
Abstract
Context
The Dry Chaco spans more than 87 million hectares across Argentina, Bolivia, and Paraguay. This unique forest system has experienced extensive loss and fragmentation due to land-use change, with different land-use histories in the three countries. This forest loss has altered landscape connectivity for the Dry Chaco’s associated biota.
Objectives
We compared patterns of deforestation-induced fragmentation and concomitant changes in structural landscape connectivity between 2000 and 2019 in the three countries to identify consistent patterns that might facilitate biome-wide conservation.
Methods
We quantified forest cover in the Dry Chaco of Argentina, Bolivia, and Paraguay for the years 2000 and 2019 at 30 m resolution. We analyzed structural connectivity at three scales. Then, we identified and visualized the most important stepping stones per country per year.
Results
Between 2000 and 2019, the overall extent of Dry Chaco forest cover decreased by 20.2% (9.5 million ha). All three counties experienced substantial reductions, with Paraguay undergoing the greatest loss and fragmentation relative to 2000. Most of the overall network metrics decreased from 2000 to 2019 for Paraguay and Bolivia, but Argentina experienced increased coalescence distance and average nodal connectance. Dispersal-level metrics showed clustering threshold distances between 1000 and 2000 m for each country in both years.
Conclusions
The large number of forest fragments and distances between them suggest that some mammals characteristic of the biome may be experiencing negative impacts from this fragmentation. Contemporary and future challenges of uncoordinated national conservation and management policies, land speculation, and increased human infrastructure will accelerate the rate of deforestation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Anthropogenic activities, particularly deforestation at the global scale, have profound effects on biodiversity (Foley et al. 2005). Over the past 25 years alone, global forest coverage has been reduced by over 129 million ha (FAO 2015), with > 25% of these losses due to land-cover changes associated with commodity production (Curtis et al. 2018). Of the ~ 4000 million ha of forest remaining, ~ 70% is within 1 km of non-forested edge, indicating extensive fragmentation (FAO 2015; Haddad et al. 2015). Rates of deforestation are spatially heterogeneous across the globe, so certain forests have been disproportionately affected (Hansen et al. 2013). Tropical and subtropical forests have been reduced by > 50%, to the point where only remnants remain (FAO 2015; Sloan and Slayer 2015; Brancalion et al. 2019). Among tropical and subtropical forests, dry forests are highly underappreciated despite their uniqueness and high species richness (DRYFLOR 2017). The most unique, most threatened, and yet largely overlooked dry forest system is the Dry Chaco of South America (Kuemmerle et al. 2017).
The Dry Chaco extends across Argentina (55.5 million ha; 63.5%), Paraguay (18.8 million ha; 21.5%), and Bolivia (13.1 million ha; 15%) (areas calculated from Olson et al. 2001) (Fig. 1) and has experienced extensive deforestation over the past few decades (TNC et al. 2005; Carranza et al. 2014; Caldas et al. 2015; Fehlenberg et al. 2017). Its biodiversity is high (Redford et al. 1990, TNC et al. 2005), is currently threatened by habitat loss and hunting (Romero-Muñoz et al. 2020a), and at the same time many of its species are still poorly understood (López–González 2005; de la Sancha and D’Elia 2015; de la Sancha et al. 2007, 2012, 2017; Teta et al. 2009, 2018). Forest loss and fragmentation are particularly concerning given the presence of endemic and near-endemic species (Prado 1993; Periago et al. 2015; Nori et al. 2016; Andrade-Díaz et al. 2019).

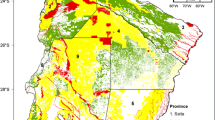
The historical boundary (gray) of the Dry Chaco of South America (A) extends into Argentina, Bolivia, and Paraguay, but (B) forest loss between 2000 and 2019 (yellow) primarily occurred in Argentina and Paraguay. Much of the remaining Dry Chaco forest in 2019 (green) consists of isolated forest remnants that are not within protected areas (pink polygons at 75% transparency). Areas within the historical boundary of the Dry Chaco that were not forested in 2000 are denoted in gray. Sources: Forest cover and loss data: Hansen et al. (2013); historical boundary of the Dry Chaco: Olson et al. (2001); and protected areas: UNEP-WCMC and IUCN (2020)
Historically, the main drivers for deforestation in the Dry Chaco were firewood and timber extraction and cattle ranching (Grau et al. 2008). This changed in the last decade to soy production for exportation (Cáceres 2015; Leguizamón 2016; Fehlenberg et al. 2017). Advance of industrial agriculture has increased road-building, further fragmenting areas, causing vehicle-collision mortalities of wildlife (Ascensão et al. 2019) and increasing human access to remote areas for hunting and poaching (Trombulak and Frissell 2000; McBride and Thompson 2019; Romero-Muñoz et al. 2020b). See Appendix A (Online Supplementary Information) for a detailed account per country on the region’s history and country-specific drivers of deforestation.
The Dry Chaco spans three nations with country-specific land-use practices that likely have contributed to differences in deforestation (Periago et al. 2015; Semper-Pascual et al. 2018), presenting substantial conservation challenges in this biome (Le Polain de Waroux et al. 2018). Although there are large tracts of intact Dry Chaco forest remaining in each country (Fig. 1, these are often limited only to protected areas as found in Paraguay (Mereles and Rodas 2014; Nativa 2020). Yet, protected areas are isolated and may not secure the conservation of large terrestrial vertebrates (Nori et al. 2016). The remaining ecoregion is subject to continued deforestation, with consequences for its biota.
The loss and fragmentation of forest into isolated remnants have likely compromised landscape connectivity within the Dry Chaco (Zak et al. 2004; Gasparri and Grau 2009; Piquer-Rodríguez et al. 2015; Mereles et al. 2020). Landscape connectivity can be defined as how the structural configuration of land-cover types facilitate or impede movement of individuals, and how organisms functionally respond to that structure (Kindlmann and Burel 2008). Landscape connectivity is positively associated with maintenance of genetic diversity, rescue of sink populations, re-establishment of extirpated populations, and minimization of extinction risk, thus making it an integral component of conservation (Taylor et al. 1993; Calabrese and Fagan 2004; Crooks and Sanjayan 2006; Lookingbill et al. 2010). Deforestation has fragmented the Dry Chaco, but to date there has been no assessment of whether connectivity is still relatively intact for its biota. This lack of information hinders conservation of remaining forest remnants and forest-dependent wildlife in this region. Engaging in management hinges upon understanding current patterns of deforestation and how they affect connectivity.
The extent and pace of deforestation in the Chaco indicate an urgent need of quantifying the number, density, and structural connectivity of forest remnants so that appropriate conservation actions can be advised. Herein, we characterize the current state of forest fragmentation in the Dry Chaco of South America via connectivity analyses for Argentina, Bolivia, and Paraguay. We estimate current patterns of structural connectivity within the three nations comprising this biome, determine how connectivity has changed through deforestation in recent time, and identify important areas for future conservation efforts.
