
Abstract
Context
Global climate change impacts forest growth and methods of modeling those impacts at the landscape scale are needed to forecast future forest species composition change and abundance. Changes in forest landscapes will affect ecosystem processes and services such as succession and disturbance, wildlife habitat, and production of forest products at regional, landscape and global scales.
Objectives
LINKAGES 2.2 was revised to create LINKAGES 3.0 and used it to evaluate tree species growth potential and total biomass production under alternative climate scenarios. This information is needed to understand species potential under future climate and to parameterize forest landscape models (FLMs) used to evaluate forest succession under climate change.
Methods
We simulated total tree biomass and responses of individual tree species in each of the 74 ecological subsections across the central hardwood region of the United States under current climate and projected climate at the end of the century from two general circulation models and two representative greenhouse gas concentration pathways.
Results
Forest composition and abundance varied by ecological subsection with more dramatic changes occurring with greater changes in temperature and precipitation and on soils with lower water holding capacity. Biomass production across the region followed patterns of soil quality.
Conclusions
Linkages 3.0 predicted realistic responses to soil and climate gradients and its application was a useful approach for considering growth potential and maximum growing space under future climates. We suggest Linkages 3.0 can also can used to inform parameter estimates in FLMs such as species establishment and maximum growing space.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climate warming is unequivocal and observed changes are unprecedented over decades to millennia (IPCC 2013). The associated changes in weather will impact the distribution and abundance of tree species globally (Allen et al. 2010). Thus, tools are needed to quantify the impact of predicted climate changes on forests to mitigate and manage ecological and economic impacts.
Forest landscape models such as LANDIS PRO (Wang et al. 2014), LANDIS II (Scheller and Mladenoff 2005), LANDCLIM (Schumacher et al. 2004), TreeMig (Lischke et al. 2006) and iLAND (Seidl et al. 2012) have been used to model forest responses to climate change. These models incorporate spatial interactions of landscape processes such seed dispersal, which facilitates species migration, and mortality from disturbances such as harvest, fire and wind. Forest landscape models often simplify physiological detail because of the immense computation cost of including spatial interactions (He 2008). Alternatively, processes that are estimated in landscape models such as growth, mortality from competition, and stand dynamics come from biophysical processes directly measured and modeled, which can be aggregated from individuals to patch level which can be aggregated to the landscape level (Lischke et al. 1998; Shifley et al. this issue).
Niche and biophysical process models have been commonly used to quantify changes to potential tree habitat, predict range shifts of tree species (Iverson et al. 2008; Morin et al. 2008; Morin and Thuiller 2009), and can be used to evaluate species suitability for assisted migration in response to global climate change. These models usually operate at relatively coarse spatial scales (e.g., 20–50 km cell size) and site-scale dynamics (e.g., tree species demography and biotic interactions in a forest stand) are either ignored or highly simplified. These site-scale processes may be more important than the direct effects of climate change in affecting tree habitats, abundances, and distribution change (Gustafson et al. 2010; Li et al. 2013). This occurs especially when the effects of disturbance are relatively weak and site-scale processes are critical determinants of future forest changes (Wang et al. 2015) such as in the central hardwood forest region (CHFR). Niche and most biophysical process models also do not usually include fine temporal (e.g., daily) weather data. Monthly, yearly, or regional averages may not be sufficient to capture extreme weather events such as drought, which can have a significant effect on forest structure and dynamics (Gu et al. 2015). Thus, not including site-scale processes and fine-scale weather data in niche and biophysical models may result in great uncertainties (Purves and Pacala 2008).
Comparisons of results from multiple model methodologies can provide some confidence in general predictions and highlight discrepancies for future investigation (Schneiderman et al. 2015; Iverson et al. 2016). Also, coupled ecosystem process and landscape models have been used to incorporate information from different model methodologies to better estimate parameters previously only crudely approximated. For example, results from the PnET biophysical model are used to estimate species establishment probabilities which are defined as the relative probability of a species to reproduce and survive under given environmental conditions. This method uses the relative biomass of each species in relation to the maximum biomass of any single species (He et al. 1999). Also, PnET is used to calculate the maximum annual net primary production in LANDIS II, a forest landscape simulation model (Scheller et al. 2007).
There is a renewed interest in applying stand dynamic models (gap models) to study forest response to climate change at regional scales (e.g. Vanderwell and Purves 2014), but many of these models are specifically designed to simulate site-scale dynamics and are limited in processing landscape and regional scale data. Here, we use the biophysical process model LINKAGES 3.0 to estimate the effects of climate change using the dominant soils from across each subsection.
LINKAGES is a plot-based forest ecosystem process model that simulates growth, competition for light, nutrients and soil moisture, seedling establishment, nutrient cycling, evapotranspiration and soil hydrology. The original LINKAGES model (Pastor and Post 1985) was developed from the gap models JABOWA (Botkin et al. 1972) and FORET (Shugart and West 1980). Whereas LINKAGES v1.0 used mean monthly climate data to drive the model, LINKAGES v 2.2 incorporated updated water and nutrient cycles used daily climate data (Wullschleger et al. 2003), which lends its use to daily downscaled climate data from general circulation models (GCM). But LINKAGES v 2.2 used an obsolete hydrology module and had some systemic programming problems that were corrected in version 3.0 (Appendix A in Supplementary material).
