
Abstract
Context
The Florida Everglades has diminished in size and its existing wetland hydrology has been altered. The endangered snail kite (Rostrhamus sociabilis) has nearly abandoned the Everglades, and its prey, the apple snail (Pomacea paludosa), has declined.
Objective
We developed a population model (EverSnail) to understand apple snail response to inter- and intra-annual fluctuations in water depths over the Everglades landscape. EverSnail was developed as a tool to understand how apple snails respond to different hydrologic scenarios.
Methods
EverSnail is an age- and size-structured, spatially-explicit landscape model of P. paludosa in the Everglades. Landscape-level inputs are water depth and air temperature. We conducted sensitivity analyses by running EverSnail with ±20 % the baseline value of eight parameters.
Results
EverSnail was sensitive to changes in survival and water depth associated with reproduction. The EverSnail population varied with changes and/or differences in depth generally consistent with empirical data; site-specific comparisons to field data proved less reliable. A simulated 3-year wet period resulted in a shift in apple snail distribution, but little change in total abundance over the landscape. In contrast, a simulated 3-year succession of relatively dry years resulted in overall lower snail abundances.
Conclusions
Comparisons of model output to empirical data indicate the need for more data to better understand, and eventually parameterize, several aspects of snail ecology in support of EverSnail. A primary value of EverSnail is its capacity to describe the relative response of snail abundance to alternative hydrologic scenarios considered for Everglades water management and restoration.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The freshwater marshes of the Everglades historically covered over 10,000 km2 in southern Florida, but were reduced to half this area in the 1900s (Kushlan 1990). The Everglades is a seasonal marsh in which the flooded area increases during the wet season and decreases in the dry season, causing expansion and contraction of the aquatic food base that governs the seasonal cycles of much of its wildlife (Kushlan 1990; Sklar et al. 2002). Alteration of the extant Everglades hydrology, due to impoundment and diversion of water for agriculture and flood control, has resulted in a decrease in hydroperiod in some areas and ponding and increased hydroperiods in others (Sklar et al. 2002). These conditions have caused habitat degradation for many species of fish and wildlife (Sklar et al. 2002). One such species is the endangered Everglades Snail Kite (Rostrhamus sociabilis plumbeus Ridgway 1894, hereafter snail kite or kite), a raptor and dietary specialist that forages exclusively on snails in flooded wetlands (Sykes et al. 1995). Understanding the interactions and shifting distribution of snails and kites associated with inter- and intra-annual fluctuations in water depths over this large landscape is well served by landscape level modeling (DeAngelis et al. 1998). Our purpose is to combine landscape data with detailed population studies through modeling to evaluate the influence of hydrology on the population of Florida apple snails (Pomacea paludosa Say 1829) (Caenogastropoda: Ampullariidae), the main prey of the kite. While the modeling here is applied to apple snails in the Everglades, the approach can easily be generalized to other Everglades aquatic fauna or to other wetlands for which similar landscape data exist.
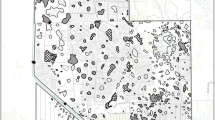
Improving habitat for the endangered Everglades Snail Kite is a key objective of Everglades restoration (NRC 2012). Snail kites in Florida exist as a contiguous population that occupies a network of wetlands from central Florida southward to Lake Okeechobee (LOKEE), the water conservation areas (WCAs), and Everglades National Park (ENP) (Sykes et al. 1995) (Fig. 1). Intra- and inter-annual shifts in kite wetland unit occupancy depend on changes in hydrology and habitat conditions that influence apple snail availability (Sykes et al. 1995). The desired outcome from an ecosystem restoration perspective is to have a network of kite foraging habitats that includes Everglades wetland units with healthy populations of apple snails (NRC 2012).

Major wetland units used by snail kites since approximately 1995. Stippling shows the area represented by EverSnail. WCA refers to Water Conservation Area. Wetland units shown are: A Lake Tohopekilega, B East Lake Tohopekilega, C Lake Kissimmee (LKISS), D Upper St. Johns Marsh, E Lake Istokpoga, F Lake Okeechobee (LOKEE), G Grassy Waters Preserve, with the following constituting the extant Everglades marshes, H WCA1, I Rotenberger and Holey Land Water Management Areas, J WCA2A, K WCA2B, L WCA3A, M WCA3B, and N portions of Everglades National Park. Scale is approximate. Two sites in WCA3A (*) represent site 13 in the south and site 18 approximately 23 km further north, referred to for site-specific comparisons of EverSnail output
How to balance the short-term needs for prey availability for kites with the long-term quality of emergent-marsh habitat is a critical question in snail kite population recovery and water management (Bennetts et al. 1998; Martin et al. 2008). Periodic drying, which renders snails unavailable to kites (Sykes 1979), is an essential driving force for development and maintenance of plant communities that support kites and other Everglades fauna (Gunderson 1994). The snails have the physiological capacity to survive, and a life history adapted to, these periodic drying events (Darby et al. 2008), and kites can move to find flooded wetlands that support their foraging needs (Rodgers et al. 1988; Takekawa and Beissinger 1989). However, extensive regional droughts significantly reduce kite foraging opportunities over large areas, resulting in negative demographic outcomes (Bennetts and Kitchens 1997; Martin et al. 2008). Consequently, research and analyses of the dry end of the hydrologic spectrum have dominated the kite and snail literature since the kites’ endangered listing in 1969 (Pomacea Project 2013). However, habitat degradation associated with extended periods of flooding, from approximately 1994–2000 in WCAs important for kite nesting, has also been described (Bennetts et al. 1998), and may have contributed to a decline in the snail kite population that began in the late 1990s (Martin et al. 2008).