Methods
Forest data
We downloaded 30-m resolution forest cover and forest loss data from Global Forest Change 2000–2019 (Hansen et al. 2013) to quantify forest cover for the years 2000 and 2019; this 2013 reference platform includes newer, updated datasets. We defined forested pixels as those with ≥ 30% forest cover, following the threshold used by recent studies (Milodowski et al. 2017; Taubert et al. 2018; Gaveau et al. 2019; Hansen et al. 2020); this cutoff value may overestimate forest coverage by including sparse trees or degraded forest patches but the threshold we used is more conservative than the ≥ 20% threshold used by some other studies (Grantham et al. 2020; Heino et al. 2015; Potapov et al. 2017). We converted raster data to vector format, which created individual forest remnant polygons (a set of contiguous pixels under an 8-neighbor rule; Turner et al. 2003) and calculated the area of each remnant in each of the 2 years for all forest remnants ≥ 0.5 ha. The geographic coordinates of the centroid of each remnant were calculated for connectivity analyses. In addition, we downloaded the extent of protected areas in Argentina, Bolivia, and Paraguay (UNEP-WCMC and IUCN 2020) and quantified the extent to which Dry Chaco forest cover was inside versus outside protected areas.
Connectivity analyses
We analyzed structural connectivity using graph theory, an efficient tool for identification of conservation targets because it does not require demographic data that most other conservation prioritization methods require but that are lacking for most species (Bunn et al. 2000; Minor and Urban 2007; Rayfield et al. 2011). In this approach, forest remnants form the nodes of a graph, and potential dispersal routes between remnants are termed links, represented as Euclidean distances between centroids. This method provides a detailed robust analysis of connectivity with modest data requirements (Calabrese and Fagan 2004). Characteristics of the overall network can be quantified, and individual nodes that are crucial in supporting overall landscape connectivity can be identified as priority conservation sites.
Structural connectivity of the Dry Chaco forest remnants was assessed in RStudio Server Pro in Azure Marketplace via the package igraph (Csardi and Nepusz 2006) in R 4.0.0 (R Core Team 2018), modifying scripts from Drake et al. (2017). Based on the size of the largest dataset (Argentina), we optimized the memory size of our virtual machine to 20 vCPUs, 160 Gb RAM, 32 data disks, 32,000 Max IOPS, and 750 Gb temporary storage. These specs were sufficient to run analyses on the network of forest remnants ≥ 10 ha in Argentina, which is a subset of the entire dataset of remnants ≥ 0.5 ha. Therefore, for comparative purposes, we conducted analyses for only those remnants ≥ 10 ha in Bolivia and Paraguay as well. Forest remnants ≥ 10 ha are large enough to support the home ranges of most of the characteristic mammals of the Dry Chaco (Online Supplementary Material Table S1).
We quantified structural connectivity for each country at three scales: that of the Dry Chaco forest present within each country (via metrics of patch density, path redundancy, and network resiliency; Tischendorf and Fahrig 2000), at potential organism dispersal scales ranging from 40 to 50,000 m (via metrics of clustering), and at the level of assessing the importance of individual forest remnants within each country’s network (via node-level metrics that can identify which remnants facilitate landscape connectivity; Ruiz et al. 2014). For each of these scales, Euclidean distances between forest remnant centroids were used rather than edge-to-edge distances. Since most of the remnants were small relative to the distances between them, using distances between centroids rather than remnant edges is more computationally efficient and still accurately represents graph structure (Galpern et al. 2011). Additionally, the suite of connectivity metrics used is known to be relatively insensitive to our choice of using distances between remnant centroids instead of edge-to-edge distances (McIntyre et al. 2018); see Table 1 for definitions and interpretation of each metric.
Overall-network metrics
We first determined the number of nodes (forest remnants) present within each country. We then calculated the coalescence distance to the nearest meter, which represents the farthest distance between nearest neighboring remnants. For an organism capable of traveling at least that distance, all forest remnants are potentially reachable. Coalescence is thus a threshold distance where the network goes from being fragmented to a single reachable cluster (Keitt et al. 1997). For the coalesced network, we calculated the number of links that connected nodes separated by less than the coalescence distance and four other network-scale metrics (Table 1).
Dispersal-level metrics
We examined connectivity among forest remnants ≥ 10 ha in area at a range of potential wildlife dispersal distances (40, 100, 300, 500, 1000, 5000, 10,000, 20,000, 30,000, 40,000, and 50,000 m); these values represent a gradient of vagility for some of the characteristic fauna of the Dry Chaco, bracketing known dispersal distances (Online Supplementary Material Table S2). At each of these distances, we identified the number of forest remnant clusters (which are a group of ≥ 2 nodes within a given dispersal distance of each other) present; distances where there are sudden changes in the number of clusters present indicate critical fragmentation thresholds that may disproportionately affect wildlife incapable of traveling that distance in a non-forested landscape matrix between forest remnants (Table 1).
Node-level metrics
We used three node-level metrics to identify which remnants were spatially situated to play important roles in supporting landscape connectivity in each country. At coalescence, we plotted the locations of the top 200-ranked stepping stones (the nodes occurring between other nodes), hubs (remnants that are situated near clusters of other remnants), and all cutpoints, (which represent nodes that if removed further fragment the network to a larger coalescence distance); for 2000 and 2019 in Argentina, Bolivia, and Paraguay (Table 1). All metrics were ranked using the ‘order’ function in the igraph package in R.
Collectively, the three node-level metrics allowed us to characterize the structure of the Dry Chaco forest network in terms of the number of forest remnants present (nodes) and at what distance they are potentially accessible by a disperser (coalescence distance, clusters at distances ranging from 40 m to 50 km), their degree of connectedness (number of links, hub score, average nodal connectance), path redundancy within the network (graph density, graph diameter, average nodal connectance), overall network topology (coalescence distance, graph density, graph diameter, transitivity), and the roles of individual nodes (as hubs, stepping stones, or cutpoints). There are dozens of metrics that quantify different aspects of structural connectivity, including many that inform more than one aspect of connectivity (e.g. average nodal connectance is used to assess degree of connectedness as well as path redundancy), but currently there is no theoretical framework to guide expectations for interpretation in different contexts (Tischendorf and Fahrig 2000; Kindlmann and Burel 2008; Kupfer 2012; McIntyre et al. 2018). Our chosen suite of metrics allowed us to examine structural connectivity in terms of the importance of individual remnants on connectivity up to overall topology of the entire Dry Chaco. Connectance and path redundancy have been associated with system resiliency (Janssen et al. 2006; Rayfield et al. 2011), and topological metrics characterize the size and dispersion of the network of remnants. See Csardi and Nepusz (2006) for formulae for each of the metrics and McIntyre et al. (2018) for their ecological interpretation.
