
Abstract
This paper analyses the importance of the South Central Andean High Puna megapatch, above 4000 masl, in the history of late Pleistocene exploration and colonization of the Atacama Desert, including the contrasting habitats that exist towards the hyperarid Pacific Ocean coast or the ecosystems bordering the tropical forests, on the western and eastern sides of the Andes, respectively. These ecosystems, which were firmly established by the end of the Pleistocene, are examined as key factors in the history of human peopling. The social, demographic and climate conditions associated with the peopling processes are discussed in relation to the appropriate technological, subsistence and settlement strategies developed by pioneer populations, who for their initial settlement selected highly productive patches where water and fauna converged. Based on concepts derived from metapopulation theory, all relevant archaeological paleoecological data from both sides of the Andean Cordillera are presented and discussed. It is concluded that the pioneer occupation of the high Puna megapatch was a gradual process, related to an emergent Andean human mobility system that connected a wide range of altitudinally staggered habitats. Moreover, we suggest that divergent cultural trajectories evolved since the early Holocene, affecting highland, lowland and coastal habitats.
Resumen
El presente trabajo analiza la importancia del megaparche de la alta puna de los Andes sur-centrales, por encima de los 4.000 m snm., en la historia de la exploración y colonización pleistocena del Desierto de Atacama, incluyendo los contrastantes hábitats que se ubican hacia la costa hiperárida del océano Pacífico o los ecosistemas yuxtapuestos hacia los bosques tropicales, en las vertientes oeste y este de los Andes, respectivamente. El papel de estos ecosistemas, mucho más continuos a finales del Pleistoceno, se examinan como factores clave en la historia del poblamiento humano. Se discuten las condiciones sociales, demográficas y climáticas asociadas a los procesos del poblamiento en relación con las estrategias tecnológicas, de subsistencia y de asentamientos más adecuadas desarrollados por las poblaciones pioneras, quienes en esta fase inicial seleccionaron parches de alta productividad donde confluían el agua y la fauna. Sobre la base de conceptos derivados de la teoría metapoblacional, se presentan y discuten todos los datos arqueológicos y paleoecológicos relevantes de ambos lados de la cordillera andina. Se concluye que las ocupaciones pioneras del megaparche de la puna ocurrieron a través de un proceso gradual, relacionado con un sistema emergente de movilidad humana andina que conectó una amplia gama de hábitats escalonados altitudinalmente. Además, se sugiere la existencia de trayectorias culturales divergentes para el Holoceno temprano, que afectaron tanto los hábitats de tierras altas como de tierras bajas y de la costa.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Si contemplan la pampa y sus rincones
verán las sequedades del silencio,
el suelo sin milagro y Oficinas vacías,
como el último desierto
If you contemplate the pampa and its surroundings
you will see the dessication of silence,
the ground without miracle and empty camps,
like the ultimate desert
Luis Advis, Cantata Santa Maria, Relato 1, 1970
The colonization of South America took thousands of years, and the process is regarded by Santoro et al. (2017) as not completely finished. The earliest generally accepted dates for this process are around 14,000–13,000 cal BP (Politis et al., 2016), but it is probable that people explored parts of South America even earlier (Dillehay et al., 2015). Human dispersal across the vast expanses of the continent may be the result of both indirect and direct movements, including back-and-forth processes. For several reasons, including human requirements for fresh water, fuel and food, it is possible that some South American habitats, including the Andean highlands, took longer than others to be fully settled. The research question is how the South American highlands were explored and colonized. We are looking at an area that goes from the Pacific Ocean up to above 4000 masl along the western Andean slope, spanning more than 12° of latitude (Latorre et al., 2013). One of our aims in this paper is to show that the pioneer occupation of the highlands above 4000 masl was a gradual process.
It is becoming clear that simple models are no longer sufficient for understanding the order in which different regions were occupied. No matter how counterintuitive it is, although habitat quality is the best determinant of move distances among hunter-gatherers (Grove, 2009), the best habitats were not rapidly settled. This is probably because only some of them were explored and recognized during the initial human forays by the end of the Pleistocene. Given the low population levels of the first explorers of the continent (Goldberg et al., 2016), many good habitats may not even have been sighted for hundreds of years. Several other places were possibly abandoned after exploration, and only a few were selected for settlement. Conversely, many regions were probably unoccupied for centuries or millennia, for instance, the core of the Atacama Desert (Latorre et al., 2013; Santoro et al., 2017; Tully et al., 2019; Workman et al., 2020). It is difficult to identify archaeological remains corresponding to exploration of new land, since with few exceptions these are elusive for reasons of low frequency, visibility and preservation (Borrero, 1989–1990; Meltzer et al., 2002).
Metapopulation thinking will be the overarching theoretical framework within which we discuss this colonization process. Accordingly, this paper does not include new archaeological data and does not formally summarize previously recorded data. Instead, it is a review of selected sites and studies that we integrate to offer a cohesive view of a large area under the umbrella of metapopulation theory (Simberloff, 1988).
As stated by Kilby, when he was trying to recognize an exploration phase during the Clovis period in North America, evidence of explorers is likely to be ‘swamped by the more abundant Clovis record of inhabitants who were familiar with their range’ (Kilby, 2014, p. 216). In other words, what we see in the archaeological record mostly results from activity by people whose long-term habitation of an area has led to a high level of knowledge and skill in management of local and extra-local environmental resources. This is basically the case for Atacama and NW Argentina, where the earliest known recorded occupations indicate good handling of the environment and its resources (Kulemeyer & López, 1997, p. 18; Aschero, 2014; Yacobaccio & Morales, 2011; Latorre et al., 2013; Santoro et al., 2016; Joly et al., 2017; Cartajena et al., 2014; Souza et al., 2021; López Mendoza et al., 2021; Loyola et al., 2017; Loyola et al., 2018; Santoro et al., 2019). A good understanding of the local environment was also apparent for other locations in the highlands, such as the Pucuncho basin in Peru (Rademaker & Hodgins, 2018), or Cueva Bautista in southwestern Bolivia (Capriles et al., 2016).
The importance of megapatches (sensu Beaton, 1991) for an understanding of the early peopling is well recognized in the archaeological literature, since it is probable that the first explorers focused on these large homogeneous units. Osorio and colleagues (Osorio, Steele, et al., 2017; Osorio, Capriles, et al., 2017) presented two main related hypotheses. The first is focused on the existence of a megapatch in the South Central Andean High Puna (SCAHP) ‘extending as far north as the highland of the Peruvian Central Andes’ (Osorio, Steele, et al., 2017; Osorio, Capriles, et al., 2017, p. 42) and up to Ipilla in the south, basically the beginning of the Salt Puna (with some differences east and west of the Andes) (see Osorio, Capriles, et al., 2017, p. 2). The SCAHP is characterized by dry environments in which there is a gradient in atmospheric pressure that declines logarithmically with altitude, while temperature decreases at a known rate (Osorio, Steele, et al., 2017; Osorio, Capriles, et al., 2017, p. 45). More importantly, these dry environments are located near large areas that are also relatively homogeneous, like the Pacific coast or the Atacama Desert. The basic resources of the SCAHP are medium-sized camelids like vicuña (Vicugna vicugna), guanaco (Lama guanicoe), and deer (Hippocamelus antisensis); small-sized animals like vizcachas (Lagidium), wild guinea pig (Cavia), and Ctenomys; and birds. The second hypothesis is that this megapatch forms a ‘late Pleistocene dispersal corridor’, perhaps parallel to the Pacific coastal corridor (Osorio, Steele, et al. 2017; Osorio, Capriles, et al., 2017, p. 42). In the end, this megapatch became a possible habitat ‘bridge’ to the eastern side of the Andean mountains (Osorio, Steele, et al., 2017; Osorio, Capriles, et al., 2017, p. 51).
All this suggests that the appropriate context to discuss the process of peopling is at the scale of the megapatch, a situation that invites us to consider the modes by which people found their way into the megapatch itself, and the ways in which it was used. We believe that the metapopulation concept offers a useful way to discuss these issues.
The process by which an area is peopled is rarely unidirectional. The direction of population advance can be considered the vector outcome of multidirectional drift. Factors like the fission of bands, extension of hunting ranges or the search for resources could be the primary causal mechanisms for moving. In what Kelly describes as ‘giving up a known environment for an unknown environment’ (Kelly, 1999, p. 124), these people certainly had a starting point outside the SCAHP megapatch. If the hyperarid coast was the starting point, in what today is southern Peru or northern Chile, then there will be a vector oriented toward the east or southeast (Fig. 1). The expected result of the initial forays would be a sparse coverage of the progressively occupied lands, not necessarily concentrated in the best places. This process may involve a geographical break—a very marked ecological and/or climatic change between previously occupied areas (probably coastal and/or the intermediate depression between the Pacific coast and the Andes, corresponding to the core of the Atacama Desert) and still empty megapatches (Figs. 2, 3, 4, 5). Osorio and colleagues (Osorio, Steele, et al., 2017; Osorio, Capriles, et al., 2017, p. 42) refer to a break at 2500 masl, ‘with dramatic shifts in precipitation … vegetation and animal communities’. This is also the break above which ‘humans begin to experience hypoxia’ (Pearsall, 2008, p. 113), and certainly entails a substantial change for human adaptation.

