
Abstract
Context
Conservation practitioners face complex decisions about the management of spatial and temporal disturbance regimes when disturbance plays a significant role in the dynamics of fragmented populations. This is particularly so if disturbance events have negative short-term consequences but are necessary for maintaining habitat quality in the longer term.
Objectives
We sought to determine the potentially interactive effects of two disturbance processes (fire and grazing) on the occupancy dynamics of the threatened Striped Legless Lizard (Delma impar) among native grassland fragments in Victoria, Australia.
Methods
We collected 9 years of occupancy data from 291 sites. We used these data to examine the influence of disturbance frequency and coincidence on rates of occupancy and persistence, plus the influence of grassland connectivity on colonisation rates.
Results
Fire and grazing were positively related to the persistence of D. impar populations at the frequencies observed during this study, but the interaction of these two disturbances had a strong negative effect on persistence. Colonisation of grassland fragments was positively related to connectivity to grassland habitat in the surrounding landscape.
Conclusions
Strategic use of fire and grazing to manage fragmented grasslands can be beneficial for the persistence of D. impar populations, but intense and concurrent application of both these disturbances may trigger population collapse. Conservation risks of inappropriate disturbance are highest in small, isolated fragments of grassland, where recolonisation is unlikely. More broadly, our study highlights the value of coupling long-term occupancy and management data to investigate the influence of disturbance processes on population persistence in fragmented landscapes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Appropriate disturbance regimes present complex challenges for threatened species conservation. In the short term, disturbance events—such as fire, floods, drought or grazing pulses—can have negative impacts on threatened species due to spikes in mortality and/or failure of recruitment, leading to population crashes (Burgman and Lindenmayer 1998). The impacts of such ‘catastrophes’ have long been the focus of population viability analyses, given the strong theoretical and empirical evidence for environmental stochasticity as a key driver of the dynamics of small populations (Beissinger 2002). Conversely, occasional, or even frequent, disturbances can be critical for species persistence. Specialists of early successional vegetation communities are important examples, as are those which rely on specific structural or biotic resources that are only provisioned by disturbance events (e.g. Brawn et al. 2001; Hunter et al. 2001).
Whether disturbance has positive or negative impacts on a species, the challenge for conservation biologists lies in understanding the functional impacts of disturbance events and identifying pathways to manage these impacts. When disturbance events cannot readily be controlled (for example, climatic phenomena such as drought and floods), managers must implement broader initiatives that ensure resilience to short-term impacts and/or facilitate longer-term prosperity during ecosystem response (Gunderson 2000). When disturbance can be controlled, such as grazing or fire regimes in grassland ecosystems, managers must identify the optimal rate, intensity and timing of disturbance events, and implement those disturbances in accord with the conceptualised optimal regime (Lunt et al. 2007).
In fragmented landscapes, the difficulties of managing the impacts of disturbance on threatened species are compounded by the risk that inappropriate disturbance may cause extinction of isolated populations, with limited or non-existent capacity for recolonisation. Metapopulation persistence in fragmented systems is an often delicate balance between the rates of population extinction and colonisation (Hanski 1999). Populations inhabiting fragments experience extinctions due to demographic and environmental stochasticity, with extinction risk being amplified in small patches which cannot support large populations and are hence more likely to experience extinction due to demographic stochasticity. These risks are exacerbated in patches with poor or declining habitat quality (Hanski et al. 1996; Franken and Hik 2004; Ferraz et al. 2007). Under equilibrium dynamics, these local extinctions are offset by patch colonisation, driven proximately by dispersal from neighbouring extant populations, and ultimately by sufficiently small inter-patch distances and matrix permeability to migrants (Ricketts 2001; Gaggiotti et al. 2004; Cosentino et al. 2012). However, such dynamic equilibria are fragile. For example, where ‘source-sink’ dynamics are prevalent, in which some key populations represent sources of recruits and migrants across the network (Pulliam 1988), loss of source populations can quickly send entire metapopulations towards collapse (Moilanen et al. 1998). Disturbance events clearly carry great risk for fragmented populations when they affect key source populations, even if those disturbance events are ultimately fundamental to broader metapopulation processes (McCarthy and Lindenmayer 1999).
The ecological role and management of disturbance for fragmented populations thus remains of considerable interest to applied ecologists. Perturbations such as fire, storms, grazing and drought are important drivers of ecosystem function in fragmented landscapes (Ross et al. 2002; Turner 2005; Laurance and Curran 2008). They are vital for metapopulation persistence for some species (Jäkäläniemi et al. 2006; Templeton et al. 2011) and detrimental for others (Lindenmayer and Possingham 1995). Likewise, both positive and negative effects of disturbance have been demonstrated for species inhabiting fragmented landscapes, resulting from a reliance on disturbance regimes to maintain key resources, but with thresholds in the scale, intensity and frequency of disturbance beyond which those mechanisms break down (Christian et al. 1997; Snäll et al. 2005).
Here, we use 9 years of occupancy data for an Australian grassland reptile, the Striped Legless Lizard (Delma impar), to investigate the role of disturbance in the persistence of fragmented populations of this species. Native temperate grassland was widespread in south-eastern Australia at the time of European settlement (c. 180 years BP), but has been systematically destroyed and degraded through intensive livestock grazing, cultivation, invasion by exotic plants and urban development (Lunt and Morgan 2002; Williams 2007). Only a small fraction of the original extent of this vegetation type remains, chiefly in the form of isolated remnants, most of which are not formally reserved or managed for conservation. Destruction of remnant grassland continues despite widespread recognition of the highly threatened status of this ecosystem, and the existence of legislative and regulatory frameworks for grassland protection (Williams et al. 2005b; Williams 2007).
The extent of historic destruction and degradation of native temperate grassland in south-eastern Australia has lead D. impar and other grassland specialists to undergo profound reductions in their extent of occurrence (Smith and Robertson 1999). Numerous grassland specialists are now listed as threatened, of which D. impar is one: the species is endangered in Victoria (DSE 2013) and vulnerable to extinction nationally (DEE 2016). In addition to habitat loss and fragmentation, inappropriate disturbance regimes—particularly fire and grazing frequency—may adversely affect population persistence of D. impar (Smith and Robertson 1999; Howland et al. 2014). However, formal, quantitative assessment of management requirements for ensuring population persistence (including disturbance regimes) have not been conducted.
