
Abstract
Context
Bumblebees are important pollinators for agricultural crops and wild plants. However, agricultural intensification and loss of semi-natural habitat may have adverse effects on colony performance. While mass-flowering crops may serve as food sources, landscapes dominated by intensive agriculture may be a poor bumblebee habitat compared to landscapes dominated by semi-natural habitat, such as forests and species-rich grasslands.
Objectives
We investigated the effect of different land use classes on bumblebee colony performance at two periods of time during the flying season. We expected mass-flowering crops to have a positive influence on colony performance during flowering. After mass-flowering, presence of semi-natural habitat was hypothesized to be essential for colony development.
Methods
We placed artificial colonies of Bombus terrestris in 28 apple and pear orchards during and after mass-flowering. We measured colony growth and took pollen samples from foraging workers. Causal relationships between land-use, pollen quality, amount of pollen collected and colony performance were analyzed.
Results
Contrary to our expectations, we found a negative correlation between orchard cover and amount of pollen collected in spring, during mass-flowering, resulting in a lower colony performance. During mass-flowering, colony performance and pollen diversity were positively correlated with cover of open semi-natural habitat, while after mass-flowering, colony performance was positively correlated with forest cover. After mass-flowering, pollen sources were more diverse and colony growth was related to land use classes at a larger spatial scale, indicating that bumblebees foraged further. We did not find a correlation between the performances of colonies at one specific site that were placed during and after mass-flowering.
Conclusions
Landscapes dominated by mass-flowering orchards represent inferior habitats for bumblebees during flowering. Heterogenous landscapes, containing a mix of various types of semi-natural habitat increase the habitat quality for bumblebees. We advise to maintain these diverse patches of semi-natural habitat, as mass-flowering crops alone are insufficient to support bumblebee colonies.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Bumblebees are important pollinators in farmlands, both for agricultural crops and wild plant species surrounding the crops (Stanley and Stout 2014). However, since the second half of last century, bumblebee populations experienced strong declines, caused by several drivers, of which habitat loss and agricultural intensification are probably the most important (Williams and Osborne 2009). Specific causes of this decline are a decrease in food resources (Gabriel and Tscharntke 2007; Holzschuh et al. 2008), degradation of elements that can provide nesting sites (Lye et al. 2009) and changing practices, such as the decreased use of leguminous fodder crops, such as red clover (Trifolium pratense L.) (Goulson et al. 2005; Scheper et al. 2014).
In temperate regions, bumblebee colonies persist throughout the spring and summer months, so they require a continuous source of pollen and nectar during this period to survive and to reproduce (Pywell et al. 2005). The absence of sufficient pollen sources seems to limit colony growth, especially late during the flying season (Fitzpatrick et al. 2007; Rundlöf et al. 2014). Bumblebee colonies require relatively large amounts of pollen, compared to solitary bee species (Rotheray et al. 2017), which makes them extra susceptible to low availability of foraging resources (Müller et al. 2006), resulting in a slower colony growht and reduced reproductive output.
Agricultural landscapes, despite often being poor in wild flowering plants (Carvell et al. 2006), can play an important role as foraging habitat for bumblebees, due to the presence of mass flowering crops, such as canola, potatoes or fruit trees (e.g. Westphal et al. 2003; Holzschuh et al. 2013). Additionally, patches of semi-natural habitat (SNH), including species-rich grasslands, forests, hedgerows and road verges, provide alternative foraging resources even when a crop is mass-flowering (Steffan-Dewenter and Tscharntke 1999; Croxton et al. 2002; Öckinger and Smith 2007). Many common bumblebee species are generalists, with colonies able to switch flower resources throughout the year. Due to their colonial lifestyle and large size, colonies of B. terrestris require large amounts of pollen and therefore often forage in large resource patches (Westphal et al. 2006; Wolf and Moritz 2008). Woody plants make up a large share of the collected pollen both during and after mass-flowering. Even individual flowering trees and shrubs often have large amounts of flowers, therefore serving as large pollen sources. Especially in spring, during mass-flowering, woody plants tend to be very important for bumblebees, as other species are not yet flowering (Kämper et al. 2016).
Earlier studies have already shown a link between landscape context and bumblebee colony performance (e.g. Hines and Hendrix 2005; Williams et al. 2012; Parmentier et al. 2013) or between pollen sources and colony performance (Génissel et al. 2002; Tasei and Aupinel 2008), but only few studies looked at both in field experiments (Kämper et al. 2016). Additionally, most research was carried out only within a short time frame, usually a few weeks, therefore ignoring the changes in floral resources during the active period of bumblebees, and possibly missing the effects of phenology of flowering plants on bumblebee colony performance.
