
Abstract
Context
The conservation value of residential landscapes is becoming increasingly apparent in our urbanizing world. The ecological characteristics of residential areas are largely determined by the decisions of many individual “managers.” In these complex socio-ecological systems, it is important to understand the factors that motivate human decision-making.
Objectives
Our first objective was to quantify wildlife resources and management activities in residential landscapes and compare vegetation in front and back yards. Our second objective was to test three hypotheses linked with variation in yards: socioeconomic characteristics, neighborhood design factors, and perceptions of neighborhood birds.
Methods
We conducted surveys of over 900 residents in 25 Chicago-area neighborhoods to examine the wildlife resources contained in front and back yards and the social factors associated with variation in yards. We used a multi-scalar approach to examine among-yard and among-neighborhood variation in residential landscapes.
Results
Results indicate that back yards contain more wildlife resources than front yards, including greater vegetation complexity, more plants with fruit/berries, and more plants intended to attract birds. Furthermore, different hypotheses explain variation in front and back yards. Perceptions of birds were most important in explaining variation in back yard vegetation and wildlife-friendly resources per parcel, while neighbors’ yards and socioeconomic characteristics best explained front yard vegetation.
Conclusions
This study demonstrates the importance of back yards as an unexplored and underestimated resource for biodiversity. In addition, the results provide insight into the complex factors linked with yard decisions, notably that residents’ connections with neighborhood birds appear to translate to on-the-ground actions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Residential landscapes exemplify social-ecological systems where environmental decisions are made by many individual “managers” (Gaston et al. 2013). Residents affect the biophysical template of urban landscapes via their yard design and management decisions, such as plant choices, maintenance activities, and incorporating wildlife-friendly features. Eighty percent of U.S. households have access to a yard or garden (U.S. EPA 2012), and the collective decisions people make in their yards can scale up to influence ecosystem service provision and biodiversity at much broader spatial scales (Kinzig et al. 2005; Belaire et al. 2014). As Wesley Schultz points out, “Drivers of phenomena such as climate change, loss of species’ habitats, and ocean acidification rarely are the result of malicious intent, but rather the consequence of the lifestyles of billions of humans” (Schultz 2011). Indeed, people seem to have limited understanding of the role their yards play in urban ecosystems (Clayton 2007; Dahmus and Nelson 2013). In these types of complex socio-ecological systems, it is important to understand the factors that motivate environmental decisions and ultimately affect ecological outcomes.
Although conventional wisdom regards urban residential landscapes as a homogeneous land cover type, research has revealed considerable heterogeneity between yards, neighborhoods, and cities (Cameron et al. 2012; Harris et al. 2012; Polsky et al. 2014; Minor et al. 2015). We do not fully understand the mechanisms driving residents’ decisions about yard design and management. Recent reviews (Chowdhury et al. 2011; Cook et al. 2012) suggest that yard behaviors are influenced by a complex suite of factors, including aesthetic tastes, socioeconomic characteristics, housing age, formal institutions (e.g., homeowners’ associations), and informal institutions (e.g., neighborhood norms). People also make different decisions or have different preferences regarding front versus back yards (Larsen and Harlan 2006; Daniels and Kirkpatrick 2006b), which further complicates matters. Ultimately, residential yard outcomes represent the influence of multi-scalar factors. Thus, different scales of analysis (e.g., household-scale versus neighborhood-scale) might reveal different patterns and influences operating simultaneously (Harris et al. 2012).
Socioeconomic characteristics appear to be important in yard design and management. For example, income has been positively associated with vegetation richness in residential neighborhoods (Martin et al. 2004; Minor et al. 2015), whereby people with greater financial resources choose landscapes with higher plant diversity (Hope et al. 2003). Perception of social status and group identity also motivate yard-related decisions (“ecology of prestige,” Grove et al. 2006). Some socioeconomic factors may be important for wildlife-friendly features in yards, such as birdfeeders. For example, householder age appears to be an important predictor for bird-feeding, while the role of income is less clear (Lepczyk et al. 2004; Davies et al. 2012; Clucas et al. 2014).
Neighborhood design can also play a role in yards. For example, recently-built neighborhoods in Baltimore had less vegetation cover than neighborhoods established 40-50 years ago (Grove et al. 2006). In the Chicago area, however, newer neighborhoods often retain some existing habitat and thus may be more vegetated than older neighborhoods (Loss et al. 2009). In addition, neighborhood housing density affects yard size (Tratalos et al. 2007), which may affect yard composition. Although bigger yards may provide more wildlife resources (Gaston et al. 2007; van Heezik et al. 2013), some research shows that density of wildlife-friendly features like birdfeeders increases with housing density (Fuller et al. 2012). At the scale of individual yards, residents appear to mimic neighbors in their yard and easement designs (Zmyslony and Gagnon 2000; Hunter and Brown 2012) and inspire one another to incorporate wildlife-friendly features into their yards (Goddard et al. 2013). Social norms at the neighborhood scale may help explain spatial similarity between nearby yards (Grove et al. 2006; Nassauer et al. 2009).