As a test area, we use the central hardwood forest region (CHFR) of the U.S. to evaluate the reasonableness of results from Linkages 3.0. Such evaluation is necessary to afford confidence in parameter estimate used in forest landscape models. The CHFR contains one of the most extensive temperate deciduous forests in the world (Johnson et al. 2009). Forests in CHFR are among the biomes that have been most severely influenced by climatic change, land use (e.g., logging and agriculture), and land management (e.g., fire suppression, harvest) (Reich and Frelich 2001). In this area, extensive modeling of tree species response to climate change has been conducted using niche models (Iverson et al. 2008) and forest landscape models (Wang et al. 2015). Outcomes from forest ecosystem process models such as LINKAGES, when contrasted with niche based models such as tree atlas (Iverson et al. 2016) that uses different methodology, provides an increased confidence in results when congruence between model results exists. These approaches can be used to help parameterize forest landscape models to reduce prediction uncertainties (Wang et al. 2015).
Our objectives were to (1) revise LINKAGES v 2.2 to improve simulation realism at landscape scales when compared to Forest Inventory and Analysis (FIA) data; (2) provide a tool to facilitate parameterization of forest landscape models such as LANDIS PRO; and (3) apply the improved model (v3.0) to predict responses of U.S. central hardwood forest to future climate scenarios in 2070–2099.
We focused on species growth (in biomass) over 100 years under current climate and climate projections for the end of the century and interpreted maximum biomass reached by year 30 and 100 as measures of early growth potential and maximum growing space (i.e., carrying capacity), respectively. Maximum growing space is defined as maximum total biomass of all species competing for soil and environmental resources beyond which self-thinning occurs. This approach let us focus on two attributes of species and ecosystems that may be particularly sensitive to climate change and that are also used as parameters in forest landscape models (Wang et al. 2015, 2016). This approach differs from approaches that simulate stand development over an actual time series of climate change. To assess the effect of temporal variability, we evaluated the relationship between mean annual precipitation and dry days to show the importance of using daily downscaled data to quantify the effects of drought. The LINKAGES 3.0 model provides new features including simulation of individual tree growth through physiological processes, a daily time step to assess water availability after runoff and evaporation, and incorporation of spatial heterogeneity from use of multiple soils per subsection. Details of model revisions appear in appendix A.
Study area
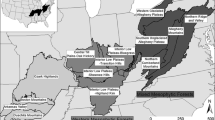
We modeled the CHFR (Braun 1950; Fralish 2003) of the United States (Fig. 1). The area encompasses nine ecological sections, 74 subsections, and a variety of vegetation, terrains, soils, and climates. Forests are dominated by oaks (Quercus spp.) with a consistent small proportion of hickories (Carya spp.) and miscellaneous deciduous species. Southern portions of the region also contain loblolly and shortleaf pine. A thorough description of forest types within the region can be found in Johnson et al. (2009, Appendix 2). The western portion of the region tends to be more xeric with shallow to very deep soils which are moderately well drained to excessively drained and the eastern portion tends to be more mesic with moderately deep to very deep soils which are well drained and clayey (USDA 2006). The CHFR climate is humid temperate and geographically falls within the central interior broadleaf forest province of the hot continental division (Cleland et al. 2007).

Ecological subsections in the central hardwood region of the United States and the five subsections (shaded and labeled) for which we provide species-specific changes in biomass forecast by LINKAGES 3.0 under climate change
Methods
Climate and soils
We selected information for climate and soil for 74 subsections from the CHFR. We compared current climate (1980–2009) to four future climate scenarios based on years 2070–2099 from two of the latest CMIP5 general circulation models (IPCC AR5) with two different CO2 emission scenarios. We used output from the MRI-CGCM3 model developed by the Meteorological Research Institute (Yukimoto et al. 2012) and the GFDL-ESM2M model by NOAA Geophysical Fluid Dynamics Laboratory (Delworth et al. 2006) and representative concentration pathways (RCP) 6.0 which is classified as a moderate CO2 scenario and 8.5 which is considered a high emission scenario (IPCC 2013, Annex III). We reference the GCM and RCP in our future climate scenarios as GCM_RCP (i.e. CGCM3_RCP60). Current trends show climates warming and CO2 emissions increasing but the long term trajectories are unknown, we therefore bracket climate scenarios between current climate and the highest emission scenario to understand the range of outcomes. By modeling forests under climates resulting from multiple GCMs and RCP scenarios we incorporated uncertainties in climate change while evaluating potential forest growth for the region.
We obtained daily maximum and minimum temperature, daily precipitation, daily wind speed, and daily solar radiation for future climate projections (Livneh et al. 2013; Reclamation 2014; Thornton et al. 2016). Resolution of observed current climate data was 1/16th degree (~6 km), except for solar radiation, which had 1 km resolution. Resolution of future climate data was 1/8-degree resolution (~12 km), except for wind speed and solar radiation, which were not spatially downscaled and available at a resolution of approximately 1.875° latitude by 1.25° longitude. We intersected ecological subsection boundaries with the climate rasters and used the climate values from a single raster cell with the median precipitation levels to represent the subsection since processing every climate raster cell with multiple soils would be extremely computationally intensive in parameterization, simulation and output interpretation.
