
Abstract
Context
Detailed information on habitat needs is integral to identify conservation measures for declining species. However, field data on habitat structure is typically limited in extent. Remote sensing has the potential to overcome these limitations of field-based studies.
Objective
We aimed to assess abiotic and biotic characteristics of territories used by the declining wood warbler (Phylloscopus sibilatrix), a forest-interior migratory passerine, at two spatial scales by evaluating a priori expectations of habitat selection patterns.
Methods
First, territories established by males before pairing, referred to as pre-breeding territories, were compared to pseudo-absence control areas located in the wider forested landscape (first spatial scale, Nterritories = 66, Ncontrols = 66). Second, breeding territories of paired wood warblers were compared to true-absence control areas located immediately close-by in the forest (second spatial scale, Nterritories = 78, Ncontrols = 78). Habitat variables predominantly described forest structure and were mainly based on first and last pulse lidar (light detection and ranging) data.
Results
Occurrence of pre-breeding territories was related to vegetation height, vertical diversity and stratification, canopy cover, inclination and solar radiation. Occurrence of breeding territories was associated to vegetation height, vertical diversity and inclination.
Conclusions
Territory selection at the two spatial scales addressed was governed by similar factors. With respect to conservation, habitat suitability for wood warblers could be retained by maintaining a shifting mosaic of stand ages and structures at large spatial scales. Moreover, leaf-off lidar variables have the potential to contribute to understanding the ecological niche of species in predominantly deciduous forests.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Detailed knowledge of the habitat needs of declining species is crucial for various applications, including habitat protection (e.g. Whittingham et al. 2005), the restoration of species to previously occupied habitat (Merrill et al. 1999), the identification of dispersal corridors (Chetkiewicz and Boyce 2009) or for predicting species distribution and locating potentially suitable habitat (e.g. Luoto et al. 2002). Studies on habitat needs and habitat suitability require information on potentially influential variables measured at the appropriate scale (spatial grain and extent). However, information on habitat variables based on field data is often limited because data collection is usually time consuming and spatially restricted. Remote sensing (hereafter RS) has therefore become an important tool for mapping ecosystems at broad spatial scales while retaining a high level of detail, thus allowing improved modeling of, for example, species’ distribution and potential habitats (e.g. Sperduto and Congalton 1996). RS includes a wide range of technologies ranging from airborne sensors to satellite-based information, global positioning systems or camera tracks (Pettorelli et al. 2014). Among others, methods currently used for modeling and mapping species distribution and richness are image texture (e.g. Bellis et al. 2008; Wood et al. 2013; Duncan et al. 2014; St-Louis et al. 2014), spectral mixture analysis (e.g. St-Louis et al. 2014) or image classification (e.g. Duncan et al. 2014). Typically, the RS data used in such models does not allow characterizing vertical habitat structure (Lefsky et al. 2002; Vierling et al. 2008). However, many species, especially forest bird species, are associated with specific three-dimensional habitat structures (e.g. Dunlavy 1935; Shaw et al. 2002). In contrast to other RS methods, light detection and ranging (lidar) is a source of high-resolution geospatial data, providing three-dimensional information about micro-topography and the structure of ecosystems over broad spatial extents (Vierling et al. 2008; Müller et al. 2009).
With regard to lidar-derived assessment of forest habitat suitability, species occurrence has successfully been investigated for various animal taxa such as mammals (Nelson et al. 2005; Coops et al. 2010; Jung et al. 2012; Palminteri et al. 2012) or arthropods (Müller and Brandl 2009; Vierling et al. 2011; Müller et al. 2014). In birds, forest habitat suitability based on lidar data has been estimated for several species, including grouse (Graf et al. 2009; Zellweger et al. 2013), owls (Zellweger et al. 2013), woodpeckers (Smart et al. 2012; Trainor et al. 2013; Vierling et al. 2013; Zellweger et al. 2013) and passerines (Broughton et al. 2006, 2012; Bellamy et al. 2009; Goetz et al. 2010; Farrell et al. 2013; Vogeler et al. 2013; Reidy et al. 2015).
Here, we address habitat selection of the wood warbler (Phylloscopus sibilatrix) at two spatial scales using RS data, and particularly lidar data. The species is a near-endemic to Europe classified as “least concern” (BirdLife International 2015). Nevertheless, the wood warbler has suffered long-term declines in many EU countries (EIONET 2015). Upon return from their sub-Saharan wintering areas (Hobson et al. 2014), male wood warblers start settling on the breeding grounds in early to mid-April, while females arrive on average 10 days later (Glutz von Blotzheim and Bauer 1991; Wesolowski and Maziarz 2009). Before getting paired, pre-breeding territories defended by males are 1–3 ha in size (Glutz von Blotzheim and Bauer 1991). After pairing has occurred, territories used for breeding average 1200–1900 m2 (Glutz von Blotzheim and Bauer 1991). Within these breeding territories, females select a nest site on the ground (Glutz von Blotzheim and Bauer 1991). To meet the species’ needs, a forest must thus offer suitable conditions at the pre-breeding territory scale as well as at the breeding territory scale. That habitat selection can differ depending on the spatial scale considered has long been recognized (Johnson 1980; Hutto 1985; Bunnell and Huggard 1999). However, while seasonal variation in territory size (e.g. Finck 1990; Möller 1990; Pasinelli et al. 2001; Wiktander et al. 2001) and seasonal changes in habitat selection (e.g. Wikar et al. 2008; Brambilla et al. 2012) have been reported, particularly for non-migratory species, surprisingly little is known about whether habitat selection by migratory songbirds prior to breeding (i.e. after settlement) and during breeding (i.e. after pair formation and during brood rearing) differs. Knowing what constitutes suitable habitats at the pre-breeding stage and the breeding stage, respectively, and managing for these habitat types, would ensure that habitats suitable for both settlement and breeding will be preserved. It is possible that the same habitat types or structures affect both settlement and breeding, as breeding territories are contained within pre-breeding territories. However, habitat types or structures in breeding territories might differ from the larger pre-breeding territories, for example when breeding territories need to provide specific food resources and/or specific habitat structure promoting successful breeding. We thus addressed the following questions: (1) do the habitat characteristics of pre-breeding territories differ from what is available in the wider forested landscape? And (2) do the habitat characteristics of breeding territories differ from what is available in the immediate forest vicinity? Based on known habitat requirements, we evaluated a priori expectations of the relationships between wood warbler occurrence and habitat factors (see Methods—“Expectations” section).
Methods
Study species
The wood warbler is a ground-nesting, long-distance migratory passerine with a distinctly European range. The species selects habitats predominantly located in broadleaf and mixed deciduous forest at low to medium altitudes. Within the forest, stands are used that feature trees with a height of at least 8–10 m, an open stem space and a rather closed canopy (Quelle and Tiedemann 1972; Glutz von Blotzheim and Bauer 1991). Older stands (Quelle and Tiedemann 1972; Reinhardt and Bauer 2009; Pasinelli et al. 2016) and stands with heterogeneous tree height (Pavlovic 2009) are suggested to be less suitable. A lush herb and shrub layer is avoided, while grass and sedge tussocks are used (Quelle and Tiedemann 1972; Schifferli et al. 1980; Bibby 1989; Glutz von Blotzheim and Bauer 1991; Pavlovic 2009; Reinhardt and Bauer 2009; Mallord et al. 2012; Pasinelli et al. 2016). Several studies reported a preference for inclined areas with eastern or southerly aspects (Quelle and Tiedemann 1972; Glutz von Blotzheim and Bauer 1991; Mallord et al. 2012; Pasinelli et al. 2016).