Results
Forest loss and fragmentation
From 2000 to 2019, the overall extent of Dry Chaco forest cover decreased by 20.2% (9.5 million ha) (Fig. 1, Table 2). Furthermore, the extent of fragmentation increased, with larger areas of forest divided into multiple, smaller forest remnants: the number of forest remnants increased by 47.0%, and mean forest remnant size decreased by 46.1%. Argentina and Paraguay both lost substantial amounts of forest (4.48 and 4.21 million ha, respectively), with Bolivia experiencing a loss of 1.02 million ha. Paraguay had the greatest percent decrease in forested area between 2000 and 2019 (29.49%), the greatest increase in the number of forest remnants (99,217 additional remnants, representing a 2.6-fold increase from the number of remnants in 2000), and the greatest decrease (72.83%) in mean remnant size (Fig. 2). Argentina experienced forest loss of 20.60% and Bolivia 9.40%. Like Paraguay, both Argentina and Bolivia had an increase in the number of forest remnants, and remnant size shrank in both countries (by 36.01% and 43.04%, respectively) (Fig. 2).

Summary of Dry Chaco forest loss and fragmentation per country, including: A the total amount of Chaco forest per country for 2000 and 2019, based on forest remnants ≥ 0.5 ha; B the total number of forest remnants ≥ 0.5 ha per country for 2000 and 2019; and C average area (± standard error) of forest remnants ≥ 0.5 ha per country for 2000 and 2019
Argentina had the greatest amount of forest cover remaining in both 2000 and 2019 (46.42% and 46.50%, respectively), followed by Paraguay (30.47%, 27.10%), then Bolivia (23.08%, 26.38%; Fig. 2). However, the total amount of Dry Chaco lost between 2000 and 2019 occurred primarily in Argentina (46.05% of the total Dry Chaco lost) and Paraguay (43.37%), followed by Bolivia (10.47%). Argentina had the greatest number of remnants in 2000 and 2019, followed by Paraguay and Bolivia. The average size of forest remnants was greatest in Bolivia, followed by Paraguay and Argentina (Fig. 2).
In 2019, 22.2% of the Dry Chaco region occurred within a protected area (UNESCO-MAB biosphere and multiple-use nature reserves, national parks and reserves, private and public reserves, wildlife refuges, and national monuments). Bolivia had the greatest percent of its Dry Chaco within a protected area (41.54%), followed closely by Paraguay (37.87%). Argentina had the least amount of its Dry Chaco within a protected area (3.51%).
Forest loss and fragmentation also occurred in the total protected areas, with 8.67% lost between 2000 and 2019. Across the entire Dry Chaco, the number of remnants increased 1.7-fold and mean size decreased by 47.39%. Within the protected areas of the three countries, Paraguay experienced the greatest percent loss of Dry Chaco between 2000 and 2019 (13.79% loss), followed by Argentina (8.99% loss) and Bolivia (3.22% loss). Within the protected areas of the three countries, the number of forest remnants increased 2.4-fold in Paraguay, followed by Bolivia (1.7-fold increase) and Argentina (1.3-fold). All three countries experienced a decrease in mean forest remnant size, with the greatest decrease in mean size in Paraguay (64.69%), followed by Bolivia (42.64%) and Argentina (27.29%).
Connectivity analyses
Overall-network metrics
Coalescence distance (the distance when the entire network is connected) varied by country between 2000 and 2019, with inconsistent trends (Table 3). Coalescence distance increased slightly in Argentina but decreased in Bolivia and Paraguay over the same span (Table 3). Graph density between 2000 and 2019 decreased for all three countries; the highest values were found in Bolivia and the lowest in Argentina (Table 3). Argentina showed the least change in graph density between 2000 and 2019 (11.6% decrease), whereas Paraguay had the largest (65.3% decrease; Table 3). Average nodal connectance decreased in Argentina between 2000 and 2019 but increased in Bolivia and Paraguay (Table 3). Graph diameter decreased between 2000 and 2019 for Argentina and Paraguay but increased for Bolivia (Table 3). Finally, transitivity consistently decreased from 2000 to 2019 for all three countries (Table 3).
Dispersal-level metrics
When we examined various dispersal distances below the coalescence distance, clustering of forest remnants was apparent and increased between 2000 and 2019 as numbers of forest remnants increased with fragmentation (Online Supplementary Material Figs. S1–S3). Critical clustering thresholds in the numbers of remnants were found at distances between 1000 and 2000 m for each country in both years (Fig. 3). This indicates that most remnants were > 1000–2000 m apart from each other and only started to form multi-remnant clusters for organisms capable of traveling at least 1000–2000 m in non-forested areas between remnants. This result, coupled with very large coalescence distances (> 40,000 m; Table 3), signals the severity of fragmentation present, even for some relatively vagile mammals characteristic of the biome (Fig. 3; Online Supplementary Material Table S1). The numbers of clusters decreased with increasing potential dispersal distance for each of the three countries and consistently between 2000 and 2019 (Fig. 3; Online Supplementary Material Table S3). The number of clusters increased between 2000 and 2019 for all countries, indicating ongoing forest fragmentation, although this effect was less variable for the greatest potential dispersal distances (Fig. 3; Online Supplementary Material Table S3).

The number of clusters based on potential wildlife dispersal distance for Argentina for 2019 (black circles) and 2000 (gray circles); Paraguay in 2019 (blue squares) and 2000 (light blue squares); and Bolivia in 2019 (red triangles) and 2000 (pink triangles) of Dry Chaco forest remnants ≥ 10 ha in size. Literature- based estimates of travel distances associated to selective species compares connectivity effects per country, shown in clockwise order from top left: Tolypeutes matacus, Alouatta caraya, Tamadua tetradactyla, Priodontes maximus, Leopardus pardalis, Tapirus terrestris, Myrmecophaga tridactyla, Tayassu pecari, and Panthera onca (also see Online Supplementary Material Table S2). Maximum distance is plotted, as this closely represents coalescence distance (distance where the entire network is connected) for each country
Node-level metrics
The locations of the top 200-ranked hubs, top 200-ranked stepping stones, and all cutpoints for each country in 2000 and 2019 highlight the forest remnants serving important functions as network topology changed (Fig. 4). The top 200 hubs shifted in Bolivia and Paraguay, but remained in consistent locations in Argentina. The top 200 hubs were densely concentrated in all three countries in 2000 and 2019, with the exception of Bolivia in 2000 (Fig. 4B), when the hubs were distributed throughout the western region of the Dry Chaco. Shifts in the locations of the top 200 stepping stones were apparent in all three countries (Fig. 4). Each country had two cutpoints in 2000 and 2019, with the exception of Paraguay in 2019, when there was one cutpoint.