We obtained measures of soil organic matter, nitrogen, wilting point, field moisture capacity, percent clay, sand and rock for soil polygons from the Natural Resources Conservation Service soil survey (Soil Survey Staff, http://soils.usda.gov/). Thirty simulations were run for each of the five most abundant soils within each subsection. Biomass averages from the 30 simulations for each soil were then used to create area-weighted average forest biomass for each subsection based on soil abundance.
Model simulation and data analysis
We simulated tree species growth for current climate (1980–2009) and the four climate change scenarios for the period 2070–2099. We ran model simulations for 100 years and recycled the 30 years of weather data for each scenario so the entire simulation was based on current climate or climate at the end of the century. We modeled dominant species determined from FIA data in each subsection and the subsections adjoining it and started simulations from bare ground. We report total forest biomass, and in some examples individual tree biomass, at simulation year 30, 70, and 100 for current climate and each climate scenario modeled. We also compared total forest biomass from LINKAGES simulation years 30, 70 and 100 under current climate to FIA biomass estimates by stand age. We binned and averaged FIA plot stand age for ages 20–40, 60–80 and 90–110 to correspond to simulation years 30, 70 and 100 respectively. We estimated maximum growing space for each soil as total biomass of all species at year 100 divided by the average total biomass for all soils within subsections being modeled at year 100.
We also report more detailed data on growth potential for five selected subsections that represented mean annual temperature range of 11.7–16.4 °C and mean annual precipitation range of 96.0–121.6 cm under current climate (Table 1). These subsections had gradients of soil qualities and capacities and represented low to high current and projected temperatures to provide a range of potential outcomes and to evaluate the relationship between mean annual temperature and dry days (Table 1). Dry days were defined as days when potential evapotranspiration demand exceeded available soil moisture; trees are increasingly drought stressed as the number of dry days increase.
Results
Climate warmed 1–6 °C (Fig. 2) and precipitation varied ±12 cm under climate change scenarios in the region. The number of dry days increased under some climate change scenarios even when mean annual precipitation increased (Table 1). The mean number of annual dry days under current climate was approximately 12 with 3 subsections having greater than 40 dry days. Under modeled climate scenarios 3 out of 4 had at least 1 subsection with greater than 40 mean annual dry days (Fig. 3). The average length of growing season averaged ~135 days for deciduous trees and ~184 days for conifer trees.

Mean increase in mean annual temperature for general circulation models: a GFDL-ESM2M RCP 6.0, b GFDL-ESM2M RCP 8.5, c MRI-CGCM3 RCP 6.0, d MRI-CGCM3 RCP 8.5, and mean change in total annual precipitation for general circulation models: e GFDL-ESM2M RCP 6.0, f GFDL-ESM2M RCP 8.5, g MRI-CGCM3 RCP 6.0, h MRI-CGCM3 RCP 8.5; for years 2070–2099 compared to current climate years 1980–2009

Average number of growing season dry days for general circulation models: a GFDL-ESM2M RCP 6.0, b GFDL-ESM2M RCP 8.5, c MRI-CGCM3 RCP 6.0, d MRI-CGCM3 RCP 8.5 based on years 2070–2099 and e current climate years 1980–2009
Forest species composition, total biomass, and growth potential were affected by climate change. Total biomass varied among climate change scenarios and was driven by species specific growth. Total biomass among scenarios ranged from greater than 75 to 197 Mg/Ha (Fig. 4). No subsections had biomasses less than 75 Mg/ha or greater than 200 Mg/ha under current climate at year 30. Departures from current climate were greater under RCP 8.5 than 6.0 and greatest with GFDL. The patterns of differences in total biomass among current climate and climate change scenarios were similar at years 30, 70, and 100.

Total biomass in simulation year 30, 70 and 100 for current climate years 1980–2009 and general circulation models MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5, GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5 based on years 2070–2099
Total biomass production from LINKAGES 3.0 under current climate at year 30 was not significantly correlated with estimates of mean biomass for FIA plots (Pearson correlation, r = 0.04, P = 0.69) but was significantly correlated at year 70 (r = 0.27, P = 0.015) and year 100 (r = 0.23, P = 0.042) under the current climate scenario. Comparison of biomass estimates from LINKAGES to FIA data produced a mean absolute error (MAE) of 82.9, 47.1, and 39.8 Mg/ha for year 30, 70, and 100, respectively.
Representative subsection results
Subsection 223Af, Current River Hills, is in the western portion of the CHFR in the heart of the historic shortleaf pine range in Missouri. The subsection increased in mean annual temperature 2 °C under CGCM3_RCP60, 3 °C under CGCM3_RCP85 and GFDL_RCP60, and 4 °C under GFDL_RCP85. There was no change in mean annual precipitation for CGCM3_RCP60, GFDL_RCP60 and GFDL_RCP85 and a 6-cm increase in CGCM3_RCP85. Biomass of white oak, post oak, mockernut hickory and red maple were greater under all future climate scenarios than current climate (Fig. 5). Bitternut, pignut and shagbark hickories, northern red oak and sugar maple grew well under current climate but had lower biomass under all climate change scenarios. Shortleaf pine grew early in all simulations but slowly declined because of its inability to regenerate under fully stocked conditions.

