isolated into the pure state in 1900 by Jokichi Takamine (Scanzano and Cosentino 2015). Three α1 (i.e., α1a, α1b, and α1d), three α2 (α2A, α2B, and α2C), and three β (i.e., β1, β2, and β3) are major types of adrenergic receptors. Furthermore, these receptors are among the best described G protein-coupled receptors (Cotecchia et al. 2012). Multiple studies have reported a relationship between these behavioral changes and alterations in the concentration of neurotransmitters (Jaroslav Flegr and Kuba 2016; Skallova et al. 2006). According to the evidence, it was shown that urinary epinephrine excretion during a cognitive challenge in ADHD subjects was at least 40% lower than that reported for the control group (Pliszka et al. 1994). Another study investigated the effect of congenital T. gondii infection on the behavior and concentration of neurotransmitters. Only the impact of congenitally T. gondii infection on the short-term memory behavior of male rats showed a significant difference, compared to that reported for the control group. The results of the aforementioned study did not confirm the changes in neurotransmitters caused by chronic T. gondii infection (Goodwin et al. 2012). Another study examined the effects of chronic toxoplasmosis on the levels of some neurotransmitters, such as dopamine, adrenaline, and noradrenaline, in human serum. The findings of the aforementioned study showed that the serum levels of adrenaline were significantly higher in patients with latent toxoplasmosis, compared to those reported for the control group (AL-Hadad et al. 2019). In addition, a systematic review by Galván-Ramírez in 2014 investigated the role of hormones in T. gondii infection. Based on the results, there was no association between adrenaline and T. gondii (Galvan-Ramirez Mde et al. 2014). MAOA with the following mechanism is effective in altering epinephrine levels.

Structure of epinephrine
Function of monoamine oxidase A
Epinephrine is one of the neurotransmitters catalyzed by the MAOA and MAOB (Grimsby et al. 1991). The changes in epinephrine levels have been observed in several mental disorders, such as AD (Tipton et al. 1984). The MAOA and MAOB are present in almost all human tissues except for the placenta that expresses most of the MAOA (Egashira 1976; Grimsby et al. 1990; Hsu et al. 1988), as well as platelets and lymphocytes that mainly show MAOB. In the fetal brain, MAOA is expressed before MAOB (Grimsby et al. 1990; Lewinsohn et al. 1980). Since T. gondii infection reduces the expression of the MAOA (87), it will probably affect epinephrine levels.
Acetylcholine
Acetylcholine is one of the most important neurotransmitters in the brain (Fig. 8) (Herlenius and Lagercrantz 2004). Multiple studies have indicated that the infection with T. gondii can interfere with several neurotransmitter systems, such as acetylcholine, leading to classical psychiatric illness (Anisman et al. 2008; Dunn and Wang 1995; Miller et al. 2009a, b). This neurotransmitter is important for the processes, such as cortical activation, pain, as well as memory, and counterbalances the effect of dopamine (Cooper et al. 2003; Johnston and Silverstein 2003). The cholinergic neurons in the brain are organized in subcortical regions, and the axons of these neurons spread in many areas of the brain, such as the cortex and hippocampus (Houser et al. 1985). These nerves play an important role in cortical growth, flexibility, and function (Hohmann 2003). The time of cholinergic nerve formation in the human fetus is about the 20th week and later than that reported for monoaminergic (Berger-Sweeney and Hohmann 1997). Cholinergic receptors are divided into nicotinic and muscarinic receptors and stimulated by nicotine or muscarinic drugs (Tiwari et al. 2013). Nicotinic acetylcholine receptors may be associated with development and flexibility. Moreover, the activation of these receptors creates synaptic contacts and wiring in the postnatal development period (Maggi et al. 2003). Chronic T. gondii infection affects cholinesterase activity and increases the level of acetylcholinesterase in the brain of infected mice. Acetylcholinesterase is a membrane-bound enzyme that hydrolyzes acetylcholine (Tonin et al. 2014; Alexandre Alberto Tonin et al. 2013). Therefore, various methods are used to improve the learning and memory impairments caused by T. gondii infection and reduce acetylcholine levels, including increasing the level of acetylcholine through synthesis promoters or using cholinesterase activity inhibitors (Giacobini 1996). One of these cholinesterase inhibitors is donepezil hydrochloride, which is used for the treatment of AD (Mahmoudvand et al. 2016). According to an animal study, it was observed that the treatment with this drug, in addition to improving memory, reduced β-amyloid in the brain of mice (Easton et al. 2013). Increased production of KA with the following mechanism affects acetylcholine levels.

Structure of acetylcholine
Increased production of kynurenic acid
The infection with T. gondii in the CNS causes the strong activation of glial cells, especially astrocytes, which increase the production of KA in the brain (Müller and Schwarz 2007). The conversion of tryptophan to KA is due to tryptophan dioxygenase and IDO activity in the brain (Dalimi and Abdoli 2012). The KA is an endogenous antagonist of the α7nACh receptor. This receptor is widely implicated in memory, learning, and cognitive processes (Schwarcz and Hunter 2007; Wonodi and Schwarcz 2010). The alteration of the concentration of endogenous KA in the brain can inhibit the α7nAch receptors for acetylcholine (Müller and Schwarz 2007). The high levels of KA may lead to cognitive impairments (Schwarcz and Hunter 2007; Wonodi and Schwarcz 2010); for example, the high levels of KA were observed in the cerebrospinal fluid of schizophrenic patients (Erhardt et al. 2001a, b; Nilsson et al. 2005; Schwarcz et al. 2001).
Nitric oxide (NO)
The NO has been identified as the first gasotransmitter. Gasotransmitters are vasoactive gaseous messenger molecules involved in different signaling processes (Fig. 9) (Mustafa et al. 2009). The NO is described as an endothelium-derived relaxing factor and produced in different types of mammalian cells. The production of NO by the NO synthase enzyme is carried out using L-arginine (Erdmann et al. 2013). This enzyme has three isoforms, including endothelial nitric oxide synthase (eNOS), neuronal nitric oxide synthase (nNOS), and inducible nitric oxide synthase (iNOS). The aforementioned isoforms are expressed in vascular endothelium and choroid plexus, neuronal cell bodies, as well as macrophages and glial cells, respectively. These three isoforms are produced in response to in vivo T. gondii infection (Dincel and Atmaca 2015; Tonin et al. 2014). An increase in intracellular Ca2+ leads to the activation of eNOS or nNOS, which results in NO production; however, iNOS-mediated NO production is independent of Ca2+ (Steinert et al. 2010). In physiological conditions, iNOS levels in the brain and arteries are low (Ortiz and Garvin 2003); nevertheless, in inflammatory conditions, such as T. gondii infection, iNOS levels are upregulated (Fujigaki et al. 2003). The NO, as a neurotransmitter in the brain, is involved in the processes of regulating memory and learning (Paul and Ekambaram 2011; Steinert et al. 2010), as well as in brain homeostasis, by cerebral blood flow during rest and under physiological conditions (Garry et al. 2015). T. gondii infection increases the production of NO (Butcher et al. 2001; Suzuki 2002) which is involved in killing and eliminating the parasite (I. A. Khan et al. 1997). Although NO production inhibits intracellular parasite replication, it has harmful and sometimes fatal effects on cells and infected mice (Sa et al. 2018). The NO production due to inflammation and resulting oxidative stress has been implicated in the etiology of psychiatric disorders, such as AD, PD, HD, as well as autoimmune diseases, such as multiple sclerosis (Bennett et al. 2009; Steinert et al. 2010). The NO and inflammatory cytokines, such as IL-2 and IL-6, are innate defenses against T. gondii infection (C. M. Miller et al. 2009a, b; Yarovinsky 2014) which in addition to their role in the etiology of psychiatric disorders contributes to the increasing levels of dopamine in the brain (Alonso et al. 1993; Petitto et al. 1997; Prast and Philippu 2001; West et al. 2002; Zalcman et al. 1994). IFN-γ alters the NO level with the following mechanisms.

Structure of nitric oxide
Role of interferon-gamma
Neuroinflammatory responses are observed in chronic T. gondii infection. These responses are not confined to the areas surrounding the cyst and exert their effects throughout the CNS. T. gondii infection induces Th1 immune response (Dupont et al. 2012) and leads to the release of several cytokines, such as IFN-γ, IL-12, IL-1, IL-6, and TNF (Biswas et al. 2015, 2017; Carruthers and Suzuki 2007; Mohle et al. 2014; Wohlfert et al. 2017). The IFN-γ stimulates transforming growth factor beta-1 which, in turn, prevents the production of NO (Rozenfeld et al. 2005). As a result, the neuronal degeneration in the infected brain with T. gondii is limited (Mohle et al. 2016; Parlog et al. 2014). Therefore, this protective NO mechanism is used for the development of a vaccine against parasite in order to reduce parasitism in the brain (Czarnewski et al. 2017). The brain cyst burden is high in iNOS knockout mice (Sa et al. 2018); however, brain inflammation is lower than their parental lines, and there is no change in the levels of IFN-γ or TNF-α (I. A. Khan et al. 1997). Furthermore, the protective role of NO against T. gondii infection is tissue-specific (Scharton-Kersten et al. 1997). Despite the increase in T. gondii replication in the brain of iNOS knockout mice, the survival time of these mice increased after the infection (I. A. Khan et al. 1997). In addition, IFN-γ knockout mice had higher iNOS levels in the lungs but with lower levels of this enzyme in the brain than those reported for wild-type mice (Fujigaki et al. 2002). The IFN-γ knockout mice are not able to inhibit the growth of parasites (Jun et al. 1993).
Conclusion and future perspectives
The pathology of mental illnesses is a result of brain dysfunction. Since the brain is the most important site for the formation of T. gondii cysts, there might be various changes during the infection resulting in various mental illnesses of the brain. There is a hypothesis about the role of T. gondii infection in the pathophysiology of mental illnesses is to make changes in different neurotransmitters. There has been limited information on chronic T. gondii infection of the brain and its effects on behavior associated with changes in neurotransmitters. It is recommended to perform further studies in this field focusing on the observed alterations in neurotransmitters and impact of these changes on mental disorders. Furthermore, further studies are required to identify the effects of T. gondii infection on neurotransmitters and interactions of the immune system and neurotransmitters. Moreover, the alteration in the levels or functions of neurotransmitters can be due to changes in neurotransmitter receptors and receptor sensitivity that may be investigated in future studies. There has been a high prevalence of latent toxoplasmosis in different countries, increasing prevalence of psychiatric disorders, increased probability of traffic accidents and working accidents, suicides, as well as other side effects of the infection associated with neurotransmitter changes. Therefore, it is required to perform further studies in countries with a higher prevalence of toxoplasmosis to elucidate the role of toxoplasmosis in neurotransmitters changes (especially less studied neurotransmitters, such as glycine, aspartate, and histamine) in psychiatric disorders. The perception of the link between T. gondii and neuropsychiatric diseases is important because it may provide an incentive to develop drugs and vaccines for the treatment or prevention from this infection, thereby eliminating one of the environmental factors that can cause problems in brain function. Future studies provide new perspectives on the prevention and treatment of these neurodegenerative disorders if they confirm the impact of T. gondii infection on neurotransmitters changes and their association with psychiatric disorders.
Authors’contributions
AD and TN review and wrote the manuscript; SS performed the literature research; AD supervised the review. All authors have read and approved the final version of the manuscript, and agree with the order of presentation of the authors.
Data availability statement
No data was used for the research described in the article.