Perceptions of and attitudes toward local wildlife may also explain variation in residential yards. Several researchers have suggested that neighborhood bird communities influence garden design and management. For example, Head and Muir (2006) noted that people with both native and exotic plants in their garden reported enjoying observations of birds so much that it inspired them to install more native vegetation. Likewise, Goddard et al. (2013) found that watching and protecting local wildlife were important motivators for wildlife gardening activities. Another study found that positive conservation attitudes and “noticing birds at least weekly” helped explain bird feed expenditures (Clucas et al. 2014). More broadly, pro-environmental attitudes have been linked with vegetation structure in yards (van Heezik et al. 2013), but the relationship between environmental concern and yard decisions is inconsistent (Larson et al. 2010; Cook et al. 2012).
Despite the importance of yards for wildlife and ecosystem services, we still lack a clear picture of the social drivers of yard design and management decisions. Here we used a social survey instrument to investigate the social factors explaining variation in yards for over 900 residents in the Chicago, IL (USA) metropolitan area. First, we quantified wildlife resources and management activities in residential landscapes, and we compared vegetation in front and back yards. Second, we used a multi-scalar approach to test several hypotheses that might explain among-yard and among-neighborhood variation in yards. Specifically, we examined the following hypotheses: (H1) Yard composition is driven by residents’ perceptions of birds in their neighborhoods; (H2) Yard composition is driven by socioeconomic characteristics; and (H3) Yard composition is driven by neighborhood design characteristics.
Methods
Study sites
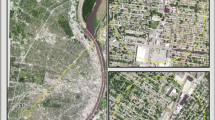
Cook County, IL is home to the city of Chicago, over 5 million residents, and almost 30,000 ha of forest preserves. We delineated 25 transects in residential neighborhoods across Cook County as study sites (Fig. 1, Belaire et al. 2014). Each transect began at the edge of a forest preserve and extended 1 km into the adjacent residential neighborhood. Transects were located at least 500 m apart to minimize spatial dependencies.

Example of a residential neighborhood in study area. The aerial view (top) shows one neighborhood block, approximately 200 m in length, with front yards visible from the street and back yards located on the opposite side of the home (out of view from the street). The street view (bottom) depicts a series of homes with front yards and sidewalks. (All imagery from Google Earth)
Data collection
We had two major goals with data collection. First, we wanted to understand the types of wildlife resources in residents’ yards and compare vegetation in front and back yards (Table 1). Second, we wanted to examine the relative importance of several hypotheses that are likely to explain variation in yards. To gather information about residents’ yards and variables related to the three hypotheses, we developed a social survey (following the guidelines of Dillman et al. 1978; full survey in Appendix S1).
We delivered surveys to all single-family residences within 50 m of the transect (n = 1751 residences) and requested that the survey be completed by one adult with some responsibility for managing the yard. Surveys were distributed with the “drop-off/pick-up” method (Steele et al. 2001; Allred and Ross-Davis 2011) during July–September 2012, and each survey included a $1 token financial incentive. After surveys were collected, we checked for non-response bias by comparing respondents to non-respondents in two ways: grass and canopy cover at the parcel scale (from 0.6 m QuickBird imagery) and socioeconomic characteristics (comparing survey responses to American Community Survey block group data). These two methods allowed us to determine if the results are influenced by lack of response from some segment of the population (Dillman et al. 1978).
The first portion of the survey focused on residents’ yard composition and contained primarily close-ended questions. Residents indicated the vegetation types in their front and back yards and presence of potential wildlife resources (e.g., bird feeder, bird house). We also asked about yard activities, such as insecticide application and whether the home had outdoor pets. We used the survey responses to characterize vegetation types, wildlife-friendly features, and management activities in respondents’ front and back yards. We calculated the percent of respondents on each transect whose yard contained each yard element or management activity. Lastly, we developed three indices for both yard- and transect-scale analyses: (1) wildlife resources per parcel, (2) front yard vegetation, and (3) back yard vegetation (Table 1). For transect-scale analyses, we aggregated each response variable by calculating the average for all respondents on each transect.