The goal of this study was to investigate how Bombus terrestris (Linnaeus, 1758) colony performance relates to mass-flowering crops and their surrounding different land-use types. The experiment was carried out in two consecutive periods: (1) spring, during mass flowering of apple and pear trees, and (2) early summer, in the absence of these mass flowering crops. We anticipated the surrounding landscape to have a strong effect on colony growth, with semi-natural habitat (forest, semi-natural grassland) having a positive influence on colony growth by providing more diverse pollen sources than agricultural crops. We expected this effect to be strongest in summer, when the fruit trees in our study do not provide food resources and the agricultural land is not expected to support a large community of flowering plants that provide pollen and nectar. Additionally, we hypothesized pollen diversity to be highest at sites with more SNH.
Materials and methods
Study area
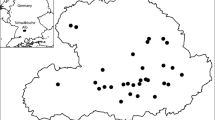
The study was carried out in the southeast of Flanders, Belgium, in a 45 × 20 km area (Fig. 1a). The study area has a temperate oceanic climate with mild winters. The region is the most important fruit-growing region of Belgium and is characterized by landscapes dominated by apple and pear orchards in the western part and cropland in the eastern part. Locally, larger forest fragments and patches of semi-natural grassland occur. The soils consist of loam in the southern half of the study region and of sandy loam in the northern half.

a Map of Belgium with the study area enclosed by the grey rectangle. The locations of the 28 bumblebee colonies are indicated by black dots. b Landscape around a single bumblebee colony. Circles show buffers with a radius of 250–2000 m around the colonies with 250 m intervals. Within these buffers the area of all land use classes was measured for further statistical analysis
Bumblebee colonies
The experiment was carried out in 2016, during two consecutive 6-week periods: the first time from the beginning of April to the end of May, during the mass-flowering of apple and pear orchards. The same setup was repeated from the beginning of June to the second half of July, to assess the effect of the land use on colony performance without the mass flowering in the orchards. 28 colonies of B. terrestris, reared at Biobest (Westerlo, Belgium), were placed in 28 different apple and pear orchards. Distance between colonies was at least 1250 m to reduce overlap in foraging range. To maximize the difference in surrounding landscapes, sites were selected to fit into four categories: landscapes dominated by apple and pear orchards, cropland, open semi-natural habitat (e.g. species rich grasslands, marshes, and heathland) and deciduous forest respectively (Online Appendix A).
To isolate the colonies against cold and moisture, styrofoam bee-coats® (Biobest, Westerlo, Belgium) were used. During mass-flowering, the initial average colony mass was 473.9 (S.D. 7.1) g, containing 25.7 (S.D. 4.3) workers (N = 28), while after mass-flowering, the colonies weighed on average 473.8 (S.D. 10.2) g, containing 22.6 (S.D. 5.6) workers (N = 28).
After 1 week all colonies were controlled for forager activity to assure that the colonies were actively foraging. During the rest of the experiment, the colonies were checked weekly and weighed every 2 weeks to measure the biomass increment. Colonies with a dead queen were removed and omitted from further analyses. In the first period of the experiment, one single colony was excluded due to the queen’s death, while during the second period, two colonies were omitted. After 6 weeks in each period, queen pupae appeared and the colonies were collected and weighed to assess colony growth. The total number of workers and queen pupae was counted to measure colony performance. As biomass increment was strongly correlated with number of workers and number of queen pupae (Online Appendix B), only biomass increment was analyzed further in general linear models.
Landscape
The land use was classified into five different land-use classes, using the Biological Valuation Map (BWK) (De Knijf et al. 2010) and the Flemish dataset of LPIS (ALV 2016): cropland (including temporary grasslands), orchards (including intensively managed apple, cherry orchards), deciduous forests, non-forest semi-natural habitat (including semi-natural grasslands, marshland, heathland, road verges) and urban (including gardens). The total cover of each land-use class was calculated within a radius ranging from 250 to 2000 m around each colony, with a 250 m increment (Fig. 1b), to include all possible foraging ranges mentioned for B. terrestris (Osborne et al. 1999; Walther-Hellwig and Frankl 2000; Knight et al. 2005; Wolf and Moritz 2008). Additionally, the total amount of semi-natural habitat was calculated in the same way, the latter being the sum of all forest and semi-natural grassland types. GIS-analyses were all carried out using QGIS 2.16 (QGIS Development Team 2015).