Dry Chaco forest remnants (in green) in Bolivia in A 2000 and B 2019; Paraguay in C 2000 and D 2019; and Argentina in E 2000 and F 2019. The top 200 stepping stones are indicated as white circles, top 200 hubs as black circles, and all cutpoints as green triangles. The symbols are enlarged relative to the size of the remnants so as to be more readily apparent
Some remnants played multiple connectivity roles (e.g. as both stepping stones and hubs, or as both cutpoints and stepping stones); the consequences of losing those remnants are especially severe because such remnants are especially crucial for conservation (Fig. 4). In several countries, these multiple connectivity roles disappeared by 2019. For example, in 2000 in Bolivia, both cutpoints also served as stepping stones, and 11 of the top 200 stepping stones and hubs overlapped. In 2019 neither cutpoints served as stepping stones, and only 5 forest remnants served as both stepping stones and hubs (Fig. 4A, B). In Paraguay in 2000 the two cutpoints were also stepping stones, but in 2019 there was no overlap between the singular cutpoint, top 200 stepping stones, or top 200 hubs (Fig. 4C, D). In Argentina, each of the two cutpoints in 2000 were the same as in 2019, and these two cutpoints in 2000 and 2019 were also stepping stones; however, there was no overlap with the top 200 hubs and stepping stones in neither 2000 nor 2019.
Discussion
Our study indicates that the current state of deforestation in the Dry Chaco is extensive, with 79.8% of the remaining forest fragmented into millions of remnants that are spatially isolated at distances beyond the dispersal capacity of many organisms. Our results are alarming for the conservation of the region and its diversity, especially considering that our estimations are optimistic. First, because we used smaller coalescence distances (Table 3) than the known dispersal distances of most species (Online Supplementary Material Table S1). Second, because we did not consider the negative effects that edges of forest patches have on wildlife individuals and populations.
Deforestation incurs forest cover loss and also creates numerous forest patch edges. The increase of edges in forest patches may decrease individuals´ fitness and also have negative demographic and genetic consequences in wildlife populations (Laurance et al. 2011; Chase et al. 2020). These edge effects are species-specific and are influenced by the nature of the landscape matrix between forest remnants (Pardini 2004). Effects on individuals and populations are the consequence of a lower probability of movement into or out of a forest remnant due to forest edges (Laurance et al. 2011). Also, forest edges may have sub-optimal habitat as compared with forested areas that have more distant to the edge (Chase et al. 2020). For example, edges of forest patches can result in novel species interactions among wildlife, livestock, and humans, which can result in increased competition and stress; and can also increase or induce emergence of diseases, cause an increase in poaching and hunting, and other consequences (Romero-Muñoz et al. 2020b; Weems et al. 2021), further complicating conservation efforts aimed at promoting connectivity among remnants.
Forest loss and fragmentation were documented in each of the three countries where the Dry Chaco is located, but Paraguay has experienced the greatest loss of forest cover and the greatest degree of fragmentation-induced loss of landscape connectivity. In 2000, the vast majority of the Dry Chaco in northern Paraguay was relatively unfragmented (Fig. 1). Many of the disconnected fragments in 2019 represent formerly large, continuous forest tracts. With these country-specific patterns of loss we also found differences in system connectivity over time and by country, with each country exhibiting different patterns of fragmentation. All three countries saw large increases in the number of nodes (i.e., forest remnants) from 2000 to 2019, indicative of extensive fragmentation. The continued fragmentation of formerly continuous habitat is shown as increased clustering in the center of the region. The principal cluster in 2000 was around the town of Filadelfia, Paraguay. By 2019 this clustering had expanded into most of central and western Paraguay with the creation of more remnants there (Figs. 1, 3, Online Supplementary Material Fig. S3). In Argentina most of the deforestation has occurred along the western and southern portions of the Dry Chaco (Figs. 1, 3, Online Supplementary Material Fig. S1). In Bolivia, most deforestation occurred along the northern portions of the Dry Chaco, with large, intact fragments remaining in the central portion of this biogeographic region (Figs. 1, 3, Online Supplementary Material Fig. S2).
These differences in forest fragmentation across the three countries of the Dry Chaco are thus reflected in different national patterns of connectivity. Although the organisms of the Dry Chaco do not perceive geopolitical boundaries, because conservation happens at the level of each nation, it is important to identify areas where each country may focus protection efforts. For example, southeastern that is isolated from the rest in that country but is contiguous with Dry Chaco in north-eastern Paraguay; this area was identified as having one of the two cutpoints for the country of Bolivia in both 2000 and 2019 (Fig. 4A, B), although they may not have been identified as such if the entire network without national borders had been examined. Nevertheless, this area is one that can be a focus for Bolivia in conserving its own natural resources.
As the number of forest remnants increased with fragmentation, overall network topology changed, with this also varying by country. For example, whereas the coalescence distance increased slightly in Argentina from 2000 to 2019, it decreased in Bolivia and in Paraguay over that same time span. This variability indicates differences in the nature of fragmentation that can be seen in Figs. 1 and 4: forest losses in Bolivia and Paraguay occurred primarily along the outer borders of the Dry Chaco, effectively shrinking the forest to a central core area, whereas in Argentina, fragmentation was interspersed. Bolivia had the lowest average nodal connectance, suggesting that it has the greatest potential dispersal efficiency within the central core remaining (apparent in Figs. 1 and 4), whereas Argentina had the lowest potential dispersal efficiency because of the perforated nature of its fragmentation. Because all three countries experienced a high degree of fragmentation, some connectivity metrics show consistent patterns across all three. For example, graph density decreased between 2000 and 2019 for all three countries, signaling that connectivity path options throughout the network decreased for all three countries. Similarly, transitivity decreased for all three, indicating less clustering/more isolation of forest remnants.
Changes to the amount and configuration of forest cover have taxon-specific effects (Núñez-Regueiro et al. 2015). For example, volant taxa such as bats or birds are likely less affected by changes to habitat availability than overland dispersers such as terrestrial mammals (Bélisle 2005). That said, habitat loss and fragmentation can also impact bats and birds (Sandoval and Barquez 2013; Názaro et al. 2020). For organisms that may not use approximately straight-line dispersal routes (e.g. overland dispersers), our approach using Euclidean distances among remnants may overestimate landscape connectivity. Our approach was based on landscape patterns rather than species-specific data, which are currently limited in this region for most species. Although some species can survive in agrarian/forest mosaics (Weiler et al. 2020), their survival may diminish over time (Núñez-Regueiro et al. 2015).