A conceptual model of wetlands occupied by snail kites in peninsular Florida that describes fluctuations in habitat quality as water levels fluctuate, posits low quality during dry downs, a peak in quality several years after reflooding, but degradation after prolonged flooded conditions (Bennetts et al. 1998). Unfortunately, we lack sufficient empirical data to describe the details of appropriate wet conditions (e.g., seasonal depths, water ascension rates, duration of inundation) and periodic dry conditions (e.g., duration, frequency, timing) that support a restoration and management plan that supports snails and kites across the Everglades landscape (Pomacea Project 2013). In light of a greater than 50 % decline in the Florida snail kite population since 1999 (Martin et al. 2007), and an associated significant decline in the numbers of kites and kite nests in the Everglades (Martin et al. 2007), any information on hydrologic conditions that directly and indirectly influence apple snails is both highly pertinent and timely for Everglades restoration and snail kite recovery.
Here we present a landscape model of apple snail populations, called EverSnail, that explores the dynamic hydrologic conditions of Everglades wetland units that appear to support apple snail population persistence and abundance. The model focuses on the underlying driver of Everglades ecosystem function, temporal and spatial variability in water depth (DeAngelis 1994). We also explore simulated hydropatterns that warrant further research to better understand how to increase apple snail populations in support of snail kite recovery. Several studies have quantified a direct relationship between snail density and kite wetland patch occupancy and foraging success (Darby et al. 2006, 2012) as well as numbers of kite nests and fledged young (Cattau et al. 2014). Therefore, when exploring alternative hydrologic scenarios in EverSnail we can characterize these scenarios as more or less favorable to kites based on increases or decreases in apple snail abundance.
Methods
Model description
The age- and size-structured apple snail population within each spatial cell is represented as a vector consisting of 500 daily age-classes, with 500 days representing the approximate life span of Florida apple snails (for details on this and other pertinent details of snail life history, see Supplementary Material, Appendix 1);
where N i (t) is the number of apple snails in age class i in a given spatial cell at time t. Snail number in each age class is updated daily using the Leslie matrix equation,
where the matrix A is
and where a i,i+1 represents the fractional survival of a given one-day age class cohort from age class i to age class i + 1, and f i is fecundity (per capita offspring) of age class i. The temporal scale of EverSnail resolution is 1 day. The number of new 1-day old snails on each day is given by the summation,
starting from the first age class capable of reproduction, where the ‘20’ represents a 20-day delay for eggs to hatch. Survival from one age class to the next is given by
The snails in each age class have an attribute of size (linear dimension in mm), which increases deterministically (snails hatch out at 3 mm and adults reach 50 mm). The values of a i,i+1 and f i depend on both snail size and environmental conditions for which daily data exist (details below). Details of these dependencies and other aspects of EverSnail are described in Supplementary Material, Appendix 1, using the model description protocol of Grimm et al. (2006, 2010) Programming and file format details are presented in Supplementary Material, Appendix 2.
Space is represented by 253,125 spatial cells, each 400 × 400 m, which are contiguous and represent the Water Conservation Areas and ENP and some surrounding wetlands (Fig. 1). The individual spatial cells are assumed independent (no movement of snails between cells is assumed), so simulations can be performed either on the whole spatial extent, or on individual cells or subsets of cells, such as those for which empirical data on snail densities are available. The analyses presented here emphasized WCA3A, where the majority of Everglades snail density data has been collected (Pomacea Project 2013).
Parameterization of EverSnail was derived, as much as possible, from published empirical data (Supplementary Material, Appendix 1). We also cite information from a peer-reviewed report (Pomacea Project 2013) that summarized snail and kite trends and associations derived from linking information from multiple sources published over 30 years. Decisions about what to include in the population model reflect the availability of data on P. paludosa and also environmental data that serve as input to the model (e.g., water depth data). Exclusion of some parameters that likely influence snail reproduction, abundance and distribution (e.g., the influence of available food sources, macrophytes and periphyton, on snail growth, see Sharfstein and Steinman 2001) reflect limited empirical data on snail response as well as a lack of readily available environmental data suitable as inputs to the model.
Simulations
Simulations were performed for the period 1991–2011, or for a subset of this period, 1995–2005, the latter in order to save computing and output processing time on the numerous runs required for sensitivity analyses (details below). Daily water depth and temperature data were available for the entire Everglades Depth Estimation Network simulator (EDEN) spatial area for 1991–2011 (EDEN details in Supplementary Material, Appendix 3). Sensitivity analyses and other detailed comparisons were restricted to the simulation results for 400 × 400 m spatial cells that represent the empirical sampling areas designated as sites 13 and 18 in WCA3A (Fig. 1), so that comparisons could be made with data collected in the field. These two sites represent an approximate 30-cm topographical gradient in WCA3A, so that water depths at site 13 are almost always higher (Supplementary Material, Appendix 3). Simulation results that represent WCA3A, which is of most interest because of its traditionally heavy use by snail kites, are presented. Presented output from the simulations was limited to that from 2001 to 2005 following a few years of computational stabilization to rid the output of transient snail abundance due to initial model conditions. The period 2001–2005 also corresponds to when the majority of empirical data were available for sampling sites in WCA3A (2002–2005; noting that 2001 seasonal water depths influenced 2002 snail populations, Supplementary Material, Appendix 1).
