
Abstract
Antimicrobial resistance in pathogenic bacteria is increasing rapidly. Therefore, investigation of novel therapeutic agents are essential to combat the growing multidrug resistance (MDR) in human pathogens. The current study was intended to identify antibacterial metabolites producing Bacillus spp. from the unexplored southern region of Pakistan. The bacterial isolates (n = 47) were isolated from soil samples and screened for antibacterial activity against a set of American Type Culture Collection (ATCC) and MDR human pathogenic bacterial strains. Among all isolated Bacillus spp. the strain MK-12.1 exhibt promising antibacterial activities agains all tested strains. The antibacterial metabolites from MK-12.1 were partially extracted and their molecular weight and toxicity were evaluated. Genomic DNA was extracted from a fresh culture of antibacterial metabolite-producing strain MK-12.1, and the 16S rRNA gene was amplified and sequenced. The growth and antibacterial metabolites production by MK-12.1 was optimized at various pH, temperature, incubation time, and medium. The strain MK-12.1 efficiently inhibit the growth of ATCC as well as MDR pathogenic bacterial strains with the zone of inhibition ranges from 14 to 18 mm. The partially extracted antibacterial compound was characterized and confirmed to be proteinogenic, non-hemolytic, and molecular weight was less than 20 kDa. The isolate MK-12.1 was identified as Bacillus safensis and revealed 99% similarity with B. safensis strain NBRC. Maximum antagonistic activity and growth of MK-12.1 were observed at pH 8, 30 °C temperature, and 48 h of incubation in a shaking incubator when fermented in an optimized (opt) medium. The current study results indicate that B. safensis MK-12.1 produces promising antibacterial compounds that could be a source of new antibacterial leads to fight against MDR bacteria in future.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Background
Before the discovery of antibiotics, individuals with bacteremia and septicemia had low chances of survival. However, the availability of antibacterial drugs and the synthesis of new chemotherapy agents upturned this scenario. Recently, the rise of multi-drug resistance (MDR) in human pathogens, challenging medical practitioners and researchers to develop or discover new antimicrobial agents to combat these organisms (Vivas et al. 2019). Urinary tract infections (UTIs) are prominent infections that occur worldwide, especially in developing countries including Pakistan, where the chances of drug-resistant are much higher due to injudicious use of antibiotics. The majority of UTIs are caused by Staphylococcus aureus, Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa. Unfortunately, these bacteria have been proven as Multidrug resistance (MDR) thus making its treatment ineffective (Shah et al. 2015). Further, the pan-drug resistance in pathogenic bacteria has turned into more serious distress to the community, gaining urgent focus of research to find a more effective chemotherapeutic agent to combat these pathogenic bacteria. Despite the salient need, the progress of developing new antibiotics is slow and the increase of newly approved antibiotics has steadily decreased in the last few decades (Vivas et al. 2019). Therefore, to treat the infections, especially in hospitals where resistance immediately may become life-threatening, the discovery of new antibacterial compounds is becoming more urgent (Dutescu and Hillie 2021). Previously several antibiotics such as penicillin, vancomycin, and tetracycline, have been discovered from environmental microorganisms. Among bacteria Actinomycetes and Bacillus spp. are known to produce a plethora of secondary metabolites that have potent antibacterial activities (Yahya et al. 2021; Selim et al. 2021). In terms of bioactivity, Actinomycetes have been extensively studied as compared to Bacillus. Bacilli are ubiquitous, endospore-forming, rod-shaped Gram-positive bacteria that are of great importance due to their ability to produce a broad spectrum of antibacterial metabolites. These bacteria are extensively used in industrial processes for different enzyme production like proteases and also used as plant growth-promoting bacteria. They produced diverse bioactive metabolites that may be used to control various pathogens (Abdelghani et al. 2021).
The current study was therefore conducted to explore the indigenous soil bacteria with broad spectrum antibacterial metabolites that could be used to combat MDR pathogens causing nosocomial infections.
Methods
Soil Sampling and Bacterial Isolates Cultivation
Soil samples (n = 9) were collected aseptically from various regions of district Kohat (33.5638o N, 71.4656o E), Pakistan at the depth of 3–8 cm. The samples were serially diluted, 200 µl from each dilution were spread over Tryptic Soy Agar (TSA) plate and incubated at 37 °C for 48 h. A single bacterial colony was obtained by repeated sub-culturing on TSA (Oxide, UK). Fresh pure bacterial cultures were deep-frozen in 20% glycerol and maintained at -80 °C until required.