Subsection 223Af biomass by species for 100 years of simulation for general circulation models GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5, MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5 based on years 2070–2099 and current climate years 1980–2009
Subsection 223Bc, Mitchell Karst Plains, is in the eastern portion of the CHFR. The section increased in mean annual temperature 3–4 °C under future climate scenarios. Mean annual precipitation increased 3–7 cm under future climate scenarios. Under current climate and RCP 6.0 emission scenarios Virginia pine dominated early growth but red maple dominated in later years (Fig. 6). Patterns of growth under RCP 6.0 closely followed current climate patterns for most species with the exception of sugar maple which wasn’t found in any of the future climate scenarios. Under the RCP 8.5 emissions scenario yellow poplar increased within the subsections reaching maximum biomass at approximately year 40. Virginia pine growth was replaced by yellow poplar as a dominant species during early to middle years under RCP 8.5 but declined as red maple increased in later years.

Subsection 223Bc biomass by species for 100 years of simulation for general circulation models GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5, MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5 based on years 2070–2099 and current climate years 1980–2009
Subsection 223Eh, Pennyroyal Karst Plains increased in mean annual temperature 2–3 °C and increased in mean annual precipitation 1–9 cm under future climate scenarios (Table 1). Virginia pine and loblolly pine dominated early growth under current climate but red maple, white oak, bur oak, chestnut oak and northern red oak dominated in later years (Fig. 7). Under future climate scenarios early growth was dominated by loblolly pine but red maple and white oak was dominant in later years.

Subsection 223Eh biomass by species for 100 years of simulation for general circulation models GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5, MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5 based on years 2070–2099 and current climate years 1980–2009
Subsection 231Cd, Sandstone Mountain increased in mean annual temperature 1–4 °C. Mean annual precipitation increased 4–5 cm under the CGCM3 climate model and decreased 1–3 cm under the GFDL climate model (Table 1). Mean annual precipitation under current climate was the lowest of the five subsections examined in detail and the number of dry days was higher under current climate and CGCM3 than GFDL. The regular periodicity in this subsection was attributed to the recycling of 30 year climate data over the 100 year simulation time (Fig. 8). Under current climate, early dominance by red maple, white ash, yellow poplar, southern red oak and American elm gave way to bitternut hickory, green ash, white oak, scarlet oak, chestnut oak, northern red oak and post oak in later years (Fig. 8). Dominance of fast growing mesic species declined due to drought induced mortality and drought tolerant species increased. Under CGCM3 RCP6.0 and RCP8.5 there was a similar decline in mesic species over time but more drought tolerant species increased compared to current climate. Under GFDL RCP 6.0 and RCP 8.5 initial dominance by loblolly pine decreases while red maple continued as a dominant species in later years. Pignut and mockernut hickories, southern hackberry, white and green ash, yellow poplar and white oak continued to increase in biomass but yellow poplar, southern red oak, water oak, cherrybark oak, willow oak, post oak, black oak and American elm peaked in middle years and then declined. The dominance and then decline of mesic species is suggestive of higher than normal precipitation followed by drought.

Subsection 231Cd biomass by species for 100 years of simulation for general circulation models GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5, MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5 based on years 2070–2099 and current climate years 1980–2009
Subsection 231Gc, Wester Arkansas Valley and Ridges increased in mean annual temperature of 2–4 °C (Table 1). Mean annual precipitation varied from an increase of 3 cm to a decrease of 12 cm. Loblolly pine dominated early years and then declined. This subsection was out of the range of many oaks and was dominated by pignut and mockernut hickories, southern hackberry and white and green ash in later years under current climate (Fig. 9). Under future climate scenarios loblolly pine had even greater early dominance but then still declined. Moving from current climate to CGCM3_RCP6 to GFDL_RCP8.5, the number of species capable of growth declined to mockernut hickory, southern hackberry, green ash, water oak and post oak. The highest number of dry days occurred under current climate (Table 1), however, the variation in the number of dry days among the five soils modeled was greater in the climate change scenarios (see Fig. 10 for example for current climate). The substantial variation among soils gives cause for judicious interpretation of individual species growth potential at the subsection level. Under current climate, for example, soils 2–5 had relatively similar species composition and total biomass production while soil 1 is distinctively different in species composition and total biomass production and has an increased number of drought events even though all soils were run with the exact same climate.

Subsection 231Gc biomass by species for 100 years of simulation for general circulation models GFDL-ESM2M RCP 6.0, GFDL-ESM2M RCP 8.5, MRI-CGCM3 RCP 6.0, MRI-CGCM3 RCP 8.5 based on years 2070–2099 and current climate years 1980–2009

Biomass by species on the five dominant soils over 100 years of current climate in subsection 231Gc
Discussion
We used an updated version of LINKAGES (v3.0) to evaluate model performance and growth potential of tree species across the CHFR region under current and future climate. The model produced reasonable estimates of biomass after we increased growth using a new equation and coefficients, added biomass limits using a stocking equation, and updated species parameters. We also corrected numerous programming anomalies that occurred when compiled on a windows platform (e.g., the random number generator, counting loops) to model tree growth from bare ground plots (see appendix A in Supplementary material).