Sensitivity analyses
In order to quantify the sensitivity of the model to estimated parameter values, we ran simulations with environmental inputs from 1995 to 2005. Four parameter values (k growth , k temp , Temp threshold , Depth max ) (Table 1) were set at +20 and −20 % relative to the baseline parameter value. In addition, the values of survival for the four snail size categories (Surv wet1, Surv wet2, Surv wet3, Surv wet4 ) were set to +0.002/day and −0.002/day relative to baseline survival (=0.99/day), hence +20 and −20 % relative to the baseline mortality. Results for two sites described earlier, 13 (representing lower ground elevations) and 18 (higher ground elevations), were used to report model sensitivity to these eight parameters. The results are reported for the total snail population (i.e., all size classes included). We compared quarterly average snail populations for baseline output to the 16 different sensitivity simulations. Sensitivity for the 16 model runs, expressed as quarterly percent change, was calculated as the percentage increase or decrease in snail population compared to baseline snail populations for 2001–2005 (representing 20 quarters) for the two sites. Overall average percent change (±SD, n = 20) for both sites was also reported.
Simulations of extended low and high water periods
Simulations were run using EDEN 1991–2011 input to estimate the total snail population response at the landscape level. We manipulated the EDEN water depth data as follows to simulate relatively dry and wet conditions in the Everglades:
-
(1)
For DrySim, simulations with the relatively low water year 2001 (details, Supplementary Material, Appendix 3, and see "Results" section) were artificially repeated in the next two subsequent years of model input (simulation year 2002 and 2003) to create a 3-year period of low water conditions,
-
(2)
For WetSim, simulations with the relatively high water year 2003 (details, Supplementary Material, Appendix 3, and see "Results" section) were artificially repeated in the two previous years of model input (simulation year 2001 and 2002) to create a 3-year period of high water conditions.
We used EverSnail to extrapolate the hydrologic influence on snail abundance and distribution at a larger landscape level and used the software EverVIEW (Romañach et al. 2014) to display snail abundance throughout WCA3A and several bordering wetland units. In order to see the landscape level effects of DrySim and WetSim, we chose to report output for a date (February 1) that reflects snail abundance prior to recruitment of hatchling snails for the breeding season (although egg production was possible starting January 15th, if temperatures were high enough) (Supplementary Material, Appendix 1). A snail population on this date represents all those snails recruited in the previous year as influenced by the previous year’s hydrologic conditions. We present a comparison of abundances over the landscape from February 1, 2001 (prior to DrySim or WetSim influences) versus February 1, 2004 (which follows simulated dry or wet conditions that occurred in simulation years 2001–2003). In addition, we calculated the percentage of landscape cells (400 m × 400 m) that exhibited a ≥ 25 % increase and ≥25 % decrease in snail abundance between 2001 and 2004 in order to quantify landscape-level effects of DrySim and WetSim.
Evaluating EverSnail performance compared to empirical data
We compared EverSnail output to available empirical data of snail density estimates from the field in two ways. First, we compared percent changes in EverSnail abundances for WCA3A site 18 (Fig. 1), which experienced a significant dry down in 2001, to the only dry down for which we have before and after empirical snail abundance data, LKISS (Fig. 1) in 1995–1996 (Darby et al. 2004). (Note that although we do have empirical data for WCA3A following dry year 2001, none of these sites had experienced a significant drying event, since they were at relatively low ground elevations in WCA3A.) We also performed a regression of empirical data from WCA3A from 2002–2005 (26 snail density estimates in 13 field sites) to EverSnail output for those same 13 sites. We used GIS-software to locate the correct EDEN cell that represented the 13 field sites, and divided the EverSnail abundance for those site-years by 160,000 m2 to yield a snail density estimate, i.e. numbers of snails per m2, for the selected 400-m × 400-m EDEN cell. Empirical snail density estimates were obtained from a well-established sampling protocol using 1-m2 throw traps. All 2002–2005 WCA3A empirical snail density estimates used for the EverSnail comparison have been reported in the literature as snail density per m2 (Darby et al. 2006) or used in statistical models (snails per m2 not shown, but analyzed) (Cattau et al. 2014).
Results
Baseline model performance
The simulated snail population exhibited intra- and inter-annual fluctuations consistent with the seasonality of egg cluster production (Supplementary Material, Appendix 1) and in response to variability in water depth within and between years (Fig. 2). The snail population response to water depths also varied by site; at Site 18 (higher elevations, lower water depths) the driest year (2001) had the greatest negative impact on the snail population, whereas at Site 13 (lower elevations, higher water depths) there was no negative effect on simulated populations (Fig. 2). Transient dynamics, prior to model output stabilization, were evident during the first three or four years of simulations and were excluded from any presentation or discussion of model output.