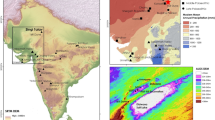
The Atacama Desert within the South Central Andean region in western South America, encompassing the high puna, the hyperarid Pacific Ocean coast and the tropical forest megapatches (key in the history of late Pleistocene exploration and colonization of this continent) of which the principal archaeological sites are listed: (1) Quebrada Maní 12 (QM12); (2) Chipana-1 quarry and workshop; (3) Pampa Ramaditas 5; (4) Quebrada Maní 35 (QM35); (5) Charaña; (6) Hakenasa; (7) Patapatane; (8) Las Cuevas; (9) Pampa El Muerto 15; (10) Ipilla; (11) Camarones; (12) Tiliviche; (13) Aragón; (14) Quebrada Blanca; (15) Cueva Bautista; (16) Cerro Kaskio; (17) Tuina; (18) Caldera Vilama; (19) Zapaleri/Laguna Blanca; (20) San Lorenzo; (21) Tulan; (22) Imilac; (23) Punta Negra; (24) Cueva Yavi; (25) Barro Negro; (26) Inca Cueva 4; (27) Pintoscayoc 1; (28) Hornillos 2; (29) León Huasi; (30) Huachichocana 3; (31) Quebrada Seca 3; (32) Salamanca; (33) Peñas de las Trampas 1; (34) Cueva Cacao IA; (35) Taller Puesto Viejo 1

Mouth of a seasonal ravine (Camiña canyon) dependent on limited rainfall in the high Andes, 150 km to the east. This hyperarid landscape of the Pacific coast contrasts with the bountiful marine ecosystem attached to the Atacama Desert in northern Chile, key for late Pleistocene human expansion

Pampa del Tamarugal basin, currently the most hyperarid area on Earth, between the Pacific coast and the Andes. By the end of the Pleistocene it was rich in vegetation and animals (now extinct or extirpated), of which a wooden and b bone remains are still found in situ

The Loa river canyon in Pampa del Tamarugal, which still transverses the core of the Atacama Desert, was crucial as a paleowetland for human expansion by the end of the Pleistocene in South America. Water from the Loa basin has been diverted for mining industry, farming, cities and tourism

a Puna Seca in the altiplano of Arica is even today rich in wild hunting resources and home of llama and alpaca herders, permanently living at 4500 masl, since around 5000–4000 years ago; b the contrasting altiplano of Puna Salada, where, as a consequence of long-term freezing annual temperatures and lower rainfall rates, no permanent human colonization has occurred
We now turn to a discussion of how the SCAHP was accessed, which was, in turn, an important step for the colonization of Atacama. This first became possible when local freshwater, in the form of seasonal riparian floodwater, paleo wetlands and lakes—surrounded by arbustive and woody vegetation—formed oases. These were places with abundant birds and mammals, including megafauna, which attracted people. These wetlands in the Chilean hyperarid coast (0–900 masl), or at Pampa del Tamarugal (900–2200 masl) in the northern Chile Atacama Desert, were probably important in the colonization of the highlands. It is postulated that movements of people within the SCAHP megapatch started somewhere in the northern highlands. However, migratory processes towards the SCAHP could also have originated in other areas of South America, such as the Atacama Desert coast or the Amazonian tropical forest.
Although megapatches are defined as homogeneous units, it is recognized that they present internal variability at different ecological scales. Accordingly, there are places within megapatches where initial establishment or longer settlement by humans is more likely to occur. This is probably due to a combination of basic conditions that require few innovations, and the presence of drinking water, fuel and food (Osorio, Steele, et al., 2017; Rademaker & Moore, 2019). Gravity models can be used to make sense of some of the incongruences in the distribution of these resources (Jochim, 1976). It is suggested that trial-and-error—expensive but necessary when exploring new lands—was among the first tactics to be selected when experimenting with new conditions (Borrero, 2011a, p. 220). Additionally, the transfer value of different classes of knowledge about particular resources such as animals or plants (Steele & Rockman, 2003) makes them more or less appropriate for the new arrivals. In any case, after some time an adjustment to the new habitat can be reached. Only later does the presence of neighbors become a major variable. Social, demographic and other compromises were probably associated with this later stage of colonization.
Metapopulation and Paleoenvironment
We will now consider the utility of the framework offered by metapopulation theory for making sense of both the multidirectional process of access to the SCAHP and the functioning of human groups within the megapatches. This body of knowledge, which is usually applied to conservation efforts, derives from studies of minimally viable populations, with a focus on refugia and connectivity (Simberloff, 1988). A metapopulation framework that takes into account the existence of several local sub-populations, distributed over different ecosystems, offers an appropriate framework for the study of disjunct distributions of social entities, which in turn refers to a ‘group of subpopulations with movement between the groups much less than movement within them’ (Simberloff, 1988, p. 480). Metapopulation, as a population of populations, spreads along ecological patches and does not depend on an external source of individuals. It should not be assumed, however, that such people were isolated from the rest of South America, because if practical help was required, this would have to be provided within the metapopulation, which highlights a degree of autonomy for day-to-day subsistence, but does not imply that interaction with people and sources outside a given metapopulation will be cut off.
Sub-populations were presumably tethered to rich life zones in Atacama, like coastal river mouths (Fig. 2) or the former late Pleistocene wetlands in the intermediate depression between the Coastal and Andean Cordilleras (Figs. 3, 4), the core of the Atacama Desert, and the highlands (Fig. 5). This may have been particularly true during the paleoecological conditions of the Central Andean Pluvial Event (CAPE) I and II (Quade et al., 2008; Placzek et al., 2009; Gayo et al., 2012; Latorre et al., 2013; Pfeiffer et al., 2108; Tully et al., 2019; Workman et al., 2020). These pluvial events occurred at the end of the Pleistocene (17,600–13,800 cal BP, 12,700–9,700 cal BP, respectively) and created distinct oases that sustained freshwater and vegetation attractive to faunas and humans. These oases also included riparian woodlands and marshy floodplains. The chronology of human settlement indicates that they are linked to the second CAPE event (Gayo et al., 2012; Tully et al., 2019; Workman et al., 2020; de Souza et al., 2022). Thus, it is particularly important to emphasize that the metapopulation concept helps us to understand human survival under adverse and contrasting conditions, like the scarcity of freshwater which characterizes the ecological belts on both sides of the Andes.
A metapopulation scheme, which is characterized by geographically separated sub-populations ‘interconnected by patterns of gene flow, extinction, and recolonization’ (Whittaker, 1998, p. 199), makes sense when considering the highly ranked nature of late Pleistocene habitats. It is also consistent with the previous observation that the known archaeological sites can only be explained ‘within a much larger system of land use and natural resource exploitation, that may have included mobility networks [with] the western … and the eastern … slopes of the Andes’ (Capriles et al., 2016, p. 473; Capriles et al., 2019). Such a framework hinges on the fact that the available technological and behavioral adaptations and basic gene pool were shared by a regionally widespread metapopulation. Indeed, Vivien Standen’s consideration of mating prospects as one of the causes for circulation between the lowlands and the highlands (Standen et al., 2018) is based on that basic survival fact. The use of corridors for behavioral necessities like the movement of foodstuffs, circulation of information, search for mates or access to buffer zones amounts to what in metapopulation theory is called the Rescue Effect. Indeed, this spatial arrangement is not necessarily always functional, given the possibility that catastrophes or negative traits may also be dispersed through corridors (Simberloff, 1988, p. 478). In any case, megapatch corridors are crucial components of the metapopulations, because their existence ensures the viability of a system of disjunct oases for human survival.
If we examine the oldest archaeological evidence (Fig. 1), it is clear that hunter-gatherers were recurrently occupying the highlands after 10 ka and perhaps earlier (Grosjean et al., 2005; Núñez et al., 2002; Santoro et al., 2017), a situation that was surely preceded by still unknown exploratory and logistical incursions.
One probable mode of exploration, which we call ‘indirect exploration’, takes the form of gradually extending the sectors of exploited land in any given year. At some point members of previously established metapopulations located below 2500 masl participate in systematic forays into the uplands, indicating long-distance logistical mobility (Lovis et al., 2005), whether from the west (Aldenderfer, 1998, p. 18) or the east (Yacobaccio, 2017, p. 39).