We collected occupancy data for D. impar from 291 grassland sites across western Victoria, Australia, between 2004 and 2013. Availability of disturbance- and fragmentation-related variables for these sites allowed us to test hypotheses concerning the influence of these factors on the species’ probabilities of local extinction and colonisation. In turn, our long-term, spatially-extensive dataset allows us to elucidate the relative contribution of disturbance regimes and fragmentation to the occupancy dynamics of D. impar, and to identify management regimes needed to conserve population networks of this threatened species in fragmented grassland ecosystems.
Materials and methods
Study area and sites
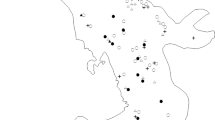
Native grassland in south-western Victoria is primarily associated with either the Western Volcanic Plain (a region of volcanic clay and basalt surface rock dating from eruptions and lava flows ending in the Late Quaternary) or alluvial plains to the north of the Western Volcanic Plains that also have clay substrates. This study was conducted across three Catchment Management Authority (CMA) areas that encompass most of south western Victoria’s remnant grassland: Glenelg-Hopkins, Corangamite and Wimmera (Fig. 1). Our sites were selected from the available extent of native grassland habitat across this region, as identified using aerial imagery, GIS layers of modelled grassland extent (see below) and ground reconnaissance. Sites included both publicly and privately-owned land and were subject to a range of land management practices. They were located on roadside and railway reserves, livestock grazing properties and formal conservation reserves.

Map of the study sites in Victoria, Australia, showing locations where Delma impar was detected and not detected during the study
Surveys
We established an array of 50 artificial cover objects (terracotta roofing tiles) at each site to survey for D. impar. The species regularly shelters under natural surface objects such as exfoliated basalt rocks and is also known to readily utilise artificial cover objects (O’Shea 2005; Peterson and Rohr 2010; Howland et al. 2016). As such, tile surveys are now the primary survey technique for the species. Tiles at each site were deployed in a regular grid pattern (5 by 10 tiles) spaced at 5 m intervals. For the purposes of our study, a site was defined as including all the area covered by the tile grid (25 m × 50 m), but would also encompass a short distance beyond that extent, within which any lizards that were present could be expected to encounter the grid of tiles and hence be at least potentially available to be detected. Grids were left in place for the duration of the study, with broken or missing tiles replaced as required.
Surveys involved visiting a site and lifting each tile in turn. They were undertaken between September and May in each survey year, coinciding with the Austral spring, summer and autumn. All vertebrate species encountered beneath tiles were identified. In addition to live individuals of D. impar, occasionally other evidence of the presence of the species was obtained, in the form of sloughed skins or eggs. Where sloughed skins and eggs could be identified as belonging to D. impar with certainty, they were treated as evidence of species presence. Sloughed skins were removed when detected to avoid double-counting during subsequent surveys at the same site. The following variables were also recorded during each tile checking event: Julian date, time of day, air temperature and under-tile temperature. It was considered likely that each of these variables could influence the probability of detecting D. impar at sites where the species was present (see below).
A total of 4672 surveys were conducted at the 291 survey sites between 2004/2005 and 2012/2013 survey seasons (Table 1). Survey data were collated by temporal periods from 1 July in each calendar year to 30 June in the following year. Survey effort, both in the number of sites surveyed and the number of surveys per site, varied substantially between years depending on available resources. Between 65 and 281 sites were surveyed in any given year (Table 1), with at least 217 sites being surveyed in all but the first and last years of the study. The mean number of surveys per site also varied substantially between years, from a minimum of 1.39 surveys per site in 2004/2005 to a maximum of 3.63 per site in 2007/2008 (Table 1).
Disturbance, fragmentation and habitat variables
Two disturbance processes—fire and grazing by livestock—were hypothesised to be particularly important determinants of occupancy dynamics of D. impar based either on existing knowledge of the lizard’s biology (Dorrough and Ash 1999; Smith and Robertson 1999; Howland et al. 2016) or the effects of these variables on the floristic and structural characteristics of native grasslands in south-eastern Australia (Morgan and Lunt 1999; Howland et al. 2014). We summarised the fire histories of each site based on the number of times each site was known to have been burnt during the study period (2004–2013) using records of the state fire-management authorities (Country Fire Authority and Department of Environment, Land, Water and Planning), our own observation of fire events and interviews with land managers. Detailed longer-term fire histories were unavailable for most sites, so the extent to which current fire regimes are representative of historic fire regimes is unknown. However, in the absence of any major shift in the management of these sites in recent decades, we assume that fire frequencies during the study period were similar to those in the recent past.
Grazing of these sites is mainly by domestic Sheep (Ovis aries) and to a much lesser extent Cattle (Bos taurus), along with invasive European Rabbits (Oryctolagus cuniculus) and native Eastern Grey Kangaroos (Macropus giganteus). In the absence of data on herbivore densities and grazing intensity, frequency of grazing during the study was scored on a simple ordinal scale for each site, using our own observations and interviews with land managers. Score were: complete or near-complete absence of grazing animals (0); occasional, light grazing (1); frequent grazing, but with fallow periods (2), and; constant grazing (3). Note that grazing of these sites was never sufficiently intense to completely modify grassland floristics or destroy tussock structure and should be considered moderate grazing pressure in the broader agricultural and ecological context. To enable interpretation of these scores, pictorial representations of vegetation structure and composition under these grazing frequencies are provided in the Supplementary Material, along with explanation of the perceived role of grazing in determining these vegetation attributes.
Surrounding habitat connectivity for D. impar was measured at each site with the aid of a GIS layer of the predicted extent of native grassland in Victoria developed by the Department of Environment, Land, Water and Planning using floristic databases, expert knowledge and statistical modelling of bioclimatic and remote-sensed variables. On the basis of the limited dispersal capacity of D. impar (Maldonado et al. 2012), we hypothesised that the predicted extent of native grassland habitat within a 1000 m radius surrounding each site would represent a useful surrogate of habitat connectivity, and its inverse, habitat fragmentation. While D. impar are known to inhabit other vegetation types, including degraded native grasslands and non-native pastures (Dorrough and Ash 1999) their association with native grassland is well-established. On this basis we hypothesised that the extent of this vegetation type in the vicinity of our sites would be a satisfactory, if imperfect surrogate for habitat availability.