The survey also included two sets of questions to investigate residents’ perceptions of neighborhood birds (H1). First, residents were asked to estimate the number of bird species on their block (open-ended). When residents estimated a range of species richness (e.g., 5–10), we retained their lowest estimate. When respondents estimated greater than 50 species, we assumed they were estimating abundance instead of richness and excluded their response from analysis. Second, residents were asked about ways in which they value or are annoyed by birds in their neighborhood. This portion of the survey was adapted from a tested set of questions regarding benefits and annoyances of trees (Schroeder and Ruffolo 1996). The survey included the statement “I value birds in my neighborhood because…”, which was followed by a list of 11 positive aspects of birds (e.g., “They have pleasant songs”). Residents were asked to indicate their level of agreement with each statement on a 5-point Likert scale ranging from “strongly disagree” to “strongly agree.” Then the focus shifted to negative aspects of birds: “I find birds in my neighborhood to be annoying or problematic because…” This statement was followed by 10 negative items (e.g., “Their droppings make a mess on my outdoor furniture, car, etc.”) and the same 5-point Likert scale. We assigned a value of 1 to “strongly disagree” and 5 to “strongly agree” for all positive statements (and vice versa for negative statements) and calculated the average for all statements. This resulted in a “bird value score” for each respondent. For transect-scale analyses, we calculated the average bird value score and average estimated bird species richness for all respondents on a transect (Table 2).
At the end of the survey, we included questions about the respondent’s socioeconomic characteristics (H2), including age, income, and education level. For transect-scale analyses, we averaged survey responses for education and age, and we used American Community Survey data from 2005 to 2009 at the block group scale for median household income (because many respondents did not provide income data) (Table 2).
We characterized neighborhood design characteristics (H3) in several ways (Table 2). For yard-scale analyses, we summarized yard characteristics of all respondents within 200 m of each parcel (i.e., the average number of vegetation types and wildlife resources in all front and back yards within 200 m) using the “Generate near table” tool in ArcGIS 10.1 (ESRI, Redlands, CA, USA). The respondent’s own yard was excluded from this summary. For transect-scale analyses, we calculated average age of housing (from American Community Survey 2005–2009 data at the block group scale) and number of parcels within 50 m of the transect. Number of parcels served as a proxy for housing density and was inversely correlated with average garden size (Pearson’s r = -0.86, p < 0.0001).
Variation partitioning
To examine the relative importance of the three hypotheses for yard variation (perception of birds, socioeconomic characteristics, and neighborhood design), we used a variation partitioning approach. Variation partitioning identifies the degree to which multiple hypotheses are redundant and/or have independent explanatory power.
This approach allowed us to decompose yard variation into eight fractions: the variation explained by each hypothesis independently (3 partitions), the variation explained by two hypotheses simultaneously (3 partitions), the variation explained by all three hypotheses jointly (1 partition), and the variation that cannot be explained by any of the hypotheses (1 partition). The process involves conducting a series of multiple regressions and computing fractions of variations from adjusted R2 values (Legendre and Legendre 2012, Sect. 10.5). We developed linear regression models for each response variable (number of wildlife resources, front yard vegetation, and back yard vegetation, Table 1) with the explanatory variables representing our three hypotheses (Table 2), at both yard-scale and transect-scale. We log-transformed the average number of wildlife resources for transect-scale analysis to meet assumptions of linear regression. For the yard-scale analyses, we excluded responses where socioeconomic information was not provided by respondents. Variation partitioning analyses were conducted in the “vegan” package in R (Oksanen et al. 2012). Tests of statistical significance for the 3 independent fractions were also conducted in the “vegan” package, by permuting the residuals of reduced models (Legendre and Legendre 2012). We used permutation tests of linear models (in the “lmPerm” package in R, Wheeler 2010) to test significance of the remaining testable fractions.
Results
Sample characteristics
We received responses from 924 residents (52.7 % response rate). Twelve surveys with missing addresses or incomplete responses were excluded from analysis (n = 912). Our study sites were primarily in mid- to high-income neighborhoods; household income ranged from around $45,000 to $191,000 (mean for all study sites = $106,720, American Community Survey data 2005–2009). This sample characteristic was an artifact of selecting sites near forest preserves for a related study (Belaire et al. 2014) and was not an intentional design. Thus, our sample population cannot be considered representative of Chicago-area residents. Respondents from all age groups were represented (normally distributed around the mean age of 55, range = 18–100), and most respondents were college-educated (69.4 % of respondents). Checks for non-response bias indicated that residents who did not respond to the survey did not differ significantly from those who did respond with respect to yard vegetation cover (grass cover and canopy cover). Similarly, there were no significant differences between respondents’ socioeconomic characteristics and those of the Census block group. We verified survey responses about yards by assessing two vegetation categories (shrubs and deciduous trees) in the front yards of 50 respondents, and found 90 % agreement between survey responses and our own assessment. This supports our use of resident-reported data for yard vegetation and wildlife-friendly features.