Wide corridors of some sort are required for such expansions to occur. These are places adequate for transit, whose topography minimizes circulation difficulties and which are not necessarily linear. In this way, strategically important transit camps could have been established along highland rivers, on lakes or adjacent to wetlands, serving to support open-site or cave camps; in time, these transitory excursions transformed into a colonial occupation of the land. These advanced processes were, perhaps, based on a point-to-point pattern of movements (cf. Binford, 1982), which seems to be adequate to the low productivity and patchy highland environments of the late Pleistocene.
The creation of empty quarters is a by-product of that mode of advance. These are places that were ignored for regular settlement because they were not productive, in the sense that they offer a limited supply of food or water (Collignon, 1996), or perhaps left as buffers because they only provided some of the critical resources. Of course, they may have been visited fleetingly or in transit, or simply not explored during the Pleistocene.
Flexible Behaviors, Risk and Uncertainty
What we know of the early inhabitants of America suggests that they possessed sufficient behavioral and technological flexibility to effectively interact simultaneously with a range of different ecologies. Indeed, generalized adaptive knowledge allowed transit through changing habitats, using a variety of shelter technologies, hunting and knapping skills, and navigational skills (Borrero, 2011a, p. 220; Kelly, 2003; Hiscock, 2008, p.122; López Mendoza et al., 2021). Together these constituted a wide spectrum of potential sleeping technologies that were probably not in use all the time, but that could be activated when required. As observed by Santoro and collaborators (Santoro et al., 2017, p. 29), ‘adaptive strategies exist in the space of possibilities for human life defined by the supply of ecosystem services’, supporting the idea that some of these strategies will fare better than others in different times and places. Indeed, many technologies are not characteristic of particular periods (as assumed by many models) but were simply used when required. Moreover, explorers must have had at their disposal a set of classic general and specific behavioral responses to risk (that is, anticipated problems), such as caching, sharing or clothing (Hiscock, 2013).
Aschero suggested that people with multi-ecological skills from different regions on the eastern side of the Andes converged on the Puna. He thought that those multiple skills helped early Puna foragers to minimize ‘risks of environmental stress’ by anticipating them (Aschero, 2014, p. 26). We use the term risk here contrastively with uncertainty, regarding a risk as a foreseeable problem that people have some idea in advance how to deal with (Cashdan, 1990). People are always prepared for risk, and at the end of the Pleistocene this issue was critical, as most of the people were constantly moving within uncertain scenarios. Conversely, coping with uncertainty that is, unexpected situations (Cashdan, 1990)—which is the inevitable result of changing megapatches, is a completely different issue.
Uncertainty deals with completely unknown environments, whose exploitation requires a learning curve. It must be remembered that for someone who does not know high altitude, the Precordillera and the Altiplano present a range of life zones that need to be decoded. Only decoding allows for adaptation, which means creating conditions for safe traveling between water sources or making adequate decisions concerning food, fuel or raw materials.
Under conditions of uncertainty, the role of embedded strategies can be extended to enable the selection of tactics and strategies that are rarely used, or the application of others that may require technical and social innovations. Needless to say, mistakes can easily be made. As a result, group size or even full adaptations might be completely redefined. There are several implications of the differentiation between the concepts of risk and uncertainty. First, human explorers always benefited from their background knowledge about how to deal with risky situations. This knowledge was probably recurrently applied throughout the exploration of megapatches. Within the Andean altitudinal gradient, however, it was difficult to be ready for all contingencies. The highlands presented unknown challenges, including hypoxia, very cold weather and changing distributions of resources. In the end, the capacity to deal with uncertainty determined the success of the colonization of the highlands.
The Attraction of the Highlands
The early human exploration of high-altitude environments was complicated not just by changes in the structure of resources, including a diminution of primary productivity in comparison with the coast, but also by human physiological requirements. Extreme forms of risk—perhaps accompanied by a measure of uncertainty—are among the classic difficulties of colonizing mountain habitats. These include changes in temperature, lower barometric pressure and rugged topography (Fig. 5). Aldenderfer lists the specific stresses produced by high altitude: ‘shortness of breath, headache, and nausea to, in very extreme instances, death due to pulmonary edema’ (Aldenderfer, 1998, p. IX). It is well known that hypoxia affects health, nutrition, work capacity and creates ‘a negative … effect on human reproduction’ (Aldenderfer, 1998, p. 6). Fehren-Schmitz et al. (2017) and Weinstein (2017) recently summarized the physiological constraints associated with high-altitude adaptations, and found a role for ‘both natural selection and developmental plasticity in response to high-altitude hypoxia’ (Weinstein, 2017, p. 14). Physiology certainly plays a role in human adaptation, since raised metabolic rates are required in cold environments (Leonard et al., 2005, p. 456), but there was probably also a need for clothing and other cultural solutions for thermal protection (Gilligan, 2008, p. 42). The evidence is not overwhelming, but at the late Pleistocene open-air camp Quebrada Maní 12 (QM12) (1240 masl), red pigment was found next to wooden sticks that were possibly used for tanning hides for clothing and shoes. Moreover, yarn and other remains of processed plant and animal fibers from late Pleistocene stratigraphic levels of Quebrada Maní 32 open camps may also have been related to the manufacture of apparel (Santoro et al., 2019).
As indicated by Pitblado and Rademaker (2017), physiological adaptations in themselves do not solve all the issues; the overall cost of colonizing the highlands has to be assessed as well. Among the general behavioral adaptations proposed for costly environments, a short effective foraging radius and higher residential mobility have been suggested as ways of reducing survival costs associated with stressful conditions (MacDonald et al., 2009).
In the lowlands, particularly at Pampa del Tamarugal, the location of sites like QM12 (Latorre et al., 2013), and open camps at Quebrada Chipana (a huge lithic quarry and a workshop, Chipana 1 and Chipana 5 respectively), Quebrada Ramaditas (Pampa Ramaditas 5 and Pampa Ramaditas 7), and Quebrada Maní 32 and Quebrada Maní 35, surrounded by an extensive and rich surface archaeological record, indicate the existence of precisely these kinds of relatively short radii, of around 20 and 30 km (Fig. 3). Even when the complex and extensive surface record near QM12 constitutes a good example of what Bailey denominates spatial palimpsests (Bailey, 2007), both the available chronology and the ecological conditions allowing for human occupation reasonably limit the period of temporal overlap of surface and stratigraphic materials within c. 800 years. The existence of at least another three contemporaneous open-air sites within 30 km suggests that QM12, an intermittently occupied site, ‘was part of a complex adaptive system of hunter-gatherers that was successfully taking advantage of the increased ecosystem services available in the Pampa del Tamarugal during the CAPE II’ (Santoro et al., 2017, p. 32; Santoro et al., 2019; Pfeiffer et al., 2018; Workman et al., 2020; Tully et al., 2019). We add that this patchy distribution helped to establish conditions under which the rescue effect—the ability of a system of nodes to compensate for losses in some of the nodes—could operate to maintain the system’s biological viability (Bocquet-Appel & Demars, 2000). Indeed, the concentration of resources tethered to water sources selects for such an interactive mode of exploitation. The functioning of such a system is based on the notion that a set of refugia connected by corridors is more viable than the same set would be without corridors (Simberloff, 1988, p. 477).
A metapopulation approach appears to be particularly suitable for understanding relationships within an ecosystem like Pampa del Tamarugal, with its surface courses and bodies of water forming various basins ‘and enhanced groundwater discharge rates toward the Pacific lowlands’ (Santoro et al., 2017, p. 31). Higher plant biomass at those places created attractive oases for both animals and humans (Quade et al., 2008; Latorre et al., 2013; Gayo et al., 2012; Martínez, 2014, p. 86; Pfeiffer et al., 2018; Workman et al., 2020; Tully et al., 2019). Metapopulation thinking postulates that long-term biological viability depends on the connections between the people of these separated patches. Mutual support works toward maintaining the viability of the human system (Wobst, 1974).
The metapopulation model in which different populations or subpopulations occupy similar niches within a megapatch, and where the change from one megapatch to another (e.g. from the coast to Pampa del Tamarugal) operates by ‘indirect exploration’, can be distinguished from other models. For instance, Aschero (2014) has suggested that people living together may come from different ecologies. In contrast, the metapopulation framework provides a way in which the rescue effect facilitates long-term survival in the different oases, basically because people not only interact genetically but they also shared knowledge regarding the conditions and resources available at each of the nodes (oases). This is an effective way of regionally sharing technological and behavioral innovations that may appear at any given node (Hopkinson et al., 2013). Such occupations are complemented by long-term logistical forays, which may even be related with direct acquisition of goods at distant places (Pintar, 2014), but interaction with similar nodes located near other wetlands was probably a basic requirement for the maintenance of the system. The low number of exotic resources recovered at each of the sites is in line with this suggestion, since they point to mechanisms like visits or other forms of social interaction among individuals from different places rather than systematic exchange (Borrero, 2011a; Borrero, 2011b; Borrero et al., 2011; see also Ortega et al., 2014).