In addition to the above measures of disturbance and habitat connectivity, we hypothesised that one additional habitat variable may help to explain the occupancy dynamics of D. impar in the study area. Published accounts commonly refer to usage of soil cracks as shelter by D. impar (O’Shea 2005), and associations between the presence of the D. impar and cracking clay-rich soils are also reported (Dorrough and Ash 1999). Hence, as a surrogate for the availability of soil cracks we obtained measures of soil-clay content at each site using a GIS layer of this variable developed by the Australian Collaborative Land Evaluation Program (ACLEP), endorsed through the National Committee on Soil and Terrain (http://www.asris.csiro.au/themes/NationalGrids.html).
Statistical analysis
A dynamic occupancy model with imperfect detectability (MacKenzie et al. 2003) was used to model the processes of population extinction and colonisation among the study sites over the 9-year survey period. We used a Bayesian state-space formulation of this model, as described by Royle and Kéry (2007). The approach treats the occupancy state of each site during year t (denoted \( {\mathbf{Z}} = z_{i,t} \)) as imperfectly-observed, using repeat survey data to parameterise an integrated model of the probabilities of initial occupancy, population persistence, colonisation and detection conditional on presence. A state-space formulation was chosen as it explicitly infers the probabilities of occupancy of each site at each time-step, conditional upon the observed data, the model and the priors (see below). Thus, it is possible to use the model to make explicit probability statements about the occupancy states of either individual sites or aggregated inferences about the occupancy of groups of sites at any given time (Royle and Kéry 2007).
At a given site i, and year t, the probability that the site is occupied is described by a Bernoulli process, with parameter \( \psi_{i,t} \):
The process model for annual changes in the occupancy status of the sites can be described by the following equation (Royle and Kéry 2007):
where \( \psi_{i,t} \) is the probability of occupancy at the ith site at time t, \( \phi_{i,t} \) is the probability that the ith occupied site remains occupied between times t − 1 and t, and \( \gamma_{i,t} \) is the probability that the ith unoccupied site is colonised between times t − 1 and t.
The initial probability of occupancy at each study site \( \left( {\psi_{i,1} } \right) \) was modelled as a function of both disturbance variables (fire frequency [firei] and grazing frequency [grazei]) as main effects, an interaction term between them, the extent of remaining native grassland habitat in the surrounding landscape (grassi, representing connectivity) and the mapped percentage of clay soil content (clayi). Hence, the initial probability of occupancy at each study site (\( \psi_{i,1} \)) was modelled as:
where β1 is an intercept term and the remainder are regression coefficients.
We hypothesised that instances of local extinction at our study sites would occur either as a consequence of prevailing fire and grazing regimes (including their interaction) or stochastic demographic processes relating to small population sizes and/or population isolation, resulting in a little or non-existent immigration, and therefore limited capacity for demographic ‘rescue’ by neighbouring populations (Brown and Kodric-Brown 1977). We assumed that our measure of connectivity (the extent of remaining native grassland habitat in the surrounding landscape) would be a useful surrogate for both local population size and immigration rate. With the addition of the mapped percentage of clay soil content, the annual probability of population persistence at each study site (\( \phi_{i,t} \)) was modelled as:
Attempts to integrate quadratic effects of both grazing and fire on the probability of persistence, to allow for the possibility that intermediate fire frequencies or grazing scores optimised persistence were undermined by poor model convergence, although initial model fitting did not indicate any clear quadratic relationships between these variables and persistence.
The annual probability of colonisation of vacant sites was modelled as function of connectivity alone. With few colonisation events observed, we were unable to pursue effects of fire, grazing and clay soil content on colonisation, despite such effects being plausible. Hence, the annual probability of colonisation of vacant study sites (\( \gamma_{i,t} \)) was simply modelled as:
Turning to the observation component of the model, the probability of detecting D. impar during a single survey was described by a Bernoulli process with parameter \( p_{j} \), conditional on the site being occupied during the year of the survey \( \left( {z_{i,t} = 1} \right) \):
Spatio-temporal variation in the probability of detecting D. impar was modelled as a function of survey-level covariates. The detection model allowed for two distinct forms of detection—detection of live lizards or detection of either sloughed skins or eggs—with different covariates and differing seasonal patterns of detection. In both cases, the annual seasonal trend in detection probability was modelled using a series of sine and cosine terms (a second-order Fourier series; Crespin et al. 2002; Heard et al. 2015). The influence of hour-of-day \( \left( h \right) \) and under-tile temperature \( \left( T \right) \) on the probability of detection was modelled using both linear and quadratic terms to allow for possible curvature in the relationships between these variables and the probability of detection. We also hypothesised that lizards may be more likely to use tiles when temperatures below them were warmer than the ambient temperature, but that this effect would be moderated at higher air temperatures. Therefore, we included the difference between the under-tile and air temperature (ΔT), and the interaction of this difference with the under-tile temperature as predictors of detection.
Therefore, for lizards, the probability of detection \( \left( {p_{j}^{liz} } \right) \) was modelled as:
For sloughed skins and eggs, the probability of detection \( \left( {p_{j}^{indirect} } \right) \) was modelled as a function of Julian date only:
Prior to analysis, Julian dates were transformed to the interval [0, 1], times of day (measured in hours) were centred around midday, and under-tile temperatures were centred around the mean value of 22 °C.
Assuming the two methods of detection were independent, we calculated the overall probability of detection \( (p^{total} ) \) when both methods of detection are applied under the same survey conditions (date, time, temperature, etc.) as follows:
\( p^{total} = 1 - [(1 - p^{indirect} )(1 - p^{liz} )] \)
To summarize the changes in occupancy across all M = 291 sites over the course of the study, we inferred the number of occupied sites at each time step [\( N_{t} \), see Royle and Kéry (2007)] from the latent occupancy states \( {\mathbf{Z}} \) of the state-space model as follows:
Similarly, the number of colonisation (\( {\text{col}}_{t} \)) and extinction (\( {\text{ext}}_{t} \)) events at each time step, and the net change in occupancy (\( \Delta N_{t} \)), was calculated as follows:
The integrated model of occupancy, persistence, colonisation and detection was fitted to the data using Bayesian Markov Chain Monte Carlo (MCMC) methods. Fitting of the model was carried out using JAGS version 4.3.0 (Plummer 2017). Convergence of the MCMC algorithm was assessed using the Brooks-Gelman-Rubin statistic (Brooks and Gelman 1998) and by visual inspection of the chain-histories of selected parameters. A burn-in period of 4000 iterations of the MCMC algorithm was used to ensure convergence. An additional 4000 iterations were then generated from each of 4 chains, retaining every 2nd iteration for inferences regarding the joint posterior distribution of the parameters and related derived quantities. Vaguely informative Cauchy (0, 2.5) priors were specified for the regression parameters of the model (Gelman 2006; Gelman et al. 2008). Data and code necessary to reproduce the analysis are provided in Scroggie et al. (2019).