Study areas
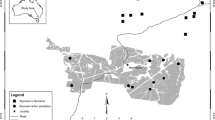
We initially determined areas with a relatively high abundance of wood warblers in the 10 years prior to 2010 throughout northern Switzerland based on (a) the common breeding bird survey provided by the Swiss Ornithological Institute (the standardized Swiss national bird monitoring program, www.vogelwarte.ch/monitoring-common-breeding-birds.html), (b) the breeding bird atlas of the canton Zurich (www.birdlifezuerich.ch), and (c) www.ornitho.ch (the official birding exchange platform in Switzerland). However, during early field work in 2010 (Gerber 2011, Grendelmeier et al. 2015), it became evident that some of these areas no longer had any breeding birds, leaving us with 16 intensively searched study areas (Fig. 1). These study areas ranged in size from 0.18 to 2.35 km2 (Appendix 1—Supplementary material) and were all part of large woodlands consisting of deciduous and mixed-forest stands dominated by European beech (Fagus sylvatica), with coniferous tree species interspersed (Picea abis, Abies alba, Pinus sylvestris). Boundaries of the study areas were determined by landscape elements such as forest edge, valleys or strong changes in forest structure (e.g. young re-growths or coniferous plantations). Elevation of the study areas ranges between 543 and 1022 m a.s.l. Study areas are characterized by cold winters, warm and wet summer months and mean yearly precipitation rates from 800 to 1500 mm (MeteoSchweiz 2014). Mean annual minimum and maximum temperature is 5.4 and 11.8 °C, respectively.

Location of the study areas (plus) in northern Switzerland (rectangular insert): 1 Bänkerjoch AG, 2 Belchen SO, 3 Blauen BL, 4 Dittingen BL, 5 Ennenda GL, 6 Erschwil SO, 7 Gündelhart TG, 8 Homberg SO, 9 Hochwald SO, 10 Kleinlützel SO, 11 Langenbruck BL, 12 Lauwil BL, 13 Montsevelier JU, 14 Oltingen BL, 15 Staffelegg SO, 16 Scheltenpass SO
Bird mapping procedure
Wood warbler occurrence was mapped between 2010 and 2012 annually from mid-April on by listening for the distinct wood warbler song. In all 16 intensively searched study areas, we used the coordinates of previous sightings as rough starting points, from where we extensively searched for the species. If no wood warblers were heard or observed, we played 10 s playbacks of wood warbler songs every 300 m to avoid overlooking territories. As soon as birds responded, we stopped the playback. On average, each study area was checked once a week until early July. Once territories had been established, they were regularly checked for the presence of females and nests. The nest position was mapped with a handheld GPS. Due to the regular observer presence, a narrow search grid and the use of playback, it is highly probable that all nests within a study area were found.
Definition of sample areas
We defined two types of sample areas differing in their occupation state, namely territories (occupied) and control areas (unoccupied). Because no exact information about the location of the nest relative to the defended area was available, we approximated territories by circles centered on the nest. The nest location was thus used as a proxy for the center of a territory. We did not aim at measuring nest-site selection within territories (fourth order selection sensu Johnson 1980). Pre-breeding territory size of the wood warbler is reported to be approximately 10,000–30,000 m2, corresponding to a circle with radius of at least 56 m. Breeding-territory size ranges from 1200–1900 m2, corresponding to a circle with mean radius of 22.25 m (Glutz von Blotzheim and Bauer 1991). To reduce the risk of including unsuitable habitat, radii were reduced to 80 % of the sizes given above. Thus, pre-breeding-territory size was approximated by a circular area of 6648 m2 and breeding territory size by a circular area of 1000 m2, both centered on the nest location.
To compare habitat characteristics of pre-breeding and breeding territories to what is available in the wider forested landscape (question 1) and in the immediate forest vicinity (question 2), respectively, two types of control areas were defined. In the following, we refer to control areas relating to question 1 as “pseudo-absences” and to control areas relating to question 2 as “field-absences”. To address question 1, for each pre-breeding territory in a specific study area, one pseudo-absence was randomly selected outside the intensively searched study area based on the following criteria: (1) located in woodland, not recently affected by large, natural disturbance (e.g. fire, wind throw), (2) located at a distance between 500 and 2000 m from the nest of the respective territory, (3) located at a distance of at least 500 m from all the other nests and centers of the field-absences (see below) of the respective study area. A range of distances was needed to fulfill criteria (1) and (3). The choice of the distance is subject to a trade-off. To allow for increased environmental variation, the distance should be as large as possible. However, with increasing distance, the probability rises that possibly existing territories are mistakenly selected as pseudo-absence. A qualitative assessment following the selection process revealed that 98 % of the defined pseudo-absences did not correspond to the species’ typical habitat description (Alex Grendelmeier, pers. comm.). To address question 2, for each breeding territory, one field-absence was defined within the intensively searched study area with its center located 200–300 m from the nest position. First, eight possible field-absences (i.e. X–Y-coordinates) 200–300 m from the nest of the respective breeding territory were defined in the four cardinal (N, E, S, W) and four intercardinal (NE, SE, SW and NW) directions. To avoid trivial results, field-absences with habitats known to be unsuitable for wood warblers (non-forest areas, forest clearings, purely coniferous forest patches, young re-growths and plantations) were ruled out. Also, field-absences closer than 50 m to other breeding territories in the same year were excluded. Of the non-excluded field-absences, one was randomly selected. Absence of wood warblers in retained field-absences was confirmed with playback.
Because some territories partly overlapped among the three study years, analyses were restricted to territories not overlapping more than 10 % with each other. Selection of overlapping territories followed two principles: (1) if two territories overlapped more than 10 % with one another, one of the two was randomly selected for use in the analyses, the other one discarded; (2) if more than two territories overlapped more than 10 % with one another, territories to be excluded were selected so as to minimize the total number of territories excluded. The same principles applied to field-absences overlapping between study years. We ended up with 66 pre-breeding territories paired to 66 pseudo-absences and 78 breeding territories paired to 78 field-absences (Appendix 1—Supplementary material).
RS variables
The RS data was processed in the Geographic Information System ESRI ArcInfo, PyScripter for Python and R. LAStools were used for the operational processing of lidar data (Isenburg 2012). The RS variables (Table 1) were grouped into lidar variables and non-lidar RS variables, respectively, and assigned to five thematic groups that comprised variables measuring similar aspects of the environment. All variables were calculated for both pre-breeding and breeding territories, respectively, and their associated control areas.
Lidar variables
First and last return lidar data was nationally available in the form of classified point clouds, namely the digital terrain model (DTM) and the digital surface model (DSM) (DTM-AV, DOM-AV © 2014 swisstopo 5704 000 000; swisstopo 2012). Mean point density was 1.5 m−2 for the sample areas analyzed. The standard deviation of height accuracy was 1.5 m in forested areas, as reported by the data provider. The interpolation of the DTM into a regular 2 m grid yields the swissALTI3D (swisstopo 2013). In addition, a normalized digital surface model (nDSM), representing vegetation height, was calculated by subtracting the DSM from the swissALTI3D. Values deviating negatively by three or more times the standard deviation from the mean of all negative values of the nDSM were treated as outliers and excluded from further analyses (Grubbs 1969).