Simulated total snail population (black lines) and water depth (gray lines) for sites 18 (a) and 13 (b) in WCA3A for baseline simulation 1995–2005. Brackets show examples of a one-year population cycle. The box highlights the 2001 dry season, where site 18 experienced 55 days of dry conditions whereas site 13 had only 3 days of dry conditions. Dotted lines show the baseline minimum depth (10 cm) and baseline maximum depth (90 cm) associated with reproduction (see EverSnail Model, Supplementary Material, Appendix 1)
Sensitivity analyses
Snail abundance (based on quarterly average population size for two representative sites over 5 years) changed by less than ±10 % for the ±20 % adjustments to k growth , k temp , Surv wet1 , Surv wet2 , and Surv wet3 . Increasing Surv wet4 had no effect on snail abundance, nor did increasing Depth max to 108 cm (depths rarely exceeded 108 cm, even at the lowest ground elevations, Fig. 2). Decreasing Temp threshold (for reproduction) by 20 % changed abundance <6 %. The model was most sensitive to decreasing Surv wet4 from 0.99 to 0.988 days−1, resulting in rapid population decline to zero (Supplementary Material, Appendix 4). Decreasing Depth max by 20 % resulted in no reproduction above 72 cm depths and an associated 62 % ± 17 % decline in abundance in Site 13; however in Site 18 abundance declined by only 12 % ± 12 %, since depths rarely exceeded 72 cm. Raising Temp threshold from 17 to 20.4 °C resulted in ≈20 % ± 15 % abundance decline in the two sites, since input temperatures had to rise to a higher level to stimulate reproduction (details, Supplementary Material, Appendix 4).
Simulations of extended low and high water level conditions: landscape-scale assessment
The response by snails to 3 years of simulated consecutive high water conditions (WetSim) was mixed, resulting in a shift in distribution rather than an overall increase or decrease over the landscape compared to 2001 conditions (Fig. 3a). Note that in 2000, conditions were sufficiently wet (Appendix 3) to support robust population recruitment evident in February 2001 abundance estimates for most of WCA3B, ENP and WCA3A (Fig. 3a). Snail abundance in the southern portion of WCA3A then decreased during WetSim (Fig. 3b). This corresponds to increasing depths (at times exceeding 100 cm) that have negative impacts on snail recruitment in the previous years leading up to the 2004 population estimate. Little change was observed in WCA3B and ENP as a result of WetSim, since depths stayed between 15 and 60 cm for the peak breeding season (water depth details, Appendix 3). We calculated that 18.4 % of landscape cells exhibited a ≥25 % decline in snail abundance from 2001 to 2004 under WetSim, attributable primarily to declines in southern WCA3A (Fig. 3a, b). However, this was offset by a ≥25 % increase in snail abundance in 17.7 % of cells, e.g., evident in northern portions of WCA3A that have higher ground elevations that benefitted from WetSim depths increasing above the 10-cm threshold needed to support snail autecology.

Estimated total snail abundances (numbers per 400 × 400 cell) in the Everglades showing a initial population on February 1st 2001 for both WetSim and DrySim, b snail abundances for WetSim year 2004, which followed 3-consecutive years of relatively high water conditions, and c snail abundances for DrySim year 2004, which followed 3-consecutive years of relatively low water conditions (c). Snail abundances are indicated in color from lowest abundances (blue) to highest abundances (red)
The impacts of DrySim had a more consistent and negative impact on snails across the landscape than did WetSim. The snail population was reduced across WCA1, WCA2 and WCA3 (both A and B), and ENP by 2004 (Fig. 3c), especially at the higher ground elevations. Depths fell below 10 cm for most of April–June for the three DrySim low water years (2001, 2002, 2003) (Supplementary Material, Appendix 3), thereby terminating egg production in much of WCA3A, WCA3B, and ENP, and dramatically reducing abundance for those populations. By our calculations, 53.0 % of the landscape cells experienced a ≥25 % decline in snail abundance, while only 1.0 % exhibited a ≥25 % increase.
Evaluating EverSnail performance compared to empirical data
We compared EverSnail output to field-derived empirical data of apple snail populations in order to provide a level of model validation. The low water conditions of 2001 resulted in an April-June drying event in parts of WCA3A (e.g., see site 18, Fig. 2, with 50 days of depths <10 cm) with a subsequent 55 % reduction in EverSnail population abundance the following year (February 2002, Fig. 2). This is consistent with an 80 % decline in empirical snail abundance found on LKISS following dry conditions, resulting from a managed water level draw down, that lasted approximately 160 days during the 1996 breeding season (Darby et al. 2004) [This 80 % decline was observed in sites that experienced no treatment other than the draw down.]
Overall there was a positive relationship between 2002 to 2005 site-specific EverSnail output and empirical data from WCA3A (R2 = 0.28, n = 26, p = 0.0007, Fig. 4). For this period we had no before-and-after empirical data to compare responses to dry downs, but we did for a relatively high water year (2003, see Fig. 2, Site 13) that influenced the 2004 population. Both EverSnail estimates for the five low ground-elevation (GE) sites (≤190 cm above mean sea level, including Site 13) and the empirical data for these same sites, declined between 2003 and 2004 (Fig. 4). However, while EverSnail densities, on average (n = 5), declined by 39 % between 2003 and 2004, the empirical data showed a decline of 71 % (Fig. 4). Whereas the EverSnail simulated population at least partially recovers within a year following such a decline (most evident in Fig. 2), the empirical data indicate much longer recovery times. For example, the LKISS snail density estimates remained below 80 % of pre-dry down densities 1.5 years after water levels rose (Darby et al. 2004); the population may have been suppressed for a longer period, but the study ended. Similarly, WCA3A snails, as of 2012 (Darby, unpublished data), had not recovered from the 70–80 % decline in southern WCA3A sites documented in 2004 (empirical data, GE ≤ 190 cm, Fig. 4). These discrepancies may be explained by a number of working hypotheses presented below.