Yard composition, features, and management activities
Yards in our study area contained diverse wildlife resources (Table 3). A third (33.4 %) of respondents reported having vegetation native to the Midwest in their yards, and 25.2 % reported having vegetation intended to attract birds. Other vegetation types that support bird diversity, especially plants with fruit or berries and evergreen trees, were present in the majority of yards (55.0 and 76.6 %, respectively). We found substantial variation between transects in percent of respondents with a particular yard element or activity. For example, 5.1 % of all respondents indicated having a cat that spent time outdoors, but this ranged from 0.0 to 16.7 % between transects. Likewise, birdfeeders were present in 35.4 % of respondents’ yards, although this ranged from 22.9 to 50.0 % across transects (Table 3). Among respondents with birdfeeders, 35.6 % said they provide food every day, 29.8 % provide food several days per week, 17.5 % provide food several days per month, and 17.2 % provide food less than once per month in their feeder.
Front and back yards differed in vegetation composition and structure. Respondents reported that their back yards, on average, contained 4.43 of the 8 vegetation types on the survey (SD = 1.96), while front yards contained 4.23 types (SD = 1.58), a small but significant difference (paired-samples t test, t(911) = −3.42, p = 0.0006). We examined this result more closely to find whether any vegetation types were more common in back yards than front yards. Generally, if a resident had a specific plant type (e.g., shrubs or bushes) on her lot, it was likely to be in both her front and back yard (Fig. 2). For example, 78.2 % of respondents had shrubs or bushes in both front and back yards. But two plant types were more likely to be in the back yard only—plants with fruit or berries and vegetation planted to attract birds. Of the respondents who had plants with fruit/berries (n = 508), over half (58.3 %) reported them in the back yard only.

Distribution of vegetation types between front and back yards. In general, if a resident had a particular vegetation type on her lot, it was likely to be in both front and back yards (e.g., the majority of residents have shrubs or bushes in both yards). However, back yards alone are the most likely location for plants with fruit/berries and vegetation planted with the goal of attracting birds
Lastly, we compared yards of residents with outdoor cats to those of residents without outdoor cats. A greater proportion of residents with outdoor cats (n = 47) have yard features intended to attract birds than residents without outdoor cats (n = 865) (Fig. 3).

A greater proportion of residents with outdoor cats (n = 47) have yard features intended to attract birds than residents without outdoor cats (n = 865). The comparison for birdhouses is statistically significant (exact binomial test, 2-tailed p = 0.04)
Role of perceptions, socioeconomics, and neighborhood design in yard composition
We used variation partitioning to test three hypotheses about yard composition at two different scales. For yard-scale analyses, our dataset included only respondents who provided socioeconomic information (n = 479). The transect-scale analyses included all respondents along 25 transects. For all yard indices at both scales of analysis, there was some redundancy among our three hypotheses (Fig. 4) but each had some significant independent effects.

Results of variation partitioning analyses at the transect scale (top) and at the yard scale (bottom) for a number of wildlife resources, b front yard vegetation, and c back yard vegetation. The three circles in each figure represent the three hypotheses related to variation in yard composition. The area of each circle is approximately proportional to the variation accounted for by that component (Micallef and Rodgers 2014, eulerAPE drawing tool); however, note that circle sizes are not sized proportionately for making comparisons between the two spatial scales. Asterisks indicate statistical significance (*p < 0.10; **p < 0.05; ***p < 0.01)
At the transect scale, the three hypotheses were fairly redundant in their ability to explain yard composition (Fig. 4). Residents’ perceptions of neighborhood birds (H1) best explained variation in wildlife resources in yards. This set of explanatory variables – which included residents’ estimates of bird diversity and their bird value scores – explained a total of 59.18 % (p < 0.001) of the variation in number of wildlife resources per parcel (averaged across the transect). The portion of variation in wildlife resources explained by perceptions of birds (H1) independently (when the other two hypotheses were “partialled out”) was marginally significant considering the small sample size of 25 transects (9.64 %, p = 0.08) (Fig. 4). Perceptions of birds also explained the greatest amount of variation in back yard vegetation (53.70 % in total, p < 0.001; 9.07 % independently, p = 0.06), but socioeconomic characteristics (H2) were most important for front yard vegetation, explaining 60.37 % (p < 0.001) of the total variation and 14.85 % independently (p = 0.02) (Fig. 4).
At the yard scale, the three hypotheses were more complementary, with each having independent explanatory power. However, the yard scale models explained less of the total variation in yard composition (Fig. 4). As at the transect scale, perceptions of neighborhood birds (H1) were most important for both wildlife resources per parcel (17.98 % in total, p < 0.001; 13.67 % independently, p = 0.005) and back yard vegetation (15.97 % in total, p < 0.001; 11.37 % independently, p = 0.005) (Fig. 4). But neighborhood design characteristics (H3)—that is, the neighbors’ front yards within 200 m—were more important in explaining front yard vegetation (10.55 % in total, p < 0.001; 5.80 % independently, p = 0.005) (Fig. 4).