Results
Occupancy, persistence and colonisation
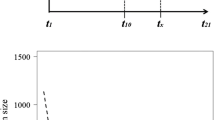
With extinctions of D. impar populations outstripping colonisations during this study, the predicted number of occupied sites declined, from 65 (95% CI 55, 77) in the first year, to 47 (95% CI 39, 56) in the final year (Table 1; Fig. 2). This represents an overall loss of 18 (95% CI 4, 32) occupied sites over the 9 years of monitoring (a decline of 27%). The net change in occupancy varied among years (and had high uncertainty; Fig. 2) but averaged a loss of 2 (95% CI 0.44, 3.6) sites per annum.

a Inferred number of sites occupied by D. impar over the course of the study. The solid line is the posterior mean prediction, and shaded areas the 75% (dark) and 95% (light) credible intervals. The dashed line denotes the number of sites where D. impar was actually observed in each year. b Inferred numbers of extinctions in each year. c Inferred number of colonisations in each year. d Inferred annual net change in the number of occupied sites
Disturbance was an important driver of occupancy dynamics, but evidently had both positive and negative relationships with persistence of D. impar populations. Main effects of fire and grazing frequency were positive for both the probability of initial occupancy and the annual probability of population persistence (Table 2; Fig. 3). Although the 95% CI for the effect of fire frequency on the probability of persistence overlapped zero (95% CI − 0.065, 0.20), this variable had an unequivocally positive effect on the probability of initial occupancy (95% CI 0.13, 0.48; Table 2; Fig. 3). Grazing frequency had clear positive effects on both probability of initial occupancy and the annual probability of persistence, with 95% CIs > 0 in both cases (Table 2; Fig. 3). However, there was a clear negative effect of the interaction of fire and grazing on the annual probability of persistence (mean = − 0.43) with 95% CI that did not overlap zero (− 0.74, − 0.16; Table 2; Fig. 3).

Posterior densities of regression parameters relating the covariates of the dynamic occupancy model to a initial occupancy (ψ1), b annual probability of persistence (ϕ), and c annual probability of colonization (γ) for populations of D. impar in remnant grassland
Habitat connectivity, as measured by the predicted extent of native grassland habitat within a 1000 m radius surrounding each site, had a positive, although uncertain, effect on the probability of initial occupancy by D. impar (mean = 0.23, 95% CI − 0.23, 0.69; Table 2; Fig. 3). Interestingly, connectivity had essentially no effect on the annual probability of population persistence (counter to our rescue-effect hypothesis), but had a positive effect on the annual probability of colonisation as expected (mean = 0.34, 95% CI 0.0014, 0.63; Table 2; Fig. 3). Soil clay content, a surrogate for the availability of soil cracks for D. impar, was positively related to the probability of initial occupancy (mean = 0.35, 95% CI − 0.044, 0.76), but essentially unrelated to the annual probability of persistence (Table 2; Fig. 3).
Relationships between the probabilities of initial occupancy, persistence and colonisation for D. impar and the above covariates are shown in Fig. 4. The probability of initial occupancy is highest with high surrounding grassland cover, high soil clay content and either relatively high fire or grazing frequency (but not both). Likewise, the annual probability of persistence is maximised when grazing frequency is high but fire frequency is low, or when fire frequency is high but grazing frequency is low. It falls to < 0.25 when fire and grazing frequency both approach their maximums, regardless of the other covariates included in the model. The annual probability of colonisation by D. impar remains low across the spectrum of surrounding grassland extent, rising from ~ 0 with no surrounding grassland to ~ 0.2 when the entire surrounding landscape consists of grassland.

Predicted effects of covariates on a the probability of initial occupancy, b the probability of persistence and c the probability of colonisation by D. impar in remnant grassland. Solid and dashed contour lines in b denote annual probabilities of persistence of 0.5 and 0.25 respectively. The solid line in c is the posterior mean prediction, and shaded areas the 75% (dark) and 95% (light) credible intervals. ‘Grazing’ ranges from 0 (complete to near-complete exclusion of grazing herbivores) to 3 (constant grazing). ‘Fire’ is the number of fires known over the course of the nine-year monitoring period
Detection
Detection probabilities for lizards and their sign (sloughed skins and eggs) displayed clear but contrasting seasonal cycles (Fig. 5). For lizards, there was a distinct peak in detection probability between September and November (the Austral spring), with progressively lower detection probabilities through summer and autumn. Detection probabilities of sloughed skins and eggs were highest in mid-summer (December–January), dropping to very low levels between winter and early spring (June–September). Combined, the probabilities of detection for D. impar by any method peaked during spring and progressively decreased through the summer and autumn (Fig. 5).

a Inferred seasonal trends in probabilities of detection for lizards (dotted line), sloughed skins and other indirect evidence (dashed line), and combined, overall probabilities of detection (solid line) for single surveys using both sources of detection for D. impar. Detection probabilities for lizards are predicted assuming that surveys are conducted at noon, with a tile temperature of 22°C and no differential between tile and air temperatures. Shading denotes the 95% credible intervals on the predictions. b Relationship between air temperature and air-tile temperature differential, and the estimated probability of detection (p) for lizards. The solid horizontal line denotes equality of air temperature and tile temperature. The dashed contour lines delimit the thermal conditions where probability of detection is > 0.6. The predictions are for tile surveys conducted on 1 November at midday
Time of day had very little effect on probabilities of detecting lizards (Table 3). Main effects of under-tile temperature and the differential between air temperature and under-tile temperature were positive; however, there was a strongly negative effect of the interaction between under-tile temperature and temperature differential (Table 3). Probabilities of detection were highest when air temperatures were low (< 20 °C) but under-tile temperatures were higher (by up to 10 °C), in which case tiles provided warm microclimates during otherwise mild conditions (Fig. 5). Detection became less likely when air temperatures exceeded 35 °C, particularly when under tile temperatures where higher than air temperatures (Fig. 5). However, if under tile temperatures were significantly lower than air temperature under these conditions (e.g., 5 °C lower), detection probability increased (that is, when tiles provided refuge from high environmental temperatures; Fig. 5).