Estimated total snail densities (m−2) from empirical data (black bars, n = 26) in WCA3A from 2002 to 2005 for 13 sites in three different categories of ground elevation (GE): GE ≤ 190 cm (bottom panel), GE = 191–214 cm (middle panel), and GE ≥ 215 (top panel), which corresponds to an approximate north (higher ground elevations) to south (lower ground elevation) depth gradient. GE is reported as centimeters above mean sea level. EverSnail output for these same 13 sites is reported as snail density (m−2) (gray bars). Regression results (top panel, embedded graph) for empirical snail density versus EverSnail density was based on the 26 density estimates exhibited in the three panels
Discussion
EverSnail performance
EverSnail apple snail populations fluctuated in response to seasonal and inter-annual changes in water depth and temperature inputs. EDEN water depth data were essential for capturing the fluctuations in hydrologic conditions at a scale relevant to snail kites in the Everglades, which in turn serve as a metric of Everglades restoration success (NRC 2012).
In general, the positive and negative responses of the EverSnail population to hydrologic extremes over time and space were consistent with expectations based on empirical data (both field and laboratory derived). EverSnail confirms the direct impacts of drying events on apple snails; i.e., snails stop all activity, including mating and laying eggs (Supplementary Material, Appendix 1) and we see snail abundance decline (site 18, Fig. 2, and DrySim, Fig. 3). Validating any potential negative impacts of high water on snail populations has more uncertainty for the following reasons. (1) We lack sufficient empirical data from different wetlands at different times that show a negative trend following high water. (2) It remains unclear to what extent high water negatively impacts snail reproduction or other aspects of snail ecology, such as growth, movements, etc. This is because, unlike dry down effects, no controlled studies of high water impacts have been reported. It would be of great value to increase the amount of field-derived empirical data that better represents the Everglades landscape in support of EverSnail validation. It would be especially valuable to collect more before and after snail response data resulting from extreme hydrological events, which to date have been very limited (e.g., one dry down event on LKISS and one dry season high water event in WCA3A).
The model was sensitive to decreasing maximum survival for snails >16 mm during flooding (=Surv wet4 ). Increasing Surv wet4 had no effect on the snail population. Changing Surv wet1, Surv wet2, and Surv wet3 to the same degree as Surv wet4 (±20 % mortality) had marginal effects on snail abundance. This reflects that snails remain in juvenile size classes for a short time (less than 3 weeks for each of three juvenile size classes) compared to snails >16 mm (over 40 weeks spent in this size class). Consequently, the cumulative effects of a decrease in daily survival rates were much lower for the three smallest size classes modeled compared to the >16 mm size class. Parameterization of size-class dependent survival rates were based on predation rates measured one time in the field in flooded conditions (Darby et al., in prep). However, these predation rates likely change; for example after a dry down that could decrease predatory fish populations (Loftus and Ecklund 1994). Such a scenario has yet to be quantified for apple snails. Concentrations of snail kites and other predators of large apple snails likely impact snail populations, but this also has not been quantified (see Valentine-Darby et al. 2014).
EverSnail output deviated from empirical data in that there was more variability among sites with similar ground elevations (empirical data) compared to EverSnail predictions (Fig. 4). This likely reflects the many other variables that might influence snail abundance on a site to site basis (e.g., predation, vegetation, etc.) that EverSnail does not account for spatially. This may in part explain the relatively low regression coefficient for EverSnail output to predict empirical site density estimates. In addition, a significant discrepancy exists between EverSnail output and field-derived empirical data when it comes to snail population recovery following a decline. EverSnail exhibits substantially faster recovery from a dry down or extreme high water event. EverSnail populations rebounded within a year or less, once favorable hydrologic conditions returned, whereas populations observed in the field take years to recover. We hypothesize that negative biological effects associated with low densities may contribute to the lack of recovery of snails in WCA3A observed in the field. For example, there may be a mate-finding Allee effect (Stephens et al. 1999), wherein snails at low densities infrequently encounter potential mates (see Stoner et al. 2012), and therefore produce few eggs despite favorable habitat conditions. The discrepancies between EverSnail output and field-derived empirical data provide an opportunity to consider additional parameterization that would benefit from controlled experiments to test these hypotheses.
EverSnail demonstrates the value of incorporating spatially explicit inputs that result in a more accurate assessment of population shifts along environmental gradients within and between Everglades wetland units. The majority of apple snail trend data available from WCA3A have been restricted to lower ground elevations in southwestern WCA3A, so empirically we have not documented these unit-wide and system-wide landscape-level shifts. However, using EverSnail we can extrapolate what we know about snails from mesocosm and lab studies and spatially limited Everglades empirical data to a broader landscape. For example, shifts in apple snail distribution under high water conditions in EverSnail (WetSim) (Fig. 3) were consistent with data that showed that snail kites shifted their distribution to higher ground elevations during high water conditions in WCA3A in the 1990s (Bennetts et al. 2002). When considering the use of large wetland units like WCA3A by kites and snails, analyses of their response to environmental changes benefits from spatially explicit environmental data within wetland units as well as between wetland units.