Screening for Antibacterial Metabolites Production
The purified bacterial isolates (n = 47) were preliminarily examined for antibacterial metabolites production against indicator strains by cross-streaking method (Williston et al. 1947). Subsequently, the antagonistic activity of the Cell-Free Supernatant (CFS) from MK-12.1 culture was determined against a set of indicator ATCC (Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus pneumonia, Salmonella typhimurium, Shigella flexneri, Klebsiella pneumoniae, Vibrio cholerae, Pseudomonas aeruginosa, and Escherichia coli) (Table S1) as well as clinical MDR bacterial strains (Acinetobacter baumannii, Staphylococcus aureus, Escherichia coli, and Pseudomonas aeruginosa) by agar well diffusion assay on Muller Hinton Agar (MHA) plate (Iqbal et al. 2021b). The Petri plates were maintained at 37 °C for 24 h and Zone Of Inhibition (ZOI) was measured in mm. The MDR bacterial indicator strains were previously collected (Ullah et al. 2016) from local tertiary care hospitals and the resistant pattern was determined according to Clinical and Laboratory Standards Institute (CLSI) 2015 protocols.
Optimization of Growth and Antibacterial Activity of Selected Strains
Various culture conditions including temperature, culture medium, aeration, pH, and incubation time were optimized for antibacterial compounds production and growth of the strain MK-12.1. In order to assess the impact of pH and temperature, 100 ml TSB and 1 ml standardize inoculum of strain MK-12.1 was poured in Erlenmeyer flasks. The pH of flasks containing 100 ml media and 1 ml inoculum were adjusted as 6, 7, 8, 9 and incubated at various temperature, i.e., 25, 30, 35, 44 °C for 48 h. For culturing medium optimization the standard inoculum was added to sterile media, i.e., Luria–Bertani (LB) broth, Brain Heart Infusion (BHI) broth, Nutrient broth (NB), Tryptic soy broth (TSB), and Optimized (OPT) medium with a few modifications (Table S2). To evaluate the impact of incubation time on bioactive metabolites production, the inhibitory activity of CFS was determined at different time intervals (every 24 h, for 96 h) against test bacterial strains. The influence of aeration on biomass and antibacterial metabolites synthesis was studied by culturing the producer isolate in shaker culture (80, 120, 160, 200 rpm) and static culture for 48 h. The antagonistic potential was determined by agar well diffusion assay and bacterial biomass was estimated by UV–vis spectrophotometer (600 nm). For each experiment, 5 ml fresh standardized (1 McFarland) culture of strain 12.1was used as the initial inoculum.
Antibacterial Metabolites Production and Partial Purification
Antibacterial exhibiting strain MK-12.1 overnight fresh culture was inoculated in Erlenmeyer flasks containing 500 ml modified OPT medium, maintained at 30 °C for 48 h as shaking culture. Subsequently, the culture of MK-12.1 was spun at 6000 rpm for 15 min. Further, the supernatant was passed via a 0.45 µM pore size membrane to remove all the cell debris. The supernatant was neutralized using 1 M NaOH and 10% HCl. The bioactive compounds were partially extracted from the supernatant using ethyl acetate (1:1). and dried in a rotary evaporator under reduced pressure (Iqbal et al., 2021a, b). Vancomycin (30 µg) and ethyl acetate were used as positive and negative control, respectively.
Determination of Minimum Inhibitory Concentration (MIC)
The MIC of partially purified metabolites from strain MK-12.1 was determined against indicator strains using a two-fold serial dilution technique (Kowalska-Krochmal and Dudek-Wicher 2021). The dried antibacterial metabolites extract was dissolved in Dimethyl sulfoxide (DMSO) at 100 mg/ml concentration. Initially, 1 ml of extract (100 mg/ml) was poured into tube no. 1 and serially diluted all test tubes (1–10) containing 1 ml of TSB. Then, 10 µl of test bacterial strain culture (final concentration equal to 0.5 McFarland) was dispensed into all tubes (1–12). The second last tube (11) containing DMSO and last tube (12) containing antibiotic streptomycin (100 µg) was used as a negative and positive control, respectively. In order to determine the MIC of positive control (streptomycin), 100 µg was dissolved in 1 ml sterile distilled water, and the same two fold dilution method was performed. The lowest amount of partially extracted antibacterial compounds that entirely restrict the proliferation of indicator strains were defined as MICs.