Abbreviations
- CNS:
-
Central nervous system
- DALYs:
-
Disability-adjusted life years
- HIV:
-
Human immunodeficiency virus
- T. gondii :
-
Toxoplasma gondii
- IgG:
-
Immunoglobulin G
- MDD:
-
Major depression disorder
- SZ:
-
Schizophrenia
- BD:
-
Bipolar disorder
- OCD:
-
Obsessive–compulsive disorder
- AD:
-
Alzheimer's disease
- PD:
-
Parkinson’s disease
- ADHD:
-
Attention-deficit hyperactivity disorder
- TH:
-
Tyrosine hydroxylase
- D2R:
-
Dopamine receptor D2
- TNF:
-
Tumor necrosis factor
- NF-κB:
-
Nuclear factor kappa light-chain enhancer of activated B cells
- IL-1β:
-
Interleukin 1 beta
- GABA:
-
Gamma-aminobutyric acid
- NO:
-
Nitric oxide
- DR:
-
Dopamine receptors
- AAH:
-
Aromatic acid hydroxylase genes
- IFN-γ:
-
Interferon-gamma
- NK:
-
Natural killer
- GTP-CH1:
-
Guanosine triphosphate cyclohydrolase 1
- BH4:
-
Tetrahydrobiopterin
- DCs:
-
Dendritic cells
- TLR:
-
Toll-like receptor
- IDO:
-
Indoleamine 2,3-dioxygenase
- KA:
-
Kynurenic acid
- QA:
-
Quinolinic acid
- NMDA:
-
N-methyl-D-aspartate
- MAPK:
-
Mitogen-activated protein kinases
- MAOA:
-
Monoamine oxidase A
- MeHg:
-
Methylmercury
- Th:
-
T helper
- EAATs:
-
Excitatory amino acid transporters
- GAD67:
-
Glutamic acid decarboxylase 67
- CCL5:
-
CC chemokine ligand 5
- CXCL9:
-
CXC chemokine ligand 9
- CXCL10:
-
CXC chemokine ligand 10
- WFS1:
-
Wolfram syndrome 1
- MePD:
-
Poster-dorsal medial amygdala
- AVP:
-
Arginine vasopressin
- eNOS:
-
endothelial nitric oxide synthase
- nNOS:
-
Neuronal nitric oxide synthase
- iNOS:
-
Inducible nitric oxide synthase
References
Abdoli A, Dalimi A, Arbabi M, Ghaffarifar F (2014) Neuropsychiatric manifestations of latent toxoplasmosis on mothers and their offspring. J Matern-Fetal Neonatal Med 27(13):1368–1374. https://doi.org/10.3109/14767058.2013.858685
Abedalthagafi M, Rushing EJ, Garvin D, Cheson B, Ozdemirli M (2009) Asymptomatic diffuse “encephalitic” cerebral toxoplasmosis in a patient with chronic lymphocytic leukemia: case report and review of the literature. Int J Clin Exp Pathol 3(1):106–109
Abg Abd Wahab DY, Gau CH, Zakaria R, Muthu Karuppan MK, BS AR, Abdullah Z, Alrafiah A, Abdullah J M, Muthuraju S, (2019) Review on cross talk between neurotransmitters and neuroinflammation in striatum and cerebellum in the mediation of motor behaviour. BioMed Res Int 2019:1767203. https://doi.org/10.1155/2019/1767203
Abo-Al-Ela HG (2019) Toxoplasmosis and psychiatric and neurological disorders: a step toward understanding parasite pathogenesis. ACS Chem Neurosci 11(16):2393–2406. https://doi.org/10.1021/acschemneuro.9b00245
Ahlquist RP (1948) A study of the adrenotropic receptors. Am J Physiol C 153(3):586–600. https://doi.org/10.1152/ajplegacy.1948.153.3.586
Ahmadpour E, Daryani A, Sharif M, Sarvi S, Aarabi M, Mizani A, Rahimi MT, Shokri A (2014) Toxoplasmosis in immunocompromised patients in Iran: a systematic review and meta-analysis. J Infect Dev Ctries 8(12):1503–1510. https://doi.org/10.3855/jidc.4796
AL-Hadad MTS, Kadhim RA, Al-Rubaye AF (2019) Effect of chronic toxoplasmosis on levels of some neurotransmitters (Dopamine, Adrenaline, and Noradrenaline) in human serum. Int J Pharm Sci Res 11(2):402–405
Alonso R, Chaudieu I, Diorio J, Krishnamurthy A, Quirion R, Boksa P (1993) Interleukin-2 modulates evoked release of [3H] dopamine in rat cultured mesencephalic cells. J Neurochem 61(4):1284–1290. https://doi.org/10.1111/j.1471-4159.1993.tb13620.x
Amouei A, Moosazadeh M, Nayeri Chegeni T, Sarvi S, Mizani A, Pourasghar M, Hosseini Teshnizi S, Hosseininejad Z, Dodangeh S, Pagheh A, Pourmand AH, Daryani A (2020) Evolutionary puzzle of Toxoplasma gondii with suicidal ideation and suicide attempts: an updated systematic review and meta-analysis. Transbound Emerg Dis 0:1–14. https://doi.org/10.1111/tbed.13550
Andrade WA, Souza Mdo C, Ramos-Martinez E, Nagpal K, Dutra MS, Melo MB, Bartholomeu DC, Ghosh S, Golenbock DT, T. Gazzinelli R, (2013) Combined action of nucleic acid-sensing Toll-like receptors and TLR11/TLR12 heterodimers imparts resistance to Toxoplasma gondii in mice. Cell Host Microbe 13:42–53. https://doi.org/10.1016/j.chom.2012.12.003
Andrews N, Barnes NM, Steward LJ, West KE, Cunningham J, Wu PY, Zangrossi H Jr, File SE (1993) A comparison of rat brain amino acid and monoamine content in diazepam withdrawal and after exposure to a phobic stimulus. Br J Pharmacol 109(1):171. https://doi.org/10.1111/j.1476-5381.1993.tb13548.x
Anisman H, Merali Z, Hayley S (2008) Neurotransmitter, peptide and cytokine processes in relation to depressive disorder: comorbidity between depression and neurodegenerative disorders. Prog Neurobiol 85(1):1–74. https://doi.org/10.1016/j.pneurobio.2008.01.004
Arango V, Underwood MD, Mann JJ (1996) Fewer pigmented locus coeruleus neurons in suicide victims: preliminary results. Biol Psychiatry 39(2):112–120. https://doi.org/10.1016/0006-3223(95)00107-7
Arnold-Schrauf C, Berod L, Sparwasser T (2015) Dendritic cell specific targeting of MyD88 signalling pathways in vivo. Eur J Immunol 45:32–39. https://doi.org/10.1002/eji.201444747
Asada H, Kawamura Y, Maruyama K, Kume H, Ding R-G, Kanbara N, Kuzume H, Sanbo M, Yagi T, Obata K (1997) Cleft palate and decreased brain γ-aminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase. Proc Natl Acad Sci USA 94(12):6496–6499. https://doi.org/10.1073/pnas.94.12.6496
Axelrod J, Kopin I (1969) The uptake, storage, release and metabolism of noradrenaline in sympathetic nerves. Prog Brain Res 31:21–32. https://doi.org/10.1016/S0079-6123(08)63224-0
Ayano G (2016) Common neurotransmitters: criteria for neurotransmitters, key locations, classifications and functions. JAPN 1(1), 1–5. https://doi.org/10.11648/j.ajpn.20160406.12
Bak LK, Schousboe A, Waagepetersen HS (2006) The glutamate/GABA-glutamine cycle: aspects of transport, neurotransmitter homeostasis and ammonia transfer. J Neurochem 98(3):641–653. https://doi.org/10.1111/j.1471-4159.2006.03913.x
Baxter MG, Murray EA (2002) The amygdala and reward. Nat Rev Neurosci 3(7):563–573. https://doi.org/10.1038/nrn875
Bayani M, Riahi SM, Bazrafshan N, Ray Gamble H, Rostami A (2019) Toxoplasma gondii infection and risk of Parkinson and Alzheimer diseases: a systematic review and meta-analysis on observational studies. Acta Trop 196:165–171. https://doi.org/10.1016/j.actatropica.2019.05.015
Bazemore AEK, Florey E (1956) Factor I and gamma-aminobutyric acid. Nature 178:1052–1053. https://doi.org/10.1038/1781052a0
Beggiato S, Antonelli T, Tomasini MC, Tanganelli S, Fuxe K, Schwarcz R, Ferraro L (2013) Kynurenic acid, by targeting α7 nicotinic acetylcholine receptors, modulates extracellular GABA levels in the rat striatum in vivo. Eur J Neurosci 37(9):1470–1477. https://doi.org/10.1111/ejn.12160
Benarroch EE (2010) Glutamate transporters: diversity, function, and involvement in neurologic disease. J Neurol 74(3):259–264. https://doi.org/10.1212/WNL.0b013e3181cc89e3
Benítez-Diaz P, Miranda-Contreras L, Mendoza-Briceño RV, Peña-Contreras Z, Palacios-Prü E (2003) Prenatal and postnatal contents of amino acid neurotransmitters in mouse parietal cortex. Dev Neurosci 25(5):366–374. https://doi.org/10.1159/000073514
Bennett S, Grant MM, Aldred S (2009) Oxidative stress in vascular dementia and Alzheimer’s disease: a common pathology. J Alzheimers Dis 17(2):245–257. https://doi.org/10.3233/jad-2009-1041
Berdoy M, Webster JP, Macdonald DW (2000) Fatal attraction in rats infected with Toxoplasma gondii. Proc Biol Sci 267(1452):1591–1594. https://doi.org/10.1098/rspb.2000.1182
Berger-Sweeney J, Hohmann CF (1997) Behavioral consequences of abnormal cortical development: insights into developmental disabilities. Behav Brain Res 86(2):121–142. https://doi.org/10.1016/s0166-4328(96)02251-6
Biswas A, Bruder D, Wolf SA, Jeron A, Mack M, Heimesaat MM, Dunay IR (2015) Ly6C (high) monocytes control cerebral toxoplasmosis. J Immunol 194(7):3223–3235. https://doi.org/10.4049/jimmunol.1402037
Biswas A, Bruder D, Wolf SA, Jeron A, Mack M, Heimesaat MM, Dunay IR (2017) Behavior of neutrophil granulocytes during Toxoplasma gondii infection in the central nervous system. Front Cell Infect Microbiol 7:259. https://doi.org/10.3389/fcimb.2017.00259
Bolton DJ, Robertson LJ (2016) Mental health disorders associated with foodborne pathogens. J Food Prot 79(11):2005–2017. https://doi.org/10.4315/0362-028x.Jfp-15-587
Brooks JM, Carrillo GL, Su J, Lindsay DS, Fox MA, Blader I (2015) Toxoplasma gondii infections alter GABAergic synapses and signaling in the central nervous system. mBio 6(6), e01428–01415. https://doi.org/10.1128/mBio.01428-15
Butcher BA, Kim L, Johnson PF, Denkers EY (2001) Toxoplasma gondii tachyzoites inhibit proinflammatory cytokine induction in infected macrophages by preventing nuclear translocation of the transcription factor NF-kappa B. J Immunol 167(4):2193–2201. https://doi.org/10.4049/jimmunol.167.4.2193
Bylund DB, Eikenberg DC, Hieble JP, Langer SZ, Lefkowitz RJ, Minneman KP, Trendelenburg U (1994) International union of pharmacology nomenclature of adrenoceptors. Pharmacol Rev 46(2):121–136
Cáceda R (2014) Suicidal behavior: risk and protective factors. J Revista De Neuro-Psiquiatría 77(1):3–18
Campos AC, Vaz GN, Saito VM, Teixeira AL (2014) Further evidence for the role of interferon-gamma on anxiety-and depressive-like behaviors: involvement of hippocampal neurogenesis and NGF production. Neurosci Lett 578:100–105. https://doi.org/10.1016/j.neulet.2014.06.039