EverSnail predictions of landscape-level snail response
The value of a model such as EverSnail is its capacity to describe relative patterns of population density across a whole landscape and across a number of years with different environmental conditions. EverSnail under WetSim conditions showed that snail populations that currently experience frequent drying events, like in portions of WCA3B, northern ENP, and northern portions of WCA3A, should respond positively to increased seasonal water depths (Fig. 3; Appendix 3). Overall, however, WetSim indicated that apple snail abundance over the entire Everglades landscape would not increase, but rather the distribution would shift as more favorable conditions were created in previously dry areas. That is, apple snails in higher ground elevation areas of the Everglades benefit from WetSim, but snail numbers at lower ground elevations in EverSnail were reduced due to negative impacts of high water on reproduction (e.g., vicinity of Site 13 in WCA3A). In contrast the DrySim scenario showed that frequent drying in these same areas of WCA3A, WCA3B and ENP resulted in negative impacts on snails that were not balanced by improved conditions elsewhere. The WetSim-based population increase in the previously dry areas was rapid in EverSnail. However, it is likely that actual population recovery would require a longer period of favorable hydrologic conditions to see a significant rebound from a current low density snail population, especially considering Allee effects. The capacity for apple snail dispersal could also limit the population rebound because of the current hydrologic disconnectivity of Everglades wetland units. This remains an unstudied aspect of apple snail ecology. Our WetSim and DrySim scenarios did not account for increased connectivity that is projected as hydrologic restoration in the Everglades proceeds (Interagency Modelling Center 2014). Due to our lack of understanding of dispersal mechanisms and associated colonization rates at large scales, we can only speculate as to how well apple snail populations recover as hydrologic conditions improve within wetlands and between wetland units as a result of increased connectivity.
Future directions to enhance EverSnail
Sensitivity analysis and discrepancies between model output and existing field data indicate that EverSnail would benefit from specific new empirical data that support more accurate quantification of snail response to abiotic and biotic influences. A fundamental concept is incorporated into our model development; that we can learn from models and use them as tools to guide future research (Orzack 2012). In the case of EverSnail, we recommend experiments and monitoring suited to support model enhancement in support of water management and apple snail population recovery in the Everglades.
The relationship between egg cluster production and depth in Everglades habitats needs to be clarified, especially given the high degree of sensitivity in the model to a negative effect of high water associated with the value set for maximum depth for reproduction (Depth max ). Empirical data supporting this relationship thus far were from low ground elevation sites in WCA3A over a 2 year period (Fig. 4). Validation of EverSnail output would benefit from comparisons with empirical data (currently not available) that were not used in developing the model (Rykiel 1996).
EverSnail does not include density dependent effects on growth or reproduction, which means that biological effects (e.g., the suspected mate-finding Allee effect) may in part explain inconsistencies between model output and empirical data. A density-dependent influence on growth and reproduction likely exists, as is common in freshwater aquatic snails (Dillon 2000), including evidence for density-dependent growth in P. paludosa and other apple snail species (see Conner et al. 2008). Additional empirical data to support parameterization of these density-dependent effects may improve EverSnail accuracy, especially for predicting the recovery of snails from a low density population.
Future iterations of EverSnail should consider the potential for the invasive exotic P. maculata (=P. insularum, Hayes et al. 2012) to have direct impacts on native apple snails or indirect effects on native snails via exotic snail alteration of habitat (reviewed in Pomacea Project 2013). So far the wetlands dominated by exotic Pomacea fall outside the area covered by EDEN and EverSnail. However, given the reliance of the majority of kite foraging and nesting on P. maculata in recent years (Cattau et al. 2010), and the fact that P. maculata have been found in the Everglades (although they are not dominant) (Darby, unpublished data from 2011 to 2014), P. maculata may become an issue in the Everglades landscape in coming years.
Predation clearly plays a role in regulating apple snail populations, but quantitative aspects of predation require more empirical data to support EverSnail parameterization. It may be that mortality rates from predation, across all size classes, can vary significantly between sites (and by more than ±20 % examined in the sensitivity analyses). It is clear that fish and several other aquatic predators target juvenile apple snails, in addition to snail kites and other large predators that target adult snails (Valentine-Darby et al. 2014). The role of predation on regulating apple snail populations should be considered as EverSnail is further developed. Doing so would also provide opportunities to model relationships between hydrology and predation rates.
Other landscape level inputs likely influence Florida apple snails, including but not limited to pH and calcium concentrations (Glass and Darby 2009), dominant vegetation type (Karunaratne et al. 2006), availability of flooded refugia from which populations may recover from dry downs (e.g. Gaff et al. 2004, Everglades fish), etc., but little or no empirical data exist to support EverSnail parameterization, and/or landscape-level inputs of most of these parameters do not exist. Even as these inputs and empirical relationships to apple snails become available, Orzack (2012) and Evans et al. (2012) recommend exercising caution against over-parameterizing simulation models. Doing so may improve model accuracy to specific locations and situations, but this may also reduce their general applicability. EverSnail was developed to explore and predict apple snail responses to intra- and interannual changes in water depths, the main driving force behind ecosystem function across the entire Everglades landscape, and future iterations will maintain that emphasis.
Conclusions
Similar to other population models that have been created for Everglades fauna (e.g., Doren et al. 2009; Jopp et al. 2010), our model has the potential to serve as a tool for understanding what influences apple snail population status and trends, as well as for facilitating management decisions, including projected ‘what-if’ hydrologic scenarios that emerge from proposed restoration activities and water management plans (DeAngelis et al. 1998). EverSnail was not designed for high precision site-specific predictions, but rather as a tool to evaluate the population-level responses of apple snails across the landscape to different hydrologic scenarios. However, there is value in comparing and contrasting site-specific simulation output to empirical data from Everglades sampling sites to identify discrepancies that may suggest a need for more research on what influences apple snail populations. Therefore, future work on EverSnail will be aimed at improving the accuracy of site-specific predictions.