Process and Human Circulation
The classic idea is that coastal habitats may have been populated first, as a result of a Pacific coastal dispersal with subsequent intermittent, perhaps seasonal, expansion into the interior (Osorio, Steele et al., 2017, p.42). Aldenderfer (2008) developed a model of progressive and logistical colonization of the high Andes that resulted in a delayed occupation of the highlands, with ‘a fairly rapid adaptive shift to permanent occupation of Andean high-altitude environments’ (Osorio, Steele, et al., 2017, pp. 43–44; see also Núñez et al., 2002; Grosjean et al., 2005; López Mendoza et al., 2021). In this scheme, coastal ecosystems were subsequently abandoned by at least some sub-populations.
In our study region the ‘earliest coastal settlements are situated both to the north and the south’ of the Atacama coasts (Santoro et al., 2017, p. 32). The evidence of eventual occupations on the Chilean northern coast may be under the oceanic water (Grosjean et al., 2007; Latorre et al., 2011). Alternative entries from the east, on the basis of the evidence from Cueva Bautista (Capriles et al., 2016), or from the south on the basis of the older human occupations recorded in southern Chile (Dillehay et al., 2008, 2015) should also be considered. The relatively late chronology of the human installation in the highlands of Peru–Bolivia–Chile–Argentina certainly allows for these possibilities, relating them with Aldenderfer’s model of gradual advance. However, it differs in the way human circulation is modeled and how occupational nodes are organized. To acknowledge the possibility of the rescue effect is a way of accepting that the process of dispersal is not unidirectional. The changing structure of subsistence resources at different elevations calls for a process of slow human dispersal. This mode of dispersal was probably not new for late Pleistocene hunter-gatherers, who, over generations, gradually became accustomed to drastically different habitats. For this reason, the concept of ‘micro-dispersals’ within a multi-generational framework is relevant here (Nicholson et al., 2021). This scheme, equivalent to our ‘indirect exploration’, is particularly appropriate to high-altitude environments in the Andes, where the occupation of widely separated oases or hotspots (Núñez et al., 2013; Rademaker & Moore, 2019; Yacobaccio, 1994) cannot be seen as a simple process. Back-and-forth processes by the end of the Pleistocene make more sense, since connectivity among nodes within or between megapatches was probably required and effective during colonization, as shown by populations at Pampa del Tamarugal connected with the Pacific coast, the high Andes and the tropical forest (Santoro et al., 2019).
The first human explorers of new lands were probably able to travel relatively narrow paths, and after completing their initial reconnaissance settled only at selected habitats. Anyway, no matter how narrow, ‘pour le chasseur, le territoire n’est plus lignes mais surfaces sur les quelles le gibier se répartit’ [for the hunter, the territory is not formed by lines but surfaces over which prey is distributed] (Collignon, 1996, p. 99). Ethnographically known cases contradict the frequent expectation that hunter-gatherers would move in a basically linear fashion (Gamble, 1999, p. 85). Linear paths are used only for transporting shelters and children. At the same time, hunters move around in wide arcs searching for food or information (cf. Clastres, 1972; Gusinde, 1982), or in movements radiating out from the main camps. This circulation scheme is even observed among the horse-adapted Aonikenk (Musters, 1964 [1971]). This situation produces a relatively good coverage of substantial tracts of land, what Gamble would call ‘surface-area territories’, and is the result of the plurifunctional character of circulation, where both movement and exploitation occur at the same time (Borrero, 2011b, p. 50). It must be stated that the width of the corridors, which undoubtedly varied, always required sufficient surface area to accommodate the hunting territories.
Occupation of the Highlands
The Pampa del Tamarugal, between 900 and 2200 masl, located between the Pacific coast and the foot of the Andes, is crucial to an understanding of human adaptation to arid environments. By the end of the Pleistocene this was not just a transit zone, but one of substantial and diverse ecosystems supporting steady human habitation. Several open camps, located in the proximal and distal sections of the alluvial fan of Quebradas Chipana, Guatacondo and Maní in the Pampa del Tamarugal, provide important information for understanding the habitats of the peopling processes and their relation to other environments (Fig. 1). For instance, human occupation at QM12, dated from 12,750 to 11,530 cal BP, occurred over a period of approximately 800 years, averaging activities at the site and its foraging radius (Joly et al., 2017; Latorre et al., 2013; Santoro et al., 2016; Ugalde et al., 2020). Even a ‘prepared fireplace dug into the hard crust of caliche’ was recorded and the preferential selection of just two species of woody taxa, S. molle and Morella pavonis (syn. M. pavonis), suggests fuel selection, among other strategies, based on prior knowledge (Joly et al., 2017; Santoro et al., 2019; Tully et al., 2019; Workman et al., 2020).
At the Precordillera–Puna, between 2200 and 3500 masl, water availability increased at the end of the Pleistocene. This zone is where the Tuina 1 and Tuina 5 sites are located in the Salt Puna, with the latter presenting one of the few pieces of evidence of megafauna (i.e. Equus spp.) in archaeological context (Núñez et al., 2002). Further, several Salt and Dry Puna sites dated around 10,200–12,600 cal BP, such as Punta Negra, Imilac, Tulan or San Lorenzo (Núñez, 1992; Cartajena et al., 2014; Núñez et al., 2016; Loyola et al., 2017, 2018; Herrera et al., 2019; Sierralta, 2019; de Souza et al., 2022), indicate scattered but regular human settlement (Fig. 1). Evidence at some of these sites indicates seasonal occupations c. 11,700 years cal BP (Núñez & Santoro, 1990; Núñez et al., 2005; Santoro, 1989; Santoro & Núñez, 1987; Santoro et al., 2005). Some of the highland places are included in what is called the Complejo Tuina-Inca Cueva, showing interaction on both sides of the Andes (Núñez et al., 2013, 2016). These places were probably somehow integrated with locations at lower altitudes in the Loa basin or even on the Pacific coast, as indicated by findings of sea shells which are not abundant but quite regularly present. Projectile point styles resembling models from the eastern Dry Puna in North Western Argentina (Hoghin & Oxman, 2015; Santoro et al., 2016, 2019; Herrera et al., 2019) were possibly the consequence of long-distance logistical mobility. Transhumance and other models have been proposed to make sense of this evidence (Núñez & Dillehay, 1979; see also Baied & Wheeler, 1993).
The earliest sites found in the Dry Puna of the Arica and Parinacota Region, located between 3100 and 4500 masl, are Las Cuevas, Hakenasa, Patapatane, Pampa El Muerto 15, Quebrada Blanca and Ipilla 2 (Baied & Wheeler, 1993; Castillo & Sepúlveda, 2017; Herrera et al., 2019; Moreno et al., 2009; Santoro et al., 2016, 2019; Fig. 1). They are all dated to the Pleistocene–Holocene transition (c. 11,500–9500 cal BP). The sites of Hakenasa and Las Cuevas are slightly older than those located below 3500 masl. Cueva Bautista (3933 masl), Bolivia, is even older than other sites at similar altitudes (10,800–10,917 BP; 12,700–12,100 cal BP) (Capriles et al., 2016). All these sites are interpreted as temporary logistical camps (Osorio, Steele, et al., 2017, p. 45; Fig. 1).
On the other side of the Andes, in the Puna of Northwest Argentina, the oldest occupations are recorded between c. 11,200 and 12,300 cal BP at sites between 3200 and 4100 masl. The list of sites includes Inca Cueva 4, Huachichocana 3, Pintoscayoc 1, León Huasi, Cueva Yavi, Peña de las Trampas 1 and Quebrada Seca 3 (Aschero, 1984, 2014; Fernández Distel, 1986, 1989; Hernández-Llosas, 2000; Kulemeyer & Laguna, 1996; Yacobaccio & Madero, 1992; Fig. 1). These sites represent basically ephemeral events (Yacobaccio, 2017), where redundancy in occupation is variable and artefacts are mostly made on local raw material (Osorio, Capriles et al., 2017; Hernández-Llosas, 2005; Martínez, 2014), as would be expected of the initial exploration of the region (Franco, 2002). Evidence of redundancy is present at later occupations on both sides of the Cordillera, as in Quebrada Seca 3, Pintoscayoc or Patapatane, which present relatively continuous Holocene sequences. Occupation at Cueva Bautista was discontinued, and the site was not reoccupied until the late Holocene (Capriles et al., 2016, p. 473; Fig. 1).