Discussion
Interacting effects of disturbance, particularly those that entail both positive and negative impacts for threatened species, serve as a reminder of the complex challenges faced by conservation managers. Our study provides evidence that fire and grazing are factors associated with persistence of populations of the threatened Striped Legless Lizard (D. impar), but the species is sensitive to the interaction of these disturbances. Across the range of fire and grazing frequency observed in our focal grassland remnants, we show that fire and grazing are positively correlated with the persistence of D. impar populations. However, populations facing both disturbance regimes intensively and concurrently have a high risk of local extinction. As such, our study points to a fragile balance in the spatial and temporal occurrence of these disturbance events for the persistence of population networks of D. impar: maintaining some burning or grazing can promote metapopulation persistence, whereas burning and grazing that is widespread and simultaneous may quickly drive population networks to regional extinction.
Our study contributes to the global literature demonstrating the importance of maintaining asynchrony in disturbance events for the persistence of fragmented populations (e.g. Harrison and Quinn 1989; Hanski et al. 1995; McCarthy and Lindenmayer 1999; Kallimanis et al. 2005; Liao et al. 2015). Species with naturally fragmented populations, particularly those with relatively high rates of population extinction and colonisation, are often adapted to spatially and temporally asynchronous disturbance (Hanski 1999). Despite the potential for significant short-term impacts (including local extinctions), disturbance events can increase long-term habitat suitability through their effects on habitat structure, food resources, microclimate, competitors and natural enemies (Thomas and Jones 1993; Moilanen and Hanski 1998; Jäkäläniemi et al. 2006; Templeton et al. 2011). As such, asynchronous disturbance events can be vital for long-term persistence of many naturally fragmented populations (Hanski 1999). The same is true for metapopulations derived from human-induced landscape change. Disturbance is pervasive in such systems, often facilitated by fragmentation itself (Lindenmayer and Fischer 2006). As for naturally fragmented populations, when disturbance events entail short-term risks to population persistence (even if they increase habitat suitability in the long-term), network persistence can only be achieved when there is adequate spatial and temporal asynchrony in disturbance regimes (Hanski 1999).
Of course, sufficient between-patch dispersal and network connectivity is fundamental to these dynamics. Where fragmentation constrains inter-patch dispersal to the extent that patch colonisation following local extinction is unlikely, metapopulations subject to disturbance will decline inexorably to extinction (Hanski 1999). Local extinctions resulting from the short-term impacts of disturbance cannot be offset, nor can the value of disturbance events be realised where they deliver long-term benefits for habitat suitability. Moreover, dispersal rates can themselves be tied to disturbance, in which case management of disturbance regimes can be fundamental to maintaining connectivity. Eastern Collared Lizards (Crotaphytus collaris collaris) in the Ozarks of eastern Missouri provide a classic example, for which forest fires not only maintain the open, rocky glades on which the species depends, but also facilitate dispersal between glades by increasing permeability of the surrounding forest (Templeton et al. 2011). Clearly managers face difficult decisions under circumstances such as these, in which they must balance the frequency, intensity and synchrony of disturbance events to mitigate short-term impacts on relic populations, facilitate long-term maintenance of habitat suitability, and encourage between-patch dispersal when disturbance increases landscape permeability.
We suspect this is the situation facing managers of our study species, D. impar. Here, we have shown that the interaction of fire and grazing has significant negative effects on population persistence for this species, yet some fire and grazing can be beneficial in isolation (Fig. 3). We also show that colonisation rates for this species are low, but increase with increasing grassland extent in the surrounding landscape (Fig. 4). Grassland edges in our study system can be diffuse; hence, it is likely that the factors driving habitat suitability within remnants also drive those outside remnants, in which case fire and grazing may also contribute to maintenance of connectivity between remnants. Given the limited dispersal ability and ‘slow’ life-history of D. impar, reliance on recolonisation of vacant habitat as a mechanism of compensating for local extinctions is likely to be a risky management strategy. Managers should instead seek to manage habitat in a way that minimises risks of patch-level extinctions, which will necessarily include the careful application of fire or grazing.
Turning to the specific mechanisms underpinning associations between fire, grazing and habitat suitability for D. impar, we hypothesise that the loss of structural and floristic diversity in grasslands that remain both unburnt and ungrazed drives the observed positive relationship between disturbance and lizard population persistence. Prior to European colonisation, the native grasslands of south-eastern Australia were subject to frequent fire caused either by lightning strikes or deliberate ignition by indigenous people, and subject to grazing by a diverse suite of native herbivores (Edney et al. 1990; Crowley and Kershaw 1994). Fire and grazing continue to play a prominent role in structuring native grassland ecosystems in south-eastern Australia (Lunt and Morgan 2002), and these ecosystems are susceptible to native species loss and exotic species accrual in the absence of these disturbance regimes (Tremont and Mcintyre 1994; Williams et al. 2005a; Lunt et al. 2007; Morgan 2015). The result can be reductions in both the structural and floristic diversity of grassland remnants with the removal of fire and grazing, particularly in long-grazed sites in which exotic flora is well established (Tremont and Mcintyre 1994; Williams et al. 2005a; Lunt et al. 2007; Morgan 2015; Supplementary Material). Although at least one study has produced equivocal results with regard to associations between D. impar and vegetation attributes (Dorrough and Ash 1999), structural and floristic diversity (particularly of tussock-forming grasses) has long been considered a key determinant of habitat suitability for D. impar (Smith and Robertson 1999; O’Shea 2005). Recent studies confirm positive relationships between this attribute and site occupancy by D. impar (Howland et al. 2014, 2016). Structurally and floristically diverse grassland remnants are likely to have higher abundance of the invertebrate prey of D. impar (Kutt et al. 1998), and offer a superior mix of both shelter and basking sites (Kutt 1992). Heavy accumulations of plant biomass may also occlude soil cracks, which our study and others (Dorrough and Ash 1999; O’Shea 2005) suggest are an important component of habitat quality for this species.