Capuron L, Miller AH (2011) Immune system to brain signaling: neuropsychopharmacological implications. Pharmacol Ther 130(2):226–238. https://doi.org/10.1016/j.pharmthera.2011.01.014
Capuron L, Schroecksnadel S, Féart C, Aubert A, Higueret D, Barberger-Gateau P, Layé S, Fuchs D (2011) Chronic low-grade inflammation in elderly persons is associated with altered tryptophan and tyrosine metabolism: role in neuropsychiatric symptoms. Biol Psychiatry 70(2):175–182. https://doi.org/10.1016/j.biopsych.2010.12.006
Carruthers VB, Suzuki Y (2007) Effects of Toxoplasma gondii infection on the brain. Schizophr Bull 33(3):745–751. https://doi.org/10.1093/schbul/sbm008
Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Taylor A, Poulton R (2002) Role of genotype in the cycle of violence in maltreated children. Science 297(5582):851–854. https://doi.org/10.1126/science.1072290
Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Taylor A, Poulton R (2003) Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301(5631):386–389. https://doi.org/10.1126/science.1083968
Cerávolo IP, Chaves AC, Bonjardim CA, Sibley D, Romanha AJ, Gazzinelli RT (1999) Replication of Toxoplasma gondii, but not Trypanosoma cruzi, is regulated in human fibroblasts activated with gamma interferon: requirement of a functional JAK/STAT pathway. Infect Immun 67(5):2233–2240
Choi DW (1994) Glutamate receptors and the induction of. J Neurosci 100:47. https://doi.org/10.2174/157015906776359540
Ciranna L (2006) Serotonin as a modulator of glutamate- and GABA-mediated neurotransmission: implications in physiological functions and in pathology. Curr Neuropharmacol 4(2):101–114. https://doi.org/10.2174/157015906776359540
Condie BG, Bain G, Gottlieb DI, Capecchi MR (1997) Cleft palate in mice with a targeted mutation in the γ-aminobutyric acid-producing enzyme glutamic acid decarboxylase 67. Proc Natl Acad Sci 94(21):11451–11455. https://doi.org/10.1073/pnas.94.21.11451
Cooper JR, Bloom FE, Roth RH (2003) The biochemical basis of neuropharmacology. Oxford University Press, USA
Cotecchia S, Stanasila L, Diviani D (2012) Protein-protein interactions at the adrenergic receptors. Curr Drug Targets 13(1):15–27. https://doi.org/10.2174/138945012798868489
Courtet P, Jollant F, Buresi C, Castelnau D, Mouthon D, Malafosse A (2005) The monoamine oxidase A gene may influence the means used in suicide attempts. Psychiatr Genet 15(3):189–193. https://doi.org/10.1097/00041444-200509000-00009
Courtois G, Gilmore TD (2006) Mutations in the NF-kappaB signaling pathway: implications for human disease. Oncogene 25(51):6831–6843. https://doi.org/10.1038/sj.onc.1209939
Croarkin PE, Levinson AJ, Daskalakis ZJ (2011) Evidence for GABAergic inhibitory deficits in major depressive disorder. Neurosci Biobehav Rev 35(3):818–825. https://doi.org/10.1016/j.neubiorev.2010.10.002
Curtis D, Watkins J (1960) The excitation and depression of spinal neurones by structurally related amino acids. J Neurochem 6(2):117–141. https://doi.org/10.1038/sj.onc.1209939
Czarnewski P, Araujo ECB, Oliveira MC, Mineo TWP, Silva NM (2017) Recombinant TgHSP70 immunization protects against Toxoplasma gondii brain cyst formation by enhancing inducible nitric oxide expression. Front Cell Infect Microbiol 7:142. https://doi.org/10.3389/fcimb.2017.00142
da Silva RC, Langoni H (2009) Toxoplasma gondii: host-parasite interaction and behavior manipulation. J Parasitol 105(4):893–898. https://doi.org/10.1007/s00436-009-1526-6
Daikhin Y, Yudkoff M (2000) Compartmentation of brain glutamate metabolism in neurons and glia. J Nutr 130(4S Suppl):1026s–1031s. https://doi.org/10.1093/jn/130.4.1026S
Dalimi A, Abdoli A (2012) Latent toxoplasmosis and human. Iran J Parasitol 7(1):1
Dard C, Sebastien Bailly S, Pepin JL, Brenier-Pinchart MP, Fricker-Hidalgo H, Peeters M, PellouxID H, Tamisier R (2020) Chronic toxoplasmosis and sleepiness in obstructive sleep apnea: Is there a link? PLoS ONE 15(7):e0235463. https://doi.org/10.1371/journal.pone.0235463
Dauvermann MR, Lee G, Dawson N (2017) Glutamatergic regulation of cognition and functional brain connectivity: insights from pharmacological, genetic and translational schizophrenia research. Br J Pharmacol 174(19):3136–3160. https://doi.org/10.1111/bph.13919
David CN, Frias ES, Szu JI, Vieira PA, Hubbard JA, Lovelace J, Michael M, Worth D, McGovern KE, Ethell IM, Stanley BG, Korzus E, Fiacco TA, BinderDK, Wilson EH (2016) GLT-1-dependent disruption of CNS glutamate homeostasis and neuronal function by the protozoan parasite Toxoplasma gondii. PLoS Pathog 12(6):e1005643. https://doi.org/10.1371/journal.ppat.1005643
Davis KL, Kahn RS, Ko G, Davidson M (1991) Dopamine in schizophrenia: a review and reconceptualization. Am J Psychiatry 148(11):1474–1486. https://doi.org/10.1176/ajp.148.11.1474
de Barros J, Barbosa IG, Salem H, Rocha NP, Kummer A, Okusaga OO, Soares JC, Teixeira AL (2017) Is there any association between Toxoplasma gondii infection and bipolar disorder? a systematic review and meta-analysis. J Affect Disord 209:59–65. https://doi.org/10.1016/j.jad.2016.11.016
Denkers EY, Gazzinelli RT (1998) Regulation and function of T-cell-mediated immunity during Toxoplasma gondii infection. Clin Microbiol Rev 11(4):569–588. https://doi.org/10.1128/cmr.11.4.569
Diamond A, Briand L, Fossella J, Gehlbach L (2004) Genetic and neurochemical modulation of prefrontal cognitive functions in children. Am J Psychiatry 161(1):125–132. https://doi.org/10.1176/appi.ajp.161.1.125
Dincel GC, Atmaca HT (2015) Nitric oxide production increases during Toxoplasma gondii encephalitis in mice. Exp Parasitol 156:104–112. https://doi.org/10.1016/j.exppara.2015.06.009
Dubey JP (1998) Advances in the life cycle of Toxoplasma gondii. Int J Parasitol 28(7):1019–1024. https://doi.org/10.1016/s0020-7519(98)00023-x
Dubey JP, Beattie CP (1988) Toxoplasmosis of animals and man. CRC Press Inc
Dunn AJ, Wang J (1995) Cytokine effects on CNS biogenic amines. NeuroImmunoModulation 2(6):319–328. https://doi.org/10.1159/000097211
Dupont CD, Christian DA, Hunter CA (2012) Immune response and immunopathology during toxoplasmosis. Semin Immunopathol 34(6):793–813. https://doi.org/10.1007/s00281-012-0339-3
Durk T, Panther E, Muller T, Sorichter S, Ferrari D, Pizzirani C, Di Virgilio F, Myrtek D, Norgauer J, Idzko M (2005) 5-Hydroxytryptamine modulates cytokine and chemokine production in LPS-primed human monocytes via stimulation of different 5-HTR subtypes. Int Immunol 17(5):599–606. https://doi.org/10.1093/intimm/dxh242
Easton A, Sankaranarayanan S, Tanghe A, Terwel D, Lin AX, Hoque N, Bourin C, Gu H, Ahlijanian M, Bristow L (2013) Effects of sub-chronic donepezil on brain Abeta and cognition in a mouse model of Alzheimer’s disease. J Psychopharmacol 230(2):279–289. https://doi.org/10.1007/s00213-013-3152-3
Egan CE, Sukhumavasi W, Butcher BA, Denkers EY (2009) Functional aspects of Toll-like receptor/MyD88 signalling during protozoan infection: focus on Toxoplasma gondii. Clin Exp Immunol 156:17–24. https://doi.org/10.1111/j.1365-2249.2009.03876.x
Egashira T (1976) Studies on monoamine oxidase. XVIII. Enzymic properties of placental monoamine oxidase. Jpn. J. Pharmacol. 26(4), 493–500. https://doi.org/10.1254/jjp.26.493
Elsheikha HM, Zhu XQ (2016) Toxoplasma gondii infection and schizophrenia: an inter-kingdom communication perspective. Curr Opin Infect Dis 29(3):311–318. https://doi.org/10.1097/qco.0000000000000265
Erdmann J, Stark K, Esslinger UB, Rumpf PM, Koesling D, De Wit C, Kaiser FJ, Braunholz D, Medack A, Fischer M (2013) Dysfunctional nitric oxide signalling increases risk of myocardial infarction. Nature 504(7480):432–436. https://doi.org/10.1038/nature12722
Erhardt S, Blennow K, Nordin C, Skogh E, Lindstrom LH, Engberg G (2001a) Kynurenic acid levels are elevated in the cerebrospinal fluid of patients with schizophrenia. Neurosci Lett 313(1–2):96–98. https://doi.org/10.1016/s0304-3940(01)02242-x
Erhardt S, Blennow K, Nordin C, Skogh E, Lindström LH, Engberg G (2001b) Kynurenic acid levels are elevated in the cerebrospinal fluid of patients with schizophrenia. Neurosci Lett 313(1–2):96–98. https://doi.org/10.1016/s0304-3940(01)02242-x
Erhardt S, Lim CK, Linderholm KR, Janelidze S, Samuelsson M, Lundberg K, Postolache TT, Braunholz D, Träskman-Bendz L, Guillemin JG, Brundin L (2013) Connecting inflammation with glutamate agonism in suicidality. Neuropsychopharmacology 38(5):743–752. https://doi.org/10.1038/npp.2012.248
Fabiani S, Pinto B, Bruschi F (2013) Toxoplasmosis and neuropsychiatric diseases: can serological studies establish a clear relationship? Neurol Sci 34(4):417–425. https://doi.org/10.1007/s10072-012-1197-4
Fekadu A, Shibre T, Cleare AJ (2010) Toxoplasmosis as a cause for behaviour disorders--overview of evidence and mechanisms. Folia Parasitol 57(2), 105–113. https://doi.org/10.14411/fp.2010.013
Feldman RS, Meyer JS, Quenzer LF, Cooper JR (1997) Principles of neuropsychopharmacology. Sinauer Associates Sunderland, MA
Fernandes Azevedo B, Barros Furieri L, Pecanha FM, Wiggers GA, Frizera Vassallo P, Ronacher Simoes M, Fiorim J, Rossi de Batista P, Fioresi M, Rossoni L, Stefanon I, Alonso MJ, Salaices M, Valentim Vassallo D (2012) Toxic effects of mercury on the cardiovascular and central nervous systems. J Biomed Sci 2012:949048. https://doi.org/10.1155/2012/949048
Flegr J (2015) Neurological and neuropsychiatric consequences of chronic Toxoplasma infection. Curr Clin Microbiol Rep 2(4):163–172