Yacobaccio points out that the areas East of the Andes located above 4000 masl in Puna Salada (Fig. 5) ‘could only have been occupied and/or visited seasonally during the summer, [as] suggested by the use of … obsidians from sources like Zapaleri/Laguna Blanca or Caldera Vilama, both above 4000 masl’ (Yacobaccio, 2017, p. 35; Fig. 1), and that the earliest sites are located below 3800 masl, with a ‘range between 12,958 and 12,363 cal. BP, whereas the sites located above 4000 masl have a range of 11,279 to 11,124 cal BP’ (Yacobaccio, 2017, p. 35), concluding that the peopling of the Puna proceeds from the lower environments, and that the earliest occupations ‘are located on the periphery of the Puna, close to the mesothermal valleys and the mountain forest, locally known as yungas’ (Yacobaccio, 2017, p. 35; Fig. 5). The same model was proposed for Puna Salada on the western side of the Andes (Núñez, 2002; Grosjean et al., 2005).
The interpretation of many sites as logistical camps points to the use of a collector strategy, that ‘may reflect a sort of highly dynamic mobility system that allowed people to connect over large regions within the remote high Andes, and thus constituted a mega-ecological patch for these early hunter-gatherers’ (Osorio, Capriles, et al., 2017, p. 10). The existence of these systems was recorded on both sides of the Andes and makes sense in terms of metapopulational thinking. This is in line with the many predictions of Strategy 3 as defined by Osorio, Capriles, et al. (2017, p. 3), an ‘intermittent or opportunistic mobility model’, which is intermediate between seasonal mobility and permanent highland foraging, and basically controlled by environmental variation.
Exotics
Exotic resources are expected at low frequencies during colonization, mostly tools or parts of tools associated with transport by individuals as personal gear. With time, more and more local resources can be incorporated, including those that required some time to be found or recognized. It is the incorporation of crucial but hard to find resources what makes for effective colonization. In a metapopulation scheme, the incorporation of a resource in any part of the system becomes, in a sense, common knowledge. Once the resource is recognized as useful, the movement of individuals between nodes will distribute the new knowledge.
Exotics of various classes were found, for example, shark teeth at early Holocene sites like alero Las Cuevas (second occupation) and Ipilla-2, and fish bones and shells at Patapatane (Osorio, Steele, et al., 2017 p. 49). Small quantities of Pacific seashells (cf. Concholepas, cf. Nassarius gayi, and cf. Argobuccinum rude) have been found at the late Pleistocene sites Chipana 1, Quebrada Maní 12, and Quebrada Maní 35, in Pampa del Tamarugal, while wood from the tropical forest has been found at Pampa Ramaditas 5 (Herrera et al., 2019; Santoro et al., 2019; Fig. 1). Likewise, small quantities of obsidian, brought from sources located in the Andean Cordillera, have been recorded in late Pleistocene sites from the southern coast of Peru (Quebrada Jaguay) and Pampa del Tamarugal (Quebrada Maní 12), in addition to the fragments found in Burial-1 of Acha-2 (c. 9500 years cal BP; Muñoz et al., 1993; Santoro et al., 2016). Some of these goods could be interpreted as belonging outside the economic or technical sphere, even when ‘scraping or cutting’ are plausible uses (Osorio, Capriles, et al., 2017, p. 9).
Strontium isotope signatures on human bones mark limited connections outside the highlands (Standen et al., 2018). Effectively, even when the isotopic values from coastal human individuals confirm that they were born and raised in the littoral zone and that their logistic mobility was concentrated along the coast, at least one individual (dated at c. 6000 BP) recovered at Patapatane in the highlands displays a strontium signal closer to marine values. Moreover, several individuals from the Late Archaic open inland camp Tiliviche, although raised in the interior, ‘were part of a broad logistic mobility pattern connected with the coast’ (Standen et al., 2018, p. 162).
Evidence of exotics—like the presence of feathers, or canes used as dart and spear shafts—is, as one would expect, more abundant in sites located East of the Andes. Also noteworthy is the presence of Strophocheilus or cervid bone at sites such as Inca Cueva or Quebrada Seca 3 (Aschero, 2014). The provenience of most of these items can be found in the eastern Yungas. On the other hand, archaeological evidence in the lower valleys of the east is not abundant, but increasingly important. At Taller Puesto Viejo 1, at El Infiernillo (3000 masl), a site that goes back to about 8300 cal BP (Fig. 1), obsidian artefacts from Puna sources were found (Martínez et al., 2013; Oliszewski et al., 2017), including at least one projectile point made of Puna vulcanite (Martínez et al., 2013; Martínez, 2014, p. 71).
Obsidian is abundant at both Puna cave sites, Hakenasa and Las Cuevas, probably transported from relatively close sources like Charaña and Parinacota, while at the younger site Pampa El Muerto it is exotic but not abundant (Osorio, Steele, et al., 2017, p. 45–46). Obsidian is also relatively abundant at Cueva Bautista, and geochemically corresponds to the Cerro Kaskio outcrop located some 15 km from the site (Capriles et al., 2016, p. 470), so that it can be considered local (Fig. 1). The presence of obsidian from other sources at several sites certainly indicates high mobility and long-distance interaction (Yacobaccio, 2017). Obsidian at early Holocene sites like Quebrada Seca 3 or Salamanca in Argentina suggested to Pintar et al. (2016) that mobility patterns during the early colonization phase (early Holocene) of the region east of the Andes included two obsidian sources, while later obsidian artifacts show the use of multiple sources ‘within a range of 40–95 km, revealing a process of landscape learning’ (Pintar, 2014, p. 16). Pintar suggests a model of land use by logistical mobility that takes advantage of adjacent ecological areas through direct access to resources—for example, direct acquisition of obsidian and wooden canes, which were used for personal gear (Pintar, 2008, p. 53; 2014, p. 17).
All this evidence indicates that highlanders had access to—and knew how to obtain—resources from coastal and lowland ecosystems. They probably interacted with other groups in a variety of configurations. The low-frequency circulation of these non-local goods, however, suggests interactions with a low level of transfer of objects between distant nodes, and would therefore indicate informal systems of exchange or movement of small human groups.
Megafauna
The paleoecological record of Atacama clearly shows that the region was attractive to humans at the end of the Pleistocene (Latorre et al., 2013; Fig. 3). Further, the fact that hunter-gatherers were preying on camelids and small pack animals like Lagidium sp. or Chinchilla sp. at sites like Inca Cueva 4, Hornillos 2 or Hakenasa (Osorio, Capriles, et al., 2017; Yacobaccio, 2017) shows that these resources were well known to Andean peoples. This knowledge probably facilitated the settlement of the highlands during the early Holocene. Pintoscayoc, for instance, is a cave site that shows an equilibrated proportion of large and small mammals, the latter including rock cavy (Galea sp.), Lagidium, Ctenomys, and Chinchilla (Elkin & Rosenfeld, 2001; Hernández-Llosas, 2000, 2005). Rodents as well as locusts were recovered at Quebrada Seca 3 (Mondini & Elkin, 2006). In other words, beyond hunting vicuñas and guanacos, for which no innovations were required (Fig. 5), the regular consumption of small pack mammals was systematically added. Their capture and consumption surely required a different kind of organizational effort (Lupo & Schmitt, 2002; Fig. 1).
It is notable that the populations of these regions apparently did not regularly consume megafauna, and in some cases even medium-sized mammals were not important at the site scale. The species recorded at most of the sites along the Andes are camelids and deer, with the occasional spectacled bear or the small pack mammals mentioned above (Osorio, Capriles, et al., 2017; Yacobaccio, 2017; Lynch & Pollock, 1981; Elkin & Rosenfeld, 2001; Mondini & Elkin, 2006; Osorio, Capriles, et al., 2017; Yacobaccio, 2017; Rademaker & Moore, 2019). It should also be noted that taphonomic analysis is lacking for many of these sites, making the lists of faunal-consumption remains tentative at best. In any case, the evidence indicates that mega mammals were not a basic food in the Andean world of the early explorers, although our knowledge of the availability of megafauna is desperately limited, a situation that is slowly changing. According to the information from both Chile and Argentina, exploitation of mega mammals is almost absent, and this extends to the Andean world both north and south of Atacama. There is evidence for the availability of megafauna, such as Hippidion, Megatherium and Paleolama, before and even contemporaneously with human presence (Caro, 2019; López Mendoza & Rojas Mondaca, 2018; Tamayo & Frassinetti, 1980). Examples are the Hippidion saldiasi found at Calama, dated 25,380–25,680 cal BP (Alberdi et al., 2007), and at Salar de Surire, dated 13,300–13,060 cal BP (Labarca et al., 2021); the ground sloth recorded in El Loa (Salinas, Naranjo & Marshall, 1991); and the bones and feces of Hippidion, Megatheriinae and Mylodontinae recovered at Peña de las Trampas 1 (3580 masl) in Argentina, dated 22,989–23,847 and 14,187–15,047 cal BP. Carnivore feces were also recovered at the latter site (Martínez et al., 2010; Martínez, 2014, p. 77). At Barro Negro (3820 m), Argentina, Fernández et al. (1991) identified the presence of Hippidion bones dated between c. 13,000 and 10,000 cal BP. Recent work by Yacobaccio and Morales (2011) in the same locality identified older presences for Hippidion, extending the temporal range to 14,133–15,172 cal BP, still without any association with humans. Even when some dates at Peña de las Trampas and Barro Negro overlap with the presence of humans in the highlands, no behavioral association was identified.