Our results lead us to support previous research that suggests that most wildlife species in the Dry Chaco are highly threatened from land-cover change (Periago et al. 2017; Aguilar et al. 2018; Názaro et al. 2020). A graph theory approach to quantifying landscape connectivity is based on the spatial arrangement of fragments, which can be used to make inferences about animal movements even when observed movement or dispersal data are unavailable. Such an approach is especially valuable when rates of deforestation and habitat loss are dramatic and rapid and when knowledge of impacts of specific taxa are limited. The resolution of our analyses allowed us to detect connectivity patterns for a large range of taxa. Species differ in their dispersal abilities and home range sizes, which has implications to determining the effects of forest fragmentation on landscape connectivity. However, we found that the degree of spatial separation of forest remnants was well beyond the dispersal range of most of the mammals of the Dry Chaco and many remnants are now too small to support home ranges of many of these species, with these effects increasing between 2000 and 2019 (Fig. 3). Entire taxonomic orders are often affected by forest loss in the same manner; although some of these species can survive in agrarian/forest mosaics (Weiler et al. 2020), their survival can diminish over time (Núñez-Regueiro et al. 2015). Additionally, the two components of fragmentation–habitat loss and habitat isolation–act in tandem. Even those species with home ranges < 10 ha may not be able to persist in these perforated landscapes, as the distance between remnants effectively isolates them.
Species appear to differ in their responses to deforestation in the Dry Chaco (Brooks 1998; Periago et al. 2017). There have been some connectivity assessments for some megafauna (e.g. Romero-Muñoz et al. 2020a, b). For example the most vagile species, the large carnivores such as pumas (Puma concolor) and jaguars (Panthera onca), require very large expanses of unmodified habitat to persist and maintain healthy populations (Maffei et al. 2004; Romero-Muñoz et al. 2020b) (Fig. 3). Forest fragmentation can result in reduced food availability for collared peccaries (Pecari tajacu; Semper-Pascual et al. 2018), anthropogenically-facilitated range expansion of capybara (Hydrochoerus hydrochaeris; Campos-Krauer and Wisely 2011), and reduced occurrence of large vertebrates and forest specialists (Núñez-Regueiro et al. 2015; Weiler et al. 2020). Primates are a good example of a group likely to be most affected in the long term, as many primate species are negatively impacted by habitat loss and fragmentation (Brown and Rumiz 1986; Stallings 1989; Estrada et al. 2017). While some species of Paraguayan primates have shown the potential to cross short gaps between forest patches (e.g., Cabral et al. 2017), they simply cannot thrive in small patches of forest over the long term (Stallings 1989). Unfortunately, for the vast majority of species, we currently have little to no data on dispersal; for example, knowledge of small mammals, which make up the majority of mammalian diversity in the region, is limited to distributional data, with major gaps for many species that are only known from a few records (López–González 2005; de la Sancha 2007, 2012, 2014, 2017, 2020; Teta et al. 2009, 2018; de la Sancha 2014; de la Sancha and D’Elia 2015; de la Sancha and Boyle 2019). Data for small mammal populations are particularly lacking an especially important consideration given that these taxa comprise the majority of mammalian diversity in these systems (de la Sancha et al. 2017; Teta et al. 2018). Despite this lack of information, our approach provides robust models for species of various sizes and dispersal abilities, as smaller fragments provide reasonable refuge for medium and small mammals, and they also serve as stepping stones for more vagile species.
In our study, we analyzed the Dry Chaco forest as a habitat-patch network described as a graph of nodes connected by actual or potential dispersal links, which we approximated as Euclidean distances between forest remnant centroids. The number and arrangement of forest remnants is a macro-scaled pattern of structural connectivity that ignores the intervening landscape matrix as well as potential differences in nodes (e.g. in terms of quality). Habitat patches do not exist in isolation, the intervening landscape mosaic is known to influence the dispersal of organisms between habitat patches (Fahrig 2007). Thus, an assessment of functional connectivity is needed for this region but would need to be conducted separately for each species or functional group of species. However, the extent and rapid pace of deforestation in the Dry Chaco indicates an urgent need to quantify the number and separation of remnants and the roles they play in overall landscape connectivity so that appropriate conservation actions can be advised. Our structural approach to understanding connectivity is thus sound and important yet incomplete, as drivers of landscape connectivity are complex and include species’ functional responses to landscape structure.
Our study is unique in documenting forest loss and changes to structural connectivity in the Dry Chaco biome, and in finding that patterns of fragmentation differ by country, meaning that country-specific conservation measures will be necessary. Important first steps for identifying conservation priorities for the Dry Chaco have been taken (TNC et al. 2005; Nativa 2020). Among priority sites for conservation identified by these efforts, areas transformed into pastures and crops between 2002 and 2017 increased by more than 100%, from 2.2 million to 5.5 million ha. Seven of 38 recently identified priority conservation sites still have more than 80% of their original forest area (e.g. Kaa Iya, Bolivia, and Defensores del Chaco, Paraguay); 24 contain more than 50% of forest area and a few others up to 30% (eg. Teniente Enciso and Pratt Gill in Paraguay) (Nativa 2020). Bolivia has the greatest percentage of its Dry Chaco protected, followed by Paraguay and Argentina. Alarmingly, Paraguay experienced substantial forest loss in its protected areas between 2000 and 2019: The expanse of much of the deforested areas are clearly within the boundaries of protected areas (Fig. 1). Although Argentina has a robust network of protected areas, these are virtually non-existent in its Chaco (Fehlenberg et al. 2017), highly isolated (Matteucci and Camino 2012) and with low representativity of large terrestrial vertebrate species (Nori et al. 2016). In general, many important areas remain unprotected, and diverse strategies that include substantial international coordination are warranted to achieve effective conservation across national borders (Guyra Paraguay 2008). We used a rapid-assessment approach to identify remnants that facilitate overall landscape connectivity for forest-dependent wildlife. Our analyses revealed some remaining areas of contiguous forest, with many isolated remnants in the northeast and along the westernmost edge of the Dry Chaco (Fig. 1). Our graph theory approach allowed us to indicate where conservation activities could be conducted so as to best support landscape-level connectivity in the Dry Chaco (specifically, the remnants identified in Fig. 4). Such activities are clearly and urgently needed in this highly imperiled biodiversity hotspot.
Data availability
All data used is either available as supporting materials or will be made available upon request. We would like to include our forest shapefiles we developed as part of our supporting material when the article is accepted.
Code availability
R code is available from the corresponding author.