Flegr J, Kuba R (2016) The relation of Toxoplasma infection and sexual attraction to fear, danger, pain, and submissiveness. Evol Psychol 14(3):1474704916659746
Flegr J, Klapilova K, Kankova S (2014) Toxoplasmosis can be a sexually transmitted infection with serious clinical consequences. Not all routes of infection are created equal. Med Hypotheses 83(3), 286–289. https://doi.org/10.1016/j.mehy.2014.05.019
Flegr J, Prandota J, Sovickova M, Israili ZH (2014) Toxoplasmosis--a global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS One 9(3), e90203. https://doi.org/10.1371/journal.pone.0090203
Frimpter GW, Andelman RJ, George WF (1969) Vitamin B6-dependency syndromes: new horizons in nutrition. Am J Clin Nutr 22(6):794–805. https://doi.org/10.1093/ajcn/22.6.794
Fuglewicz AJ, Piotrowski P, Stodolak A (2017) Relationship between toxoplasmosis and schizophrenia: a review. Adv Clin Exp Med 26(6), 1031–1036. https://doi.org/10.17219/acem/61435
Fujigaki S, Saito K, Takemura M, Maekawa N, Yamada Y, Wada H, Seishima M (2002) L-tryptophan-L-kynurenine pathway metabolism accelerated by Toxoplasma gondii infection is abolished in gamma interferon-gene-deficient mice: cross-regulation between inducible nitric oxide synthase and indoleamine-2,3-dioxygenase. Infect Immun 70(2):779–786. https://doi.org/10.1128/iai.70.2.779-786.2002
Fujigaki S, Takemura M, Hamakawa H, Seishima M, Saito K (2003) The mechanism of interferon-gamma induced anti Toxoplasma gondii by indoleamine 2,3-dioxygenase and/or inducible nitric oxide synthase vary among tissues. Adv Exp Med Biol 527:97–103. https://doi.org/10.1007/978-1-4615-0135-0_11
Fuks JM, Arrighi RB, Weidner JM, Mendu SK, Jin Z, Wallin RP, Rethi B, Birnir B, Barragan A (2012) GABAergic signaling is linked to a hypermigratory phenotype in dendritic cells infected by Toxoplasma gondii. PLoS pathogens 8(12). https://doi.org/10.1371/journal.ppat.1003051
Furtado JM, Smith JR, Belfort R Jr, Gattey D, Winthrop KL (2011) Toxoplasmosis: a global threat. J Glob Infect Dis 3(3):281–284. https://doi.org/10.4103/0974-777x.83536
Galvan-Ramirez Mde L, Gutierrez-Maldonado A, Verduzco-Grijalva F, Jimenez JM (2014) The role of hormones on Toxoplasma gondii infection: a systematic review. Front Microbiol 5:503. https://doi.org/10.3389/fmicb.2014.00503
Garry PS, Ezra M, Rowland MJ, Westbrook J, Pattinson KT (2015) The role of the nitric oxide pathway in brain injury and its treatment–from bench to bedside. Exp Neurol 263:235–243. https://doi.org/10.1016/j.expneurol.2014.10.017
Gaskell EA, Smith JE, Pinney JW, Westhead DR, McConkey GA (2009) A unique dual activity amino acid hydroxylase in Toxoplasma gondii. PLoS ONE 4(3):e4801. https://doi.org/10.1371/journal.pone.0004801
Gaspar P, Cases O, Maroteaux L (2003) The developmental role of serotonin: news from mouse molecular genetics. Nat Rev Neurosci 4(12):1002–1012. https://doi.org/10.1038/nrn1256
Gatkowska J, Wieczorek M, Dziadek B, Dzitko K, Dlugonska H (2012) Behavioral changes in mice caused by Toxoplasma gondii invasion of brain. Parasitol Res 111(1):53–58. https://doi.org/10.1007/s00436-011-2800-y
Gatkowska J, Wieczorek M, Dziadek B, Dzitko K, Dlugonska H (2013) Sex-dependent neurotransmitter level changes in brains of Toxoplasma gondii infected mice. Exp Parasitol 133(1):1–7. https://doi.org/10.1016/j.exppara.2012.10.005
Giacobini E (1996) New trends in cholinergic therapy for Alzheimer disease: nicotinic agonists or cholinesterase inhibitors? Prog Brain Res 109:311–323. https://doi.org/10.1016/s0079-6123(08)62114-7
Goff DC (2015) Drug development in schizophrenia: are glutamatergic targets still worth aiming at? Curr Opin Psychiatry 28(3):207–215. https://doi.org/10.1097/yco.0000000000000152
Gohardehi S, Sharif M, Sarvi S, Moosazadeh M, Alizadeh-Navaei R, Hosseini SA, Amouei A, Pagheh A, Sadeghi M, Daryani A (2018) The potential risk of toxoplasmosis for traffic accidents: a systematic review and meta-analysis. Exp Parasitol 191:19–24. https://doi.org/10.1016/j.exppara.2018.06.003
Gonzalez LE, Rojnik B, Urrea F, Urdaneta H, Petrosino P, Colasante C, Pino S, Hernandez L (2007) Toxoplasma gondii infection lower anxiety as measured in the plus-maze and social interaction tests in rats A behavioral analysis. Behav Brain Res 177(1):70–79. https://doi.org/10.1016/j.bbr.2006.11.012
Goodwin D, Hrubec TC, Klein BG, Strobl JS, Werre SR, Han Q, Zajac AM, Lindsay DS (2012) Congenital infection of mice with Toxoplasma gondii induces minimal change in behavior and no change in neurotransmitter concentrations. J Parasitol 98(4):706–712. https://doi.org/10.1645/ge-3068.1
Graeff FG, Viana MB, Mora PO (1997) Dual role of 5-HT in defense and anxiety. Neurosci Biobehav Rev 21(6):791–799. https://doi.org/10.1016/s0149-7634(96)00059-0
Grimsby J, Lan NC, Neve R, Chen K, Shih JC (1990) Tissue distribution of human monoamine oxidase A and B mRNA. J Neurochem 55(4):1166–1169. https://doi.org/10.1111/j.1471-4159.1990.tb03121.x
Grimsby J, Chen K, Wang L-J, Lan NC, Shih JC (1991) Human monoamine oxidase A and B genes exhibit identical exon-intron organization. Proc Natl Acad Sci 88(9):3637–3641
Guidetti P, Bates GP, Graham RK, Hayden MR, Leavitt BR, MacDonald ME, Slow EJ, Wheeler VC, Woodman B, Schwarcz R (2006) Elevated brain 3-hydroxykynurenine and quinolinate levels in Huntington disease mice. Neurobiol Dis 23(1):190–197. https://doi.org/10.1016/j.nbd.2006.02.011
Guillemin GJ, Smythe G, Takikawa O, Brew BJ (2005) Expression of indoleamine 2,3-dioxygenase and production of quinolinic acid by human microglia, astrocytes, and neurons. Glia 49(1):15–23. https://doi.org/10.1002/glia.20090
Gulaj E, Pawlak K, Bien B, Pawlak D (2010) Kynurenine and its metabolites in Alzheimer’s disease patients. Adv Med Sci 55(2):204–211. https://doi.org/10.2478/v10039-010-0023-6
Hamie M, Najm R, Deleuze-Masquefa C, Antoine Bonnet P, Dubremetz JF, El Sabban M, El Hajj H (2021) Imiquimod targets toxoplasmosis through modulating host Toll-like receptor-MyD88 signaling. Front Immunol 12:629917. https://doi.org/10.3389/fimmu.2021.629917
Hamon M, Blier P (2013) Monoamine neurocircuitry in depression and strategies for new treatments. Prog Neuropsychopharmacol Biol Psychiatry 45:54–63. https://doi.org/10.1016/j.pnpbp.2013.04.009
Hari Dass SA, Vyas A (2014) Toxoplasma gondii infection reduces predator aversion in rats through epigenetic modulation in the host medial amygdala. Mol Ecol 23(24):6114–6122
Haroon E, Raison CL, Miller AH (2012) Psychoneuroimmunology meets neuropsychopharmacology: translational implications of the impact of inflammation on behavior. Neuropsychopharmacology 37(1):137–162. https://doi.org/10.1038/npp.2011.205
Hayashi T (1952) A physiological study of epileptic seizures following cortical stimulation in animals and its application to human clinics. Jpn J Physiol 3:46–64. https://doi.org/10.2170/jjphysiol.3.46
Henriquez SA, Brett R, Alexander J, Pratt J, Roberts CW (2009) Neuropsychiatric disease and Toxoplasma gondii infection. NeuroImmunoModulation 16(2):122–133. https://doi.org/10.1159/000180267
Herlenius E, Lagercrantz H (2004) Development of neurotransmitter systems during critical periods. Exp Neurol 190:8–21. https://doi.org/10.1016/j.expneurol.2004.03.027
Hilmas C, Pereira EF, Alkondon M, Rassoulpour A, Schwarcz R, Albuquerque EX (2001) The brain metabolite kynurenic acid inhibits α7 nicotinic receptor activity and increases non-α7 nicotinic receptor expression: physiopathological implications. J Neurosci 21(19):7463–7473. https://doi.org/10.1523/JNEUROSCI.21-19-07463.2001
Hinze-Selch D (2015) Toxoplasma gondii infection and neuropsychiatric disease: current insight. Reports Parasitol 4:43–51. https://doi.org/10.2147/RIP.S52980
Hofmann S, Philbrook C, Gerbitz K-D, Bauer MF (2003) Wolfram syndrome: structural and functional analyses of mutant and wild-type wolframin, the WFS1 gene product. Hum Mol Genet 12(16):2003–2012. https://doi.org/10.1093/hmg/ddg214
Hohmann CF (2003) A morphogenetic role for acetylcholine in mouse cerebral neocortex. Neurosci Biobehav Rev 27(4):351–363. https://doi.org/10.1016/s0149-7634(03)00066-6
Houser C, Crawford G, Salvaterra P, Vaughn J (1985) Immunocytochemical localization of choline acetyltransferase in rat cerebral cortex: a study of cholinergic neurons and synapses. J Comp Neurol 234(1):17–34. https://doi.org/10.1002/cne.902340103
Hsu YPP, Weyler W, Chen S, Sims KB, Rinehart WB, Utterback MC, Powell JF, Breakefield XO (1988) Structural features of human monoamine oxidase A elucidated from cDNA and peptide sequences. J Neurochem 51(4):1321–1324. https://doi.org/10.1111/j.1471-4159.1988.tb03105.x
Hsu PC, Groer M, Beckie T (2014) New findings: depression, suicide, and Toxoplasma gondii infection. J Am Assoc Nurse Pract 26(11):629–637. https://doi.org/10.1002/2327-6924.12129
Hull EM, Du J, Lorrain DS, Matuszewich L (1995) Extracellular dopamine in the medial preoptic area: implications for sexual motivation and hormonal control of copulation. J Neurosci 15(11):7465–7471
Hull EM, Muschamp JW, Sato S (2004) Dopamine and serotonin: influences on male sexual behavior. Physiol Behav 83(2):291–307. https://doi.org/10.1016/j.physbeh.2004.08.018
Hunter CA, Candolfi E, Subauste C, Van Cleave V, Remington JS (1995) Studies on the role of interleukin-12 in acute murine toxoplasmosis. Immunology 84(1):16–20
Hunter C, Suzuki Y, Subauste C, Remington J (1996) Cells and cytokines in resistance to Toxoplasma gondii, Toxoplasma gondii. Springer 113–125.