Pollen collection and plant source identification
Three weeks after the colonies were put in the orchards, pollen was sampled from foraging bumblebees. At each location, the nest entrance was closed and five workers returning to the nest were caught using an insect net. The pollen load of each bumblebee was visually scored on a 0–5 scale (0 = carrying no pollen; 1 = carrying small amounts of pollen, not forming a solid clump, 2 = pollen clumps small, covering less than half the tibial width, 3 = pollen clumps covering less than the whole tibial width, 4 = pollen clumps covering more than the full tibial width, 5 = pollen clumps covering more than double the tibial width). Cumulative pollen load (‘pollen quantity’) of the five trapped foragers was used as a measure of pollen availability.
To collect pollen from foragers, they were held with clean tweezers and the pollen clumps were removed from both corbiculae, using a clean insect pin. The pollen was stored in a dry eppendorf tube and frozen before further identification. All sampling took place between 10:00 and 16:00 under dry, sunny weather (Peat and Goulson 2005). The pollen samples were processed following the standard protocols, stained with basic fuchsin and mounted in glycerine (Moore et al. 1994). To achieve homogenization, pollen clumps were thoroughly mixed in the glycerine. Pollen were identified to the highest possible taxonomical resolution under × 1250 magnification with phase contrast (JENAMED, Carl Zeiss, Jena), using a reference collection of insect-pollinated plants, collected in the study region and Beug (2004) for identification. In each pollen slide, five straight transects with random starting point were followed, and on each transect, 50 pollen grains were identified, up to a total of 250 pollen grains per sample. The amount of each pollen type was multiplied by the pollen load of the sample and all pollen samples per colony were summed to estimate the composition of the collected pollen per colony. The Simpson diversity index of the total collected pollen was calculated and used in further analyses as a measure for pollen diversity. During the second run of the experiment, only nectar flights were observed in two colonies, which were omitted from analyses that include pollen composition.
Statistical analysis
All analyses were carried out using R 3.4.3 (R Core Team 2018). To achieve normality, colony biomass increment, number of queen pupae and number of workers were squareroot-transformed.
The colony performance during the two periods of the experiment was analysed separately. To assess the optimal spatial scale, a general linear model was created, with the total area of orchards, non-forest semi-natural habitat and forest as fixed variables and the biomass increment as response variable. This analysis was performed on all analysed scales from 250 to 2000 m, with 250 m increments, both during and after mass-flowering. The scale with the highest R2 was selected as the spatial scale to use in the analyses (Holland et al. 2004). During mass-flowering, the landscape composition at a scale of 750 m had the strongest influence on colony biomass increase, while after mass-flowering, it was strongest at a 1000 m radius (Online Appendix C).
First, general linear models (glm) were performed to assess the effect of the different land use classes on biomass increment. The area of orchards, forests, non-forest SNH and urban habitat were included as land use classes in the models. Area of cropland was strongly negatively correlated with all other land use classes (except for orchards at the 750 m radius) and was therefore omitted from further analyses (Online Appendix B). A backwards model selection was performed, and variables with a p value below 0.10 were retained. Following the same procedure, additional glm were performed to detect a correlation between amount of pollen collected or pollen diversity and land use.
As land use classes were expected to influence the colony performance through the available pollen resources, we also fitted glms on the relationship between (i) land use class and pollen availability, (ii) land use class and pollen diversity and (iii) pollen quantity and pollen diversity on colony growth. Models testing the effect of pollen diversity contained both the Simpson diversity of the collected pollen and the share of the three most commonly collected pollen types in each period.
To assess whether the cover of different land use classes has a consistent effect throughout the year, the correlation between the biomass increments of the colonies at the same location during the two consecutive periods was calculated. Only locations where the colonies developed during both periods were taken into account (n = 25).
Based on the correlations between landscape composition, pollen quantity, pollen composition and colony performance, a causal relationship was hypothesized. To test this, structural equation models (SEM) were made (Grace 2006) using the lavaan-package in R (Rosseel 2012). Separate models were made for the colonies during and after mass-flowering. All variables were standardized by subtracting by their mean and dividing through their standard error.
First, a model was created, based on our hypotheses on how these factors influence each other. Latent variables were created for land use classes cover, pollen composition, pollen quantity and colony performance. A Bollen-Stine test with 1000 bootstraps was performed to test for normality (Kim and Milsap 2014). Nonsignificant paths (p > 0.10) were dropped and the best SEM was selected based on its AICc. At each path, we indicated the standardized regression coefficients, which represent the amount of change on a parameter if the other one changes one standard deviation. The model fit was tested using a Chi squared test (p > 0.05) and the comparative fit index (CFI > 0.90) (Bentler 1990).