For many tropical and subtropical wetlands, varying water levels govern resource availability for the fauna they support (e.g., Dodd 1993; Sykes et al. 1995; Swarts 2000). Management for conservation of wetland species like snail kites and other predators should be informed by accurate modeling of the changing food base (in this case apple snails) on the landscape (Doren et al. 2009; Jopp et al. 2010). Note that native Florida apple snails also serve as prey for a myriad of other Florida wetlands fauna (see Valentine-Darby et al. 2014), so interest in how snail abundance (and/or density) responds to water and habitat management extends beyond the needs of the endangered snail kite. EverSnail provides a tool to explore ecological trade-offs as hydrologic conditions range from relatively dry to wet, and our modeling framework could apply to many species that respond to some optimal wetland condition in between hydrologic extremes. EverSnail also provides a tool to explore trade-offs between alternative restoration plans that result in shifts in snail distribution and abundance across the Everglades landscape.
References
Bennetts RE, Kitchens WM (1997) Population dynamics and conservation of snail kites in Florida: the importance of spatial and temporal scale. Colon Waterbirds 20:324–329
Bennetts RE, Kitchens WM, DeAngelis DL (1998) Recovery of the snail kite in Florida: beyond a reductionist paradigm. Trans N Am Wildl Nat Resour Conf 63:486–501
Bennetts RE, Kitchens WM, Dreitz VJ (2002) Influence of extreme high water event on survival, reproduction and distribution of snail kites in Florida, US. Wetlands 22:366–373
Cattau CE, Martin J, Kitchens WM (2010) Effects of an exotic prey species on a native specialist: example of the snail kite. Biol Conserv 143:513–520
Cattau CE, Darby PC, Fletcher RJ Jr, Kitchens WM (2014) Reproductive responses of the endangered snail kite to variations in prey density. J Wildl Manag 78:620–631
Conner SL, Pomory CM, Darby PC (2008) Density effects of native and exotic snails on growth in juvenile apple snails Pomacea paludosa (Gastropoda: Ampullariidae): a laboratory experiment. J Molluscan Stud 74:355–362
Darby PC, Valentine-Darby PL, Percival HF, Kitchens WM (2004) Florida apple snail (Pomacea paludosa Say) responses to lake habitat restoration activity. Arch Hydrobiol 161:561–575
Darby PC, Bennetts RE, Karunaratne LB (2006) Apple snail densities in habitats used by foraging snail kites. Fla Field Nat 34:37–47
Darby PC, Bennetts RE, Percival HF (2008) Dry down impacts on apple snail (Pomacea paludosa) demography: implications for wetland water management. Wetlands 28:204–214
Darby PC, Fujisaki I, Mellow DJ (2012) The effects of prey density on capture times and foraging bout success of course-hunting adult snail kites. Condor 114:755–763
DeAngelis DL (1994) Synthesis: spatial and temporal characteristics of the environment. In: Davis SM, Ogden JC (eds) Everglades: the ecosystem and its restoration. St. Lucie Press, Delray Beach, pp 307–320
DeAngelis DL, Gross LJ, Huston MA, Wolff WF, Fleming DM, Comiskey EJ, Sylvester S (1998) Landscape modeling for Everglades ecosystem restoration. Ecosystems 1:64–75
Dillon RT (2000) Population dynamics and competition. In: Dillon RT (ed) The ecology of freshwater molluscs. Cambridge University Press, Cambridge, pp 171–226
Dodd CK (1993) Costs of living in an unpredictable environment: the ecology of striped newts Notophthalmus perstriatus during a prolonged drought. Copeia 1993:605–614
Doren RF, Trexler J, Gottleib A, Harwell M (2009) Ecological indicators for system-wide assessment of the greater Everglades ecosystem restoration program. Ecol Indic 9:2–16
Evans MR, Norris KJ, Benton TG (2012) Predictive ecology: systems approaches. Philos Trans R Soc 367:163–169
Gaff H, Chick J, Trexler J, DeAngelis D, Gross L, Salinas R (2004) Evaluation of and insights from ALFISH: a spatially explicit landscape-level simulation of fish populations in the Everglades. Hydrobiologia 520:73–87
Glass NH, Darby PC (2009) The effect of calcium and pH on Florida apple snail Pomacea paludosa (Gastropoda: Ampullariidae) shell growth and crush weight. Aquat Ecol 43:1085–1093
Grimm V, Berger U, Bastiansen F, Eliassen S, Ginot V, Giske J, Goss-Custard J, Grand T, Heinz SK, Huse G, Huth A, Jepsen JU, Jørgensen C, Mooij WM, Müller B, Péer G, Piou C, Railsback SF, Robbins AM, Robbins MM, Rossmanith E, Rüger N, Strand E, Souissi S, Stillman RA, Vabø R, Visser U, DeAngelis DL (2006) A standard protocol for describing individual-based and agent-based models. Ecol Model 198:115–126
Grimm V, Berger U, DeAngelis DL, Polhill JG, Giske J, Railsback SF (2010) The ODD protocol: a review and first update. Ecol Model 221:2760–2768
Gunderson L (1994) Vegetation of the Everglades: determinants of community composition. In: Davis SM, Ogden JC (eds) Everglades, the ecosystem and its restoration. St. Lucie Press, Delray Beach, pp 323–340
Hayes KA, Cowie RH, Thiengo SC, Strong EE (2012) Comparing apples with apples: clarifying the identities of two highly invasive Neotropical Ampullariidae (Caenogastropoda). Zool J Linn Soc 166:723–753
Interagency Modelling Center (2014) Central Everglades Planning Project (CEPP) tentatively selected plan model documentation report (DRAFT). US Army Corps of Engineers, Jacksonville, FL and South Florida Water Management District, West Palm Beach, FL