Hurley RA, Taber KH (2012) Latent Toxoplasma gondii: emerging evidence for influences on neuropsychiatric disorders. J Neuropsychiatry Clin Neurosci 24(4):376–383. https://doi.org/10.1176/appi.neuropsych.12100234
Ihara F, Nishimura M, Muroi Y, Mahmoud ME, Yokoyama N, Nagamune K, Nishikawa Y (2016) Toxoplasma gondii infection in mice impairs long-term fear memory consolidation through dysfunction of the cortex and amygdala. Infect Immun 84(10):2861–2870. https://doi.org/10.1128/iai.00217-16
Javitt DC (2014) Current and emergent treatments for symptoms and neurocognitive impairment in schizophrenia. Curr Treat Options Psychiatry 1(2):107–120. https://doi.org/10.1007/s40501-014-0010-9
Johnston MV, Silverstein FS (2003) The newborn brain: neuroscience and clinical applications. Cambridge University Press115
Jones JL, Lopez A, Wilson M (2003) Congenital Toxoplasmosis. Am Fam Physician 67(10):2131–2138
Jun CD, Kim SH, Soh CT, Kang SS, Chung HT (1993) Nitric oxide mediates the toxoplasmastatic activity of murine microglial cells in vitro. Immunol Invest 22(8):487–501. https://doi.org/10.3109/08820139309084178
Kalueff AV, Nutt DJ (2007) Role of GABA in anxiety and depression. J Depress Anxiety 24(7):495–517. https://doi.org/10.1002/da.20262
Kannan G, Crawford JA, Yang C, Gressitt KL, Ihenatu C, Krasnova IN, Cadet JL, Yolken RH, Severance EG, Pletnikov MV (2016) Anti-NMDA receptor autoantibodies and associated neurobehavioral pathology in mice are dependent on age of first exposure to Toxoplasma gondii. Neurobiol Dis 91:307–314. https://doi.org/10.1016/j.nbd.2016.03.005
Kannan G, Gressitt K, Yang S, Stallings C, Katsafanas E, Schweinfurth L, Savage C, Adamos M, Sweeney K, Origoni A (2017) Pathogen-mediated NMDA receptor autoimmunity and cellular barrier dysfunction in schizophrenia. Transl Psychiatry 7(8):e1186–e1186. https://doi.org/10.1038/tp.2017.162
Karolewicz B, Maciag D, O’Dwyer G, Stockmeier CA, Feyissa AM, Rajkowska G (2010) Reduced level of glutamic acid decarboxylase-67 kDa in the prefrontal cortex in major depression. Int J Neuropsychopharmacol 13(4):411–420. https://doi.org/10.1017/S1461145709990587
Kegel ME, Bhat M, Skogh E, Samuelsson M, Lundberg K, Dahl ML, Sellgren C, Schwieler L, Engberg G, Schuppe-Koistinen I, Erhardt S (2014) Imbalanced kynurenine pathway in schizophrenia. Int J Tryptophan Res 7:15–22. https://doi.org/10.4137/ijtr.S16800
Khan IA, Schwartzman JD, Matsuura T, Kasper LH (1997) A dichotomous role for nitric oxide during acute Toxoplasma gondii infection in mice. Proc Natl Acad Sci USA 94(25):13955–13960. https://doi.org/10.1073/pnas.94.25.13955
Khan IA, MacLean JA, Lee FS, Casciotti L, DeHaan E, Schwartzman JD, Luster AD (2000) IP-10 is critical for effector T cell trafficking and host survival in Toxoplasma gondii infection. Immunity 12(5):483–494. https://doi.org/10.1016/s1074-7613(00)80200-9
King MD, Lindsay DS, Holladay S, Ehrich M (2003) Neurotoxicity and immunotoxicity assessment in CBA/J mice with chronic Toxoplasma gondii infection and single-dose exposure to methylmercury. Int J Toxicol 22(1):53–61. https://doi.org/10.1080/10915810305075
Kitagami T, Yamada K, Miura H, Hashimoto R, Nabeshima T, Ohta T (2003) Mechanism of systemically injected interferon-alpha impeding monoamine biosynthesis in rats: role of nitric oxide as a signal crossing the blood-brain barrier. Brain Res 978(1–2):104–114. https://doi.org/10.1016/s0006-8993(03)02776-8
Klaren VN, Kijlstra A (2002) Toxoplasmosis, an overview with emphasis on ocular involvement. Ocul Immunol Inflamm 10(1):1–26. https://doi.org/10.1076/ocii.10.1.1.10330
Klein SL (2003) Parasite manipulation of the proximate mechanisms that mediate social behavior in vertebrates. Physiol Behav 79(3):441–449. https://doi.org/10.1016/s0031-9384(03)00163-x
Koblansky AA, Jankovic D, Oh H, Hieny S, Sungnak W, Mathur R, Hayden MS, Akira S, Sher A, Ghosh S (2013) Recognition of profilin by Toll-like receptor 12 is critical for host resistance to Toxoplasma gondii. Immunity 38:119–130. https://doi.org/10.1016/j.immuni.2012.09.016
Kõks S, Planken A, Luuk H, Vasar E (2002) Cat odour exposure increases the expression of wolframin gene in the amygdaloid area of rat. Neurosci Lett 322(2):116–120. https://doi.org/10.1016/s0304-3940(02)00110-6
Konradt C, Ueno N, Christian DA, Delong JH, Pritchard GH, Herz J, Bzik DJ, Koshy AA, McGavern DB, Lodoen MB, Hunter CA (2016) Endothelial cells are a replicative niche for entry of Toxoplasma gondii to the central nervous system. Nat Microbiol 1:16001. https://doi.org/10.1038/nmicrobiol.2016.1
Kucera K, Koblansky AA, Saunders LP, Frederick KB, De La Cruz EM, Ghosh S, Modisa Y (2010) Structure-based analysis of Toxoplasma gondii profilin: a parasitespecific motif is required for recognition by Toll-like receptor 11. J Mol Biol 403:616–629. https://doi.org/10.1016/j.jmb.2010.09.022
Kusbeci OY, Miman O, Yaman M, Aktepe OC, Yazar S (2011) Could Toxoplasma gondii have any role in Alzheimer disease? Alzheimers Dis Assoc Disord 25:1–3. https://doi.org/10.1097/WAD.0b013e3181f73bc2
Lamadrid-Romero M, Solís KH, Cruz-Reséndiz MS, Pérez JE, Díaz NF, Flores-Herrera H, García-López G, Perichart O, Reyes-Munoz E, Arenas-Huertero F, Eguía-Aguilar P, Molina-Hernández A (2018) Central nervous system development-related microRNAs levels increase in the serum of gestational diabetic women during the first trimester of pregnancy. Neurosci Res 130:8–22. https://doi.org/10.1016/j.neures.2017.08.003
Lang D, Schott BH, van Ham M, Morton L, Kulikovskaja L, Herrera-Molina R, Pielot R, Klawonn F, Montag D, Jänsch L (2018) Chronic Toxoplasma infection is associated with distinct alterations in the synaptic protein composition. J Neuroinflammation 15(1):216. https://doi.org/10.1186/s12974-018-1242-1
Laruelle M, Abi-Dargham A, van Dyck CH, Gil R, D’Souza CD, Erdos J, McCance E, Rosenblatt W, Fingado C, Zoghbi SS, Baldwin RM, Seibyl JP, Krystal JH, Charney DS, Innis RB (1996) Single photon emission computerized tomography imaging of amphetamine-induced dopamine release in drug-free schizophrenic subjects. Proc Natl Acad Sci USA 93(17):9235–9240. https://doi.org/10.1073/pnas.93.17.9235
Lau JY, Goldman D, Buzas B, Fromm SJ, Guyer AE, Hodgkinson C, Monk CS, Nelson EE, Shen PH, Pine DS, Ernst M (2009) Amygdala function and 5-HTT gene variants in adolescent anxiety and major depressive disorder. Biol Psychiatry 65(4):349–355. https://doi.org/10.1016/j.biopsych.2008.08.037
Lavoie A, Twyman R (1996) Direct evidence for diazepam modulation of GABAA receptor microscopic affinity. Neuropharmacology 35(9–10):1383–1392. https://doi.org/10.1016/s0028-3908(96)00077-9
Leite AZ, Rodrigues NC, Gonzaga MI, Paiolo JCC, de Souza CA, Stefanutto NAV, Omori WP, Pinheiro DG, Brisotti JL, Matheucci Junior E, Mariano VS, de Oliveira GLV (2017) Detection of increased plasma interleukin-6 levels and prevalence of Prevotella copri and Bacteroides vulgatus in the feces of type 2 diabetes patients. Front Immunol 8:1107. https://doi.org/10.3389/fimmu.2017.01107
Lesch KP, Bengel., Heils A, Sabol S., Greenberg BD, Petri S, Benjamin J, Muller CR, Hamer D H, Murphy DL, (1996) Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274(5292):1527–1531. https://doi.org/10.1126/science.274.5292.1527
Levinson AJ, Fitzgerald PB, Favalli G, Blumberger DM, Daigle M, Daskalakis ZJ (2010) Evidence of cortical inhibitory deficits in major depressive disorder. Biol Psychiatry 67(5):458–464. https://doi.org/10.1016/j.neubiorev.2010.10.002
Lewinsohn R, Glover V, Sandler M (1980) Development of benzylamine oxidase and monoamine oxidase A and B in man. Biochem Pharmacol 29(9):1221–1230. https://doi.org/10.1016/0006-2952(80)90278-6
Li YE, Kannan G, Pletnikov MV, Yolken RH, Xiao J (2015) Chronic infection of Toxoplasma gondii downregulates miR-132 expression in multiple brain regions in a sex-dependent manner. Parasitology 142(4):623–632. https://doi.org/10.1017/s003118201400167x
Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, Bartel DP, Linsley PS, Johnson JM (2005) Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 433(7027):769–773. https://doi.org/10.1038/nature03315
Litteljohn D, Nelson E, Hayley S (2014) IFN-γ differentially modulates memory-related processes under basal and chronic stressor conditions. Front Cell Neurosci 8:391. https://doi.org/10.3389/fncel.2014.00391
Lucchese G (2017) From toxoplasmosis to schizophrenia via NMDA dysfunction: peptide overlap between Toxoplasma gondii and N-methyl-d-aspartate receptors as a potential mechanistic link. Front Psychiatry 8:37. https://doi.org/10.3389/fpsyt.2017.00037
MacRae JI, Sheiner L, Nahid A, Tonkin C, Striepen B, McConville MJ (2012) Mitochondrial metabolism of glucose and glutamine is required for intracellular growth of Toxoplasma gondii. Cell Host Microbe 12(5):682–692. https://doi.org/10.1016/j.chom.2012.09.013
Maggi L, Le Magueresse C, Changeux J-P, Cherubini E (2003) Nicotine activates immature “silent” connections in the developing hippocampus. J Proc Natl Acad Sci 100(4):2059–2064. https://doi.org/10.1073/pnas.0437947100
Mahmoud ME, Ihara F, Fereig RM, Nishimura M, Nishikawa Y (2016) Induction of depression-related behaviors by reactivation of chronic Toxoplasma gondii infection in mice. Behav Brain Res 298(Pt B):125–133. https://doi.org/10.1016/j.bbr.2015.11.005
Mahmoud M E, Fereig R, Nishikawa Y (2017) Involvement of host defense mechanisms against Toxoplasma gondii infection in anhedonic and despair-like behaviors in mice. Infect Immun 85(4). doi:https://doi.org/10.1128/iai.00007-17