Logically, any promotion of structural and floristic diversity of grassland remnants by fire and grazing will be mediated by their frequency, intensity and coincidence. Very frequent or intense fires, and/or heavy livestock grazing, could eliminate structural and floristic diversity of grasslands on which the lizard populations rely. If our hypothesis holds that the positive effects of fire and grazing demonstrated here for D. impar relate to the maintenance of structural and floristic diversity of grassland remnants in south western Victoria (most of which have been grazed for many decades and are susceptible to exotic domination), we may also hypothesise that the structural and floristic diversity of grassland remnants declines sharply as the coincidence of these two disturbances increases (given that our data demonstrate a strong negative effect of the interaction of fire and grazing on population persistence by D. impar). In particular, we hypothesise that tussock-forming perennial grasses, with which D. impar is particularly associated (Kutt 1992; Howland et al. 2014, 2016), may be exterminated by concurrent and repeated bouts of fire and grazing. Tussock-forming perennials in south-eastern Australian grasslands are relatively robust to grazing (Dorrough and Scroggie 2008) and readily re-sprout after fire (Morgan and Lunt 1999). However, the combination of frequent grazing and fire produces significant physiological stress for these plants (Tremont and Mcintyre 1994) and could lead to widespread mortality. As post-fire recruitment of exotic annual grasses can be extremely high in southern Australian grasslands (Lunt 1990), state-transition to exotic domination could result from intense bouts of both fire and grazing.
Our study reveals that populations of D. impar in western Victoria continue to decline, with a predicted 27% reduction in occupancy among our sites over a 9-year period. Stabilising and recovering remnant populations of this threatened species will require a focus on maintaining suitable fire and grazing regimes, in concert with retention and restoration of grassland habitat to facilitate connectivity and bolster (re)colonisation rates. As above, our study suggests that fire and grazing may be effective tools for maintaining the structural and floristic requirements of these lizards (see also the Supplementary Material). However, we caution that grazing applied at a frequency and/or intensity that eliminates native perennial tussock-forming grasses (in particular) is detrimental for D. impar, as has been shown elsewhere (Howland et al. 2014, 2016). Moreover, populations subject to concurrent fire and grazing are highly susceptible to extinction. We recommend development of an experimental approach to fire and grazing management for this species, building on previous work by O’Shea (2005). Experimental management should seek to identify optimal burning and grazing regimes to maintain the floristic and structural attributes of grassland flora required by D. impar, the contextual and ecological factors underpinning these relationships, and the spatial and temporal asynchrony in disturbance among sites required to ensure metapopulation persistence. With regard to context, we recommend that livestock grazing should only be used at sites where it is already the primary management tool, as the introduction of livestock to ungrazed grasslands can compact soils, increase nutrient inputs and facilitate weed incursion (Wong and Morgan 2007; Supplementary Material). At sites where fire is already being applied, it should remain the preferred management tool when disturbance is required.
At the landscape scale, retention and restoration of grassland habitat to facilitate connectivity for D. impar will require significant conservation investment. Further loss of remnant grassland habitat will exacerbate the ongoing decline of this species. As such, we recommend improved protection and management of remnant grassland throughout the species’ range as the highest priority conservation action for this species. Opportunities to expand grassland habitats should also be pursued where feasible. Native temperate grasslands in southern Australia are difficult to restore; however, significant recent advances have been made, drawing upon the now extensive literature on the ecology of this ecosystem (Williams et al. 2015). This includes successful efforts directed at conserving another grassland-affiliated legless lizard in southern Australia (McDougall et al. 2016). We encourage further investment in grassland restoration projects across southern Australia, both for their benefits for D. impar and wider contribution to conserving this critically endangered ecosystem and its distinctive biota.
For some isolated and unoccupied habitat remnants, the technical feasibility of translocating D. impar should also be explored. Such efforts should be restricted to areas of high suitability for the species, where persistence of translocated populations is likely in the long term. As our model allows the probability of persistence to be predicted from habitat and disturbance variables, it could be used to prioritise sites for translocation. We strongly encourage translocation trials for this species; however, these initiatives must be additional to, not replacements for, investment in the retention and management of remnant populations. The relatively high costs and generally low success rates of translocation programs for threatened reptiles (Dodd and Seigel 1991; Germano and Bishop 2009) means they should be seen as a secondary, and unproven conservation priority.
For each of the conservation initiatives listed above, rigorous monitoring programs that allow their success or failure to be assessed will be vital. Our study provides a sound basis for designing those monitoring programs. Our data demonstrate the value of artificial cover objects for monitoring D. impar and show that detection rates using this method are maximal when cover objects are checked during late spring and early summer, when temperatures are mild and when under-tile temperatures are warmer than ambient. These results allow survey effort to be optimised both seasonally and meteorologically to maximise detection probabilities, and thereby maximise statistical power to assess occupancy trends (Field et al. 2005; Guillera-Arroita and Lahoz-Monfort 2012).
Thus, to conclude, we see four main priorities for conservation efforts directed at D. impar: (i) experimental management of disturbance regimes that seeks to maximise habitat occupancy by maintaining the required floristic and structural attributes of grassland flora; (ii) significant investment in grassland protection and restoration to facilitate connectivity and re-expansion of this species; (iii) re-introduction trials within isolated but otherwise high-quality grassland remnants, and; (iv) rigorous monitoring programs that maximise detection probabilities and statistical power to critique and iteratively revise these management interventions. Collectively, these initiatives have the potential to reverse the ongoing decline of this cryptic grassland specialist.