Results
Colony growth
During the first period, the bumblebee colonies grew on average with 197.7 (S.D. 209.2) g, with 15 out of 27 active colonies producing queen pupae after 6 weeks. Colony growth was strongly correlated to the cumulative amount of pollen collected (Fig. 2; Table 1). However, we found no significant effect of pollen composition or pollen diversity on colony growth. Presence of orchards had a marginally negative effect on colony growth.

Relationship between cumulative pollen load of sampled workers and colony growth a during and b after mass-flowering
During the second period, the colony biomass increment was 153.0 (S. D. 147.9) g, with 17 out of 26 active colonies producing queen pupae after 6 weeks. Colony growth was marginally positively correlated with pollen diversity, but negatively correlated with urban land.
Colony growth during the first and the second period were not correlated to each other (r = 0.03, p = 0.88).
Pollen collection
During the first period of the experiment, nine morphotypes of pollen were collected from foraging workers. Most pollen was collected from Rosaceae (68.2%), followed by Salix (15.7%) and Acer (12.1%). These three plant sources accounted for 96% of all collected pollen during this period. As the colonies were placed in apple and pear orchards and no other Rosaceae were abundantly flowering in the study area at the time of pollen collection, we consider it highly likely that the large majority of Rosaceae pollen that was collected belongs to these two species.
During the second period, most of the collected pollen belonged to Tilia (53.4%), followed by Papaver rhoeas-type (14.6%) and Lysimachia vulgaris-type (13.4%). Seventeen other morphotypes accounted for 18.5% of the total collected pollen (Online Appendix D). While during the first period 96.9% of all pollen was collected from woody plants, only 57.9% of all pollen collected during the second period originated from shrubs or trees. Additionally, 5.6% of all pollen collected during the second period belonged to plants that are primarily wind pollinated, such as Castanea, Urticaceae and grasses.
The amount of pollen collected during the first period was strongly negatively correlated to the area of orchards (Table 1). Land use also had a strong influence on pollen diversity and composition, with pollen diversity being negatively associated with orchard cover. Furthermore, the amount of Rosaceae pollen collected increased with orchard cover.
During the second period, no significant relationship was found between the amount of collected pollen, pollen composition or pollen diversity and land use.
Structural equation models
The structural equation models (Fig. 3) synthesise the results from the general linear models and confirm the strong connections between land use classes, pollen quantity, pollen composition and colony performance. During mass-flowering, the land use had a significant effect on both pollen quantity and pollen composition, with the total area of orchards decreasing pollen diversity and pollen load in foragers, while non-forest SNH had a positive effect on both. Only the amount of pollen collected had a direct influence on the colony performance, while land use had only an indirect effect through pollen quantity. After mass-flowering, forest cover had a positive effect on both the amount of pollen collected and the colony growth. No correlations between land use and pollen quality or pollen quality and colony performance were found after mass-flowering.

SEM explaining the relationships between landscape context, pollen quantity, pollen composition and colony performance a during (n = 27, χ2(7) = 12.36, p = 0.09, CFI = 0.93) and b after mass-flowering (n = 24, χ2(7) = 2.74, p = 0.91, CFI = 1.00). Observed variables are given in rectangular boxes, while latent variables are displayed in oval boxes. Positive correlations are indicated by green arrows, while negative correlations are indicated by red arrows. Arrow thickness is proportional to the standardized coefficient of the path. Grey, dashed lines indicate non-significant paths. Non-significant paths were not included in the final model. Standardized coefficients are indicated on the arrows. The R2 of dependent variables is displayed in the boxes. (Color figure online)
Discussion
Our study showed that bumblebees forage in different habitat types depending on the time of the year. Therefore, to maintain a healthy bumblebee population, it is important to provide a diverse landscape several types of SNH, which can provide pollen during different times of the season. During mass-flowering, bumblebees foraged on apple and pear trees, leading to a smaller scale of the land use effect than after mass-flowering.
We found that landscapes dominated by large, homogenous areas of orchards are negatively correlated with bumblebee colony growth. While other studies show that pollinators can benefit from mass-flowering crops (Holzschuh et al. 2013; Riedinger et al. 2014), we showed that preservation of semi-natural habitat in agro-landscapes has beneficial effects on B. terrestris colony performance by providing more pollen. While orchards can serve as foraging habitat for bumblebees during mass-flowering, bumblebees fare better in more diverse landscapes. Pure dependence on a succession of mass-flowering crops is therefore probably less effective to maintain bumblebee colonies in agricultural landscapes than providing sufficient semi-natural habitat.