For 14 out of 16 study areas, lidar data were collected in 2001, for the other two in 2002. Eleven study areas were flown in February, four in April, and one in December and January, respectively. Forest structure determined from lidar data very likely reflects conditions during wood warbler territory mapping conducted 10 years later because areas strongly impacted by disturbances were not suitable for territory establishment anyway and, if disturbances existed, excluded as both field-absences and pseudo-absences (see above). Moreover, most study areas are located in forests managed last 20–50 years ago (Duc et al. 2010). Further, a case study showed that a 6-year difference between the collection of lidar data and bird data had a minimal effect on mapped avian distribution patterns in an undisturbed coniferous forest (Vierling et al. 2014). That lidar data collection occurred outside the growing season could be critical in terms of tree species composition (deciduous versus non-deciduous) and temporal displacement between habitat selection and habitat description. For deciduous trees, lidar signals under leaf-off conditions can only be intercepted by branches and stems, leading to a higher proportion of ground reflections than under leaf-on conditions (Brandtberg et al. 2003). Further, under leaf-off conditions, variability of canopy height is significantly higher and many canopy height and density measures are lower for deciduous forests compared to leaf-on conditions (Brandtberg et al. 2003; Næsset 2005). In contrast, non-deciduous trees will not be affected by changes from leaf-off to leaf-on conditions (Ørka et al. 2010). For the application in this study, however, the most important issue is that relative differences in forest structure characteristics are captured by the lidar variables (i.e. a closed stand can be distinguished from a scattered stand). Moreover, recent studies demonstrated that leaf-off lidar sampled for the purpose of terrain mapping is valuable for predicting forest characteristics even for deciduous forests (Hawbaker et al. 2010; Wasser et al. 2013; Parent and Volin 2014).
Height and vertical diversity variables
Variables of this thematic group describe vegetation height (VH) and canopy height (CH), and the vertical diversity within VH and CH. VH equates to the nDSM. CH is represented by the maximum lidar signal for each cell in a 2 × 2 m grid laid over every sample area. On this basis, the following variables were calculated for the pre-breeding and breeding territories as well as their associated control areas (Table 1): mean vegetation height (meanVH), mean canopy height (meanCH), maximum vegetation height (maxVH), the 95 % percentile of vegetation height (VH95), the standard deviation of vegetation height (sdVH) and the standard deviation of canopy height (sdCH). MeanVH is an index of vegetation height in general, whereas meanCH refers specifically to the upper canopy layer. The standard deviation provides a measure of the vertical variation of VH and CH, respectively. A small standard deviation arises from sample areas with a homogeneous vegetation or tree height, while a high standard deviation reflects a heterogeneous vegetation or tree height (Müller et al. 2009). Additionally, the following variables refer to the tree layer: mean vegetation height above 3 m (meanVH>3m), mean canopy height above 3 m (meanCH>3m) and the standard deviation of canopy height above 3 m (sdCH>3m). These three variables were calculated including only laser signals reflecting from objects more than 3 m above ground. Mean vegetation height less than 3 m (meanVH<3m) refers to the shrub and regeneration layer and is represented by the arithmetic mean of all lidar signals less than 3 m above ground.
Vertical stratification variables
Stratification describes the distribution of different life forms or age classes existing at specific heights, e.g. trees, shrubs and herbs (Parker and Brown 2000; Park 2007). In this study, three vegetation layers were analyzed based on their penetration rates: pen50_2 refers to the tree layer, pen10_2 to the mid-story and pen5_1 to the shrub and regeneration layer. A high penetration rate indicates a sparse development of the corresponding vegetation layer, while a low penetration rate originates when the vegetation layer of interest is well developed (Müller et al. 2009). To calculate pen50_2, the sum of all lidar signals below 2 m above ground was divided by the sum of all lidar signals below 50 m above ground. Pen10_2 is described by the ratio of the sum of all lidar signals below 2 m and the sum of all lidar signals below 10 m. Pen5_1 equals to the ratio of the sum of all lidar signals below 1 m and the sum of all lidar signals below 5 m (Müller et al. 2009).
Canopy cover variables
This group contains variables describing canopy cover at four height levels, namely at 3 m (meanCC), 10 m (meanCC_10m), 15 m (meanCC_15m) and 20 m (meanCC_20m) above ground. The calculation of canopy cover was based on the data of CH. The respective canopy cover results from the ratio of the number of 2 × 2 m cells with values above the particular height level and the total number of 2 × 2 m cells per sample area. Canopy cover is higher, the more lidar signals are present above the particular height level.
Geography-related variables
This group only comprises one lidar variable, namely slope. Slope values were derived from the swissALTI3D and averaged per sample area.
Non-lidar RS variables
In addition to the lidar variables, the following non-lidar RS variables were calculated and assigned to the geography-related variables: potential direct solar radiation in March (r_march), forest type (forest_type) and distance to forest edge (dist_f). R_march values were derived for from the digital height model DHM25 © 2014 swisstopo 5704 000 000 (swisstopo 2015) following Zimmermann and Kienast (1999). Values were resampled to a resolution of 5 × 5 m and averaged per sample area. Forest_type values were derived from the vegetation classification provided by the Swiss Federal Statistical Office (2010), classifying 25-m forest squares into the four categories coniferous, coniferous mixed, broadleaf mixed and broadleaf (BFS GEOSTAT). The classification values were resampled to a resolution of 5 × 5 m and averaged per sample area. Dist_f represents the shortest distance from the center of the sample area (i.e. nest locations for territories and center points for control areas) to forest edge, extracted from the swissTLM3D 1.1 © 2014 swisstopo 5704 000 000 (swisstopo 2014).
Expectations
Given the habitat requirements summarized above (Methods/Study species) and the RS variables just introduced, a priori expectations were derived for the probability of wood warbler occurrence and potentially influential habitat variables (Fig. 2; Table 1). Since habitat needs of this species addressed in the past have not distinguished pre-breeding and breeding territory selection, no scale-specific expectations were possible.

Expected relationships a priori derived from the literature between independent variables (x-axis) and the probability of wood warbler occurrence (y-axis). Bold: names of thematic groups and associated expectations (numerals)
Data analysis
To analyze if habitat characteristics of pre-breeding territories (N = 66) and breeding territories (N = 78) differ from what is available in the forested landscape and in the immediate forest vicinity, respectively, pre-breeding territories were compared to their associated pseudo-absences 500–2000 m away (see above), while breeding territories were compared to their associated field-absences 200–300 m away. Generalized linear mixed-effect models (GLMMs) with logit link and assuming a binomial error distribution were applied with two random effects, namely study area and territory-control pairs nested within study area. For all analyses, the binomial response variable was specified as unoccupied control area (coded as 0) versus either pre-breeding territory or breeding territory (coded as 1 in either case). All RS variables were standardized (mean = 0, standard deviation = 1) prior to analyses.