At Cueva Cacao IA (3720 masl), Argentina, a Glossotherium rib ‘with possible cutmarks in association with five obsidian and dacite artefacts’ was recovered by Aschero (Pintar, 2014, p. 13). A recent claim for human association with megafauna c. 39,000–42,000 14C BP at Cacao IA (Aschero, 2017) remains unpublished. Human occupations at Cueva Bautista, Bolivia, show that most of the artiodactyl remains correspond to vicuña and guanaco. The possibility remains open, however, that ‘extinct taxa such as gracile llama (Lama gracilis), American horses (Hippidium saldiasi), macrauchenia (Macrauchenia patachonica), and xenarthrans’ (Capriles et al., 2016, p. 470), were seen and eventually hunted or even scavenged by late Pleistocene hunter-gatherers. Villavicencio et al. (2019) have shown certain contemporaneity of human and megafauna, most of which later became extinct. A possible sloth coprolite was also identified above the human occupation surface. The only bona fide evidence for human interaction was found at Tuina 5, where a single horse bone was recovered (Núñez et al., 2002).
In sum, the early record for humans in the highlands on both sides of the Andes does not show any important interaction with Pleistocene mega mammals. The chronology of the disappearance of this fauna from the Puna is not well known, but there is clear evidence for the existence of mega mammals before the arrival of humans (Labarca et al., 2021; Martínez, 2014), with perhaps some temporal overlap (Yacobaccio & Morales, 2011). During the Pleistocene/Holocene transition several Andean habitats were attractive to humans, including Pampa del Tamarugal and the Precordillera–Puna ecosystems (Capriles et al., 2016; Osorio, Capriles, et al., 2017; Rademaker & Moore, 2019). However, the evidence for human colonization of the upper highlands is not abundant and only weak signals of interaction with megafauna exist. In contrast, interaction with modern fauna is unequivocal. Taken together, the evidence indicates a delayed process of occupation of the high Andes in comparison with other regions of South America (Politis et al., 2016).
Early Holocene Divergence
At the beginning of the Holocene there was a sudden change in the general distribution of the occupational nodes in the highlands between 900 and > 3500 masl. The imposition of ecological barriers transformed a previously interconnected distribution of nodes across the Andean ecosystems into a series of disconnected dots. In biogeographical terms they can be considered as cases of vicariance. Divergent cultural trajectories either side of the Andes were the basic result, despite the fact that people maintained some communication and sociocultural relationship (Santoro et al., 2016; Yacobaccio, 2017; de Souza et al., 2022). Likewise, the long-term sociocultural processes along the coast were locally distinct, with strong attachment to the hyper-arid habitats of the Pacific littoral in social and genetic terms, despite the fact that connections with inland groups never ceased. Indeed, these became more active and dynamic towards the end of the pre-European epoch, as interregional traffic of goods intensified (Berenguer & Pimentel, 2017; Carter, 2016; Castro et al., 2016; Gallardo, 2013; Horta, 2015; Knudson et al., 2012; Núñez & Briones, 2022; Núñez et al., 2010; Pimentel et al., 2017; Santoro et al., 2020).
The paleoecological history of the Pampa del Tamarugal clearly points to a drastic decrease in the availability of water after 10,000–9500 14C BP; this became chronic, generating ‘an absence of intermittent or even perennial surface discharge and extremely reduced bioproductivity’ (Santoro et al., 2017, p. 35). Under these conditions it was difficult to maintain a permanent hunter-gatherer way of life in the desert; sites were abandoned early in the Holocene, and people probably migrated to the coastal valleys and to the east towards the ‘sierra’ and Altiplano highlands.
Similar conditions have been recorded in the southern Atacama Desert (Núñez et al., 2013; de Souza et al., 2021). In other words, after seven to six thousand years the availability of fresh water collapsed, ending the effective process of human colonization of the Intermediate Depression, the core of the Atacama Desert. Populations that remained in the highlands were faced with the choice of reversing their eastward movement and returning to the coastal areas, or, alternatively, adapting in completely new ways ‘near the few and highly localized sources of permanent water’, like ‘the quebradas of Sapiga … and Tiliviche, where the sites of Aragón and Tiliviche 1 are situated respectively’ (Santoro et al., 2017, p. 33). The occupation of these lower nodes closer to the coast began c. 9500 cal BP and was relatively continuous throughout the Holocene, with an economic dependence on the resources of the Pacific coast, where specialized marine hunter-gatherers developed costly adaptations (like the Chinchorro tradition) (Arriaza et al., 2005; Pestle et al., 2015; Roberts et al., 2013; Standen et al., 2014, 2018). Craniometric data and genetic mtDNA indicated that the skeletal material from Acha (9500 cal BP), at the beginning of the Chinchorro Tradition, is related to an early coastal migration (Manríquez et al., 2011; Rothhammer, 2014; Rothhammer et al., 2009; Varela et al., 2004, 2008). In contrast, skeletal remains from the Camarones 14 site, as well as stable isotope signals, indicate closer coastal-population interaction with the highlands (Santoro et al., 2012, p. 640; Rothhammer, 2014; Standen et al., 2018).
The question of ‘how the Chinchorro carried on with their way of life isolated at geographically restricted perennial river mouths with fresh water along the Atacama Desert in the Pacific coast of South America’ (Santoro et al., 2012, p. 637) can also be approached via the concept of metapopulation. One indicator of connections between coastal valleys may be the high arsenic levels recorded in hair from Chinchorro mummies from the Morro site in Arica, ‘perhaps reflecting mobility and interaction among communities in arsenic rich areas (i.e. Camarones Valley)’ (Byrne et al., 2010, p. 34), which would clearly imply the existence of corridors, in this case along the coast. An analysis of inter-valley connectivity between the different coastal nodes shows that it diminishes with time (Santoro et al., 2012, p. 645), which can be understood as a reflection of an increase in the operating costs of the rescue effect through time. This helps to explain their ‘disintegration’ at the end of the Holocene. These costs can perhaps be measured in terms of the high number of pathologies that resulted from increasing isolation. Chinchorro was a costly adaptation to start with, as shown by their resilience in an area characterized by toxic levels of arsenic in the water (Arriaza, 2005; Byrne et al., 2010; Swift et al., 2015). For example, the highest arsenic levels in the Americas (> 1000 mg/L) are recorded at Quebrada Camarones (Apata et al., 2017). A high percentage of chronic arsenic poisoning, probably from drinking water, was identified in a 3000-year sequence (Swift et al., 2015). Despite all the negative effects of arseniasis, the Camarones people persisted in this adverse environment for some seven thousand years. Indeed, mummification as an emotional response to perceived adversity can be seen as an important component of that adjustment (Arriaza, 2005).
Conclusions
The pioneer occupations of the highlands as ‘part of an emergent Andean human mobility system that connected adjacent ecological environments spanning across different elevations’ (Osorio, Capriles, et al., 2017, p. 10), can be understood using metapopulation theory. Early occupations are found in patches where drinking water and other resources are concentrated (Rademaker & Moore, 2019; Yacobaccio, 1994; Yacobaccio et al., 2017). Corridors can be seen as a basic component of the environment during the relatively short late Pleistocene bonanza intervals, ensuring the connectivity required to provide security through the rescue effect, throughout vast territories of South America, and despite very low population density. Then, when critical demographic, technical or subsistence resources failed, the network of corridors and nodes facilitated the maintenance of a steady occupation within high Andean ecosystems. The rescue effect is not measured by the circulation of exotics, but by the circulation of the basic knowledge of technologies and managing subsistence resources.
The distribution of the oldest sites on either side of the Andean Cordillera is slightly different, but it can be plausibly argued that c. 11,000 cal. BP people were selectively using all well-known habitats along and across the Andes. We suggest that these patterns could be the result of gradual processes of highland colonization, and, to a large extent, are the result of the ability of relatively isolated groups of people to support each other through the rescue effect. The circulation of goods or genes helped to keep the metapopulation biologically viable as they adapted to the high-altitude environments (Jodry & Santoro, 2017).