LINKAGES 3.0 produced realistic results that followed expected patterns in stand development and the soil and climate gradients among the five subsections that we examined in detail. The more xeric soils of the western CHFR produced less total biomass than the more mesic soils in the eastern region at year 30 and year 70 under current climate. Subsections in the eastern and more mesic portion of the CHFR increased in total biomass under the future climate scenarios. Subsections that exhibited little or no change in mean annual temperature and total precipitation still exhibited changes in species composition and biomass over time as a result of succession. These results followed generally expected patterns of stand development (Johnson et al. 2009). (2) As species niches disappeared due to changes in climate other species were often able to utilize newly available resources and total biomass changed very little. This pattern was observed in subsection 231Bc, which had more mesic soils and consequently fewer dry days than the other subsections in the eastern portion of the CHFR region (Table 1). The pattern did not hold in subsection 231Cd, where total biomass increased or decreased from current climate depending on the scenario, and 231Gc, where total biomass increased from current climate due to increased climate suitability of species capable of producing much higher basal areas. Early growth was dominated by loblolly pine and shortleaf pine in subsection 223Af, 223Bc, and 223Eh, but red maple and white oak were dominant in later years. This likely occurred because pine is not shade tolerant and was unable to regenerate and sustain its biomass as older trees died. We saw a decline in shade intolerant species across subsections because these species cannot reproduce under full canopy and we did not simulate large-scale disturbance such as fire. In addition, pines usually occur in fire-prone systems and we expected to see pines replaced by more shade tolerant or mesic species over time without fire (Guyette and Kabrick 2000). These types of successional dynamics may not be predicted by niche and biophysical models but are incorporated in forest landscape simulation models.
The daily accounting of evaporation and hydrology in LINKAGES differs from models that use monthly, seasonal, or annual precipitation. LINKAGES is able to simulate hydrology and evapotranspiration that occur when daily precipitation is extreme. Future climates are thought to contain such events (IPCC 2013) and behavior of forest growth within subsections will vary dependent on soil characteristics. Some subsections (e.g., 231Gc) had soils that were well drained to excessively well drained and had little water holding capacity, so species with higher drought tolerances were at a competitive advantage over drought intolerant species. This can lead to reduced species richness but potentially higher biomasses due to elevated temperatures that favor growth of some species. For example, white oak and post oak often increased in biomass over time under climate change scenarios while drought-intolerant species decreased in biomass, and species richness eventually declined.
Mean annual precipitation can mask the importance of water availability during the growing season. Monthly or growing season averages also may not capture the effects of extreme weather events and can appear to provide water that is not captured. LINKAGES demonstrated that greater temperatures increased evaporation and the number of dry days (i.e., decreased soil water availability). Decreased soil water availability reduced growth and increased mortality of drought-sensitive species on all but the most mesic sites.
We simulated the dominant species in a subsection on a plot and averaged results from the five dominant soils within each subsection to provide a general assessment of model performance. An important application of LINKAGES is to parameterize FLMs such as LANDIS-II (Scheller et al. 2007; Thompson et al. 2011) and LANDIS PRO (Wang et al. 2014, 2015). Forest landscape simulation models which include competition for resources and succession dynamics might more appropriately use results from single species simulations to calculate seedling establishment probabilities (He et al. 1999; Wang et al. 2014, 2015) but growing space available can be modeled by methods described here. Variation in tree species response to variation in soils within subsections indicates that species early growth and possibly growing space should be evaluated at finer scales than at the subsection level. Forest landscape simulation models are often run at the landform level and summarized at the subsection level (Wang et al. 2014). This method would be conducive to running LINKAGES using the dominant soils for each landform and using the results to parameterize the forest landscape model at that spatial scale.
Biomass estimates from the LINKAGES model were higher than the subsection averages from FIA data for simulation years 30, 70 and 100. Although correlations between total biomass at year 70 and 100 with FIA data were significant, R values were low. One factor that likely contributed to this apparent over estimation is LINKAGES does not simulate mortality, species establishment or growth due to disturbances such as wind, fire, disease and insects. Harvest and disturbance have had important effects on CHFR second growth forest (MacCleery 1992) and other model-based approaches that account for disturbance determined harvest and disturbance had greater impacts on forest composition and structure than climate change (Gustafson et al. 2010; Wang et al. 2015). An additional confounding problem is that stand age as described in the FIA database user manual “is difficult to measure” and “may have large measurement errors” (O’Connell et al. 2013). Defining an FIA plot’s stand age can be nebulous since FIA plots can be multi-aged and the classification of FIA plots may represent higher or lower biomass than those of single aged plots. Although LINKAGES overestimates biomass when compared to FIA biomass estimated by stand age, biomass estimates when stand age is derived from age of oldest tree are higher (McGarvey et al. 2015) and more similar to LINKAGES results. Results for current climate biomass from LINKAGES simulations at year 100 ranged between 75 and 196 Mg/Ha with the majority of subsections falling into the range 125–250 Mg/Ha estimated from FIA data in other studies for mature forests within the eastern US (Brown et al. 1997; Schroeder et al. 1997; Jenkins et al. 2001). Total biomass ranges for the western CHFR and the eastern CHFR correspond to biomass levels simulated by the FORENA forest stand model for western deciduous and eastern deciduous forests (Solomon 1986), respectively. Species assemblages for the CHFR are the result of early 20th century disturbance and do not necessarily represent optimum species groups when disturbance is absent.