Model selection
Model selection and model averaging were based on Akaike Information Criterion (AIC) (Akaike 1974; Everitt 2002), following an approach described by Burnham and Anderson (2002). This multi-model inference approach accounts for model selection uncertainty and leads to more robust inferences compared to inferences based on only one single best model (Burnham and Anderson 2002). For this study, AIC corrected for small sample size (AICc) and AICc differences (ΔAICc) were calculated for all candidate models (Hurvich and Tsai 1989). Models with ΔAICc values <2 compared to the best supported one are considered to have similar support, while the others are considered to be less supported by the data (Burnham and Anderson 2002). In addition to ΔAICc values, model-averaged estimates, model-averaged standard errors (SE) and 95 % confidence intervals (95 % CI) were calculated for all variables. A 95 % CI excluding 0 indicates that the occurrence of the study species is either positively or negatively related to the respective variable (Mazerolle 2006).
For each thematic group, firstly, a set of candidate models was constructed (within-group analysis). These sets consisted of models with all variable combinations of the particular group, except that variables correlating more than |0.5| were never included together in the same model, but were instead evaluated in separate (but otherwise identical) models (Appendix 2—Supplementary material). Interactions between the variables were not considered. According to the expectations formulated a priori (Fig. 2), quadratic effects (x ± x2) were considered for meanVH, meanCH, meanVH>3m, meanCH>3m, maxVH, VH95, pen50_2, meanCC, meanCC_10m, meanCC_15m and meanCC_20m. The null model included the intercept and the two random effects. No more than six RS variables were analyzed jointly in the same model. Including more than six variables could have resulted in overfitting of the models, given the sample sizes available. A variable was considered relevant if 1) it was included in the best supported model or in a model with a ΔAICc value <2 compared to the best supported one and 2) had a model-averaged estimate across all models per group larger than its model-averaged SE. Secondly, an across-group analysis was performed including only the relevant variables of each thematic group. The candidate model set consisted of models with all combinations of the relevant variables. Again, variables correlating more than |0.5| were never included together in the same model, and six RS variables at most were jointly analyzed in the same model.
Model fit
Predictive performance and robustness of the best performing models were evaluated based on area under the receiver operating characteristic curve (AUC) and true skill statistic (TSS). AUC is a threshold-independent measure indicating the model’s ability to discriminate presences and absences (Zweig and Campell 1993; Fielding and Bell 1997). Following labels were assigned to AUC values: 0.5 = no discrimination, 0.7–0.79 = acceptable, 0.8–0.9 = excellent, and >0.9 = outstanding (Hosmer and Lemeshow 2000). TSS measures overall model performance by correcting the overall accuracy of model predictions by the accuracy expected to occur by chance. TSS is a special case of Cohen’s kappa (Cohen 1960), but not dependent on prevalence, which is described by the proportion of observed presences. TSS is defined as sum of sensitivity (proportion of observed presences correctly predicted) and specificity (proportion of observed absences correctly predicted) less one (Allouche et al. 2006). Following labels were assigned to the ranges of Cohen’s kappa: <0.00 = poor, 0.00–0.20 = slight, 0.21–0.40 = fair, 0.41–0.60 = moderate, 0.61–0.80 = substantial, 0.81–1.00 = almost perfect (Landis and Koch 1977).
The statistical analyses were conducted in R (https://www.r-project.org/) using the packages AICcmodavg (Mazerolle 2012), lme4 (Bates et al. 2012) and PresenceAbsence (Freeman 2012).
Results
Pre-breeding territories versus pseudo-absences
Within-group analysis
For each thematic group, the best supported models (ΔAICc < 2) always received strong support relative to the null model (Appendix 4—Supplementary material). In the vegetation height group, these models included maxVH, meanVH<3m, and the quadratic effects of meanCH and maxVH. Further, these models included sdVH, sdCH>3m, pen50_2, pen5_1, meanCC_10m, meanCC_20m, slope, r_march and dist_f, and the quadratic effects of pen50_2 and meanCC_20m. After considering the model-averaged estimates and SE, the following variables were retained for the across-group analyses: the quadratic effects of meanCH, maxVH and pen50_2, and the linear effects of maxVH, sdVH, sdCH>3m, pen50_2, pen5_1, meanCC_10m, meanCC_20m, slope, r_march and forest_type.
Across-group analysis
The best supported models had ΔAICc values >74.7 to the null model and included sdVH, pen50_2, pen5_1, meanCC_10m, meanCC_20m, slope, r_march and dist_f (Table 2). According to the 95 % CI (Table 3), pre-breeding territory occurrence was negatively related to maxVH, sdVH, sdCH>3m and meanCC_20m, and positively related to pen50_2, meanCC_10m, slope and r_march, respectively (Fig. 3). Further, pre-breeding territory occurrence showed a negative quadratic relationship with both meanCH and maxVH.

Wood warbler pre-breeding territory occurrence in relation to the variables of the best supported GLMM (filled circle territories, open circle pseudo-absences). Model-averaged estimates of the across-group analysis were used as coefficients. Lines represent the fitted responses for the respective variable (where the other variables were set to their means). Variable description is available in Table 1. Nterritories = 66, Npseudo-absences = 66
Breeding territories versus field-absences
Within-group analysis
For the breeding territories, results of within-group model selection were similar to those of the pre-breeding territories (Appendix 4—Supplementary material). However, the best supported models neither included pen5_1, r_march and dist_f nor the quadratic effects of meanCH and meanCC_20m. However, meanVH and forest_type, and the quadratic effect of VH95 appeared in the best supported models. The following variables were kept for the across-group analyses: meanVH, maxVH, meanVH<3m, sdVH, sdCH>3m, pen50_2, meanCC_10m, meanCC_20m, slope, forest_type and the quadratic effect of VH95.
Across-group analysis
The best supported models had ΔAICc > 58.1 to the null model and included meanVH, meanVH<3m, maxVH, sdVH, sdCH>3m, pen50_2, slope, forest_type and the quadratic effect of VH95 (Table 2). Breeding territory occurrence was negatively related to meanVH, maxVH, sdVH, sdCH>3m and meanCC_20m and positively related to slope, respectively (Fig. 4; Table 3).

Wood warbler breeding territory occurrence in relation to the variables of the best supported GLMM (filled circle territories, open circle field-absences). Model-averaged estimates of the across-group analysis were used as coefficients. Lines represent the fitted responses for the respective variable (where the other variables were set to their means). Variable description is available in Table 1. Nterritories = 78, Nfield-absences = 78
Model fit
Discrimination ability of models (best model and models with ∆AICc values <2 to the best) based on AUC was considered excellent to outstanding with values between 0.84 and 0.93 (Appendix 5—Supplementary material). Model predictive performances based on TSS ranged from moderate to substantial with values between 0.59 and 0.77.
Discussion
Territory selection of the wood warbler at two spatial scales was governed by similar factors. Pre-breeding territories were located in areas with intermediate vegetation height, lower vertical diversity, higher penetration rate of the tree layer, larger canopy cover between 10 and 20 m above ground and lower canopy cover above 20 m, increased inclination and higher potential direct solar radiation in March compared to control areas in the wider forested landscape (Table 3). Breeding territories differed from control areas in the immediate forest vicinity by lower vegetation height, lower vertical diversity, lower canopy cover above 20 m and increased inclination.