References
Aguilar R, Calviño A, Ashworth L, Aguirre-Acosta N, Carbone LM, Albrieu-Llinás G, Nolasco M, Ghilardi A, Cagnolo L (2018) Unprecedented plant species loss after a decade in fragmented subtropical Chaco Serrano forests. PLoS ONE 13:e0206738
Andrade-Díaz MS, Sarquis JA, Loiselle BA, Giraudo AR, Díaz-Gómez JM (2019) Expansion of the agricultural frontier in the largest South American dry forest: identifying priority conservation areas for snakes before everything is lost. PLoS ONE 14:e0221901
Ascensão F, Yoguid D, Alvesd M, Medicig EP, Desbiez A (2019) Predicting spatiotemporal patterns of road mortality for medium-large mammals. J Environ Manag 248:109320
Bélisle M (2005) Measuring landscape connectivity: the challenge of behavioral landscape ecology. Ecology 86:1988–1995
Brancalion PHS, Niamir A, Broadbent E, Crouzeilles R, Barros FSM, Almeyda Zambrano AM, Baccini A, Aronson J, Goetz S, Reid JL, Strassburg BBN, Wilson S, Chazdon RL (2019) Global restoration opportunities in tropical rainforest landscapes. Sci Adv 5:eaav3223
Brooks DM (1998) Habitat variability as a predictor of rarity in large Chacoan mammals. Vida Silvestre Neotrop 7:115–120
Brown AD, Rumiz DI (1986) Distribucion y conservacion de los primates en Bolivia - estado actual de conocimiento. In: de Mello MT (eds) A primatologia no Brasil, vol 2. Sociedade Brasileira de Primatologia. Brasilia, Brazil, pp 335–363
Bunn AG, Urban DL, Keitt TH (2000) Landscape connectivity: a conservation application of graph theory. J Environ Manag 59:265–278
Cáceres DM (2015) Accumulation by dispossession and socio-environmental conflicts caused by the expansion of agribusiness in Argentina. J Agrar Change 15:116–147
Cabral-B H, Yanosky A, Romero-N L, Bueno D, Brooks DM (2017) A new locality in Paraguay for the Black-tailed Marmoset, Mico melanurus (Geoffroy Saint-Hilaire, 1812) (Primates, Callitrichidae). Check List 13:1–4.
Calabrese JM, Fagan WF (2004) A comparison-shopper’s guide to connectivity metrics. Front Ecol Environ 2:529–536
Caldas MM, Goodin D, Sherwood S, Campos Krauer JM, Wisely SM (2015) Land-cover change in the Paraguayan Chaco: 2000–2011. J Land Use Sci 10:1–18
Campos-Krauer JM, Wisely SM (2011) Deforestation and cattle ranching drive rapid range expansion of capybara in the Gran Chaco ecosystem. Glob Change Biol 17:206–218.
Carranza ML, Frate L, Acosta AT, Hoyos L, Ricotta C, Cabido M (2014) Measuring forest fragmentation using multitemporal remotely sensed data: three decades of change in the dry Chaco. Eur J Remote Sens 47:793–804
Chase JM, Blowes SA, Knight TM, Gerstner K, May F (2020) Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature 584:238–243
Crooks KR, Sanjayan M (2006) Connectivity conservation: maintaining connections for nature. In: Crooks KR, Sanjayan M (eds) Connectivity conservation. Cambridge University Press, Cambridge, pp 1–19
Csardi G, Nepusz T (2006) The igraph software package for complex network research. InterJ Complex Syst 1695 5:1
Curtis PG, Slay CM, Harris NL, Tyukavina A, Hansen MC (2018) Classifying drivers of global forest loss. Science 361:1108–1111
de la Sancha NU (2014) Patterns of small mammal diversity in fragments of subtropical Interior Atlantic Forest in eastern Paraguay. Mammalia 78:437–449
de la Sancha NU, Boyle SA (2019) Predictive sampling effort and species-area relationship models for estimating richness in fragmented landscapes. PLoS ONE 14(12):e0226529.
de la Sancha NU, D’Elía G (2015) Additions to the Paraguayan mammal fauna: the first records of two marsupials (Didelphimorphia, Didelphidae) with comments on the alpha taxonomy of Cryptonanus and Philander. Mammalia 79:343–356
de la Sancha NU, Solari S, Owen RD (2007) First records of Monodelphis kunsi Pine (Didelphimorphia, Didelphidae) from Paraguay, with an evaluation of its distribution. Mastozool Neotrop 14:241–247
de la Sancha NU, D’Elía G, Teta P (2012) Systematics of the subgenus of mouse opossums Marmosa (Micoureus) (Didelphimorphia, Didelphidae) with noteworthy records from Paraguay. Mamm Biol-Zeitschrift Für Säugetierkunde 77:229–236
de la Sancha NU, Higgins CL, Presley SJ, Strauss RE (2014) Metacommunity structure in a highly fragmented forest: has deforestation in the Atlantic Forest altered historic biogeographic patterns? Divers Distrib 20:1058–1070
de la Sancha NU, López-González C, D’Elía G, Myers P, Valdez L, Ortiz ML (2017) An annotated checklist of the mammals of Paraguay. Therya 8:241–260
de la Sancha NU, Maestri R, Bovendorp RS, Higgins CL (2020) Disentangling drivers of small mammal diversity in a highly fragmented forest system. Biotropica 52:182–195.
Drake JC, Griffis-Kyle K, McIntyre NE (2017) Using nested connectivity models to resolve management conflicts of isolated water networks in the Sonoran Desert. Ecosphere 8:e01652
DRYFLOR, Pennington RT, Banda-R K, Delgado-Salinas A, Dexter KG, Linares-Palomino R, Maturo HM, Mogni V, Oakley L, Olivera-Filho A, Prado D, Quintana C, Riina R, Särkinen T (2017) Forest conservation: remember Gran Chaco—response. Science 355:465–466
Estrada A, Garber PA, Rylands AB, Roos C, Fernandez-Duque E, Di Fiore A, Nekaris KAI, Nijman V, Heymann EW, Lambert JE, Rovero F, Barelli C, Setchell JM, Gillespie TR, Mittermeier RA, Arregoitia LV, de Guinea M, Gouveia S, Dobrovolski R, Shanee S, Shanee N, Boyle SA, Fuentes A, MacKinnon KC, Amato KR, Meyer ALS, Wich S, Sussman RW, Pan R, Kone I, Baoguo L (2017) Impending extinction crisis of the world’s primates: why primates matter. Sci Adv 3:e1600946
Fahrig L (2007) Non-optimal animal movement in human-altered landscapes. Funct Ecol 21:1003–1015
FAO (United Nations Food and Agriculture Organization) (2015) Global Forest Resources Assessment 2015. FAO Forestry Paper No. 1. U.N. Food and Agriculture Organization, Rome, Italy
Fehlenberg V, Baumann M, Gasparri NI, Piquer-Rodriguez M, Gavier-Pizarro G, Kuemmerle T (2017) The role of soybean production as an underlying driver of deforestation in the South American Chaco. Glob Environ Change 45:24–34