References
Beissinger SR (2002) Population viability analysis: past, present and future. In: Beissinge SA, McCullough DR (eds) Population viability analysis. The University of Chicago Press, Chicago, pp 5–17
Brawn JD, Robinson SK, Thompson FR (2001) The role of disturbance in the ecology and conservation of birds. Annu Rev Ecol Syst 32:251–276
Brooks SP, Gelman A (1998) Alternative methods for monitoring convergence of iterative simulations. J Comput Gr Stat 7:434–455
Brown JH, Kodric-Brown A (1977) Turnover rates in insular biogeography: effect of immigration on extinction. Ecology 58:445–449
Burgman MA, Lindenmayer DB (1998) Conservation biology for the Australian environment. Surrey Beatty and Sons, Chipping Norton
Christian S, Michael R, Volker G, Christian W (1997) Modelling persistence in dynamic landscapes: lessons from a metapopulation of the grasshopper Bryodema tuberculata. J Anim Ecol 66:508–518
Cosentino BJ, Phillips CA, Schooley RL, Lowe WH, Douglas MR (2012) Linking extinction–colonization dynamics to genetic structure in a salamander metapopulation. Proc R Soc B 279:1575–1582
Crespin L, Verhagen R, Stenseth NC, Yoccoz NG, Prévot-Julliard A-C, Lebreton J-D (2002) Survival in fluctuating bank vole populations: seasonal and yearly variations. Oikos 98:467–479
Crowley GM, Kershaw AP (1994) Late quaternary environmental change and human impact around Lake Bolac, western Victoria, Australia. J Quat Sci 9:367–377
DEE (2016) Striped legless lizard (Delma impar): conservation advice. Department of Environment and Energy, Canberra
Dodd CKJ, Seigel RA (1991) Relocation, repatriation, and translocation of amphibians and reptiles: are they conservation strategies that work? Herpetologica 47:336–350
Dorrough J, Ash JE (1999) Using past and present habitat to predict the current distribution and abundance of a rare cryptic lizard, Delma impar (Pygopodidae). Aust J Ecol 24:614–624
Dorrough J, Scroggie MP (2008) Plant responses to agricultural intensification. J Appl Ecol 45:1274–1283
DSE (2013) Advisory list of threatened vertebrate fauna in Victoria. Department of Sustainability and Environment, Melbourne
Edney PA, Kershaw AP, De Deckker P (1990) A late Pleistocene and Holocene vegetation and environmental record from Lake Wangoom, Western Plains of Victoria, Australia. Palaeogeogr Palaeoclimatol Palaeoecol 80:325–343
Ferraz G, Nichols JD, Hines JE, Stouffer PC, Bierregaard RO Jr, Lovejoy TE (2007) A large-scale deforestation experiment: effects of patch area and isolation on Amazon birds. Science 315:238–241
Field SA, Tyre AJ, Possingham HP (2005) Optimizing the allocation of survey effort under economic and observational constraints. J Wildl Manag 69:473–482
Franken RJ, Hik DS (2004) Influence of habitat quality, patch size and connectivity on colonization and extinction dynamics of collared pikas Ochotona collaris. J Anim Ecol 73:889–896
Gaggiotti OE, Brooks SP, Amos W, Harwood J (2004) Combining demographic, environmental and genetic data to test hypotheses about colonization events in metapopulations. Mol Ecol 13:811–825
Gelman A (2006) Prior distributions for variance parameters in hierarchical models (comment on article by Browne and Draper). Bayesian Anal 1:515–534
Gelman A, Jakulin A, Pittau MG, Su Y-S (2008) A weakly informative default prior distribution for logistic and other regression models. Ann Appl Stat 2:1360–1383
Germano JM, Bishop PJ (2009) Suitability of amphibians and reptiles for translocation. Conserv Biol 23:7–15
Guillera-Arroita G, Lahoz-Monfort JJ (2012) Designing studies to detect differences in species occupancy: power analysis under imperfect detection. Methods Ecol Evol 3:860–869
Gunderson LH (2000) Ecological resilience in theory and application. Annu Rev Ecol Syst 31:425–439
Hanski I (1999) Metapopulation ecology. Oxford University Press, Oxford
Hanski I, Pakkala T, Kuussaari M, Lei G (1995) Metapopulatin persistence of an endangered butterfly in a fragmented landscape. Oikos 72:21–28
Hanski I, Moilanen A, Pakkala T, Kuussaari M (1996) The quantitative incidence function model and the persistence of an endangered butterfly metapopulation. Conserv Biol 10:578–590
Harrison S, Quinn JF (1989) Correlated environments and the persistence of metapopulations. Oikos 56:293–298
Heard GW, Canessa S, Parris KM (2015) Interspecific variation in the phenology of advertisement calling in a temperate Australian frog community. Ecol Evol 5(18):3927–3938
Howland BAW, Stojanovic D, Gordon IJ, Manning AD, Fletcher D, Lindenmayer DB (2014) Eaten out of house and home: impacts of grazing on ground-dwelling reptiles in Australian grasslands and grassy woodlands. PLoS ONE 9:e105966
Howland BAW, Stojanovic D, Gordon IJ, Don Fletcher, Snape M, Stirnemann IA, Lindenmayer DB (2016) Habitat preference of the Striped Legless Lizard: implications of grazing by native herbivores and livestock for conservation of grassland biota. Aust Ecol 41:455–464
Hunter WC, Buehler DA, Canterbury RA, Confer JL, Hamel PB (2001) Conservation of disturbance-dependent birds in eastern North America. Wildl Soc Bull 29:440–455
Jäkäläniemi A, Tuomi J, Siikamäki P (2006) Conservation of species in dynamic landscapes: divergent fates of Silene tatarica populations in riparian habitats. Conserv Biol 20:844–852
Kallimanis AS, Kunin WE, Halley JM, Sgardelis SP (2005) Metapopulation extinction risk under spatially autocorrelated disturbance. Conserv Biol 19:534–546
Kutt AS (1992) Microhabitat selection and mobility of the Striped Legless Lizard Delma impar. University of Melbourne, Parkville
Kutt AS, Coulson G, Wainer J (1998) Diet of the Striped Legless Lizard Delma impar (Squamata: Pygopodidae) in a western (basalt) plains grassland, Victoria. Aust Zool 30:412–418
Laurance WF, Curran TJ (2008) Impacts of wind disturbance on fragmented tropical forests: a review and synthesis. Aust Ecol 33:399–408
Liao J, Ying Z, Hiebeler DE, Wang Y, Takada T, Nijs I (2015) Species extinction thresholds in the face of spatially correlated periodic disturbance. Sci Rep 5:15455
Lindenmayer DB, Fischer J (2006) Habitat fragmentation and landscape change: an ecological and conservation synthesis. Island Press, Washington
Lindenmayer DB, Possingham HP (1995) Modelling the impacts of wildfire on the viability of metapopulations of the endangered Australian species of arboreal marsupial, Leadbeater’s Possum. For Ecol Manag 74:197–222