Mahmoudvand H, Sheibani V, Keshavarz H, Shojaee S, Esmaeelpour K, Ziaali N (2016) Acetylcholinesterase inhibitor improves learning and memory impairment induced by Toxoplasma gondii infection. Iran J Parasitol 11(2):177
Marín O, Rubenstein JL (2001) A long, remarkable journey: tangential migration in the telencephalon. Nat Rev Neurosci 2(11):780–790. https://doi.org/10.1038/35097509
McConkey GA, Martin HL, Bristow GC, Webster JP (2013) Toxoplasma gondii infection and behaviour - location, location, location? J Exp Biol 216(Pt 1):113–119. https://doi.org/10.1242/jeb.074153
Melamede R (2009) Parasitic brain infection, endocannabinoids, and schizophrenia. Med Hypotheses 72(2):220–222. https://doi.org/10.1016/j.mehy.2008.08.025
Meldrum BS (2000) Glutamate as a neurotransmitter in the brain: review of physiology and pathology. J Nutr 130(4):1007S-1015S. https://doi.org/10.1093/jn/130.4.1007S
Mentzer A, Perry M, Fitzgerald N, Barrington S, Siddiqui A, Kulasegaram R (2012) Is it all cerebral toxoplasmosis? Lancet 379(9812):286. https://doi.org/10.1016/s0140-6736(11)61579-1
Miller AH, Maletic V, Raison CL (2009a) Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry 65(9):732–741
Miller CM, Boulter NR, Ikin RJ, Smith NC (2009b) The immunobiology of the innate response to Toxoplasma gondii. Int J Parasitol 39(1):23–39. https://doi.org/10.1016/j.ijpara.2008.08.002
Miller BH, Zeier Z, Xi L, Lanz TA, Deng S, Strathmann J, Willoughby D, Kenny PJ, Elsworth JD, Lawrence MS, Roth RH, Edbauer D, Kleiman RJ, Wahlestedt C (2012) MicroRNA-132 dysregulation in schizophrenia has implications for both neurodevelopment and adult brain function. Proc Natl Acad Sci USA 109(8):3125–3130. https://doi.org/10.1073/pnas.1113793109
Moghaddam B, Javitt D (2012) From revolution to evolution: the glutamate hypothesis of schizophrenia and its implication for treatment. Neuropsychopharmacology 37(1):4–15. https://doi.org/10.1038/npp.2011.181
Mohle L, Parlog A, Pahnke J, Dunay IR (2014) Spinal cord pathology in chronic experimental Toxoplasma gondii infection. Eur J Immunol (bp) 4(1):65–75. https://doi.org/10.1556/EuJMI.4.2014.1.6
Mohle L, Israel N, Paarmann K, Krohn M, Pietkiewicz S, Muller A, Lavrik IN, Buguliskis JS, Schott BH, Schluter D, Gundelfinger ED, Montag D, Seifert U, Pahnke J, Dunay IR (2016) Chronic Toxoplasma gondii infection enhances beta-amyloid phagocytosis and clearance by recruited monocytes. Acta Neuropathol Commun 4:25. https://doi.org/10.1186/s40478-016-0293-8
Montoya JG, Liesenfeld O (2004) Toxoplasmosis. Lancet 363:1965–1975
Mortensen PB, Norgaard-Pedersen B, Waltoft BL, Sorensen TL, Hougaard D, Yolken RH (2007) Early infections of Toxoplasma gondii and the later development of schizophrenia. Schizophr Bull 33(3):741–744. https://doi.org/10.1093/schbul/sbm009
Müller N, Schwarz M (2007) The immunological basis of glutamatergic disturbance in schizophrenia: towards an integrated view. J Neural Transm 269-80.https://doi.org/10.1007/978-3-211-73574-9_33
Muller N, Schwarz M (2006) Schizophrenia as an inflammation-mediated dysbalance of glutamatergic neurotransmission. Neurotox Res 10(2):131–148. https://doi.org/10.1007/bf03033242
Munoz M, Liesenfeld O, Heimesaat MM (2011) Immunology of Toxoplasma gondii. J Immunol Rev 240(1):269–285. https://doi.org/10.1111/j.1600-065X.2010.00992.x
Murakami Y, Hoshi M, Hara A, Takemura M, Arioka Y, Yamamoto Y, Matsunami H, Funato T, Seishima M, Saito K (2012) Inhibition of increased indoleamine 2,3-dioxygenase activity attenuates Toxoplasma gondii replication in the lung during acute infection. Cytokine 59(2):245–251. https://doi.org/10.1016/j.cyto.2012.04.022
Murphy DL, Garrick NA, Aulakh CS, Cohen RM (1984) New contributions for basic science to understanding the effects of monoamine oxidase inhibiting antidepressants. J Clin Psychiatry 45:37–43
Mustafa AK, Gadalla MM, Snyder SH (2009) Signaling by gasotransmitters. J Sci Signal 2(68), re2-re2. https://doi.org/10.1126/scisignal.268re2
Nakamura T, Sato A, Kitsukawa T, Momiyama T, Yamamori T, Sasaoka T (2014) Distinct motor impairments of dopamine D1 and D2 receptor knockout mice revealed by three types of motor behavior. Front Integr Neurosci 8:56. https://doi.org/10.3389/fnint.2014.00056
Nath A, Hobson DE, Russel A (1993) Movement disorders with cerebral toxoplasmosis and AIDS. Mov Disord 8(1):107–112. https://doi.org/10.1002/mds.870080119
Nayeri T, Sarvi S, Moosazadeh M, Hosseininejad Z, Sharif M, Amouei A, Daryani A (2020b) Relationship between toxoplasmosis and autism: a systematic review and meta-analysis. Microb Pathog 147:104434. https://doi.org/10.1016/j.micpath.2020.104434
Nayeri T, Sarvi S, Moosazadeh M, Hosseininejad Z, Sharif M, Amouei A, Daryani A (2020c) Association between Toxoplasma gondii infection and headache: a systematic review and meta-analysis. Infect Disord Drug Targets 20:1–8. https://doi.org/10.2174/1871526520666200617135851
Nayeri Chegeni T, Sarvi S, Amouei MM, Hosseininejad Z, Aghayan S, Daryani A (2019b) Relationship between toxoplasmosis and obsessive compulsive disorder: a systematic review and meta-analysis. PLOS Negl Trop Dis 13(4):e0007306. https://doi.org/10.1371/journal.pntd.0007306
Nayeri Chegeni T, Sarvi S, Moosazadeh M, Sharif M, Aghayan SA, Amouei A, Hosseininejad Z, Daryani A (2019c) Is Toxoplasma gondii a potential risk factor for Alzheimer’s disease? a systematic review and meta-analysis. Microb Pathog 137:103751. https://doi.org/10.1016/j.micpath.2019.103751
Nayeri Chegeni T, Sharif M, Sarvi S, Moosazadeh M, Montazeri M, Aghayan SA, Balalami NJ, Gholami S, Hosseininejad Z, Saberi R, Anvari D, Gohardehi S, Daryani A (2019a) Is there any association between Toxoplasma gondii infection and depression? a systematic review and meta-analysis. PLoS ONE 14(6):e0218524. https://doi.org/10.1371/journal.pone.0218524
Nayeri T, Sarvi S, Moosazadeh M, Hosseininejad Z, Amouei A, Daryani A (2020a) Toxoplasma gondii infection and risk of attention-deficit hyperactivity disorder: a systematic review and meta-analysis. Pathog Glob Health 1-10.https://doi.org/10.1080/20477724.2020.1738153
Neurauter G, Schrocksnadel K, Scholl-Burgi S, Sperner-Unterweger B, Schubert C, Ledochowski M, Fuchs D (2008) Chronic immune stimulation correlates with reduced phenylalanine turnover. Curr Drug Metab 9(7):622–627. https://doi.org/10.2174/138920008785821738
Ngô HM, Zhou Y, Lorenzi H, Wang K, Kim T-K, Zhou Y, El Bissati K, Mui E, Fraczek L, Rajagopala SV (2017) Toxoplasma modulates signature pathways of human epilepsy, neurodegeneration & cancer. Sci Rep 7(1):11496. https://doi.org/10.1038/s41598-017-10675-6
Ngoungou EB, Bhalla D, Nzoghe A, Darde ML, Preux PM (2015) Toxoplasmosis and epilepsy–systematic review and meta analysis. PLOS Negl Trop Dis 9(2):e0003525. https://doi.org/10.1371/journal.pntd.0003525
Nilsson L, Linderholm K, Engberg G, Paulson L, Blennow K, Lindström L, Nordin C, Karanti A, Persson P, Erhardt S (2005) Elevated levels of kynurenic acid in the cerebrospinal fluid of male patients with schizophrenia. J Schizophr Res 80(2–3):315–322. https://doi.org/10.1016/j.biopsych.2005.06.024
Nilsson KW, Sjoberg RL, Damberg M, Leppert OJ, Alm PO, Lindstrom L, Oreland L (2006) Role of monoamine oxidase A genotype and psychosocial factors in male adolescent criminal activity. Biol Psychiatry 59(2):121–127. https://doi.org/10.1016/j.biopsych.2005.06.024
Nilsson KW, Wargelius HL, Sjoberg RL, Leppert J, Oreland L (2008) The MAO-A gene, platelet MAO-B activity and psychosocial environment in adolescent female alcohol-related problem behaviour. Drug Alcohol Depend 93(1–2):51–62. https://doi.org/10.1016/j.drugalcdep.2007.08.022
Notarangelo FM, Wilson EH, Horning KJ, Thomas MA, Harris TH, Fang Q, Hunter CA, Schwarcz R (2014) Evaluation of kynurenine pathway metabolism in Toxoplasma gondii-infected mice: implications for schizophrenia. Schizophr Res 152(1):261–267. https://doi.org/10.1016/j.schres.2013.11.011
Ohno Y (2011) Therapeutic role of 5-HT1A receptors in the treatment of schizophrenia and Parkinson’s disease. CNS Neurosci Ther 17(1):58–65. https://doi.org/10.1111/j.1755-5949.2010.00211.x
Ordway GA, Widdowson PS, Smith KS, Halaris A (1994) Agonist binding to α2-adrenoceptors is elevated in the locus coeruleus from victims of suicide. J Neurochem 63(2):617–624. https://doi.org/10.1046/j.1471-4159.1994.63020617.x
Ortiz PA, Garvin JL (2003) Cardiovascular and renal control in NOS-deficient mouse models. Am J Physiol Regul Integr Comp Physiol 284(3), R628–638. https://doi.org/10.1152/ajpregu.00401.2002
Oxenkrug G (2013) Serotonin-kynurenine hypothesis of depression: historical overview and recent developments. Curr Drug Targets 14(5):514–521. https://doi.org/10.2174/1389450111314050002
Palmer BS( 2007) Meta-analysis of three case controlled studies and an ecological study into the link between cryptogenic epilepsy and chronic toxoplasmosis infection. Seizure 16(8), 657–663. https://doi.org/10.1016/j.seizure.2007.05.010
Papouin T, Dunphy JM, Tolman M, Dineley KT, Haydon PG (2017) Septal cholinergic neuromodulation tunes the astrocyte-dependent gating of hippocampal NMDA receptors to wakefulness. Neuron 94(4), 840–854. e847. https://doi.org/10.1016/j.neuron.2017.04.021
Parlog A, Harsan LA, Zagrebelsky M, Weller M, von Elverfeldt D, Mawrin C, Korte M, Dunay IR (2014) Chronic murine toxoplasmosis is defined by subtle changes in neuronal connectivity. Dis Models Mech 7(4):459–469. https://doi.org/10.1242/dmm.014183
Parlog A, Schluter D, Duna IR (2015) Toxoplasma gondii-induced neuronal alterations. Parasite Immunol 37(3):159–170. https://doi.org/10.1111/pim.12157
Patri M (2019) Synaptic transmission and amino acid neurotransmitters. Neurochemical Basis of Brain Function and Dysfunction. IntechOpen.