Interpretation of models should take into consideration model limitations, strengths, and weaknesses. LINKAGES is a hybrid empirical-physiological model with empirical parameters based on tree growth, and mortality relationships determined under current climate; which may not hold true under future climates (Gustafson and Keene 2014), particularly given CO2 fertilization. Similarly, process-based models such as PnET-II rely on equations developed under current conditions and niche-based models rely on statistical relationships resulting from current climate. We believe a strength of LINKAGES is it uses daily climate data, whereas many process-based and niche-based models use monthly climate data and may underestimate the effects of drought. Although LINKAGES provides a direct effect of mortality from drought, it does not incorporate indirect effects such as insect or disease mortality that can be exacerbated by drought (Gustafson and Shinneman 2015). LINKAGES does not model landscape disturbance, management or spatial ecological processes. Thus, some deviations of LINKAGES results are probably due to lack of landscape disturbance, management and spatial ecological processes. One approach to addressing these limitations is to pair LINKAGES with a forest landscape simulation models such as LANDIS PRO (Wang et al. 2015).
Conclusions
Our revised version LINKAGES 3.0 is easier to apply to various landscapes and climates than earlier versions. LINKAGES 3.0 results reflected the full range of spatial heterogeneity present in the soil and climate of the central hardwood region. Our modeling framework provided a useful approach for considering growth potential and maximum growing space under future climates. Soil-climate interactions indicated that LINKAGES 3.0 was sensitive to changes in soil properties. LINKAGES 3.0 uses daily downscaled data so it is likely more sensitive to extreme precipitation and short term drought than models that use monthly or yearly averages which view all precipitation as water available to trees. To increase LINKAGES reliability we suggest future research to calibrate and validate LINKAGES 3.0, investigate its sensitivity to input parameters, and its further application to forest landscape simulation models such as LANDIS. We believe Linkages 3.0 can be a useful tool for estimating parameters such as species establishment and maximum growing space in FLMs such as LANDIS PRO.
References
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SG, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684
Arner SL, Woudenber S, Waters S, Vissage J, MacLean C, Thompson M, Hansen M (2001) National algorithms for determining stocking class, stand size class, and forest type for forest inventory and analysis plots. http://www.fs.fed.us/fmsc/ftp/fvs/docs/gtr/Arner2001.pdf
Botkin DB (1993) Forest dynamics an ecological model, 1st edn. Oxford University Press, New York
Botkin DB, Janak JF, Wallis JR (1972) Some ecological consequences of a computer model of forest growth. J Ecol 60(3):849
Braun EL (1950) Deciduous forests of eastern North America. Blakiston CO., Philadelphia, Toronto, pp 596
Brown SL, Schroeder P, Birdsey R (1997) Aboveground biomass distribution of US eastern hardwood forests and the use of large trees as an indicator of forest development. For Ecol Manag 96:37–47
Brown SL, Schroeder P, Kern JS (1999) Spatial distribution of biomass in forests of the eastern USA. For Ecol Manag 123:81–90
Burns RM, Honkala BH (1990) Silvics of North America: 1. Conifers; 2. Hardwood. Agriculture Handbook 654, USDA Forest Service, Washington, DC
Cleland DT, Freeouf JA, Keys JE, Nowacki GJ, Carpenter C, McNab WH (2007) Ecological subregions: sections of the coterminous United States. USDA forest service, Gen Tech Rep WO-76, Washington DC
Delworth TL, Broccoli AJ, Rosati A, Stouffer RJ, Balaji V, Beesley JA (2006) GFDL’s CM2 global coupled climate models. Part I: formulation and simulation characteristics. J Clim 19:643–674
Federer CA (1995) BROOK90: a simulation model for evaporation, soil water, and streamflow, Version 3.1. USDA Forest Service
Federer CA (2015) The BROOK90 hydrologic model for evapotranspiration, soil water and streamflow. http://www.ecoshift.net/brook/brook90.html
Fralish JS (2003) The central hardwood forest: its boundaries and physiographic provinces, Proceedings 13th central hardwoods conference. In: Van Sambeek JW, Dawson JO, Ponder F, Lowenstein EF, Fralish JS (eds) USDA Forest Service, North Central Research Station, Gen Tech Rep NC-234, St. Paul
Gu L, Pallardy SG, Hosman KP, Sun Y (2015) Drought influenced mortality of tree species with different predawn leaf water dynamics in a decade-long study of a central US forest. Biogeosciences 12:2831–2845
Gustafson EJ, Keene R (2014). Predicting changes in forest composition and dynamics—landscape-scale process models. U.S. Department of Agriculture, Forest Service, Climate Change Resource Center
Gustafson EJ, Shinneman DJ (2015) Approaches to modeling landscape-scale drought-induced forest mortality. In: Perera AH, Sturtevant BR, Buse LJ (eds) Simulation modeling of forest landscape disturbances, Chap 3. Springer, New York, pp 45–71