Effect of land use on pollen quantity and quality and colony growth
We hypothesized that during mass-flowering of the orchards, these would serve as important foraging habitat and that the effect of semi-natural habitat would be minimal due to the large abundance of pollen resources. In our study, apple and pear trees were indeed the most important pollen source during spring. However, contrary to our expectations, in landscapes dominated by orchards, bumblebees collected less pollen, leading to a lower colony performance, in line with what Mattila and Otis (2006) found for honey bees. One possible reason for the low amount of pollen collected is competition with managed honeybees. Although orchards may provide large amounts of pollen and nectar during flowering season, high densities of honeybee hives are usually placed during flowering. Due to these unnaturally high densities (Geldmann and González-varo 2018), bumblebees and other wild pollinators may suffer from competition with honeybees (Thomson 2004; Goulson and Sparrow 2009; Shavit et al. 2009, but see Steffan-Dewenter and Tscharntke 2000) or pathogen spillover (Fürst et al. 2014). The negative effect of competition with honeybees is stronger in homogeneous landscapes (Herbertsson et al. 2016), making bumblebees in intensive agricultural landscapes dominated by orchards more susceptible than in more diverse agro-landscapes. Additionally, wild pollinator densities can increase in presence of mass-flowering crops (Holzschuh et al. 2013; Kovács-Hostyánszki et al. 2013), potentially further increasing the competition for pollen. Despite the ample presence of foraging resources, which, based on the identity of the collected pollen, attract the majority of foraging workers, flowering orchards could therefore be considered as ecological traps for bumblebees (Kallioniemi et al. 2017), as these attract foraging workers, while not providing sufficient pollen. Although we expected that mass-flowering apple trees would have a positive effect on bumblebees, colonies in landscapes with only a small area of orchards performed better.
Effect of semi-natural habitat on colony performance
We studied the role of land use on bumblebee foraging. Both during and after mass-flowering, colony performance was positively correlated with presence of semi-natural habitat. In spring, during mass-flowering, more pollen were collected in areas with more non-forest semi-natural habitat. After mass-flowering, colony growth was slightly better at locations with a higher surrounding forest cover, where Tilia and Castanea are important pollen sources. Bees are known to forage in different habitats (Mandelik et al. 2012). Although semi-natural habitat can serve as an important pollen and nectar source throughout the season (Scheper et al. 2014; Földesi et al. 2015; Requier et al. 2015), B. terrestris, having a very broad diet, is able to forage in agricultural (Odoux et al. 2012) and urban (Chapman et al. 2003) habitats. During mass-flowering, most pollen was collected from fruit-bearing trees in orchards, while after mass-flowering, the main pollen sources were plants typical for forest and urban (Tilia, Lysimachia vulgaris-type) and agricultural (Papaver rhoeas-type) habitat. These land use classes may therefore also serve as suitable foraging habitat for B. terrestris during specific periods of the year (Goulson et al. 2002, but see Bukovinszky et al. 2017).
As the bumblebees were placed in artificial bumblebee nests, we did not investigate the role of the landscape as nesting habitat. While our study found a relatively weak positive effect of SNH on colony performance, it may have a strong effect on nesting place availability, leading to a higher colony density in landscapes with a higher cover of SNH.
Seasonal aspect of foraging habitat
A gap in pollen availability during the flying season can limit bumblebee colony growth or survival (Vanbergen and the Insect Pollinators Initiative 2013). We showed that colony growth during and after mass-flowering on a single location were not correlated. In our study, colonies at various locations performed differently during and after mass-flowering, and the amount of pollen collected differed strongly between the two periods. This implies that while a landscape could provide sufficient pollen resources to support a bumblebee colony at a given time, only 2 months later, the same location could be a hostile environment for bumblebees. While one type of land use could provide the necessary foraging resources for bumblebees during part of the season, it could be poor in floral resources at other times of the year (Williams et al. 2012). For long-term survival of a bumblebee population, it is therefore necessary that sufficient foraging resources are continuously available within flying distance of bumblebee colonies during the flying season, as also observed for other groups of social bees (Franzén and Nilsson 2008; Decourtye et al. 2010; Aleixo et al. 2016), although the exact locations and land use classes that provide pollen and nectar may change throughout the year.