Jopp F, DeAngelis DL, Trexler JC (2010) Modeling seasonal dynamics of small fish cohorts in fluctuating freshwater marsh landscapes. Landsc Ecol 25:1041–1054
Karunaratne LB, Darby PC, Bennetts RE (2006) The effects of wetland habitat structure on Florida apple snail density. Wetlands 26:1143–1150
Kushlan JA (1990) Freshwater marshes. In: Myers RL, Ewel JJ (eds) Ecosystems of Florida. University of Central Florida Press, Orlando, pp 324–363
Loftus WM, Ecklund AM (1994) Long-term dynamics of an Everglades small-fish assemblage. In: Davis SM, Ogden JC (eds) Everglades, the ecosystem and its restoration. St. Lucie Press, Delray Beach, pp 323–340
Martin J, Kitchens WM, Hines JE (2007) Importance of well-designed monitoring programs for the conservation of endangered species: case study of the snail kite. Conserv Biol 21:472–481
Martin J, Kitchens WM, Cattau CE, Oli MK (2008) Relative importance of natural disturbances and habitat degradation on snail kite population dynamics. Endanger Species Res 6:25–39
NRC (National Research Council) (2012) Progress toward restoring the Everglades: the fourth biennial review. The National Academy Press, Washington
Orzack SH (2012) The philosophy of modeling or does the philosophy of biology have any use? Philoso Trans R Soc 367:170–180
Pomacea Project, Inc (2013) Literature review of Florida apple snails and snail kites, and recommendations for their adaptive management. Final Report. Submitted to the National Park Service, Everglades National Park. Available from http://www.nps.gov/ever/learn/nature/upload/ASS10-3FinalReportSecure.pdf. Accessed 5 May 2015
Rodgers JA, Schwikert ST Jr, Wenner AS (1988) Status of the snail kite in Florida: 1981–1985. Am Birds 42:30–35
Romañach SS, McKelvy M, Conzelmann C, Suir K (2014) EverVIEW data viewer: a visualization framework and exploration tool for decision support of large-scale biological planning. Environ Model Softw 62:221–229. doi:10.1016/j.envsoft.2014.09.008
Rykiel EJ Jr (1996) Testing ecological models: the meaning of validation. Ecol Model 90:229–244
Sharfstein B, Steinman AD (2001) Growth and survival of the Florida apple snail (Pomacea paludosa) fed 3 naturally occurring macrophyte assemblages. J North Am Benthol Soc 20:84–95
Sklar F, Melloy C, VanZee R, Gawlik DE, Tarboton K, Rudnick D, Miao S, Armentano T (2002) The effects of altered hydrology on the ecology of the Everglades. In: Porter JW, Porter KG (eds) The Everglades, Florida Bay, and coral reefs of the Florida Keys. CRC Press, Boca Raton, pp 39–82
Stephens PA, Sutherland WJ, Freckleton RP (1999) What is the Allee effect? Oikos 87:185–190
Stoner AW, Davis MH, Booker CJ (2012) Negative consequences of Allee effect are compounded by fishing pressure: comparison of queen conch reproduction in fishing grounds and marine protected area. Bull Mar Sci 88:89–104
Swarts FA (2000) The Pantanal: understanding and preserving the world’s largest wetland. Paragon House, St. Paul
Sykes PW Jr (1979) Status of the Everglade kite in Florida- 1968–1978. Wilson Bull 91:495–511
Sykes PW Jr, Rodgers JA Jr, Bennetts RE (1995) Snail Kite (Rostrhamus sociabilis). In: Poole A, Gill F (eds) The birds of North America. The Academy of Natural Sciences, Philadelphia, and The American Ornithologists’ Union No 171, Washington
Takekawa JE, Beissinger SR (1989) Cyclic drought, dispersal, and the conservation of the snail kite in Florida: Lessons in critical habitat. Conserv Biol 3:302–311
Valentine-Darby PL, Kell SE, Darby PC (2014) Predation on Florida apple snails (Pomacea paludosa) by native and non-native aquatic fauna, and predator-prey size relationships. Fla Sci 78:47–56
Acknowledgments
We appreciate the support of Ronnie Best (USGS, retired) and Craig Conzelmann (USGS) in facilitating the collaboration for this effort. We thank Alicia LoGalbo (National Park Service, Everglades), Rena Borkhataria (University of Florida), and Wingrove Duverney (USGS contractor) for their input into the formulation of this model. Pam Telis (US Army Corps of Engineers) provided helpful information regarding the use of EDEN water depth data. Bethany Wight (University of West Florida) provided technical assistance regarding site-specific hydrologic and snail density data. David Bucklin (University of Florida) created the Figure-1 map. We appreciate the comments from three anonymous reviewers and James Beerens (USGS) on drafts of this manuscript. Funding from the USGS National Climate Change and Wildlife Science Center project “La Florida: A Land of Flowers on a Latitude of Deserts” provided some support for DLD. SSR and DLD were supported by the USGS’s Greater Everglades Priority Ecosystem Science program. Views expressed here do not necessarily represent the views of the U.S. Fish and Wildlife Service. Any use of trade, product, or firm names in this article is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Darby, P.C., DeAngelis, D.L., Romañach, S.S. et al. Modeling apple snail population dynamics on the Everglades landscape. Landscape Ecol 30, 1497–1510 (2015). https://doi.org/10.1007/s10980-015-0205-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0205-5