Characterization of Antibacterial Metabolites
To estimate the molecular weight of antibacterial metabolites, the partially purified antibacterial metabolites were dispersed on a dialysis membrane having a pore size < 20 Kilo Dalton (kDa). All the setup was kept on an agitator for 18 h at 25 °C and subsequently, antibacterial activity was determined. To confirm the nature of antibacterial metabolites, the partially purified metabolites were mixed with proteinase K, and later on, antagonistic activity was assessed. The temperature stability of antibacterial metabolites was evaluated by heating the partially purified metabolites from 20 to 121 °C for 20 min and antagonistic activity was evaluated against indicator strains. Hemolytic activity was determined on a blood agar plate. The antibacterial metabolites (100 µg/ml) were aseptically dispensed into the pre-made agar well and the plates were maintained at 37 °C for 24 h.
Identification of Soil Isolate MK-12.1
Initially, the antibacterial metabolites producing isolate was tentatively identified by colony morphology, Gram reaction, and biochemical testing as per Bergey's manual of determinative bacteriology. Subsequently, the soil isolate MK-12.1 was molecularly identified based on 16S rRNA gene sequencing. Total genomic DNA was extracted using phenol:chloroform method with a little modification (Wright et al. 2017) and the aforementioned gene was amplified by 9F (GAGTTTGATCCTGGCTCAG) and 1510R (GGCTACCTTGTTACGA) as forward and reverse primers. In PCR initial denaturation was performed at 94 °C for 120 s., annealing temperature was 52 °C for 90 s. and extension time was 2 min at 72 °C, and then the reaction was repeated 35 times, then a final extension was performed at 72 °C for 10 min. The amplified PCR product was separated on 0.9% agarose gel. The amplified product was recovered and purified using PCR purification kit (Roche, Germany)and sequenced using the di-deoxy chain termination method (Macrogen, Korea).The obtain sequence was BLASTn in NCBI (https://www.ncbi.nlm.nich.gov/BLAST/) and Eztaxon (https://www.ezbiocloud.net/identify) databases. The 16S rRNA gene of the related strains were downloaded from Ezbioloud. Multiple sequence alignments were performed using Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/). The phylogenetic tree was constructed based on the neighbor-joining method using a computational tool MEGA-X (Kumar et al. 2018). The reliability of the phylogenetic trees was determined using the bootstrap technique on 1000 replicates.
Results
Soil Sampling and Plate Count
The soil samples were collected from various localities including hilly areas, lawns, gardens, industrial areas, open cultivated fields, riverbanks, stream banks, rhizosphere, and ponds surrounding area. Plate count on TSA ranged from 4 × 105 to 7 × 105 CFU/g of soil samples. A minimum count of 4 × 105 CFU/g was obtained for the mill effluent soil sample, Kohat.
Antibacterial Activity
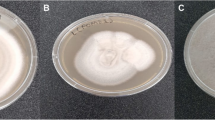
Among all 47 soil isolates, 12 isolates (25.53%) exhibited antagonistic activities either against Gram-positive bacterial strains or Gram-negative bacterial strains or both (Figure S1). Among these, three (25%) isolates designated as MK-9, MK-10 and MK-32, showed antagonistic activity only against Gram-positive test bacterial strains while nine isolates (75%), exhibited antagonistic activity against both Gram-positive as well as Gram-negative bacterial strains (data not shown here). One isolate B. safensis MK-12.1 exhibited promising antibacterial activity against all test strains (Fig. 1).

Antagonistic activities of B. safensis strain MK-12.1 against indicator strains
The partial extract of B. safensis MK-12.1 efficiently inhibits the growth of S. typhimurium (ZOI = 18 + 0.5 mm) as compared to other test strains such as K. pneumoniae (ZOI = 15 mm).
Optimization of Growth and Antibacterial Metabolites
Maximum antibacterial activity of CFS was achieved at 30 °C with 18 mm ZOI followed by 35 °C with 17 mm ZOI. On the other hand, lower antagonistic activity was observed at 25 and 40 °C, indicating that the soil isolate B. safensis MK-12.1 produces the highest amount of antibacterial metabolites at 30 °C (Fig. 2). The current study result revealed that nitrogen-rich media, i.e., BHI and TSB enhanced the growth and production of antibacterial metabolites production as compared to a simple nutrient broth medium. The CFS collected from OPT medium exhibited 19 mm ZOI, followed by BHI and TSB medium which exhibited 18 mm ZOI each against indicator strain. On the other hand, a high amount of biomass was obtained by cultivating the strain MK-12.1 in OPT medium followed by TSB and BHI medium, indicating that nutrients have a considerable effect on antibacterial metabolites production (Fig. 3). The BHI and TSB culture media are rich in carbon and nitrogen sources as compared to the nutrient broth as previously demonstrated earlier (Morais and Suárez 2016).