Discussion
We examined vegetation and wildlife resources in over 900 yards in the Chicago area. Availability of ecological resources varied substantially between neighborhoods and between front and back yards. Furthermore, different factors explained variation in front and back yards. Perceptions of birds were most important for back yard vegetation and wildlife-friendly resources, while neighbors’ yards and socioeconomic characteristics best explained front yard vegetation.
Resource heterogeneity in residential landscapes
Residential landscapes around Chicago provide a surprising number of resources for wildlife in the “matrix” between forest preserves and other remnant habitat. However, there were large differences in adoption rates of wildlife-friendly features and vegetation types across transects. Studies of wildlife resources in yards across the U.S. and Europe have also noted substantial variation in adoption rates within a single city (Gaston et al. 2007) and between cities (Loram et al. 2008; Lepczyk et al. 2012). This heterogeneity has important implications for landscape connectivity. Yards and neighborhoods with more wildlife resources may offer relatively little resistance to moving organisms, which can reduce the effective isolation of existing habitat patches (Rudd et al. 2002; Vergnes et al. 2012). The importance of yards for urban biodiversity is becoming increasingly apparent (Daniels and Kirkpatrick 2006a; Lerman and Warren 2011; Goddard et al. 2013; Belaire et al. 2014), suggesting that residential matrix management is a viable conservation strategy in urban areas (Cooper et al. 2007).
Back yards in particular may represent an underestimated resource for urban biodiversity. Most research in urban areas has excluded back yards, because they aren’t visible from the street or easy to access. We found that back yards contained more vegetation types and were more likely to contain important wildlife resources (namely, plants with fruit/berries and vegetation planted to attract birds). Research in Australia also demonstrates differences between front and back yards, with front yards containing more “showy” plants and back yards containing more vegetables, fruits, and herbs (Daniels and Kirkpatrick 2006b). Back yards are where residents’ preferences and desires are more likely to be realized (Larsen and Harlan 2006; Larson et al. 2009), probably because they are less visible to neighbors and less subject to constraints from formal and informal institutions (e.g., homeowners associations, neighborhood norms).
We were especially interested in the unexpected differences between residents with and without outdoor cats. Residents with cats outdoors were more likely to have a birdfeeder, birdhouse, and plants intended to attract birds. Similar results have been reported for residents in Michigan and Arizona (Lepczyk et al. 2012), suggesting a broader trend. In general, pet owners report greater fondness for animals than people without pets (Bjerke et al. 2003), which may explain why they want to attract or support local birds. Although the negative effects of cats on wildlife have been documented in scientific literature (e.g., Stracey 2011; Loss et al. 2013) and public media outlets (e.g., “Cats kill up to 3.7 billion birds annually,” USA Today 30 Jan. 2013 and “The Evil of the Outdoor Cat,” New York Times 21 Mar. 2014), outdoor cat owners may not believe that their own pets harm birds or other species (McDonald et al. 2015), or they may believe that their cats are “just doing what’s natural” (Conniff 2014). Residents may also be unaware that the mere presence of cats can harm birds through indirect effects (e.g., birds reduced food delivery to nestlings after a domestic cat model was placed nearby, Bonnington et al. 2013). Previous work in our study area suggests that cats have an overriding negative effect on bird richness (Belaire et al. 2014), despite the fact that outdoor cats are also associated with resources such as birdhouses and bird feeders (Fig. 3).
Our results showed that between-yard variation is more challenging to explain than between-neighborhood variation. The yard-scale models had rather poor fit (R2 of 0.17–0.23), which suggests individual yard decisions are governed by a more complicated and obscure mix of factors. Our analysis at the transect scale “averages away” the complex drivers of individual yard variation—at this scale, our models explained substantially more variation (e.g., full model R2 of 0.71—0.83). Recent work suggests the degree of heterogeneity in residential landscapes depends on scale of analysis (Harris et al. 2012). This result also points to multi-scalar drivers of residential yard decisions, ranging from individual desires up to municipal policies or regional economic influences (Cook et al. 2012).
Importance of human-nature interactions for yards (and beyond?)
Perceptions of neighborhood birds explained significant variation in wildlife resources and back yard vegetation at both individual yard scale and neighborhood scale (Fig. 3). Our results align with recent research in the U.K. suggesting that wildlife strongly motivates yard activities. In that study, residents expressed satisfaction and pride that their yards attracted wildlife, and some felt responsibility to protect local species; in some cases, passion for wildlife could override social pressures related to yard neatness (Goddard et al. 2013). In addition, several studies have pointed to the positive feedback or reward mechanism that occurs when residents notice wildlife in their yards, leading them to further improve the wildlife resources in the yard (e.g., Head and Muir 2006; van Heezik et al. 2012; Cosquer et al. 2012; Goddard et al. 2013).