Vegetation height (expectation 1)
The results imply that both pre-breeding and breeding territory selection of wood warblers were sensitive to stand height. As expected, probability of pre-breeding territory occurrence showed a negative quadratic relationship with mean canopy and maximum vegetation height, indicating an optimum at intermediate heights. In addition, the negative relationship with maximum vegetation height indicates that pre-breeding territories were established in areas with smaller trees than available in the wider forested landscape. Likewise, for the breeding territories, the negative relationships with the vegetation height variables suggest that areas were selected that featured smaller trees than what was available in the immediate forest vicinity. Intermediate stand height corresponds to late pole wood (mean diameter at breast height of the hundred thickest trees per hectare dbh100 = 20–30 cm) and young timber (dbh100 = 31–40 cm) (Cioldi et al. 2010). Stands of these development stages are usually single-layered and characterized by a closed canopy (Korpel 1995; Pontailler et al. 1997; Meyer et al. 2003; Rozenbergar et al. 2007). This allows for an open stem space and inhibits the development of dense regeneration, herbs and shrubs. A relatively open under- and mid-story is conducive to the wood warbler’s courtship behavior, which includes song-flights from low branches between tree trunks (Glutz von Blotzheim and Bauer 1991). Moreover, a closed canopy allows for development of a sparse and low-growing ground vegetation cover, a key habitat feature affecting both breeding habitat selection (Pasinelli et al. 2016) and reproductive performance (Grendelmeier et al. 2015) of the wood warbler.
Vertical diversity (expectation 2)
Wood warbler territory occurrence was negatively related to vertical diversity of vegetation and canopy height. Thus, expectation 2 was supported for both scales, suggesting that stands are preferred that are characterized by low tree height diversity of the canopy trees and few gaps. These conclusions are closely connected to the findings from expectation 1 because stands of medium height are characterized by homogeneous stand height (Korpel 1995; Pontailler et al. 1997; Meyer et al. 2003; Rozenbergar et al. 2007). In general, avian diversity in forests is positively related to structural diversity (Scherzinger 1996; Müller 2005; Winter et al. 2005; Mollet et al. 2006; Goetz et al. 2007; Müller et al. 2009). Thus, while measures aiming at increasing structural diversity within forests appear to promote bird species richness, specialized species such as the wood warbler preferring homogeneously structured forests will not benefit.
Vertical stratification (expectation 3)
Unexpectedly, pre-breeding territory selection was positively related to the penetration rate of the tree layer. This indicates that pre-breeding territories were located in forest stands enabling lidar signals to pass through them more easily than through other forest stands available in the landscape. Penetration rate of the tree layer was not correlated with the lidar-derived canopy cover (Appendix 2—Supplementary material), but negatively related with the field variable number of conifers (rs = −0.51, Appendix 3—Supplementary material) gathered for addressing other questions within the wood warbler project (Pasinelli et al. 2016). This implies that male wood warblers select territories that contain fewer conifers than control areas in the forested landscape. However, the forest type variable (expectation 10) did not receive support. It is likely that the spatial resolution of the data and the classification into four groups representing coniferous to broadleaf forest types was not sufficiently accurate to test for a preference of broadleaf forest stands. Rather, lidar data collection outside the growing season appears to be suited for capturing the proportion of coniferous trees because of their higher amount of canopy reflections compared to deciduous trees.
Canopy cover (expectation 6)
According to this study, canopy cover favoring wood warbler occurrence was high 10 m above ground (pre-breeding territories) but low 20 m above ground (pre-breeding and breeding territories). Again, this finding is closely connected with the results concerning vegetation height (expectation 1) and vertical diversity (expectation 2) because pole wood and young timber are characterized by a closed canopy.
Slope (expectation 7)
In line with previous studies, wood warbler occurrence at both the pre-breeding and breeding territory scales was positively related to slope. Several reasons for a preference of slopes by breeding wood warblers have been proposed. First, slope may act as surrogate for some main characteristics of habitat suitability. For example, relatively dense middle-aged forest stands along inclined terrain might allow more direct sun radiation to reach the ground, thereby positively affecting ground vegetation cover (i.e., grass tussocks) important for wood warbler nest site choice and nest survival (Grendelmeier et al. 2015; Pasinelli et al. 2016), compared to similar forest stands in flat terrain. Second, Mallord et al. (2012) argued that nowadays wood warblers mainly settle in the upland areas of the UK, where grazing has remained the dominant form of land management, maintaining the open woodland structure preferred by wood warblers. Also in Switzerland, wood warbler occurrence has strongly declined in the lowlands (Schmid et al. 1998). However, the argument of grazing is not applicable to Switzerland because wood pasture was already discontinued after the first half of the 19th century due to changes in agricultural practice (Bürgi 1999). Third, when comparing pre-breeding territories to control areas in the wider forested landscape, another explanation for the positive relationship between wood warbler occurrence and slope could be that disturbance due to recreational activity or forest management intensity decreases with increasing inclination. However, recreational activities hardly explain the preference for inclined areas found because all study areas investigated were relatively remote. In addition, distance to trails and paths did not differ between breeding territories and field-absences (Pasinelli et al. 2016). Concerning forest management, inclined areas are often treated less intensively due to increased costs and logistic challenges of timber harvesting. Many study areas are located in areas where the last treatment, a good indicator for forest management intensity, took place 20–50 years ago (Duc et al. 2010), and some study areas are situated in forest nature reserves. However, forest management hardly explains the positive relationship between wood warbler occurrence and inclination when comparing breeding territories to what is available in the immediate forest vicinity. A final explanation for the observed relationship with slope might thus be found in the nesting ecology of the wood warbler. Nest entrances are typically oriented horizontally in this species. Nests built at inclined locations, with entrances facing away from the slope, could allow wood warblers to easily escape from nests without getting entangled in the vegetation (Piotrowska and Wesolowski 1989). Nest predation accounts for the majority of nest losses in this ground-nesting species (Grendelmeier et al. 2015).
Potential direct solar radiation in March (expectation 8)
Pre-breeding territory selection was positively related to potential direct solar radiation in March. Because solar radiation is highest on south-facing slopes and lowest on north-facing slopes, this finding may explain why wood warbler territories with southern aspect are more common than territories with northern or western aspects (Glutz von Blotzheim and Bauer 1991; Hölzinger 1999; Reinhardt and Bauer 2009). Further, solar radiation might positively influence food availability. Higher food availability on south-facing slopes than on north-facing slopes could attract wood warblers, as Jedrzejewska and Jedrzejewski (1998) and Kühn (2015) reported positive correlations between wood warbler population size and food abundance.
Conclusions
Many conservation efforts in forests aim at developing open and diversely structured forest stands to promote biodiversity. Structurally rich stands also promote bird species diversity because habitat suitability for many avian forest species is strongly determined by three-dimensional vegetation structure (e.g. Dunlavy 1935; Müller et al. 2009; Zellweger et al. 2013). Nevertheless, not all species thrive in structurally rich stands, if these species require specific developmental forest stages. The wood warbler seems to be an example of such a species, because it favors a rather uniform environment. Our study with RS variables corroborates previous field-based studies relating wood warbler habitat to homogeneously structured forests of intermediate age, featuring a closed canopy and low canopy height diversity. Forest management may locally contribute to the deterioration of suitable areas, for example when relatively closed forests are opened up due to harvesting. Likewise, forestry practices that aim either at bringing more light into woodland by selective thinning or that generate forest stands permanently covered with shrubs, bushes and trees of various sizes (continuous cover forestry) are not beneficial to the wood warbler. On the other hand, maturing of forest stands also deteriorates habitat quality for this species in the long run. Therefore, forest management at a regional scale should maintain a shifting mosaic of forest patches of different development stages by focusing on sustainable regeneration and a balanced age structure so that suitable stands of even age are always present and new suitable stands are steadily developing. Moreover, areas characterized by abiotic factors identified as relevant for wood warbler occurrence, namely slope and solar radiation, should be identified and managed appropriately. Such areas could represent “safe havens” for this species, given that abiotic factors important for wood warbler occurrence do not change over time, and should thus not be targets of forest management aiming at increasing forest biodiversity, for example by excessive thinning to favor light-demanding species.