Optimization of incubation time and temperature for growth and bioactive compound production by B.safensis MK-12.1

Culturing medium optimization for B. safensis MK-12.1 growth and bioactive compounds production. Trypticase soy broth: TSB, Nutrients broth: NB, Luria Bertani LB, Brain heart infusion: BHI and Optimized: OPT
The effect of pH on antibacterial metabolite production was evaluated by cultivating the strain 12.1 at various pH ranges from 6–9. Maximum ZOI against indicator strains was obtained at pH 8, while lower ZOI was recorded when the strain was cultured at 6, 7, and 9 pH. These results indicates that pH 8 favors the synthesis of antibacterial metabolite. Similarly, maximum growth was also observed at pH 8 after 48 h of incubation (Fig. 4).

Optimization of pH for growth and bioactive compounds production by B.safensis MK-12.1
Partially Purification and Characterization of Bioactive Compounds
The partially purified bioactive compounds exhibited increased antibacterial activity against the test organism as compared to CFS. The molecular weight of antibacterial metabolites was estimated as > 20 kDa. The antibacterial metabolites were treated with proteinase K lost their antibacterial activity validating that it is a protein in nature. When the CFS was heated from 20–121 °C for 20 min, the antagonistic activity against test strains decreased at 45–80 °C and denatured when heated up to 100 °C. The sensitivity of antibacterial metabolites to proteinase K and high temperature confirm that it is protein in nature. Hemolytic activity of the partially purified extract was evaluated and no activity was observed against human erythrocytes.
Determination of MIC
The MIC of partially purified bioactive compounds against ATCC bacterial strains ranged from 2.35–4.66 mg/ml. While against local MDR bacterial strains the MIC ranged from 3.12 to 6.25 mg/ml as shown in Table 1.
Morphological, Biochemical, and Molecular Identification of Strain MK-12.1
The colony morphology of B.safensis MK-12.1 was irregular in shape, with an undulant margin, raised elevation, light yellow in color, and opaque opacity. The phenotypic analysis showed that the strain MK-12.1 is Gram-positive, rod-shaped, motile, and spore-forming bacilli. Biochemical results indicate that B. safensis MK-12.1 is positive for ureas, catalase, oxidase production, and MR test while negative for indole, citrate, and VP test. The 16S rRNA gene of strain 12.1 was successfully amplified and > 1200 bp amplified product was obtained (Figure S2).The 16S rRNA gene sequence of strain MK-12.1 has been submitted to GenBank (NCBI) database under the accession number MN519460. The 16S rRNA gene sequence BLASTn analysis with other related strains showed maximum similarity (99.73%) with B.safensis strain NBRC 100,820. The phylogenetic tree indicates the close relation of strain MK-12.1 with other B. safensis strains and shared a clade with B. safensis NBRC and B. safensis FO-36b (Fig. 5).