These results highlight the potential for human-bird interactions to catalyze stewardship activities. In general, people find birds easy to like and connect with (Bjerke and Ostdahl 2004), and people who feel more connected to nature may be more likely to engage in ecologically-friendly behaviors (Mayer and Frantz 2004). Previous research has shown that yard behaviors do not necessarily reflect people’s broader environmental values (Larsen et al. 2010; Cook et al. 2012), but local birds may represent a more relatable facet of the environment to which people can develop attachments. Dickinson et al. (2013a) suggested that people’s strong emotions about birds and other organisms may motivate environmental stewardship. For example, framing climate change in terms of potential harm to birds significantly increases people’s interest in carbon-footprint reducing activities (Dickinson et al. 2013a). Our results suggest that birds may motivate stewardship, in that values for birds appear to translate into on-the-ground effects on yards.
As our global population grows, the collective decisions of many individuals will increasingly drive environmental conditions in ways we are only beginning to understand. Ultimately, conservation requires that we understand and harness the power of people and the choices they make (Balmford and Cowling 2006; Schultz 2011). Yards represent a real opportunity to “imagine human actors as a force for environmental good” (Head and Muir 2007). Front yards, which are governed more by variables related to status, could become increasingly wildlife-friendly over time with shifting descriptive norms (observable behavior patterns that indicate what is typical or normal, Cialdini et al. 1990) in a neighborhood. Norms have been strongly correlated to pro-environmental behaviors such as recycling (Schultz 1999) and energy conservation (Nolan et al. 2008), and their potential to shift yard-related behaviors has been suggested by several researchers (e.g., Nassauer et al. 2009; Dickinson et al. 2013b). The conservation potential of back yards seems especially promising, since people tend to incorporate more vegetation types in their back yards, especially plants that support birds and other wildlife.
References
Allred SB, Ross-Davis A (2011) The drop-off and pick-up method: an approach to reduce nonresponse bias in natural resource surveys. Small Scale For 10:305–318
Balmford A, Cowling R (2006) Fusion or failure? The future of conservation biology. Conserv Biol 20:692–695
Belaire JA, Whelan CJ, Minor ES (2014) Having our yards and sharing them too: the collective effects of yards on native bird species in an urban landscape. Ecol Appl 24(8):2132–2143
Bjerke T, Ostdahl T (2004) Animal-related attitudes and activities in an urban population. Anthrozoös 17(2):109–129
Bjerke T, Ostdahl T, Kleiven J (2003) Attitudes and activities related to urban wildlife: pet owners and non-owners. Anthrozoos 16:252–262
Bonnington C, Gaston KJ, Evans KL (2013) Fearing the feline: domestic cats reduce avian fecundity through trait-mediated indirect effects that increase nest predation by other species. J Appl Ecol 50:15–24
Cameron RWF, Blanuša T, Taylor JE, Salisbury A, Halstead AJ, Henricot B, Thompson K (2012) The domestic garden—its contribution to urban green infrastructure. Urban For Urban Green 11(2012):129–137
Chowdhury R, Larson KL, Grove JM, Polsky C, Ogden L, Onsted J, Cook E (2011) A multi-scalar approach to theorizing socio-ecological dynamics of urban residential landscapes. Cities Environ 4(1):6–21
Cialdini RB, Reno RR, Kallgren CA (1990) A focus theory of normative conduct: recycling the concept of norms to reduce littering in public places. J Personal Soc Psychol 58(6):1015–1026
Clayton S (2007) Domesticated nature: motivations for gardening and perceptions of environmental impact. J Environ Psychol 27:215–224
Clucas B, Rabotyagov S, Marzluff JM (2014) How much is that birdie in my backyard? A cross-continental economic valuation of native urban songbirds. Urban Ecosyst. doi:10.1007/s11252-014-0392-x
Conniff R 2014. The evil of the outdoor cat. New York Times. 24 Mar. 2014
Cook EM, Hall SJ, Larson KL (2012) Residential landscapes as social-ecological systems: a synthesis of multi-scalar interactions between people and their home environment. Urban Ecosyst (6 August 2011):1-34
Cooper CB, Dickinson J, Phillips T, Bonney R (2007) Citizen science as a tool for conservation in residential ecosystems. Ecol Soc 12(2): 11. http://www.ecologyandsociety.org/vol12/iss2/art11/