While B. terrestris colonies are active for a large time of the year (Rasmont et al. 2008), most floral resources are only present during a short period. Except for small amounts of Crepis-type, we did not find a single pollen morphotype that was collected during both periods of the experiment. With changing flower resources, the role of the landscape as foraging habitat also changes. While most pollen during the first period was collected on mass-flowering fruit-trees, linden trees, which in the study region are mostly associated with (sub)urban habitat, were the major pollen source during the second period.
The major foraging habitat within a certain period also influences the foraging distance, hence altering the spatial scale on which the landscape influences the colonies. During mass-flowering, when most pollen was collected in the orchards themselves, the land use effects were strongest within a 750 m radius, while after mass-flowering, when several more distant foraging habitats were used, land use effects were strongest at a 1000 m scale (Danner et al. 2016).
References
Aleixo KP, Menezes C, Imperatriz Fonseca VL, da Silva CI (2016) Seasonal availability of floral resources and ambient temperature shape stingless bee foraging behavior (Scaptotrigona aff. depilis). Apidologie 48:117–127
ALV (2016) Landbouwgebruikspercelen [Agricultural land use parcels map]
Bentler P (1990) Comparative fit indices in structural models. Quant Methods Psychol 107:238–246
Beug HJ (2004) Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Friedrich Pfeil
Bukovinszky T, Verheijen J, Zwerver S, Klop E, Biesmeijer JC, Wäckers FL, Prins HHT, Kleijn D (2017) Exploring the relationships between landscape complexity, wild bee species richness and reproduction, and pollination services along a complexity gradient in the Netherlands. Biol Conserv 214:312–319
Carvell C, Roy DB, Smart SM, Pywell RF, Preston CD, Goulson D (2006) Declines in forage availability for bumblebees at a national scale. Biol Conserv 132:481–489
Chapman RE, Wang J, Bourke AFG (2003) Genetic analysis of spatial foraging patterns and resource sharing in bumble bee pollinators. Mol Ecol 12:2801–2808
Croxton PJ, Carvell C, Mountford JO, Sparks TH (2002) A comparison of green lanes and field margins as bumblebee habitat in an arable landscape. Biol Conserv 107:365–374
Danner N, Molitor AM, Schiele S, Härtel S, Steffan-Dewenter I (2016) Season and landscape composition affect pollen foraging distances and habitat use of Honey bees. Ecol Appl 26:1920–1929
De Knijf G, De Saeger S, Vriens L, Oosterlynck P, Paelinckx D (2010) Wetenschappelijke ondersteuning van het gebiedsgericht beleid: De Biologische Waarderingskaart. Mededeling van het Instituut voor Natuur- en Bosonderzoek, Brussels
Decourtye A, Mader E, Desneux N (2010) Landscape enhancement of floral resources for honey bees in agro-ecosystems. Apidologie 41:264–277
Fitzpatrick Ú, Murray TE, Paxton RJ, Breen J, Cotton D, Santorum V, Brown MJF (2007) Rarity and decline in bumblebees—a test of causes and correlates in the Irish fauna. Biol Conserv 136:185–194
Földesi R, Kovács-Hostyánszki A, Korösi Á, Somay L, Elek Z, Markó V, Sárospataki M, Bakos R, Varga Á, Nyisztor K, Báldi A (2015) Relationships between wild bees, hoverflies and pollination success in apple orchards with different landscape contexts. Agric For Entomol. https://doi.org/10.13140/rg.2.1.3702.8566
Franzén M, Nilsson SG (2008) How can we preserve and restore species richness of pollinating insects on agricultural land? Ecography (Cop.) 31:698–708
Fürst MA, Mcmahon DP, Osborne JL, Paxton RJ, Brown MJF (2014) Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 506:364–366
Gabriel D, Tscharntke T (2007) Insect pollinated plants benefit from organic farming. Agric Ecosyst Environ 118:43–48
Geldmann J, González-varo JP (2018) Conserving honey bees does not help wildlife. Science 80:359–392–393
Génissel A, Aupinel P, Bressac C, Tasei J-N, Chevrier C (2002) Influence of pollen origin on performance of Bombus terrestris micro-colonies. Entomol Exp Appl 104:329–336
Goulson D, Hanley ME, Darvill B, Ellis JS, Knight ME (2005) Causes of rarity in bumblebees. Biol Conserv 122:1–8