That territory selection during both pre-breeding and breeding is governed by similar factors is a novel and important finding for the conservation of this declining passerine. Analyzing habitat needs at two spatial scales allows transferring the inferences of this study to both fine- and broad-scale forest and/or conservation management without struggling with scaling issues. The results suggests that promoting forests consisting of relatively uniform habitat, as outlined above, will serve both to attract prospecting males prior to breeding and to subsequently allow establishment of breeding territories. This simplifies management measures needed to promote suitable habitats for this species as opposed to the situation when different structures affect habitat selection at different spatial scales.
Finally, lidar-derived RS data based on first and last return data acquired during leaf-off conditions appear to be suitable to describe habitat suitability during the breeding season of forest birds, as for example the wood warbler, even at small spatial scales.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43:1223–1232
Bates D, Maechler M, Bolker B (2012) Linear mixed-effects models using S4 classes (lme4). Package version 0.999999-0. http://cran.rstudio.com/bin/windows/contrib/2.14/. Accessed Dec 2012
Bellamy PE, Hill RA, Rothery P, Hinsley SA, Fuller RJ, Broughton RK (2009) Willow Warbler Phylloscops trochilus habitat in woods with different structure and management in southern England. Bird Stud 56:338–348
Bellis LM, Pidgeon AM, Radeloff VC, St-Louis V, Navarro JL, Martella MB (2008) Modeling habitat suitability for greater rheas based on satellite image texture. Ecol Appl 18:1956–1966
Bibby CJ (1989) A survey of breeding wood warblers Phylloscopus sibilatrix in Britain, 1984–1985. Bird Stud 36:56–72
BirdLife International (2015) European red list of birds. Office for Official Publications of the European Communities, Luxembourg
Brambilla M, Falco R, Negri I (2012) A spatially explicit assessment of within-season changes in environmental suitability for farmland birds along an altitudinal gradient. Anim Conserv 15:638–647
Brandtberg T, Warner TA, Landenberger RE, McGraw JB (2003) Detection and analysis of individual leaf-off tree crowns in small footprint, high sampling density lidar data from the eastern deciduous forest in North America. Remote Sens Environ 85:290–303
Broughton RK, Hinsley SA, Bellamy PE, Hill RA, Rothery P (2006) Marsh Tit Poecile palustris territories in a British broad-leaved wood. Ibis 148:744–752
Broughton RK, Ross AH, Freeman SN, Bellamy PE, Hinsley SA (2012) Describing habitat occupation by woodland birds with territory mapping and remotely sensed data: an example using the marsh tit (Poecile palustris). Condor 114:812–822
Bunnell FL, Huggard DJ (1999) Biodiversity across spatial and temporal scales: problems and opportunities. For Ecol Manag 115:113–126
Bürgi M (1999) A case study of forest change in the Swiss lowlands. Landscape Ecol 14:567–575
Burnham K, Anderson D (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, Berlin
Chetkiewicz C-LB, Boyce MS (2009) Use of resource selection functions to identify conservation corridors. J Appl Ecol 46:1036–1047
Cioldi F, Baltensweiler A, Brändli U-B, Duc P, Ginzler C, Herold Bonardi A, Thürig E, Ulmer U (2010) Waldressourcen. In: Brändli U-B (ed) Schweizerisches Landesforstinventar: Ergebnisse der dritten Erhebung 2004–2006. Eidgenössische Forschungsanstalt für Wald, Schnee und Landschaft, WSL, Birmensdorf and Bundesamt für Umwelt, BAFU, Bern
Cohen J (1960) A coefficient of agreement for nominal scales. Educ Psychol Meas 20:37–46
Coops NC, Duffe J, Koot C (2010) Assessing the utility of lidar remote sensing technology to identify mule deer winter habitat. Can J Remote Sens 36:81–88
Duc P, Brändli U-B, Bornardi AH, Rösler E, Thürig E, Ulmer U, Frutig F, Rosset C, Kaufmann E (2010) Holzproduktion. In: Brändli U-B (ed) Schweizerisches Landesforstinventar: Ergebnisse der dritten Erhebung 2004-2006. Eidgenössische Forschungsanstalt für Wald, Schnee und Landschaft, WSL, Birmensdorf and Bundesamt für Umwelt, BAFU, Bern
Duncan C, Kretz D, Wegmann M, Rabeil T, Pettorelli N (2014) Oil in the Sahara: mapping anthropogenic threats to Saharan biodiversity from space. Philos Trans R Soc B 369:20130191
Dunlavy JC (1935) Studies on the phyto-vertical distribution of birds. Auk 52:425–431
EIONET (2015) Population status and trends at the EU and Member State levels. http://bd.eionet.europa.eu/article12/summary?period=1&subject=A314. Accessed Nov 2015
Everitt B (2002) The Cambridge dictionary of statistics, 2nd edn. Cambridge University Press, Cambridge
Farrell SL, Collier BA, Skow KL, Long AM, Campomizzi AJ, Morrison ML, Hays KB, Wilkins RN (2013) Using LiDAR-derived vegetation metrics for high-resolution, species distribution models for conservation planning. Ecosphere 4:1–18
Fielding AH, Bell JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24:38–49
Finck P (1990) Seasonal variation of territory size with the little owl (Athene noctua). Oecologia 83:68–75
Freeman E (2012) Presence-Absence Model Evaluation (PresenceAbsence). Package version 1.1.9. http://cran.rstudio.com/bin/windows/contrib/2.14/. Accessed Dec 2012
Gerber M (2011) Territory choice of the Wood Warbler Phylloscopus sibilatrix in Switzerland in relation to habitat structure and rodent density. Master Thesis, University of Zurich, Zurich
Glutz von Blotzheim UN, Bauer KM (1991) Handbuch der Vögel Mitteleuropas. AULA-Verlag GmbH, Wiesbaden
Goetz SJ, Steinberg D, Betts MG, Holmes RT, Doran PJ, Dubayah R, Hofton M (2010) Lidar remote sensing variables predict breeding habitat of a Neotropical migrant bird. Ecology 91:1569–1576
Goetz S, Steinberg D, Dubayah R, Blair B (2007) Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens Environ 108:254–263
Graf RF, Mathys L, Bollmann K (2009) Habitat assessment for forest dwelling species using LiDAR remote sensing: Capercaillie in the Alps. For Ecol Manag 257:160–167
Grendelmeier A, Arlettaz R, Gerber M, Pasinelli G (2015) Reproductive performance of a declining forest passerine in relation to environmental and social factors: implications for species conservation. PLoS ONE 10:e0130954
Grubbs FE (1969) Procedures for detecting outlying observations in samples. Technometrics 11:1–21