Neither the highland nor the coastal adaptations show perfect adjustment to the environment; rather, they are examples of the high costs associated with making a living at places where risk and uncertainty are frequent. As such, they are a powerful demonstration of how resilient human populations can be. In the case of people from the Atacama Desert, this meant that they coped with critical periods of potential collapse caused by shortage of water and the concomitant reduction in bioproductivity. In the end, the discontinuous use of resource patches across ecosystems resulted in several local sub-populations distributed over these fractured environments from the Pacific coast to the highlands, ensuring the long-term viability of the colonization process, the remains and traces of which can still be seen in (to paraphrase Luis Advis) the arid silence of the pampa free of miracle.
References
Alberdi, M. T., Prado, J. L., López, P., Labarca, R., & Martínez, I. (2007). Hippidion saldiasi Roth, 1899 (Mammalia, Perissodactyla) en el Pleistoceno tardío de Calama, norte de Chile. Revista Chilena De Historia Natural, 80(2), 157–171.
Aldenderfer, M. S. (1998). Montane foragers: Asana and the South Central Andean Archaic. University of Iowa Press.
Aldenderfer, M. S. (2008). High elevation foraging societies. In H. Silverman & W. H. Isbell (Eds.), Handbook of South American archaeology (pp. 131–144). Springer.
Apata, M., Arriaza, B. T., Llop, E., & Moraga, M. (2017). Human adaptation to arsenic in Andean populations of the Atacama Desert. American Journal of Physical Anthropology, 163(1), 192–199.
Arriaza, B. T. (2005). Arseniasis as an environmental hypothetical explanation for the origin of the oldest artificial mummification practice in the world. Chungara Revista de Antropología Chilena, 37(2), 255–260.
Arriaza, B. T., Doubrava, M., Standen, V. G., & Haas, H. (2005). Differential mortuary treatment among the Andean Chinchorro fishers: Social inequalities or in situ regional cultural evolution? Current Anthropology, 46(4), 662–671.
Aschero, C. A. (1984). El sitio ICC-4: Un asentamiento precerámico en la quebrada de Inca Cueva. Estudios Atacameños. Arqueología y Antropología Surandinas, 7, 67–72.
Aschero, C. A. (2014). Hunter-gatherers of the Puna in a temporal perspective (10,500–3500 BP): The case of Antofagasta de la Sierra (Catamarca, Argentina). In E. Pintar (Ed.), Hunter-gatherers from a high-elevation desert: People of the Salt Puna (Northwest Argentina). BAR, International Series 2641 (pp. 27–44). Archaeopress.
Aschero, C. A. (2017). Cacao 1: Lithic evidence and mobility ranges during the Pleistocene in the Atacama Puna (Antofagasta de la Sierra, Catamarca, Argentina). 11th International Symposium on Knappable Materials, ‘From toolstone to stone tools’, p. 116. Buenos Aires: IMHICIHU-CONICET.
Baied, C., & Wheeler, J. (1993). Evolution of high Andean puna ecosystems: Environment, climate, and culture change over the last 12.000 years in the central Andes. Mountain Research and Development, 13, 145–156.
Bailey, G. (2007). Time perspectives, palimpsests and the archaeology of time. Journal of Anthropological Archaeology, 26, 198–223.
Beaton, J. M. (1991). Colonizing continents: Some problems from Australia and the Americas. In T. D. Dillehay & D. J. Meltzer (Eds.), The first Americans: Search and research (pp. 209–230). CRC Press.
Berenguer, J., & Pimentel, G. (2017). Introducción al estudio de los espacios internodales y su aporte a la historia, naturaleza y dinámica de las ocupaciones humanas en zonas áridas. Estudios Atacameños Arqueología y Antropología Surandinas, 56, 3–11.
Binford, L. R. (1982). The archaeology of place. Journal of Anthropological Archaeology, 1, 5–31.
Bocquet-Appel, J.-P., & Demars, P.-Y. (2000). Population kinetics in the Upper Palaeolithic in Western Europe. Journal of Archaeological Science, 27, 551–570.
Borrero, L. A. (1989–1990). Evolución cultural divergente en la Patagonia Austral. Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 19, 133–140.
Borrero, L. A. (2011a). The theory of evolution, other theories, and the process of human colonization of America. Evolution and Education Outreach, 4(2), 218–222.
Borrero, L. A. (2011b). La arqueología de cazadores-recolectores: Ambiente y conocimiento. Revista De Cazadores-Recolectores, 4, 43–58.
Borrero, L. A., Martin, F. M., & Barberena, R. (2011). Visits, ‘Fuegians,’ and information networks. In R. Whallon, W. A. Lovis, & R. Hitchcock (Eds.), Information and its role in hunter-gatherer bands (pp. 249–265). Los Angeles: The Cotsen Institute of Archaeology at UCLA.
Byrne, S., Amarasiriwardena, D., Bandak, B., Bartkus, L., Kane, J., Jones, J., et al. (2010). Were Chinchorros exposed to arsenic? Arsenic determination in Chinchorro mummies’ hair by laser ablation inductively coupled plasma-mass spectrometry (LA-ICP-MS). Microchemical Journal, 94, 28–35.
Capriles, J. M., Albarracin-Jordan, J., Lombardo, U., Osorio, D., Herrera, K. A., Maley, B., et al. (2016). High-altitude adaptation and late Pleistocene foraging in the Bolivian Andes. Journal of Archaeological Science, Report, 6, 463–474.
Capriles, J. M., Lombardo, U., Maley, M., Zuna, C., Veit, H., & Kennett, D. J. (2019) Persistent Early to Middle Holocene tropical foraging in southwestern Amazonia. Science Advances, 5(4), eaav5449.
Caro, F. (2019). Registro de Megatherium sp. (Mammalia, Xenarthra, Megatheriidae) en Quebrada Maní 35, un sitio arqueológico del Pleistoceno tardío en la Pampa del Tamarugal, Chile. Memoria para optar al título de arqueólogo, Departamento de Antropología, Universidad de Tarapacá, Arica.
Cartajena, I., Loyola, R., Núñez, L., & Faúndez, W. (2014). Problemas y perspectivas en la interpretación del registro espacial de Punta Negra e Imilac. In F. Falabella, L. Sanhueza, L. E. Cornejo, & I. Correa (Eds.), Distribución espacial en sociedades no aldeanas: del registro arqueológico a la interpretación social (pp. 143–162). Santiago: Sociedad Chilena de Arqueología.
Carter, C. P. (2016). The economy of prehistoric coastal northern Chile: Case study: Caleta Vitor. The Australian National University, Canberra.
Cashdan, E. (1990). Risk and uncertainty in tribal and peasant economies. Boulder: Westview.
Castillo, C., & Sepúlveda, M. (2017). Objetos ‘misceláneos’ y dinámicas sociales en contextos cazadores recolectores de la precordillera de Arica, extremo norte de Chile. Chungara Revista de Antropología Chilena, 49(2), 159–174.
Castro, V., Berenguer, J., Gallardo, F., Llagostera, A. & Salazar, D. (2016). Vertiente occidental circumpuneña: Desde las sociedades post arcaicas hasta las preincas (ca. 1500 a.C.– 1470 d.C.) In C. Aldunate, F. Falabella, L. Sanhueza, J. Hidalgo, & M. Uribe (Eds.), Prehistoria en Chile desde sus primeros habitantes hasta los Incas (pp. 239–283). Santiago: Editorial Universitaria.
Clastres, P. (1972). Chronique des Indiens Guayaki. Libraire Plon.
Collignon, B. (1996). Les Inuit : Ce qu’ils savent du territoire. L’Harmattan.
de Souza, P., Cartajena, I., Riquelme, R., Maldonado, A., De Porras, E., Santander, B., et al. (2021). Late Pleistocene–early Holocene human settlement and environmental dynamics in the southern Atacama Desert highlands (24.0°S–24.5°S, northern Chile). Geoarchaeology, An International Journal. Doi: https://doi.org/10.1002/gea.2184.
Dillehay, T. D., Ocampo, C., Saavedra, J., Oliveira Sawakuchi, A., Vega, R. M., Pino, M., et al. (2015). New archaeological evidence for an early human presence at Monte Verde, Chile. Plos ONE, 10(11), e0141923.
Dillehay, T. D., Ramírez, C., Pino, M., Collins, M. B., Rossen, J., & Pino-Navarro, J. (2008). Monte Verde: Seaweed, food, medicine, and the peopling of South America. Science, 320, 784–786.
Elkin, D. C., & Rosenfeld, S. A. (2001). Análisis faunístico de Pintoscayoc 1 (Jujuy). In G. Mengoni Goñalons, D. Olivera, & H. Yacobaccio (Eds.), El uso de los camélidos a través del tiempo (pp. 29–64). Buenos Aires: Ediciones del Tridente.
Fehren-Schmitz, L., Harkins, K. M., & Llamas, B. (2017). A paleogenetic perspective on the early population history of the high-altitude Andes. Quaternary International, 15, 25–33.