Gustafson EJ, Shvidenko AZ, Sturtevant BR, Scheller RM (2010) Predicting global change effects on forest biomass and composition in south-central Siberia. Ecol Appl 20:700–715
Guyette R, Kabrick JM (2000) The legacy and continuity of forest disturbance, succession, and species at the MOFEP sites. In: Shifley SR, Kabrick JM (eds) Proceedings of the second Missouri Ozark forest ecosystem project symposium: post treatment results of the landscape experiment. U.S. Department of Agriculture, Forest Service, GTR NC-227 26-44
He HS (2008) Forest landscape models, definition, characterization, and classification. For Ecol Manag 254:484–498
He HS, Mladenoff DJ (1999) Spatially explicit and stochastic simulation of forest landscape fire disturbance and succession. Ecology 80:81–99
He HS, Mladenoff DJ, Crow TR (1999) Linking an ecosystem model and a landscape model to study forest species response to climate warming. Ecol Model 114:213–233
IPCC (2013) Annex III: glossary [Planton S. (ed.)]. In: Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Iverson LR, Prasad AM, Matthews SN, Peters M (2008) Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For Ecol Manag 254:390–406
Iverson LR, Thompson FR III, Mathews S, Peters M, Prasad A, Dijak WD, Fraser JS, Wang WJ, Hanberry B, He HS, Janowiak M, Butler P, Brandt L, Swanston C (2016) Multi-model comparison on the effects of climate change on tree species in the eastern U.S.: results from an enhanced niche model and process-based ecosystem and landscape models. Landscape. doi:10.1007/s10980-016-0404-8
Jenkins JC, Birdsey RA, Pan Y (2001) Biomass and NPP estimation for the mid-Atlantic region (USA) using plot-level forest inventory data. Ecol Appl 11:1174–1193
Johnson PS, Shifley SR, Rogers R (2009) The ecology and silviculture of oaks, 2nd edn. CAB International, Oxfordshire
Kabrick JM, Jensen RG, Shifley SR, Larsen (2002) Woody vegetation following even-aged, uneven-aged and no-harvest treatments on the Missouri Ozarks forest ecosystem project sites. USDA General Technical Report NC-227
Li X, Niu J, Xie B, Johnson SJ (2013) Study on hydrological functions of litter layers in North China. PLoS ONE 8(7):e70328
Lischke H, Loffler TJ, Fischlin A (1998) Forest succession models: capturing variability with height structured, random, spatial distributions. Theor Popul Biol 54:213–226
Lischke H, Zimmermann NE, Bolliger J, Rickebusch S, Loffler TJ (2006) TreeMig: a forest-landscape model for simulating spatio-temporal patterns from stand to landscape scale. Ecol Model 199:409–420
Little EL Jr (1971) Atlas of United States trees, volume 1, conifers and important hardwoods: U.S. Department of Agriculture Miscellaneous Publication 1146, 200 maps
Little EL Jr (1977) Atlas of United States trees, volume 4, minor Eastern hardwoods: U.S. Department of Agriculture Miscellaneous Publication 1342, 230 maps
Livneh B, Rosenberg EA, Lin C, Nijssen B, Mishra V, Andreadis KM, Maurer EP, Lettenmaier DP (2013) A long-term hydrologically based dataset of land surface fluxes and states for the conterminous United States: update and extensions. J Clim 26:9384–9392
MacCleery DW. (1992) American forests: a history of resilience and recovery. USDA forest Service FS-540
McGarvey JC, Thompson JR, Epstein HE, Shugart HH Jr (2015) Carbon storage in old-growth forests of the Mid-Atlantic: toward better understanding the eastern forest carbon sink. Ecology 96:311–317
Morin X, Thuiller W (2009) Comparing niche- and process-based models to reduce prediction uncertainty in species range shifts under climate change. Ecology 90:1301–1313
Morin X, Viner D, Chuine I (2008) Tree species range shifts at a continental scale: new predictive insights from a process-based model. J Ecol 96(4):784–794
Niinemets U, Valladares F (2006) Tolerance to shade, drought and waterlogging of temperate northern hemisphere trees and shrubs. Ecol Monogr 76:521–547
O’Connell, BM, LaPoint EB, Turner JA, Ridley T, Boyer D, Wilson AM, Waddell KL, Conkling BL (2013) The forest inventory and analysis database: database description and user guide for Phase 2 (version 6.0.2). http://www.fia.fs.fed.us
Pastor J, Post WM (1985) Development of a linked forest productivity-soil process model. In: ORNL/TM-9519. Oak Ridge National Laboratory, Oak Ridge
Purves D, Pacala S (2008) Predictive models of forest dynamics. Science 320(5882):1452–1453
Reclamation (2014) http://cmip-pcdmi.llnl.gov/cmip5/availability.html
Reich PB, Frelich LE (2001) Temperate deciduous forests. Encyclopedia of global change. Macmillan Reference USA, Biology for students
Saxton KE, Rawls WJ, Romberger JS, Papendick RI (1986) Estimating generalized soil-water characteristics from texture. Soil Sci Soc Am J 50:1031–1036
Scheller RM, Mladenoff DJ (2005) A spatially dynamic simulation of the effects of climate change, harvesting, wind, and tree species migration on the forest composition, and biomass in northern Wisconsin, USA. Glob Change Biol 11:307–321
Scheller RM, Van Tuyl S, Clark KL, Hayden NG, Hom J, Mladenoff DJ (2007) Simulation of forest change in the New Jersey Pine Barrens under current and pre-colonial conditions. For Ecol Manage 255:1489–1500