Paul V, Ekambaram P (2011) Involvement of nitric oxide in learning & memory processes. Indian J Med Res 133:471–478
Petitto JM, McCarthy DB, Rinker CM, Huang Z, Getty T (1997) Modulation of behavioral and neurochemical measures of forebrain dopamine function in mice by species-specific interleukin-2. J Neuroimmunol 73(1–2):183–190. https://doi.org/10.1016/s0165-5728(96)00196-8
Pfefferkorn ER (1984) Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proc Natl Acad Sci USA 81(3):908–912. https://doi.org/10.1073/pnas.81.3.908
Plattner F, Yarovinsky F, Romero S, Didry D, Carlier MF, Sher A, Soldati-Favre D (2008) Toxoplasma profilin is essential for host cell invasion and TLR11-dependent induction of an interleukin-12 response. Cell Host Microbe 3:77–87. https://doi.org/10.1016/j.chom.2008.01.001
Pliszka SR, Maas JW, Javors MA, Rogeness GA, Baker BS, J, (1994) Urinary catecholamines in attention-deficit hyperactivity disorder with and without comorbid anxiety. J Am Acad Child Adolesc Psychiatry 33(8):1165–1173. https://doi.org/10.1097/00004583-199410000-00012
Prandovszky E, Gaskell E, Martin H, Dubey JP, Webster JP, McConkey GA (2011) The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS ONE 6(9):e23866. https://doi.org/10.1371/journal.pone.0023866
Prast H, Philippu A (2001) Nitric oxide as modulator of neuronal function. Prog Neurobiol 64(1):51–68. https://doi.org/10.1016/S0301-0082(00)00044-7
Remenyi J, van den Bosch MW, Palygin O, Mistry RB, McKenzie C, Macdonald A, Hutvagner G, Arthur JS, Frenguelli BG, Pankratov Y (2013) miR-132/212 knockout mice reveal roles for these miRNAs in regulating cortical synaptic transmission and plasticity. PLoS ONE 8(4):e62509. https://doi.org/10.1371/journal.pone.0062509
Rommelfanger K, Edwards G, Freeman K, Liles L, Miller G, Weinshenker D (2007) Norepinephrine loss produces more profound motor deficits than MPTP treatment in mice. Proc Natl Acad Sci 104(34):13804–13809. https://doi.org/10.1073/pnas.0702753104
Rothstein JD, Patel S, Regan MR, Haenggeli C, Huang YH, Bergles DE, Jin L, Hoberg MD, Vidensky S, Chung DS (2005) β-Lactam antibiotics offer neuroprotection by increasing glutamate transporter expression. Nature 433(7021):73–77. https://doi.org/10.1038/nature03180
Rozenfeld C, Martinez R, Seabra S, Sant’anna C, Goncalves JG, Bozza M, Moura-Neto V, SouzaW De (2005) Toxoplasma gondii prevents neuron degeneration by interferon-gamma-activated microglia in a mechanism involving inhibition of inducible nitric oxide synthase and transforming growth factor-beta1 production by infected microglia. Am J Pathol 167(4):1021–1031. https://doi.org/10.1016/s0002-9440(10)61191-1
Sa Q, Tiwari A, Ochiai E, Mullins J, Suzuki Y (2018) Inducible nitric oxide synthase in innate immune cells is important for restricting cyst formation of Toxoplasma gondii in the brain but not required for the protective immune process to remove the cysts. Microbes Infect 20(4):261–266. https://doi.org/10.1016/j.micinf.2017.12.004
Sadeghi M, Riahi SM, Mohammadi M, Saber V, Aghamolaie S, Moghaddam SA, Aghaei S, Javanian M, Gamble HR, Rostami A (2019) An updated meta-analysis of the association between Toxoplasma gondii infection and risk of epilepsy. Trans R Soc Trop Med Hyg 113(8):453–462. https://doi.org/10.1093/trstmh/trz025
Saito K, Markey SP, Heyes MP (1992) Effects of immune activation on quinolinic acid and neuroactive kynurenines in the mouse. Neuroscience 51(1):25–39. https://doi.org/10.1016/0306-4522(92)90467-g
Sanecka A, Frickel EM (2012) Use and abuse of dendritic cells by Toxoplasma gondii. Virulence 3:678–689. https://doi.org/10.4161/viru.22833
Scanzano A, Cosentino M (2015) Adrenergic regulation of innate immunity: a review. Front Pharmacol 6:171. https://doi.org/10.3389/fphar.2015.00171
Scharton-Kersten TM, Yap G, Magram J, Sher A (1997) Inducible nitric oxide is essential for host control of persistent but not acute infection with the intracellular pathogen Toxoplasma gondii. J Exp Med 185(7):1261–1274. https://doi.org/10.1084/jem.185.7.1261
Schwarcz R, Hunter CA (2007) Toxoplasma gondii and schizophrenia: linkage through astrocyte-derived kynurenic acid? Schizophr Bull 33(3):652–653. https://doi.org/10.1093/schbul/sbm030
Schwarcz R, Pellicciari R (2002) Manipulation of brain kynurenines: glial targets, neuronal effects, and clinical opportunities. J Pharmacol Exp Ther 303(1):1–10. https://doi.org/10.1124/jpet.102.034439
Schwarcz R, Rassoulpour A, Wu H-Q, Medoff D, Tamminga CA, Roberts RC (2001) Increased cortical kynurenate content in schizophrenia. Biol Psychiatry 50(7):521–530. https://doi.org/10.1016/s0006-3223(01)01078-2
Schwarcz R, Bruno JP, Muchowski PJ, Wu HQ (2012) Kynurenines in the mammalian brain: when physiology meets pathology. Nat Rev Neurosci 13(7):465–477. https://doi.org/10.1038/nrn3257
Shen J (2013) Modeling the glutamate–glutamine neurotransmitter cycle. Front Neuroenergetics 5:1–13. https://doi.org/10.3389/fnene.2013.00001
Sher A, Tosh K, Jankovic D (2017) Innate recognition of Toxoplasma gondii in humans involves a mechanism distinct from that utilized by rodents. Cell Mol Immunol 14:36–42. https://doi.org/10.1038/cmi.2016.12
Sjoberg RL, Nilsson KW, Nordquist N, Ohrvik J, Leppert J, Lindstrom L, Oreland L (2006) Development of depression: sex and the interaction between environment and a promoter polymorphism of the serotonin transporter gene. Int J Neuropsychopharmacol 9(4):443–449. https://doi.org/10.1017/s1461145705005936
Skallova A, Kodym P, Frynta D, Flegr J (2006) The role of dopamine in Toxoplasma-induced behavioural alterations in mice: an ethological and ethopharmacological study. Parasitology 133(Pt 5):525–535. https://doi.org/10.1017/s0031182006000886
Sperner-Unterweger B, Kohl C, Fuchs D (2014) Immune changes and neurotransmitters: possible interactions in depression? Prog Neuropsychopharmacol Biol Psychiatry 48:268–276. https://doi.org/10.1016/j.pnpbp.2012.10.006
Stein V, Nicoll RA (2003) GABA generates excitement. Neuron 37(3):375–378
Steiner J, Walter M, Gos T, Guillemin GJ, Bernstein HG, Sarnyai Z, Mawrin C, Brisch R, Bielau H, Meyer zu Schwabedissen L, Bogerts B, Myint AM, (2011) Severe depression is associated with increased microglial quinolinic acid in subregions of the anterior cingulate gyrus: evidence for an immune-modulated glutamatergic neurotransmission? Neuroinflammation 8:94. https://doi.org/10.1186/1742-2094-8-94
Steinert JR, Chernova T, Forsythe ID (2010) Nitric oxide signaling in brain function, dysfunction, and dementia. Neuroscientist 16(4):435–452. https://doi.org/10.1177/1073858410366481
Stibbs HH (1985) Changes in brain concentrations of catecholamines and indoleamines in Toxoplasma gondii infected mice. Ann Trop Med Parasitol 79(2):153–157. https://doi.org/10.1080/00034983.1985.11811902
Strobl JS, Goodwin DG, Rzigalinski BA, Lindsay DS (2012) Dopamine stimulates propagation of Toxoplasma gondii tachyzoites in human fibroblast and primary neonatal rat astrocyte cell cultures. J Parasitol 98(6):1296–1299. https://doi.org/10.1645/ge-2760.1
Sugden K, Moffitt TE, Pinto L, Poulton R, Williams BS, Caspi A (2016) Is Toxoplasma gondii infection related to brain and behavior impairments in humans? Evidence from a population-representative birth cohort. PLoS ONE 11(2):e0148435. https://doi.org/10.1371/journal.pone.0148435
Sullivan JL, Cavenar J, Maltbie A, Lister P, Zung W (1979) Familial biochemical and clinical correlates of alcoholics with low platelet monoamine oxidase activity. Biol Psychiatry 14:385–394
Sundström E, Kölare S, Souverbic F, Samuelsson E-B, Pschera H, Lunell N-O, Seiger Å (1993) Neurochemical differentiation of human bulbospinal monoaminergic neurons during the first trimester. Dev Brain Res 75(1):1–12
Sutterland AL, Kuin A, Kuiper B, van Gool T, Leboyer M, Fond G, de Haan L (2019) Driving us mad: the association of Toxoplasma gondii with suicide attempts and traffic accidents - a systematic review and meta-analysis. Psychol Med 49(10):1608–1623. https://doi.org/10.1017/s0033291719000813
Sutterland AL, Fond G, Kuin A, Koeter MW, Lutter R, van Gool T, Yolken R, Szoke A, Leboyer M, de Haan L (2015) Beyond the association. Toxoplasma gondii in schizophrenia, bipolar disorder, and addiction: systematic review and meta-analysis. Acta Psychiatr Scand 132(3), 161–179. https://doi.org/10.1111/acps.12423
Suzuki Y (2002) Immunopathogenesis of cerebral toxoplasmosis. J Infect Dis 186(Suppl 2):S234-240. https://doi.org/10.1086/344276
Suzuki Y, Sa Q, Gehman M, Ochiai E (2011) Interferon-gamma-and perforin-mediated immune responses for resistance against Toxoplasma gondii in the brain. Expert Rev Mol Med 13:e31. https://doi.org/10.1086/344276
Swift RG, Polymeropoulos MH, Torres R, Swift M (1998) Predisposition of Wolfram syndrome heterozygotes to psychiatric illness. Mol Psychiatry 3:86–91
Tan L, Yu JT, Tan L (2012) The kynurenine pathway in neurodegenerative diseases: mechanistic and therapeutic considerations. J Neurol Sci 323(1–2), 1–8. https://doi.org/10.1016/j.jns.2012.08.005
Taupin P (2007). The hippocampus: neurotransmission and plasticity in the nervous system. Nova Publishers.