Evolutionary analysis of B. safensis strain MK-12.1 with related Bacillus species. Phylogenetic tree was generated based 16S rRNA with neighbor-joining method using MEGA X
Discussion
The prevalence of MDR bacteria has increased at an alarming rate in the past decades. Therefore, it is an urgent need to investigate new drugs to combat these MDR pathogenic bacteria. In the present study, environmental samples were collected from the unexplored region in terms of bioactive metabolites producing bacteria. A total of 12 (25.53%) out of 47 bacterial isolates exhibited antibacterial activity in primary screening. This result indicates a relatively low number of antibacterial strains previously reported (Mutaz Al-Ajlani and Hasnain 2010). Previously CFS of L. plantarum was evaluated against pathogenic E. coli and 26 mm ZOI was recorded (Kaewchomphunuch et al. 2022). Recently, CFS was collected from Lactobacillus paracasei strain CH88 efficiently inhibits the growth of Gardnerella vaginalis (Moon et al. 2022). The results of the current study are fascinating as the bioactive compounds synthesized by strain MK-12.1 exhibited promising antibacterial activity against both ATCC and MDR clinical pathogens. According to the current study result, vancomycin showed 24 mm ZOI against MDR clinical strains which is higher than partially purified metabolites from the soil isolates. However, it is expected that further complete purification of the antibacterial metabolites might be comparable to vancomycin. The current study result corroborates with the earlier, where B. subtilis strain RS10 was isolated from soil and exhibit potent antimicrobial activity againt ATCC indicator strains including S. aureus (14 mm ZOI) (Iqbal et al. 2021a). Optimization for biomass and bioactive compound synthesis revealed that CFS of MK-12.1 culture exhibited maximum antibacterial with 18.6 mm ZOI at 30 °C followed by 37 °C with 18 mm of ZOI. While lowest inhibitory activity was noted at lower and higher temperature, which indicate that the optimum temperature for B.safensis MK-12.1 is 30 °C and temperature fluctuations play a major role in bacterial growth and production of metabolites. These results match with the other study in which Lactobacillus spp. yield highest amount of bacitracin at 25 °C and 30 °C (Malheiros et al. 2015). Temperature significantly affects the antibacterial metabolite production and may inactivate it to some level. In the earlier investigation, Bacillus sp. was found to produce antimicrobial compounds that are stable at high temperature such as 75 °C. A slight loss of activity (20%) was observed at 80 °C (Chalasani et al. 2015). As compared to general-purpose media such as nutrient broth medium, the nitrogen-rich BHI and OPT media enhance the growth and bioactive compounds synthesis. The ZOI of CFS taken from OPT culture medium was higher (25 mm) than the nutrient broth (15 mm), demonstrating the significant role of nutrients in the production of antibacterial metabolites. The OPT, BHI and TSB medium is rich in terms of carbon and nitrogen source as compared to the nutrient broth medium as conferred in an aerlier study (Holzapfel 2002). The MDR Gram-positive strains such as S. aureus are often causing UTIs and nosocomial infections and confer resistant to antibiotics by modifying its teichoic acid in the cell wall with D-alanine as a result of increased MIC. The result of the current study is worth observing that the MIC values (mg/ml) against MDR strains did not increase and are comparable to other tested bacterial strains. In previous study, the MIC values against different indicator strains were determined and ranges from 0.3 to 16 μg/ml (for MRSA 5) in BHI and CA-MHB to 120 μg/ml (Chalasani et al. 2015). The antibacterial activity of extracted bioactive compounds from B. safensis MK-12.1 is impeded at high temperature and it did not exhibit hemolytic activity on the blood agar.
In conclusion, the current study results showed that the B. safensis strain 12.1 isolated from the southern region in Pakistan has potential for the discovery of novel antibacterial compounds. The strain 12.1 exhibits promising antibacterial activities against both ATCC as well as local clinical MDR strains. The partially extracted bioactive compounds were determined to be protein in nature and have a molecular size of less than 20 kDa. The broad spectrum of antibacterial activities are either due to the broad spectrum of bioactive compound production or, to several bioactive compounds with various potencies. Therefore, further analysis is required to purify and elucidate their structures using various analytical techniques.
-
Employment: None.
-
Personal financial interests: None declared
-
Stocks or shares in companies: No
-
Consultation fees: No
-
Patents: None
-
Personal or professional relations with organizations and individuals (parents and children, wife and husband, family relationships, etc.): No personal relation