Lunt ID (1990) Impact of an autumn fire on a long-grazed Themeda triandra (Kangaroo Grass) grassland: implications for management of invaded, remnant vegetations. Vic Nat 107:45–51
Lunt I, Morgan JW (2002) Grasslands of southeastern Australia. In: Bradstock RA, Williams JE, Gill MA (eds) Flammable Australia: the fire regimes and biodiversity of a continent. Cambridge University Press, Cambridge, pp 177–196
Lunt ID, Eldridge DJ, Morgan JW, Witt GB (2007) A framework to predict the effects of livestock grazing and grazing exclusion on conservation values in natural ecosystems in Australia. Aust J Bot 55:401–415
MacKenzie DI, Nichols JD, Hines JE, Knutson MG, Franklin AB (2003) Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 84:2200–2207
Maldonado SP, Melville J, Peterson GNL, Sumner J (2012) Human-induced versus historical habitat shifts: identifying the processes that shaped the genetic structure of the threatened grassland legless lizard, Delma impar. Conserv Genet 13:1329–1342
McCarthy MA, Lindenmayer DB (1999) Incorporating metapopulation dynamics of greater gliders into reserve design in disturbed landscapes. Ecology 80:651–667
McDougall A, Milner RNC, Driscoll DA, Smith AL (2016) Restoration rocks: integrating abiotic and biotic habitat restoration to conserve threatened species and reduce fire fuel load. Biodivers Conserv 25:1529–1542
Moilanen A, Hanski I (1998) Metapopulation dynamics: effects of habitat quality and landscape structure. Ecology 79:2503–2515
Moilanen A, Smith AT, Hanski I (1998) Long-term dynamics in a metapopulation of the American Pika. Am Nat 152:530–542
Morgan JW (2015) Biomass management in native grasslands. In: Williams NSG, Marshal A, Morgan JW (eds) Land of sweeping plains: managing and restoring the native grasslands of South-Eastern Australia. CSIRO Publishing, Collingwood, pp 201–222
Morgan JW, Lunt ID (1999) Effects of time-since-fire on the tussock dynamics of a dominant grass (Themeda triandra) in a temperate Australian grassland. Biol Conserv 88:379–386
O’Shea MB (2005) Methods for assessment and techniques for management of Striped Legless Lizard Delma impar populations in South-Eastern Australia. Victoria Univerisity of Technology, Melbourne
Peterson GNL, Rohr D (2010) Delma impar (Striped Legless Lizard) repeated use of communal nesting site. Herpetol Rev 41:78–79
Plummer M (2017) JAGS User Manual. Version 4.3.0 Available from https://sourceforge.net/projects/mcmc-jags/files/Manuals/4.x/jags_user_manual.pdf. Accessed 28 June 2017
Pulliam HR (1988) Sources, sinks and population regulation. Am Nat 132:652–661
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158:87–99
Ross KA, Fox BJ, Fox MD (2002) Changes to plant species richness in forest fragments: fragment age, disturbance and fire history may be as important as area. J Biogeogr 29:749–765
Royle JA, Kéry M (2007) A Bayesian state-space formulation of dynamic occupancy models. Ecology 88:1813–1823
Scroggie MP, Peterson GNL, Rohr DH, Nicholson E, Heard GW (2019) Code and data for: disturbance has benefits as well as costs for fragmented populations of a cryptic grassland reptile. Zenodo Digit Arch. https://doi.org/10.5281/zenodo.3259372
Smith WJS, Robertson P (1999) National recovery plan for the Striped Legless Lizard (Delma impar): 1999-2003. Unpublished report to Environment Australia, Canberra
Snäll T, Ehrlén J, Rydin H (2005) Colonization-extinction dynamics of an epiphyte metapopulation in a dynamic landscape. Ecology 86:106–115
Templeton AR, Brazeal H, Neuwald JL (2011) The transition from isolated patches to a metapopulation in the eastern collared lizard in response to prescribed fires. Ecology 92:1736–1747
Thomas CD, Jones TM (1993) Partial recovery of a Skipper Butterfly (Hesperia comma) from population refuges: lessons for conservation in a fragmented landscape. J Anim Ecol 62:472–481
Tremont R, Mcintyre S (1994) Natural grassy vegetation and native forbs in temperate Australia: structure, dynamics and life-histories. Aust J Bot 42:641–658
Turner MG (2005) Landscape ecology: what is the state of the science? Annu Rev Ecol Evol Syst 36:319–344
Williams NSG (2007) Environmental, landscape and social predictors of native grassland loss in western Victoria, Australia. Biol Conserv 137:308–318
Williams NSG, McDonnell MJ, Seager EJ (2005a) Factors influencing the loss of an endangered ecosystem in an urbanising landscape: a case study of native grasslands from Melbourne, Australia. Landscape and Urban Planning 71:35–49
Williams NSG, Morgan JW, McDonnell MJ, McCarthy MA (2005b) Plant traits and local extinctions in natural grasslands along an urban–rural gradient. J Ecol 93:1203–1213
Williams NSG, Marshal A, Morgan JW (eds) (2015) Land of sweeping plains: managing and restoring the native grasslands of South-eastern Australia. CSIRO Publishing, Collingwood
Wong N, Morgan JW (2007) Review of grassland management in South-eastern Australia. Parks Victoria technical series number 39. Parks Victoria, Melbourne
Acknowledgements
Funding was provided by the Victorian Department of Environment, Land, Water and Planning, and the Australian Commonwealth Government via the Corangamite, Glenelg-Hopkins and Wimmera Catchment Management Authorities. This work would not have been possible without the cooperation of numerous land managers who kindly provided access to the field sites. We thank the many staff and volunteers who collected data, especially Cath Grant, Donna McMaster, Lauren Koehler, Melinda Thompson, James Booth and Gemma Candy. David Ramsey, Michael Kearney and Ben Phillips provided helpful modelling advice, and Graeme Newell and Matt White kindly provided access to grassland spatial data. We thank Deirdre Lucas, Jim Thomson, Lindy Lumsden and Steve Sinclair for helpful comments on drafts of the manuscript. Our work was conducted under permits granted by the Department of Environment, Land, Water and Planning.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Scroggie, M.P., Peterson, G.N.L., Rohr, D.H. et al. Disturbance has benefits as well as costs for fragmented populations of a cryptic grassland reptile. Landscape Ecol 34, 1949–1965 (2019). https://doi.org/10.1007/s10980-019-00865-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00865-0