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski JH, Holloway T, Howard EA, Kucharik CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309:570–574
Galpern P, Manseau M, Fall A (2011) Patch-based graphs of landscape connectivity: a guide to construction, analysis and application for conservation. Biol Conserv 144:44–55
Gasparri NI, Grau HR (2009) Deforestation and fragmentation of Chaco dry forest in NW Argentina (1972–2007). For Ecol Manag 258:913–921
Gaveau DLA, Locatelli B, Salim MA, Yaen H, Pacheco P, Shell D (2019) Rise and fall of forest loss and industrial plantations in Borneo (2000–2017). Conserv Lett 12:e12622
Grantham HS, Duncan A, Evans JKR, Beyer HL, Schuster R, Walston J, Ray JC, Robinson JG, Callow M, Clements T, Costa HM, DeGemmis A, Elsen PR, Ervin J, Franco P, Goldman E, Goetz S, Hansen A, Hofsvang E, Jantz P, Jupiter S, Kang A, Langhammer P, Laurance WF, Lieberman S, Linkie M, Malhi Y, Maxwell S, Mendez M, Mittermeier R, Murray NJ, Possingham H, Radachowsky J, Saatchi S, Samper C, Silverman J, Shapiro A, Strassburg B, Stevens T, Stokes E, Taylor R, Tear T, Tizard R, Venter O, Visconti P, Wang S, Watson JEM (2020) Anthropogenic modification of forests means only 40% of remaining forests have high ecosystem integrity. Nat Commun 11:5978
Grau HR, Gasparri NI, Aide TM (2008) Balancing food production and nature conservation in the Neotropical dry forests of northern Argentina. Glob Change Biol 14:985–997
Guyra Paraguay (2008) Áreas de importancia para la conservación de las aves en Paraguay. Guyra Paraguay/BirdLife International, Asunción
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song DX, Townshend JR (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:e1500052
Hansen MC, Potapov PV, Moore R, Hancher M, Turubanova SA, Tyukavina A, Thau D, Stehman SV, Goetz SJ, Loveland TR, Kommareddy A, Egorov A, Chini L, Justice CO, Townshend JRG (2013) High resolution global maps of 21st-century forest cover change. Science 342:850–853
Hansen MC, Wang L, Song X-P, Tyukavina A, Turubanova S, Potapov SSV (2020) The fate of tropical forest fragments. Sci Adv 6:eaax8574
Heino M, Kummu M, Makkonen M, Mulligan M, Verburg PH, Jalava M, Räsänen TA (2015) Forest loss in protected areas and intact forest landscapes: a global analysis. PLoS ONE 10:e0138918
Janssen MA, Bodin Ö, Anderies JM, Elmqvist T, Ernstson H, McAllister RRJ, Olsson P, Ryan P (2006) Toward a network perspective on the resilience of social-ecological systems. Ecol Soc 11:15
Keitt TH, Urban DL, Milne BT (1997) Detecting critical scales in fragmented landscapes. Conserv Ecol 1:4
Kindlmann P, Burel F (2008) Connectivity measures: a review. Landsc Ecol 23:879–890
Kuemmerle T, Altrichter M, Baldi G, Cabido M, Camino M, Cuellar E, Cuellar RL, Decarre J, Díaz S, Gasparri I, Gavier-Pizarro G, Ginzburg R, Giordano AJ, Grau HR, Jobbágy E, Leynaud G, Macchi L, Mastrangelo M, Matteucci SD, Noss A, Paruelo J, Piquer-Rodríguez M, Romero-Muñoz A, Semper-Pascual A, Thompson J, Torrella S, Torres R, Volante JN, Yanosky A, Zak M (2017) Forest conservation: remember Gran Chaco. Science 355:465–465
Kupfer JA (2012) Landscape ecology and biogeography: rethinking landscape metrics in a post-FRAGSTATS world. Prog Phys Geogr 36:400–420
Laurance WF, Camargo JLC, Luizāo RCC, Laurance SG, Pimm SL, Bruna EM, Stouffer PC, Williamson BG, Benítez-Malvido J, Vasconcelos HL, Van Houtan KS, Zartman CE, Boyle SA, Didham RK, Andrade A, Lovejoy TE (2011) The fate of Amazonian forest fragments: a 32-year investigation. Biol Conserv 144:56–67
Leguizamón A (2016) Disappearing nature? Agribusiness, biotechnology and distance in Argentine soybean production. J Peasant Stud 43:313–330
Le Polain de Waroux Y, Baumann M, Gasparri NI, Gavier-Pizarro G, Godar J, Kuemmerle T, Müller R, Vázquez F, Volante JN, Meyfroidt P (2018) Rents, actors, and the expansion of commodity frontiers in the Gran Chaco. Ann Am Assoc Geogr 108:204–225
Lookingbill TR, Gardner RH, Ferrari J, Keller C (2010) Combining a dispersal model with network theory to assess habitat connectivity. Ecol Appl 20:427–441
López–González C (2005) Murciélagos del Paraguay. Biosfera, Publicaciones del Comité Español del Programa MaB y de la Red IberoMaB de la UNESCO. Sevilla, Spain
Maffei L, Cuéllar E, Noss A (2004) One thousand jaguars (Panthera onca) in Bolivia’s Chaco? Camera trapping in the Kaa-Iya National Park. J Zool 262:295–304
Matteucci SD, Camino M (2012) Protected Areas Isolation in the Chaco Region, Argentina. J Geogr Geol 4:15–28
McBride RT Jr, Thompson JJ (2019) Spatial ecology of Paraguay’s last remaining Atlantic Forest jaguars (Panthera onca): implications for their long-term survival. Biodiversity 20:20–26
McIntyre NE, Collins SD, Heintzman LJ, Starr SM, van Gestel N (2018) The challenge of assaying landscape connectivity in a changing world: a 27-year case study in the southern Great Plains (USA) playa network. Ecol Ind 91:607–616
Mereles MF, Céspedes G, Cartes JL, Goerzen R, de Egea-Elsam J, Rodríguez L, Yanosky A, Villalba L, Weiler A, Cacciali P (2020) Biological corridors as a connectivity tool in the region of the Great American Chaco: identification of biodiversity hotspots in the ecoregions of the Paraguayan Chaco. Res Ecol 2:27–36
Mereles MF, Rodas O (2014) Assessment of rates of deforestation classes in the Paraguayan Chaco (Great South American Chaco) with comments on the vulnerability of forest fragments to climate change. Clim Change 127:55–71
Milodowski DT, Mitchard ETA, Williams M (2017) Forest loss maps from regional satellite monitoring systematically underestimate deforestation in two rapidly changing parts of the Amazon. Environ Lett 12:094003
Minor ES, Urban DL (2007) Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol Appl 17:1771–1782
Nativa (2020) Evaluación Ecorregional – Actualización al 2018 – documento resumen. https://nativabolivia.org/wp-content/uploads/2020/05/Resumen-Evaluaci%C3%B3n-Ecorregional-actualizaci%C3%B3n-2018.pdf.