-
Unpaid membership in a government or non-governmental organization: No
-
Are you one of the editorial board members or a reviewer of this journal: No
References
Abdelghani Z, Hourani N, Zaidan Z et al (2021) (2021) Therapeutic applications and biological activities of bacterial bioactive extracts. Arch Microbiol 2038(203):4755–4776. https://doi.org/10.1007/S00203-021-02505-1
Chalasani AG, Dhanarajan G, Nema S et al (2015) An antimicrobial metabolite from Bacillus sp.: significant activity against pathogenic bacteria including multidrug-resistant clinical strains. Front Microbiol 6:1335. https://doi.org/10.3389/FMICB.2015.01335/BIBTEX
Dutescu IA, Hillie SA (2021) Encouraging the development of new antibiotics: are financial incentives the right way forward? A systematic review and case study. Infect Drug Resist 14:415. https://doi.org/10.2147/IDR.S287792
Holzapfel W (2002) Bacteriocin production by Enterococcus faecium RZS C5 is cell density limited and occurs in the very early growth phase. Int J Food Microbiol
Iqbal S, Ullah N, Janjua HA (2021a) In vitro evaluation and genome mining of Bacillus subtilis strain RS10 reveals its biocontrol and plant growth-promoting potential. Agriculture 11:1273
Iqbal S, Vohra MS, Janjua HA (2021b) Whole-genome sequence and broad-spectrum antibacterial activity of Chryseobacterium cucumeris strain MW-6 isolated from the Arabian Sea. 3 Biotech. https://doi.org/10.1007/S13205-021-03039-5
Kaewchomphunuch T, Charoenpichitnunt T, Thongbaiyai V et al (2022) Cell-free culture supernatants of Lactobacillus spp. and Pediococcus spp. inhibit growth of pathogenic Escherichia coli isolated from pigs in Thailand. BMC Vet Res 18:1–13. https://doi.org/10.1186/S12917-022-03140-8/TABLES/7
Kowalska-Krochmal B, Dudek-Wicher R (2021) The minimum inhibitory concentration of antibiotics: methods, interpretation, clinical relevance. Pathogens 10:1–21. https://doi.org/10.3390/PATHOGENS10020165
Kumar S, Stecher G, Li M et al (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/MOLBEV/MSY096
Malheiros PS, Sant’Anna V, Todorov SD, Franco BDGM (2015) Optimization of growth and bacteriocin production by Lactobacillus sakei subsp. sakei2a. Braz J Microbiol 46:825–834. https://doi.org/10.1590/S1517-838246320140279
Moon EC, Park MS, Lim T et al (2022) (2022) Antibacterial effect of cell-free supernatant fraction from Lactobacillus paracasei CH88 against Gardnerella vaginalis. Sci Reports 121(12):1–10. https://doi.org/10.1038/s41598-022-08808-7
Morais V, Suárez N (2016) Economic Evaluation of Streptococcus Pneumoniae Culture Media. Am J Biochem Biotechnol 12:133–138. https://doi.org/10.3844/AJBBSP.2016.133.138
Mutaz Al-Ajlani M, Hasnain S (2010) Bacteria exhibiting antimicrobial activities; screening for antibiotics and the associated genetic studies. TOPROCJ 1:230–238
Selim MSM, Abdelhamid SA, Mohamed SS (2021) Secondary metabolites and biodiversity of actinomycetes. J Genet Eng Biotechnol. https://doi.org/10.1186/S43141-021-00156-9
Shah DA, Wasim S, Abdullah FE (2015) Antibiotic resistance pattern of Pseudomonas aeruginosa isolated from urine samples of urinary tract infections patients in Karachi, Pakistan. Pakistan J Med Sci 31:341–345. https://doi.org/10.12669/PJMS.312.6839
Ullah A, Qasim M, Rahman H et al (2016) High frequency of methicillin-resistant Staphylococcus aureus in Peshawar Region of Pakistan. Springerplus. https://doi.org/10.1186/S40064-016-2277-3
Vivas R, Barbosa AAT, Dolabela SS, Jain S (2019) Multidrug-resistant bacteria and alternative methods to control them: an overview. Microb Drug Resist 25:890–908. https://doi.org/10.1089/MDR.2018.0319
Williston EH, Zia-Walrath P, Youmans GP (1947) Plate methods for testing antibiotic activity of actinomycetes against virulent human type tubercle bacilli. J Bacteriol Res 54(5):563–568.
Wright MH, Adelskov J, Greene AC (2017) Bacterial DNA extraction using individual enzymes and phenol/chloroform separation. J Microbiol Biol Educ. https://doi.org/10.1128/JMBE.V18I2.1348
Yahya G, Ebada A, Khalaf EM et al (2021) Soil-associated bacillus species: a reservoir of bioactive compounds with potential therapeutic activity against human pathogens. Microorganisms. https://doi.org/10.3390/MICROORGANISMS9061131
Funding
The current study did not receive any funding.
Author information
Authors and Affiliations
Contributions
SI and MQ designed and SI performed the experiments. SI and FB wrote the manuscript. FB and MQ review and edits the manuscript. MQ supervised the study.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Iqbal, S., Begum, F. & Qasim, M. Screening, Characterization and Optimization of Bioactive Peptides with Antibacterial Activities Against Multi-Drug Resistant Pathogens, Produced by Bacillus safensis Strain MK-12.1. Int J Pept Res Ther 28, 161 (2022). https://doi.org/10.1007/s10989-022-10469-z
Accepted:
Published:
DOI: https://doi.org/10.1007/s10989-022-10469-z