Goulson D, Hughes WOH, Derwent LC, Stout JC (2002) Colony growth of the bumblebee, Bombus terrestris, in improved and conventional agricultural and suburban habitats. Oecologia 130:267–273
Goulson D, Sparrow KR (2009) Evidence for competition between honeybees and bumblebees; effects on bumblebee worker size. J Insect Conserv 13:177–181
Grace JB (2006) Structural equation modeling and natural systems. Cambridge University Press, New York
Herbertsson L, Lindström SAM, Rundlöf M, Bommarco R, Smith HG (2016) Competition between managed honeybees and wild bumblebees depends on landscape context. Basic Appl Ecol 17:609–616
Hines HM, Hendrix SD (2005) Bumble bee (Hymenoptera: Apidae) diversity and abundance in tallgrass prairie patches: effects of local and landscape floral resources. Environ Entomol 34:1477–1484
Holland JD, Bert DG, Fahrig L, Holland JD, Bert DG, Fahrig L (2004) Determining the spatial scale of species’ response to habitat. Bioscience 54:227–233
Holzschuh A, Dormann CF, Tscharntke T, Steffan-Dewenter I (2013) Mass-flowering crops enhance wild bee abundance. Oecologia 172:477–484
Holzschuh A, Steffan-Dewenter I, Tscharntke T (2008) Agricultural landscapes with organic crops support higher pollinator diversity. Oikos 117:354–361
Kallioniemi E, Åström J, Rusch GM, Dahle S, Åström S, Gjershaug JO (2017) Local resources, linear elements and mass-flowering crops determine bumblebee occurrences in moderately intensified farmlands. Agric Ecosyst Environ 239:90–100
Kämper W, Werner PK, Hilpert A, Westphal C, Blüthgen N, Eltz T, Leonhardt SD (2016) How landscape, pollen intake and pollen quality affect colony growth in Bombus terrestris. Landscape Ecol 31:2245–2258
Kim H, Milsap R (2014) Using the Bollen-Stine bootstrapping method for evaluating approximate fit indices. Multivar Behav Res 49:581–596
Knight ME, Martin AP, Bishop S, Osborne JL, Hale RJ, Sanderson RA, Goulson D (2005) An interspecific comparison of foraging range and nest density of four bumblebee (Bombus) species. Mol Ecol 14:1811–1820
Kovács-hostyánszki A, Haenke S, Batáry P, Jaukjer B, Báldi A, Tscharntke T, Holzschuh A (2013) Contrasting effects of mass-flowering crops on bee pollination of hedge plants at different spatial and temporal scales. Ecol Appl 23:1938–1946
Lye GC, Park K, Osborne JL, Holland JM, Goulson D (2009) Assessing the value of Rural Stewardship schemes for providing foraging resources and nesting habitat for bumblebee queens (Hymenoptera: Apidae). Biol Conserv 142:2023–2032
Mandelik Y, Winfree R, Neeson T, Kremen C (2012) Complementary habitat use by wild bees in agro-natural landscapes. Ecol Appl 22:1535–1546
Mattila H, Otis G (2006) Influence of pollen diet in spring on development of honey bee (Hymenoptera: Apidae) colonies. J Econ Entomol 99:604–613
Moore PD, Collinson M, Webb JA (1994) Pollen analysis, 2nd edn. Wiley, Hoboken
Müller A, Diener S, Schnyder S, Stutz K, Sedivy C, Dorn S (2006) Quantitative pollen requirements of solitary bees: implications for bee conservation and the evolution of bee-flower relationships. Biol Conserv 130:604–615
Öckinger E, Smith HG (2007) Semi-natural grasslands as population sources for pollinating insects in agricultural landscapes. J Appl Ecol 44:50–59
Odoux JF, Feuillet D, Aupinel P, Loublier Y, Tasei J-N, Mateescu C (2012) Territorial biodiversity and consequences on physico-chemical characteristics of pollen collected by honey bee colonies. Apidologie 43:561–575
Osborne JL, Clark SJ, Morris RJ, Williams IH, Riley JR, Smith AD, Reynolds DR, Edwards AS (1999) A landscape-scale of bumble bee foraging study range and constancy, using harmonic radar. J Appl Ecol 36:519–533
Parmentier L, Meeus I, Cheroutre L, Mommaerts V, Louwye S, Smagghe G (2013) Commercial bumblebee hives to assess an anthropogenic environment for pollinator support: a case study in the region of Ghent (Belgium). Environ Monit Assess 185:1–11
Peat J, Goulson D (2005) Effects of experience and weather on foraging rate and pollen versus nectar collection in the bumblebee, Bombus terrestris. Behav Ecol Sociobiol 58:152–156