Názaro MG, Dos Santos DA, Torres R, Baumann M, Blendinger PG (2020) Untangling the imprints of climate, geography and land use/cover on bird diversity in the South American Gran Chaco. J Biogeogr 47:1439–1454
Nori J, Torres R, Lescano JN, Cordier JM, Periago ME, Baldo D (2016) Protected areas and spatial conservation priorities for endemic vertebrates of the Gran Chaco, one of the most threatened ecoregions of the world. Divers Distrib 22:1212–1219
Núñez-Regueiro MM, Branch L, Fletcher RJ Jr, Marás GA, Derlindati E, Tálamo A (2015) Spatial patterns of mammal occurrence in forest strips surrounded by agricultural crops of the Chaco region, Argentina. Biol Conserv 187:19–26
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GVN, Underwood EC, D'Amico JA, Itoua I, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience 51:933–938
Pardini R (2004) Effects of forest fragmentation on small mammals in an Atlantic Forest landscape. Biodivers Conserv 13:2567–2586
Periago ME, Chillo V, Ojeda RA (2015) Loss of mammalian species from the South American Gran Chaco: empty savanna syndrome? Mammal Rev 45:41–53
Periago ME, Tamburini DM, Ojeda RA, Cáceres DM, Díaz S (2017) Combining ecological aspects and local knowledge for the conservation of two native mammals in the Gran Chaco. J Arid Environ 147:54–62
Piquer-Rodríguez M, Torella S, Gavier-Pizarro G, Volante J, Somma D, Ginzburg R, Kuemmerle T (2015) Effects of past and future land conversions on forest connectivity in the Argentine Chaco. Landsc Ecol 30:817–833
Potapov P, Mansen MC, Laestadius L, Turubanova S, Yaroshenko A, Thies C, Smith W, Zhuravleva I, Komarova A, Minnemeyer S, Esipova E (2017) The last frontiers of wilderness: tracking loss of intact forest landscapes from 2000 to 2013. Sci Adv 3:e1600821
Prado D (1993) What is the Gran Chaco vegetation in South America? I: A review. Contribution to the study of flora and vegetation of the Chaco. Candollea 48:145–172
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Rayfield B, Fortin M-J, Fall A (2011) Connectivity for conservation: a framework to classify network measures. Ecology 92:847–858
Redford KH, Taber A, Simonetti JA (1990) There is more to biodiversity than the tropical rain forests. Conserv Biol 4:328–330
Romero-Muñoz A, Fandos G, Benítez-López A, Kuemmerle T (2020a) Habitat destructions and overexploitation drive widespread declines in all facets of mammalian diversity in the Gran Chaco. Glob Change Biol. https://doi.org/10.1111/gcb.15418
Romero-Muñoz A, Benítez-López A, Zurell D, Baumann M, Camino M, Decarre J, del Castillo H, Giordano AJ, Gómez-Valencia B, Levers C, Noss AJ, Quiroga V, Thompson JJ, Torres H, Velilla M, Weiler A, Kuemmerle T (2020b) Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography 43:954–966
Ruiz LJ, Parikh NN, Heintzman LJ, Collins SD, Starr SM, Wright CK, Henebry GM, van Gestel N, McIntyre NE (2014) Dynamic connectivity of temporary wetlands in the southern Great Plains. Landsc Ecol 29:507–516
Sandoval ML, Barquez RM (2013) La identidad de la fauna de murciélagos del Chaco: patrones de congruencia distribucional e implicancias en su conservación. Rev Chil Hist Nat 86:75–94
Semper-Pascual A, Macchi L, Sabatini FM, Decarre J, Baumann M, Blendinger PG, Gómez-Valencia B, Mastrangelo ME, Kuemmerle T (2018) Mapping extinction debt highlights conservation opportunities for birds and mammals in the South American Chaco. J Appl Ecol 55:1218–1229
Sloan S, Slayer JA (2015) Forest Resources Assessment of 2015 shows positive global trends but forest loss and degradation persist in poor tropical countries. For Ecol Manag 352:134–145
Stallings J (1989) Status y conservación de primates en el Paraguay. In: Saavedra CJ, Mittermeier RA, Santos IB (eds) La Primatología en Latinoamérica. World Wildlife Fund, Washington, D.C., USA, pp 133–151
Taubert F, Fischer R, Groeneveld J, Lahmann S, Müller MS, Rödig E, Wiegand T, Huth A (2018) Global patterns of tropical forest fragmentation. Nature 554:519–522
Taylor PD, Fahrig L, Henein K, Merriam G (1993) Connectivity is a vital element of landscape structure. Oikos 68:571–573
Teta P, D’Elía G, Flores D, de la Sancha NU (2009) Diversity and distribution of the mouse opossums of the genus Thylamys (Didelphimorphia, Didelphidae) in northeastern and central Argentina. Gayana 73:180–199
Teta P, Abba AM, Cassini GH, Flores DA, Galliari CA, Lucero SO, Ramírez M (2018) Lista revisada de los mamíferos de Argentina. Mastozool Neotrop 25:163–198
Tischendorf L, Fahrig L (2000) How should we measure landscape connectivity? Landsc Ecol 15:633–641
TNC (The Nature Conservancy), Fundación Vida Silvestre Argentina, Fundación para el Desarrollo Sustentable del Chaco (DeSdel Chaco), Wildife Conservation Society Bolivia (WCS) (2005) Evaluación Ecorregional del Gran Chaco Americano / Gran Chaco Americano Ecoregional Assessment. Fundación Vida Silvestre Argentina, Buenos Aires
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
Turner MG, Gardner RH, O’Neill RV (2003) Landscape ecology in theory and practice: pattern and process. Springer, New York
UNEP-WCMC, IUCN (2020) Protected planet: the world database on protected areas (WDPA). http://www.protectedplanet.net. Accessed 4 Dec 2020.
Weems EI, de la Sancha NU, Anderson LJ, Zambrana-Torrelio C, Ferraris RP (2021) Centering microbes in the emerging role of integrative biology in understanding environmental change. Integr Comp Biol. https://doi.org/10.1093/icb/icab047
Weiler A, Núñez K, Silla F (2020) Forest matters: use of water reservoirs by mammal communities in cattle ranch landscapes in the Paraguayan Dry Chaco. Glob Ecol Conserv 23:e01103
Zak MR, Cabido M, Hodgson JG (2004) Do subtropical seasonal forests in the Gran Chaco, Argentina, have a future? Biol Conserv 120:589–598
Funding
Partial financial support to ND came from the Grainer Bioinformatics Center at The Field Museum.
Author information
Authors and Affiliations
Contributions
Conceptualization: NUdlS, SAB, NEM, DMB, AY, ECS, FM, MC, RDS; Data curation: NUdlS, SAB, Formal analysis: NUdlS, SAB, Funding acquisition: NUdlS, SAB; Methodology: NUdlS, SAB, NEM; Project administration: NUdlS, SAB, NEM; Validation: NUdlS, SAB, NEM; Visualization: NUdlS, SAB; Writing—original draft: NUdlS, SAB, NEM, DMB, AY, ECS, FM, MC, RDS; Writing—review and editing: NUdlS, SAB, NEM, DMB, RDS. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Consent to participate
All authors consented to participate on this manuscript.
Consent for publication
All authors have appropriate consent to publish this data from their institutions.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
de la Sancha, N.U., Boyle, S.A., McIntyre, N.E. et al. The disappearing Dry Chaco, one of the last dry forest systems on earth. Landscape Ecol 36, 2997–3012 (2021). https://doi.org/10.1007/s10980-021-01291-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-021-01291-x