Cosquer A, Raymond R, Prevot-Julliard A (2012) Observations of everyday biodiversity: a new perspective for conservation? Ecol Soc 17(4):2
Dahmus ME, Nelson KC (2013) Yard stories: examining residents’ conceptions of their yards as part of the urban ecosystem in Minnesota. Urban Ecosyst 17(1):173–194
Daniels GD, Kirkpatrick JB (2006a) Does variation in garden characteristics influence the conservation of birds in suburbia? Biol Conserv 133:326–335
Daniels GD, Kirkpatrick JB (2006b) Comparing the characteristics of front and back domestic gardens in Hobart, Tasmania, Australia. Landsc Urban Plan 78:344–352
Davies ZG, Fuller RA, Dallimer M, Loram A, Gaston KJ (2012) Household factors influencing participation in bird feeding activity: a national scale analysis. PLoS One 7(6):e39692
Dickinson JL, Crain R, Yalowitz S, Cherry TM (2013a) How framing climate change influences citizen scientists’ intentions to do something about it. J Environ Educ 44(3):145–158
Dickinson JL, Crain R, Reeve HK, Schuldt JP (2013b) Can evolutionary design of social networks make it easier to be ‘green’? Trends Ecol Evol 28(9):561–569
Dillman DA (1978) Mail and telephone surveys: the total design method. Wiley, New York
Fuller RA, Irvine KN, Davies ZG, Armsworth PR, Gaston KJ (2012) Interactions between people and birds in urban landscapes. In: Lepczyk CA, Warren PS (eds) Urban bird ecology and conservation. University of California Press, Berkeley
Gaston KJ, Fuller RA, Loram A, MacDonald C, Power S, Dempsey N (2007) Urban domestic gardens (XI): variation in urban wildlife gardening in the United Kingdom. Biodivers Conserv 16:3227–3238
Gaston KJ, Avila-Jimenez ML, Edmondson JL (2013) Managing urban ecosystems for goods and services. J Appl Ecol 50(4):830–840
Goddard MA, Dougill AJ, Benton TG (2013) Why garden for wildlife? Social and ecological drivers, motivations and barriers for biodiversity management in residential landscapes. Ecol Econ 86:258–273
Grove JM, Troy AR, O’Neil-Dunne JPM, Burch WR Jr, Cadenasso ML, Pickett STA (2006) Characterization of households and its implications for the vegetation of urban ecosystems. Ecosystems 9(4):578–597
Harris EM, Polsky C, Larson KL, Garvoille R, Martin DG, Brumand J, Ogden L (2012) Heterogeneity in residential yard care: evidence from Boston, Miami, and Phoenix. Hum Ecol 40:735–749
Head L, Muir P (2006) Suburban life and the boundaries of nature: resilience and rupture in Australian backyard gardens. Trans Inst Br Geogr 31(4):505–524
Head L, Muir P (2007) Edges of connection: reconceptualising the human role in urban biogeography. Aust Geogr 37(1):87–101
Hope D, Gries C, Zhu W, Fagan WF, Redman CL, Grimm NB, Nelson AL, Martin C, Kinzig A (2003) Socioeconomics drive urban plant diversity. PNAS 100(15):8788–8792
Hunter MCR, Brown DG (2012) Spatial contagion: gardening along the street in residential neighborhoods. Landsc Urban Plan 105:407–416
Kinzig AP, Warren P, Martin C, Hope D and Katti M (2005) The effects of human socioeconomic status and cultural characteristics on urban patterns of biodiversity. Ecol Soc 10(1): 23. http://www.ecologyandsociety.org/vol10/iss1/art23/5
Larsen L, Harlan SL (2006) Desert dreamscapes: residential landscape preference and behavior. Landsc Urban Plan 78:85–100
Larson KL, Casagrande D, Harlan S, Yabiku S (2009) Residents’ yard choices and rationales in a desert city: social priorities, ecological impacts, and decision tradeoffs. Environ Manag 44:921–937
Larson KL, Cook E, Strawhacker C, Hall SJ (2010) The influence of diverse values, ecological structure, and geographic context on residents' multifaceted landscaping decisions. Hum Ecol 38:747–761
Legendre P, Legendre L (2012) Numerical ecology, 3rd edn. Elsevier, Boston
Lepczyk CA, Mertig AG, Liu J (2004) Assessing landowner activities related to birds across rural-to-urban landscapes. Environ Manag 33(1):110–125
Lepczyk CA, Warren PS, Machabee L, Kinzig AP, Mertig AG (2012) Who feeds the birds? A comparison across regions. In: Lepczyk CA, Warren PS (eds) Urban Bird Ecology and Conservation. University of California Press, Berkeley
Lerman SB, Warren PS (2011) The conservation value of residential yards: linking birds and people. Ecol Appl 21(4):1327–1339
Loram A, Warren PH, Gaston KJ (2008) Urban domestic gardens (XIV): the characteristics of gardens in five cities. Environ Manag 42:361–376
Loss SR, Ruiz MO, Brawn JD (2009) Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol Conserv 142:2578–2585
Loss SR, Will T, Marra P (2013) The impact of free-ranging domestic cats on wildlife of the United States. Nat Commun 4:1396