Pywell RF, Warman EA, Carvell C, Sparks TH, Dicks LV, Bennett D, Wright A, Critchley CNR, Sherwood A (2005) Providing foraging resources for bumblebees in intensively farmed landscapes. Biol Conserv 121:479–494
QGIS Development Team (2015) QGIS 2.16 Geographic Information System. Open Source Geospatial Foundation Project. http://qgis.osgeo.org
R Core Team (2018) R: a language and environment for statistical computing
Rasmont P, Coppee A, Michez D, De Meulemeester T (2008) An overview of the Bombus terrestris (L. 1758) subspecies (Hymenoptera: Apidae). Ann. la Société Entomol. Fr. 44:243–250
Requier F, Odoux JF, Tamic T, Moreau N, Henry M, Decourtye A, Bretagnolle V (2015) Honey bee diet in intensive farmland habitats reveals an unexpectedly high flower richness and a major role of weeds. Ecol Appl 25:881–890
Riedinger V, Renner M, Rundlöf M, Steffan-Dewenter I, Holzschuh A (2014) Early mass-flowering crops mitigate pollinator dilution in late-flowering crops. Landscape Ecol 29:425–435
Rosseel Y (2012) lavaan: an R Package for Structural Equation Modeling. J Stat Softw 48:1–36
Rotheray EL, Osborne JL, Goulson D (2017) Quantifying the food requirements and effects of food stress on bumble bee colony development. J Apic Res 56:288–299
Rundlöf M, Persson AS, Smith HG, Bommarco R (2014) Late-season mass-flowering red clover increases bumble bee queen and male densities. Biol Conserv 172:138–145
Scheper J, Reemer M, van Kats R, Ozinga WA, van der Linden GTJ, Schaminée JHJ, Siepel H, Kleijn D (2014) Museum specimens reveal loss of pollen host plants as key factor driving wild bee decline in The Netherlands. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.1412973111
Shavit O, Dafni A, Neeman G (2009) Competition between honeybees (Apis mellifera) and native solitary bees in the Mediterranean region of Israel—implications for conservation. Isr J Plant Sci 57:171–183
Stanley DA, Stout JC (2014) Pollinator sharing between mass-flowering oilseed rape and co-flowering wild plants: implications for wild plant pollination. Plant Ecol 215:315–325
Steffan-Dewenter I, Tscharntke T (1999) Effects of habitat isolation on pollinator communities and seed set. Oecologia 121:432–440
Steffan-Dewenter I, Tscharntke T (2000) Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia 122:288–296
Tasei J-N, Aupinel P (2008) Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 39:397–409
Thomson D (2004) Competitive interactions between the invasive European honey bee and native bumble bees. Ecology 85:458–470
Vanbergen AJ (2013) The Insect Pollinators Initiative. Threats to an ecosystem service: pressures on pollinators. Front Ecol Environ 11:251–259
Walther-Hellwig K, Frankl R (2000) Foraging distances of Bombus muscorum, Bombus lapidarius, and Bombus terrestris (Hymenoptera, Apidae). J Insect Behav 13:239–246
Westphal C, Steffan-Dewenter I, Tscharntke T (2003) Mass flowering crops enhance pollinator densities at a landscape scale. Ecol Lett 6:961–965
Westphal C, Steffan-Dewenter I, Tscharntke T (2006) Foraging trip duration of bumblebees in relation to landscape-wide resource availability. Ecol Entomol 31:389–394
Williams PH, Osborne JL (2009) Bumblebee vulnerability and conservation world-wide. Apidologie 40:367–387
Williams NM, Regetz J, Kremen C, Ecology S (2012) Landscape-scale resources promote colony growth but not reproductive performance of bumble bees. Ecology 93:1049–1058
Wolf S, Moritz RFA (2008) Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 39:419–427
Acknowledgements
We are very grateful to Tim Beliën (pcfruit) for his help in finding suitable locations for the experiment and to the farmers that allowed the experiment to be carried out in their orchards. We thank Felix Wäckers (Biobest) for providing artificial bumblebee colonies. The first author was supported by a grant by FWO-Vlaanderen (Grant No. FWO14/ASP/195).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Proesmans, W., Smagghe, G., Meeus, I. et al. The effect of mass-flowering orchards and semi-natural habitat on bumblebee colony performance. Landscape Ecol 34, 1033–1044 (2019). https://doi.org/10.1007/s10980-019-00836-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00836-5