Hawbaker TJ, Gobakken T, Lesak A, Trømborg Contrucci K, Radeloff V (2010) Light detection and ranging-based measures of mixed hardwood forest structure. For Sci 56(3):313–326
Hobson KA, Van Wilgenburg SL, Wesolowski T, Maziarz M, Bijlsma RG, Grendelmeier A, Mallord JW (2014) A multi-isotope (δ2H, δ13C, δ15 N) approach to establishing migratory connectivity in Palearctic-Afrotropical migrants: an example using wood warblers Phylloscopus sibilatrix. Acta Ornithol 49:57–69
Hölzinger J (1999) Die Vögel Baden-Württembergs Singvögel 1. Eugen Ulmer GmbH & Co, Stuttgart
Hosmer D, Lemeshow S (2000) Applied logistic regression, 2nd edn. Wiley, New York
Hurvich CM, Tsai C-L (1989) Regression and time series model selection in small samples. Biometrika 76(2):297–307
Hutto RL (1985) Habitat selection by nonbreeding, migratory land birds. In: Cody ML (ed) Habitat selection in birds. Academic Press, Orlando, pp 455–476
Isenburg M (2012) LAStools: award winning software for rapid LIDAR processing. http://www.cs.unc.edu/~isenburg/lastools/. Accessed Nov 2012
Jedrzejewska B, Jedrzejewski W (1998) Predation in vertebrate communities: the Bialowiezka primeval forest as a case study. Ecol Stud 135:1–450
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71
Jung K, Kaiser S, Böhm S, Nieschulze J, Kalko EKV (2012) Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J Appl Ecol 49:523–531
Korpel S (1995) Die Urwälder der Westkarpaten. Fischer, Stuttgart
Kühn B (2015) The role of caterpillar abundance and phenology for breeding success, territory choice and population size of the declining wood warbler (Phylloscopus sibilatrix). Master Thesis, University of Zurich, Zurich
Landis JR, Koch GG (1977) The measurement of observer agreement for categorical data. Biometrics 33:159–174
Lefsky MA, Cohen WB, Parker GG, Harding DJ (2002) Lidar remote sensing for ecosystem studies. BioSience 52:19–30
Luoto M, Kuusaari M, Toivonen T (2002) Modelling butterfly distribution based on remote sensing data. J Biogeogr 29:1027–1037
Mallord JW, Charman EC, Cristinacce A, Orsman CJ (2012) Habitat associations of Wood Warblers Phylloscopus sibilatrix breeding in Welsh oakwoods. Bird Stud 59:403–415
Mazerolle MJ (2006) Improving data analysis in herpetology: using Akaike’s Information Criterion (AIC) to assess the strength of biological hypotheses. Amphibia-Reptilia 27:169–180
Mazerolle MJ (2012) Model selection and multimodel inference based on (Q) AIC (c) (AICcmodavg), Version 1.28. http://cran.rstudio.com/bin/windows/contrib/2.14/. Accessed Dec 2012
Merrill T, Mattson DJ, Wright RG, Quigley HB (1999) Defining landscapes suitable for restoration of grizzly bears Ursus arctos in Idaho. Biol Conserv 87:231–248
MeteoSchweiz (2014) Normwerte 1981-2010: Niederschlagssumme. Bundesamt für Meteorologie und Klimatologie MeteoSchweiz, Zürich
Meyer P, Tabaku V, von Lüpke B (2003) Die Struktur albanischer Rotbuchen-Urwälder: Ableitungen für eine naturnahe Buchenwirtschaft. Forstw Cbl 122:47–58
Möller AP (1990) Changes in the size of avian breeding territories in relation to the nesting cycle. Anim Behav 40:1070–1079
Mollet P, Birrer S, Naef-Daenzer B, Naef-Daenzer L, Spaar R, Zbinden N (2006) Situation der Vogelwelt im Schweizer Wald. Schweizerische Vogelwarte, Sempach
Müller J (2005) Waldstrukturen als Steuergrösse für Artengemeinschaften in kollinen bis submontanen Buchenwäldern. Dissertation, Technische Universität München, Munchen
Müller J, Bae S, Röder J, Chao A, Didham RK (2014) Airborne LiDAR reveals context dependence in the effects of canopy architecture on arthropod diversity. For Ecol Manag 312:129–137
Müller J, Brandl R (2009) Assessing biodiversity by remote sensing in mountainous terrain: the potential of LiDAR to predict forest beetle assemblages. J Appl Ecol 46:897–905
Müller J, Moning C, Bässler C, Heurich M, Brandl R (2009) Using airborne laser scanning to model potential abundance and assemblages of forest passerines. Basic Appl Ecol 10:671–681
Næsset E (2005) Assessing sensor effects and effects of leaf-off and leaf-on canopy conditions on biophysical stand properties derived from small-footprint airborne laser data. Remote Sens Environ 98:356–370
Nelson R, Keller C, Ratnaswamy M (2005) Locating and estimating the extent of Delmarva fox squirrel habitat using an airborne LiDAR profiler. Remote Sens Environ 96:292–301
Ørka HO, Næsset E, Bollandsås OM (2010) Effects of different sensors and leaf-on and leaf-off canopy conditions on echo distributions and individual tree properties derived from airborne laser scanning. Remote Sens Environ 114:1445–1461
Palminteri S, Powell GVN, Asner GP, Peres CA (2012) LiDAR measurements of canopy structure predict spatial distribution of a tropical mature forest primate. Remote Sens Environ 127:98–105
Parent JR, Volin JC (2014) Assessing the potential for leaf-off lidar data to model canopy closure in temperate deciduous forests. ISPRS J Photogramm Remote Sens 95:134–145
Park C (2007) A dictionary of environment and conservation, 1st edn. Oxford University Press, New York
Parker GG, Brown MJ (2000) Forest canopy stratification—is it useful? Am Nat 4:473–484
Pasinelli G, Grendelmeier A, Gerber M, Arlettaz, R (2016) Rodent-avoidance, topography and forest structure shape territory selection of a forest bird. BMC Ecol (in press)
Pasinelli G, Hegelbach J, Reyer H-U (2001) Spacing behavior of the middle spotted woodpecker in central Europe. J Wildl Manag 65:432–441
Pavlovic A (2009) Raumnutzung von Waldstrukturen durch Phylloscopus-Arten anhand von Scannerdaten. Diplomarbeit, Fachhochschule Weihenstephan, Freising
Pettorelli N, Kamran S, Turner W (2014) Satellite remote sensing, biodiversity research and conservation of the future. Philos Trans R Soc B 369:20130190
Piotrowska M, Wesolowski T (1989) The breeding ecology and behaviour of the chiffchaff Phylloscopus collybita in primaeval and managed stands of Bialowieza Forest (Poland). Acta Ornithol 25:25–76
Pontailler J-Y, Faille A, Lemée G (1997) Storms drive successional dynamics in natural forests: a case study in Fontainebleau forest (France). For Ecol Manag 98:1–15
Quelle M, Tiedemann G (1972) Strukturanalyse von Waldlaubsängerrevieren im Raum Bielefeld. Abh Landesmus Nat Münst Westfal 34:95–102
Reidy JL, Thompson FR III, Amundson C, O’Donnell L (2015) Landscape and local effects on occupancy and densities of an endangered wood-warbler in an urbanizing landscape. Landscape Ecol. doi:10.1007/s10980-015-0250-0