Schneiderman JE, He HS, Thompson FR, Dijak WD, Fraser JS (2015) Comparison of a species distribution model and a process model from a hierarchical perspective to quantify effects of projected climate change on tree species. Landscape Ecol 30(10):1879–1892
Schroeder P, Brown S, Mo JM, Birdsey R, Cieszewski C (1997) Biomass estimation for temperate broadleaf forests of the United States using inventory data. For Sci 42:424–434
Schumacher S, Bugmann H, Mladenoff DJ (2004) Improving the formulation of tree growth and succession in a spatially explicit landscape model. Ecol Model 180:175–194
Seidl R, Rammer W, Scheller RM, Spies T (2012) An individual-based process model to simulate landscape-scale forest ecosystem dynamics. Ecol Model 231:87–100
Shifley SR, He HS, Lischke H, Wang W, Jin W, Gustafson EJ, Thompson JR, Thompson FR III, Dijak WD, Wang J (in press) The past and future of modeling forest dynamics: from growth and yield curves to forest landscape models. Landscape Ecology
Shugart HH, West DC (1980) Forest succession models. BioScience 30(5):308–313
Solomon AM (1986) Transient response of forest to CO2-induced climate change: simulation modeling experiments in eastern North America. Oecologia 68:567–579
Taylor JA (1967) Growing season as affected by land aspect and soil texture. In: Taylor JA (ed) Weather and agriculture. Pergamon Press, Oxford, pp 15–36
Thompson JR, Foster DR, Scheller RM, Kittredge D (2011) The influence of land use and climate change on forest biomass and composition in Massachusetts, USA. Ecol Appl 21(7):2425–2444
Thornton PE, Thornton MM, Mayer BW, Wilhelmi N, Wei Y, Deveraconda R, Cook RB (2016) Daymet: daily surface weather data on a 1 km grid for North America, 1980–2012. (http://daymet.ornl.gov/) Accessed 13 Jul 2016, Oak Ridge National Laboratory Distributed Active Archive Center, Oak Ridge
United States Department of Agriculture, Natural Resources Conservation Service. (2006) Land Resource Regions and Major Land Resource Areas of the United States, the Caribbean, and the Pacific Basin. U.S. Department of Agriculture Handbook 296
Vanclay JK (1991) Aggregating tree species to develop diameter increment equations for tropical rainforests. For Ecol Manag 42:143–168
Vanderwel MC, Purves DW (2014) How do disturbances and environmental heterogeneity affect the pace of forest distribution shifts under climate change? Ecography 37:10–20
Wang WJ, He HS, Spetich MA, Shifley SR, Thompson FR III (2014) LANDIS PRO: a landscape model that predicts forest composition and structure changes at regional scales. Ecography 37:225–229
Wang WJ, He HS, Thompson FR III, Fraser JS, Hanberry BB, Dijak WD (2015) Importance of succession, harvest, and climate change in determining future composition in U.S. central hardwood forests. Ecosphere 6(12):1–18
Wang WJ, He HS, Thompson FR III, Fraser JS, Dijak WD (2016) Changes in forest biomass and tree species distribution under climate change in the northeastern United States. Landscape Ecol. doi:10.1007/s10980-016-0429-z
Way D, Montgomery R (2015) Photoperiod constraints on tree phenology, performance and migration in a warming world. Plant Cell Environ 38:1725–1736
Wullschleger SD, Gunderson CA, Tharp ML, West DC, Post WM (2003) Simulated patterns of forest succession and productivity as a consequence of altered precipitation. In: Hanson PJ, Wullschleger SD (eds) North American temperate deciduous forest responses to changing precipitation regimes. Springer, New York, pp 433–446
Yukimoto S, Adachi Y, Hosaka M, Sakami T, Yoshimura H, Hirabara M, Tanaka TY, Shindo E, Tsujino H, Deushi M, Mizuta R, Yabu S, Obata A, Nakano H, Koshiro T, Ose T, Kitoh A (2012) A new global climate model of the meteorological research institute: MRI-CGCM3—model description and basic performance. J Meterol Soc Japn 90A:23–64
Acknowledgements
We thank Stanley Wullschleger and Wilfred Post for help in initializing LINKAGES v2.2 and Stephen Shifley, John Kabrick and Dan Dey for support and knowledge provided about forest ecology and forest soils. We thank Steve Pallardy and Oak Ridge National Laboratory for access to Ameriflux data. This project was funded by the U.S.D.A. Forest Service Northern Research Station, a cooperative agreement with the United States Geological Survey Northeast Climate Science Center, Department of Interior USGS Northeast Climate Science Center graduate and post-graduate fellowships, and the University of Missouri-Columbia. Its contents are solely the responsibility of the authors and do not necessarily represent views of the Northeast Climate Science Center or the USGS. This manuscript is submitted for publication with the understanding that the United States Government is authorized to reproduce and distribute reprints for Governmental purposes.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Dijak, W.D., Hanberry, B.B., Fraser, J.S. et al. Revision and application of the LINKAGES model to simulate forest growth in central hardwood landscapes in response to climate change. Landscape Ecol 32, 1365–1384 (2017). https://doi.org/10.1007/s10980-016-0473-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0473-8