Tedford E, McConkey G (2017) Neurophysiological changes induced by chronic Toxoplasma gondii infection. Pathogens 6(2). https://doi.org/10.3390/pathogens6020019
Tenter AM, Heckeroth AR, Weiss LM (2000) Toxoplasma gondii: from animals to humans. J Int J Parasitol 30(12–13):1217–1258. https://doi.org/10.1016/S0020-7519(00)00124-7
Tipton KF, Dostert P, Strolin Benedetti M (1984) Monoamine oxidase and disease. Academic, London
Tiwari P, Dwivedi S, Singh MP, Mishra R, Chandy A (2013) Basic and modern concepts on cholinergic receptor: a review. Asian Pac J Trop Dis 3(5):413–420. https://doi.org/10.1016/S2222-1808(13)60094-8
Tonin AA, da Silva AS, Thorstenberg ML, Castilhos LG, França RT, Leal DBR, Duarte MMMF, Vogel FSF, de La Rue ML, dos Anjos Lopes ST (2013) Influence of Toxoplasma gondii acute infection on cholinesterase activities of Wistar rats. Korean J Parasitol 51(4):421. https://doi.org/10.3347/kjp.2013.51.4.421
Tonin AA, Da Silva AS, Thomé GR, Sangoi MB, Oliveira LS, Flores MM, Schetinger MRC, Fighera RA, Moresco RN, Camillo G (2014) Influence of toxoplasmosis on acetylcholinesterase activity, nitric oxide levels and cellular lesion on the brain of mice. Pathol Res Pract 210(8):526–532. https://doi.org/10.1016/j.prp.2014.04.025
Torrey EF, Yolken RH (2003) Toxoplasma gondii and schizophrenia. Emerging Infect Dis 9(11):1375. https://doi.org/10.3201/eid0911.030143
Torrey EF, Bartko JJ, Lun ZR, Yolken RH (2007) Antibodies to Toxoplasma gondii in patients with schizophrenia: a meta-analysis. Schizophr Bull 33(3):729–736. https://doi.org/10.1093/schbul/sbl050
Treiman DM (2001) GABAergic mechanisms in epilepsy. Epilepsia 42:8–12. https://doi.org/10.1046/j.1528-1157.2001.042suppl.3008.x
Tyebji S, Seizova S, Hannan AJ, Tonkin CJ (2019) Toxoplasmosis: a pathway to neuropsychiatric disorders. Neurosci Biobehav Rev 96:72–92. https://doi.org/10.1016/j.neubiorev.2018.11.012
van Staden JF, van Staden RI (2012) Flow-injection analysis systems with different detection devices and other related techniques for the in vitro and in vivo determination of dopamine as neurotransmitter. A Review Talanta 102:34–43. https://doi.org/10.1016/j.talanta.2012.05.017
Vyas A (2015) Mechanisms of host behavioral change in Toxoplasma gondii rodent association. PLoS Pathog 11(7):e1004935. https://doi.org/10.1371/journal.ppat.1004935
Wagner MJ, Kim TH, Savall J, Schnitzer MJ, Luo L (2017) Cerebellar granule cells encode the expectation of reward. Nature 544(7648):96–100. https://doi.org/10.1038/nature21726
Waly M, Olteanu H, Banerjee R, Choi SW, Mason JB, Parker BS, Sukumar S, Shim S, Sharma A, Benzecry JM, Power-Charnitsky VA, Deth RC (2004) Activation of methionine synthase by insulin-like growth factor-1 and dopamine: a target for neurodevelopmental toxins and thimerosal. Mol Psychiatry 9(4):358–370. https://doi.org/10.1038/sj.mp.4001476
Wanet A, Tacheny A, Arnould T, Renard P (2012) miR-212/132 expression and functions: within and beyond the neuronal compartment. Nucleic Acids Res 40(11):4742–4753. https://doi.org/10.1093/nar/gks151
Wang F, Lidow MS (1997) α2A-adrenergic receptors are expressed by diverse cell types in the fetal primate cerebral wall. J Comp Neurol 378(4):493–507
Wang Y-M, Xu F, Gainetdinov RR, Caron MG (1999) Genetic approaches to studying norepinephrine function: knockout of the mouse norepinephrine transporter gene. Biol Psychiatry 46(9):1124–1130. https://doi.org/10.1016/s0006-3223(99)00245-0
Wang HL, Wang GH, Li QY, Shu C, Jiang MS, Guo Y (2006) Prevalence of Toxoplasma infection in first-episode schizophrenia and comparison between Toxoplasma-seropositive and Toxoplasma-seronegative schizophrenia. Acta Psychiatr Scand 114(1):40–48. https://doi.org/10.1111/j.1600-0447.2006.00780.x
Wang T, Tang Z-h, Li J-f, Li X-n, Wang X, Zhao Z-j (2013) A potential association between Toxoplasma gondii infection and schizophrenia in mouse models. Exp Parasitol 135(3):497–502. https://doi.org/10.1016/j.exppara.2013.08.012
Wang ZT, Harmon S, O’Malley KL, Sibley LD (2015) Reassessment of the role of aromatic amino acid hydroxylases and the effect of infection by Toxoplasma gondii on host dopamine. Infect Immun 83(3):1039–1047. https://doi.org/10.1128/iai.02465-14
Wang ZT, Verma SK, Dubey JP, Sibley LD (2017) The aromatic amino acid hydroxylase genes AAH1 and AAH2 in Toxoplasma gondii contribute to transmission in the cat. PLoS Pathog 13(3):e1006272. https://doi.org/10.1371/journal.ppat.1006272
Wang T, Sun X, Qin W, Zhang X, Wu L, Li Y, Zhou C, Zhou H, He S, Cong H (2019) From inflammatory reactions to neurotransmitter changes: Implications for understanding the neurobehavioral changes in mice chronically infected with Toxoplasma gondii. Behav Brain Res 359:737–748. https://doi.org/10.1016/j.bbr.2018.09.011
Webster JP, Lamberton PH, Donnelly CA, Torrey EF (2006) Parasites as causative agents of human affective disorders? The impact of anti-psychotic, mood-stabilizer and anti-parasite medication on Toxoplasma gondii’s ability to alter host behaviour. Proc Biol Sci 273(1589):1023–1030. https://doi.org/10.1098/rspb.2005.3413
West AR, Galloway MP, Grace AA (2002) Regulation of striatal dopamine neurotransmission by nitric oxide: effector pathways and signaling mechanisms. Synapse 44(4):227–245. https://doi.org/10.1002/syn.10076
White K, Shih J, Fong T, Young H, Gelfand R, Boyd J, Simpson G, Sloane R (1980) Elevated platelet monoamine oxidase activity in patients with nonendogenous depression. Am J Psychiatry 137:1258–1259. https://doi.org/10.1176/ajp.137.10.1258
Wohlfert EA, Blader IJ, Wilson EH (2017) Brains and brawn: Toxoplasma infections of the central nervous system and skeletal muscle. Trends Parasitol 33(7):519–531. https://doi.org/10.1016/j.pt.2017.04.001
Wolfe N, Katz DI, Albert ML, Almozlino A, Durso R, Smith MC, Volicer L (1990) Neuropsychological profile linked to low dopamine: in Alzheimer’s disease, major depression, and Parkinson’s disease. J Neurol Neurosurg Psychiatry 53(10):915–917. https://doi.org/10.1136/jnnp.53.10.915
Wonodi I, Schwarcz R (2010) Cortical kynurenine pathway metabolism: a novel target for cognitive enhancement in schizophrenia. Schizophr Bull 36(2):211–218. https://doi.org/10.1093/schbul/sbq002
Wyatt RJ, Murphy DL (1976) Low platelet monoamine oxidase activity and schizophrenia. Schizophr Bull 2(1):77. https://doi.org/10.1093/schbul/2.1.77
Xiao J, Li Y, Prandovszky E, Karuppagounder SS, Talbot CC Jr, Dawson VL, Dawson TM, Yolken RH (2014) MicroRNA-132 dysregulation in Toxoplasma gondii infection has implications for dopamine signaling pathway. Neuroscience 268:128–138. https://doi.org/10.1016/j.neuroscience.2014.03.015
Xiao J, Li Y, Prandovszky E, Kannan G, Viscidi RP, Pletnikov MV, Yolken RH (2016) Behavioral abnormalities in a mouse model of chronic toxoplasmosis are associated with MAG1 antibody levels and cyst burden. PLOS Negl Trop Dis 10(4):e0004674. https://doi.org/10.1371/journal.pntd.0004674
Xu F, Gainetdinov RR, Wetsel WC, Jones SR, Bohn LM, Miller GW, Wang Y-M, Caron MG (2000) Mice lacking the norepinephrine transporter are supersensitive to psychostimulants. Nat Neurosci 3(5):465–471. https://doi.org/10.1038/74839
Xu M-J, Zhou D-H, Nisbet AJ, Huang S-Y, Fan Y-F, Zhu X-Q (2013) Characterization of mouse brain microRNAs after infection with cyst-forming Toxoplasma gondii. Parasites Vectors 6:154. https://doi.org/10.1186/1756-3305-6-154
Yang D, Li T, Wang Y, Tang Y, Cui H, Tang Y, Zhang X, Chen D, Shen N, Le W (2012) miR-132 regulates the differentiation of dopamine neurons by directly targeting Nurr1 expression. J Cell Sci 125(Pt 7):1673–1682. https://doi.org/10.1242/jcs.086421
Yap G, Pesin M, Sher A (2000) Cutting edge: IL-12 is required for the maintenance of IFN-gamma production in T cells mediating chronic resistance to the intracellular pathogen Toxoplasma Gondii. J Immunol 165(2):628–631. https://doi.org/10.4049/jimmunol.165.2.628
Yarovinsky F (2014) Innate immunity to Toxoplasma gondii infection. Nat Rev Immunol 14(2):109–121. https://doi.org/10.1038/nri3598
Yarovinsky F, Zhang D, Andersen JF, Bannenberg GL, Serhan CN, Hayden MS, Hieny S, Sutterwala FS, Flavell RA, Ghosh S, Sher A (2005) TLR11 activation of dendritic cells by a protozoan profilin-like protein. Science 308:1626–1629. https://doi.org/10.1126/science.1109893
Zalcman S, Green-Johnson JM, Murray L, Nance DM, Dyck D, Anisman H, Greenberg AH (1994) Cytokine-specific central monoamine alterations induced by interleukin-1, -2 and -6. Brain Res 643(1–2):40–49. https://doi.org/10.1016/0006-8993(94)90006-x
Zhang YH, Chen H, Chen Y, Wang L, Cai YH, Li M, Wen HQ, Du J, An R, Luo QL, Wang XL, Lun ZR, Xu YH, Shen JL (2014) Activated microglia contribute to neuronal apoptosis in toxoplasmic encephalitis. Parasit Vectors 7:372. https://doi.org/10.1186/1756-3305-7-372
Zhang Y, Tan F, Xu P, Qu S (2016) Recent advance in the relationship between excitatory amino acid transporters and Parkinson’s disease. Neural Plast 2016.https://doi.org/10.1155/2016/8941327
Zhu CB, Carneiro AM, Dostmann WR, Hewlett WA, Blakely RD (2005) p38 MAPK activation elevates serotonin transport activity via a trafficking-independent, protein phosphatase 2A-dependent process. J Biol Chem 280(16):15649–15658. https://doi.org/10.1074/jbc.M410858200
Zhu CB, Lindler KM, Owens AW, Daws LC, Blakely RD, Hewlett WA (2010) Interleukin-1 receptor activation by systemic lipopolysaccharide induces behavioral despair linked to MAPK regulation of CNS serotonin transporters. Neuropsychopharmacology 35(13):2510–2520. https://doi.org/10.1038/npp.2010.116
Zimmerman AW, Jyonouchi H, Comi AM, Connors SL, Milstien S, Varsou A, Heyes MP (2005) Cerebrospinal fluid and serum markers of inflammation in autism. Pediatr Neurol 33(3):195–201. https://doi.org/10.1016/j.pediatrneurol.2005.03.014
Feigin VLAA, Abate KH, Abd-Allah F, Abdulle AM, Abera SF, Abyu GY, Vos T (2017) Global, regional, and national burden of neurological disorders during 1990–2015: a systematic analysis for the global burden of disease study 2015. Lancet Neurol 16(11):877–97. https://doi.org/10.1016/s1474-4422(17)30299-5
Acknowledgements
This work is the approved plan (No. 7378) from the Student Research Committee of Mazandaran University of Medical Sciences, Sari, Iran.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
The code of ethics of this plan is (IR. MAZUMS.REC.1399.386).
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Nayeri, T., Sarvi, S. & Daryani, A. Toxoplasmosis: Targeting neurotransmitter systems in psychiatric disorders. Metab Brain Dis 37, 123–146 (2022). https://doi.org/10.1007/s11011-021-00824-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11011-021-00824-2