Reinhardt A, Bauer H-G (2009) Analyse des starken Bestandesrückgangs beim Waldlaubsänger Phylloscopus sibilatrix im Bodenseegebiet. Vogelwarte 47:23–39
Rozenbergar D, Mikac S, Anic I, Diaci J (2007) Gap regeneration patterns in relationship to light heterogeneity in two old-growth beech-fir forest reserves in South East Europe. Forestry 80:431–443
Scherzinger W (1996) Naturschutz im Wald—Qualitätsziele einer dynamischen Waldentwicklung. Ulmer, Stuttgart
Schifferli A, Géroudet P, Winkler R, Jacquat B (1980) Verbreitungsatlas der Brutvögel der Schweiz: kartographische Darstellung des Brutvorkommens aller einheimischen Vogelarten in den Jahren 1972 bis 1976. Schweizerische Vogelwarte, Sempach
Schmid H, Luder R, Naef-Daenzer B, Graf R, Zbinden N (1998) Schweizer Brutvogelatlas: Verbreitung der Brutvögel in der Schweiz und im Fürstentum Liechtenstein 1993–1996. Schweizerische Vogelwarte, Sempach
Shaw DC, Freeman EA, Flick C (2002) The vertical occurrence of small birds in an old-growth douglas-fir-western hemlock forest Stand. Northwest Sci 76:322–334
Smart LS, Swenson JJ, Christensen NL, Sexton JO (2012) Three-dimensional characterization of pine forest type and red-cockaded woodpecker habitat by small-footprint, discrete-return lidar. For Ecol Manag 281:100–110
Sperduto MB, Congalton RG (1996) Predicting rare orchid (small whorled pogonia) habitat using GIS. Photogram Eng Remote Sens 62:1269–1279
St-Louis V, Pidgeon AM, Kuemmerle T, Sonnenschein R, Radeloff VC, Clayton MK, Locke BA, Bash D, Hostert P (2014) Modelling avian biodiversity using raw, unclassified satellite imagery. Philos Trans R Soc B 369:20130197
Swiss Federal Statistical Office (2010) Waldmischungsgrad. http://www.bfs.admin.ch/bfs/portal/de/index/dienstleistungen/geostat/datenbeschreibung/waldmischungsgrad.html. Accessed Dec 2012
swisstopo (2012) DOM. http://www.swisstopo.admin.ch/internet/swisstopo/de/home/products/height/dom_dtm-av.html. Accessed Dec 2012
swisstopo (2013) swissALTI3D. http://www.swisstopo.admin.ch/internet/swisstopo/de/home/products/height/swissALTI3D.html. Accessed Apr 2013
swisstopo (2014) swiss TLM3D. http://www.swisstopo.admin.ch/internet/swisstopo/de/home/products/landscape/swissTLM3D.html. Accessed Feb 2014
swisstopo (2015) DHM25. http://www.swisstopo.admin.ch/internet/swisstopo/en/home/products/height/dhm25.html. Accessed Aug 2015
Trainor AM, Walters JR, Morris WF, Sexton J, Moody A (2013) Empirical estimation of dispersal resistance surfaces: a case study with red-cockaded woodpeckers. Landscape Ecol 28:767–775
Vierling KT, Bässler C, Brandl R, Vierling LA, Weiss I, Müller J (2011) Spinning a laser web: predicting spider distributions using LiDAR. Ecol Appl 21:577–588
Vierling KT, Swift CE, Hudak AT, Vogeler JC, Vierling LA (2014) How much does the time lag between wildlife field-data collection and LiDAR-data acquisition matter for studies of animal distributions? A case study using bird communities. Remote Sens Lett 5:185–193
Vierling KT, Vierling LA, Gould W, Martinuzzi S, Clawges RM (2008) Lidar: shedding new light on habitat characterization and modeling. Front Ecol Environ 6:90–98
Vierling LA, Vierling KT, Adam P, Hudak AT (2013) Using satellite and airborne LiDAR to model woodpecker habitat occupancy at the landscape scale. PLoS ONE 8:1–13
Vogeler JC, Hudak AT, Vierling LA, Vierling KT (2013) Lidar-derived canopy architecture predicts Brown Creeper occupancy of two western coniferous forests. Condor 115:614–622
Wasser L, Day R, Chasmer L, Taylor A (2013) Influence of vegetation structure on lidar-derived canopy height and fractional cover in forested riparian buffers during leaf-off and leaf-on conditions. PLoS ONE 8:e54776
Wesolowski T, Maziarz M (2009) Changes in breeding phenology and performance of Wood Warblers Phylloscopus sibilatrix in a primeval forest: a thirty-year perspective. Acta Ornithol 44:69–80
Whittingham MJ, Swetnam RD, Wilson JD, Chamberlan DE, Freckleton RP (2005) Habitat selection by yellowhammers Emberiza citrinella on lowland farmland at two spatial scales: implications for conservation management. J Appl Ecol 42:270–280
Wikar D, Ciach M, Bylicka M, Bylicka M (2008) Changes in habitat use by the common buzzard (Buteo buteo L.) during non-breeding season in relation to winter conditions. Pol J Ecol 56:119–125
Wiktander U, Olsson O, Nilsson SG (2001) Seasonal variation in home-range size, and habitat area requirement of the lesser spotted woodpecker (Dendrocopos minor) in southern Sweden. Biol Conserv 100:387–395
Winter S, Flade M, Schumacher H, Kerstan E, Möller G (2005) The importance of near-natural stand structures for the biocoenosis of lowland beech forests. For Snow Landsc Res 79:127–144
Wood EM, Pidgeon AM, Radeloff VC, Keuler NS (2013) Image texture predicts avian density and species richness. PLoS ONE 8:e63211
Zellweger F, Braunisch V, Baltensweiler A, Bollmann K (2013) Remotely sensed forest structural complexity predicts multi species occurrence at the landscape scale. For Ecol Manag 307:303–312
Zimmermann NE, Kienast F (1999) Predictive mapping of alpine grasslands in Switzerland: species versus community approach. J Veg Sci 10:469–482
Zweig MH, Campell G (1993) Receiver-operating characteristic (ROC) plots: a fundamental evaluation tool in clinical medicine. Clin Chem 39:561–577
Acknowledgments
Many thanks to Alexander Grendelmeier and Michael Gerber for collecting data on wood warbler nests and territory locations. We thank Rafael Wüest for his support with the statistical analyses and Peter Rotach for a valuable discussion. We are grateful for helpful comments to previous versions of the manuscript provided by Kurt Bollmann, Jeff Hepinstall-Cymerman and three anonymous reviewers. For financial support, we thank the Swiss National Science Foundation (grant no. 31003A_143879 to G. P. and Raphaël Arlettaz), Hilfsfonds für die Schweizerische Vogelwarte Sempach, PD-Stiftung der Universität Zürich, Styner-Stiftung, Stotzer-Kästli-Stiftung and Lotteriefonds des Kantons Solothurn.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Huber, N., Kienast, F., Ginzler, C. et al. Using remote-sensing data to assess habitat selection of a declining passerine at two spatial scales. Landscape Ecol 31, 1919–1937 (2016). https://doi.org/10.1007/s10980-016-0370-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0370-1