Martin CA, Warren PS, Kinzig AP (2004) Neighborhood socioeconomic status is a useful predictor of perennial landscape vegetation in residential neighborhoods and embedded small parks of Phoenix, AZ. Landsc Urban Plan 69:355–368
Mayer FS, Frantz CM (2004) The connectedness to nature scale: a measure of individuals’ feeling in community with nature. J Environ Psychol 24:503–515
McDonald JL, Maclean M, Evans MR, Hodgson DJ (2015) Reconciling actual and perceived rates of predation by domestic cats. Ecol Evol 5(14):2745–2753
Micallef L, Rodgers P (2014) eulerAPE: drawing area-proportional 3-Venn diagrams using ellipses. PLoS ONE 9(7):e101717
Minor ES, Belaire JA, Davis A, Franco M, Lin M (2015) Socioeconomics and neighbor mimicry drive urban yard and neighborhood vegetation patterns. In: Francis R, Millington J (eds) Urban landscape ecology: science, policy and practice. Taylor & Francis, Oxon
Nassauer JI, Wang Z, Dayrell E (2009) What will the neighbors think? Cultural norms and ecological design. Landsc Urban Plan 92:282–292
Nolan JM, Schultz PW, Cialdini RB, Goldstein NJ, Griskevicius V (2008) Normative social influence is underdetected. Pers Soc Psychol Bull 34:913
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2012) vegan: Community ecology package. R package version 2.0-3. http://CRAN.R-project.org/package=vegan
Polsky C, Grove JM, Knudson C, Groffman PM, Bettez N, Cavender-Bares J, Hall SJ, Heffernan JB, Hobbie SE, Larson KL, Morse JL, Neill C, Nelson KC, Ogden LA, O'Neil-Dunne J, Pataki DE, Chowdhury RR, Steele M (2014) Assessing the homogenization of urban land management with an application to US residential lawn care. PNAS 111(12):4432–4437
Rudd H, Vala J, Schaefer V (2002) Importance of backyard habitat in a comprehensive biodiversity conservation strategy: a connectivity analysis of Urban green spaces. Restor Ecol 10(2):368–375
Schroeder HW, Ruffolo SR (1996) Householder evaluations of street trees in a Chicago suburb. J Arboric 22(1):35–43
Schultz PW (1999) Changing behavior with normative feedback interventions: a field experiment on curbside recycling. Basic Appl Soc Psychol 27(1):25–36
Schultz PW (2011) Conservation means behavior. Conserv Biol 25(6):1080–1083
Steele J, Bourke L, Luloff AE, Liao P, Theodori GL, Krannich RS (2001) The drop-off/pick-up method for household survey research. J Community Dev Soc 32(2):238–250
Stracey CM (2011) Resolving the urban nest predator paradox: the role of alternative foods for nest predators. Biol Conserv 144:1545–1552
Tratalos J, Fuller RA, Warren PH, Davies RG, Gaston KJ (2007) Urban form, biodiversity potential and ecosystem services. Landsc Urban Plan 83(4):308–317
U.S. EPA. 2012. Healthy yards, healthy lawns, healthy environment. http://www.epa.gov/reg3wcmd/pesticideslawn.htm. Accessed Nov 2013
van Heezik YM, Dickinson KJM, Freeman C (2012) Closing the gap: communicating to change gardening practices in support of native biodiversity in urban private gardens. Ecol Soc 17(1):34
van Heezik Y, Freeman C, Porter S, Dickinson KJ (2013) Garden size, householder knowledge, and socio-economic status influence plant and bird diversity at the scale of individual gardens. Ecosystems 16(8):1442–1454
Vergnes A, Le Viol I, Clergeau P (2012) Green corridors in urban landscapes affect the arthropod communities of domestic gardens. Biol Conserv 145:171–178
Wheeler B. 2010. lmPerm: permutation tests for linear models. R package version 1.1–2. http://CRAN.R-project.org/package=lmPerm
Zmyslony J, Gagnon D (2000) Path analysis of spatial predictors of front-yard landscape in an anthropogenic environment. Landscape Ecol 15:357–371
Acknowledgments
This study was based upon work supported by the National Science Foundation Grant DGE-0549245 and the University of Illinois at Chicago’s Department of Biological Sciences Elmer Hadley Graduate Research Award. We thank C. Watkins for advice in social survey design and delivery, H. Gin for help with survey distribution, and L. Vonderlinden for verifying yard characteristics of respondents. We also thank the many residents of Cook County, Illinois who graciously participated in the survey.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Belaire, J.A., Westphal, L.M. & Minor, E.S. Different social drivers, including perceptions of urban wildlife, explain the ecological resources in residential landscapes. Landscape Ecol 31, 401–413 (2016). https://doi.org/10.1007/s10980-015-0256-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0256-7