Fernández, J., Markgraf, V., Panarello, H. O., Albero, M., Angiolini, F. E., Valencio, S., & Arriaga, M. (1991). Late Pleistocene/Early Holocene environments and climates, fauna, and human occupation in the Argentine Altiplano. Geoarchaeology, an International Journal, 6(3), 251–272.
Fernández Distel, A. A. (1986). Las cuevas de Huachichocana, su posición dentro del precerámico con agricultura incipiente del noroeste Argentino. Beiträge Zur Allg. Vgl. Archäologie, 8, 353–430.
Fernández Distel, A. A. (1989). Una nueva cueva con maíz acerámico en N.O. argentino: León Huasi 1, excavación. Revista Comunidades Científicas, 1(1), 4–17.
Franco, N. V. (2002). ¿Es posible diferenciar los conjuntos líticos atribuidos a la exploración de un espacio de los correspondientes a otras etapas del poblamiento? El caso del extremo sur de Patagonia. Werken, 3, 119–132.
Gallardo, F. (2013). Sobre el comercio y mercado tradicional entre los lupaca del siglo XVI: Un enfoque económico sustantivo. Chungara Revista de Antropología Chilena, 45(4), 599–612.
Gamble, C. (1999). The Palaeolithic societies of Europe. Cambridge University Press.
Gayo, E. M., Latorre, C., Jordan, T. E., Nester, P. L., Estay, S., Ojeda, K. F., & Santoro, C. M. (2012). Late Quaternary hydrological and ecological changes in the hyperarid core of the northern Atacama Desert (21° S). Earth Science Review, 113, 120–140.
Gilligan, I. (2008). Clothing and climate in aboriginal Australia. Current Anthropology, 49(3), 487–495.
Goldberg, A., Mychajliw, A. M., & Hadly, E. H. (2016). Post-invasion demography of prehistoric humans in South America. Nature, 532, 232–235.
Grosjean, M., Núñez, L., & Cartajena, I. (2005). Palaeoindian occupation of the Atacama Desert, northern Chile. Journal of Quaternary Science, 20(7–8), 643–653.
Grosjean, M., Santoro, C. M., Thompson, L., Núñez, L., & Standen, V. G. (2007). Mid-Holocene climate and culture change in the south-central Andes. In D. G. Anderson, K. A. Maasch, & D. H. Sandweiss (Eds.), Climate change and cultural dynamics: A global perspective on mid-Holocene transitions (pp. 51–115). Elsevier.
Grove, M. (2009). Hunter-gatherer movement patterns: Causes and constraints. Journal of Anthropological Archaeology, 28, 222–233.
Gusinde, M. (1982). Los indios de Tierra del Fuego. I. Los Selknam. Buenos Aires: CONICET-CAEA.
Hernández-Llosas, M. I. (2000). Quebradas altas de Humahuaca a través del tiempo: El caso Pintoscayoc. Estudios Sociales del NOA, 4(2), 167–224.
Hernández-Llosas, M. I. (2005). Pintoscayoc and the archaeology of the arid Humahuaca rift valley, north-western Argentina. In M. Smith & P. Hesse (Eds.), 23° south, archaeology and environmental history of the southern deserts (pp. 186–198). National Museum of Australia Press.
Herrera, K. A., Pelegrin, J., Gayo, E. M., & Santoro, C. M. (2019) Circulation of objects and raw material in the Atacama Desert, northern Chile by the end of the Pleistocene. PaleoAmerica. A Journal of Early Human Migration and Dispersal, 5(4), 335–348.
Hiscock, P. (2008). Archaeology of ancient Australia. Routledge.
Hiscock, P. (2013). Occupying new lands: Global migrations and cultural diversification with particular reference to Australia. In K. E. Graf, C. V. Ketron, & M. R. Waters (Eds.), Paleoamerican odyssey (pp. 3–11). Center for the Study of the First Americans, Texas A&M University.
Hoguin, R., & Oxman, B. (2015). Palaeoenvironmental scenarios and lithic technology of the first human occupations in the Argentine Dry Puna. Quaternary International, 330, 171–182.
Hopkinson, T., Nowell, A., & White, M. (2013). Life histories, metapopulation ecology, and innovation in the Acheulian. PaleoAnthropology, 2013, 61–76.
Horta, H. (2015). El señorío Arica y los reinos altiplánicos. Quillqa, Universidad Católica del Norte.
Jochim, M. (1976). Hunter-gatherer subsistence and settlement: A predictive model. Academic Press.
Jodry, M. A., & Santoro, C. M. (2017). Walking closer to the sky: High-altitude landscapes and the peopling of the New World. Quaternary International, 461, 102–107.
Joly, D., Santoro, C. M., Gayo, E. M., Ugalde, P. C., March, R. J., Carmona, R., et al. (2017). Fuel management and human colonization of the Atacama Desert, northern Chile, during the Pleistocene–Holocene transition. Latin American Antiquity, 28(1), 144–160.
Kelly, R. L. (1999). Hunter-gatherer foraging and colonization of the western hemisphere. L’anthropologie, 37(1), 143–153.
Kelly, R. L. (2003). Colonization of new land by hunters-gatherers: Expectations and implications based on ethnographic data. In M. Rockman & J. Steele (Eds.), Colonization of unfamiliar landscapes: The archaeology of adaptation (pp. 44–57). Routledge.
Kilby, J. D. (2014). Direction and distance in Clovis caching: The movement of people and lithic raw materials on the Clovis-age landscape. In B. B. Huckell & J. D. Kilby (Eds.), Clovis caches: Recent discoveries and new research (pp. 201–216). University of New Mexico Press.
Knudson, K. J., Pestle, W. J., Torres-Rouff, C., & Pimentel, G. (2012). Assessing the life history of an Andean traveler through biogeochemistry: Stable and radiogenic isotope analyses of archaeological human remains from northern Chile. International Journal of Osteoarchaeology, 22, 435–451.
Kulemeyer, J. A., & Laguna, L. R. (1996). La cueva de Yavi: Cazadores-recolectores del borde oriental de la puna de Jujuy (Argentina) entre los 12.500 y 8.000 años 14C B.P. Ciencia y Tecnología, 1, 37–46.
Kulemeyer, J. J., & López, F. L. (1997). Reconocimiento de las materias primas líticas del sitio arqueológico La Cueva de Yavi (Jujuy). Estudios Atacameños. Arqueología y Antropología Surandinas, 12, 15–24.
Labarca, R., Caro, F. J., Villavicencio, N. A., Capriles, J. M., Briones, E., Latorre, C., et al. (2021). A partially complete skeleton of Hippidion saldiasi Roth, 1899 (Mammalia: Perissodactyla) from the late Pleistocene of the high Andes in northern Chile. Journal of Vertebrate Paleontology. https://doi.org/10.1080/02724634.2020.1862132
Latorre, C., González, A. L., Quade, J., Fariña, J. M., Pinto, R., & Marquet, P. A. (2011). Establishment and formation of fog-dependent Tillandsia landbeckii dunes in the Atacama Desert: Evidence from radiocarbon and stable isotopes. Journal of Geophysical Research, 116, G03033.
Latorre, C., Santoro, C. M., Ugalde, P. C., Gayo, E. M., Osorio, D., Salas-Egaña, C., et al. (2013). Late Pleistocene human occupation of the hyperarid core in the Atacama Desert, northern Chile. Quaternary Science Reviews, 77, 19–30.
Leonard, W. R., Snodgrass, J. J., & Sorensen, M. V. (2005). Metabolic adaptation in indigenous Siberian populations. Annual Review of Anthropology, 34, 451–471.
López Mendoza, P., Carrasco González, C., Loyola Muñoz, R., Flores-Aqueveque, V., Santana-Sagredo, F., Maldonado Castro, A., et al. (2021). Develando terra incognita: Una búsqueda arqueológica de las primeras ocupaciones humanas en los salares de Infieles y Pedernales (3000–4100 msnm, 25–26 S), Región de Atacama, Chile. Intersecciones en Antropología, 22(1), 11–23.
López Mendoza, P., & Rojas Mondaca, O. (2018). El Pleistoceno de la cuenca de Calama: Evidencias paleontológicas y arqueológicas. Calama: Monografías del Museo de Historia Natural y Cultural del Desierto de Atacama.
Lovis, W. A., Donahue, R. E., & Holman, M. B. (2005). Long-distance logistic mobility as an organizing principle among northern hunter-gatherers: A great lakes middle Holocene settlement system. American Antiquity, 70(4), 669–693.
Loyola, R., Núñez, L., Aschero, C., & Cartajena, I. (2017). Tecnología lítica del Pleistoceno final y la colonización del Salar de Punta Negra (24,5° S), Desierto de Atacama. Estudios Atacameños. Arqueología y Antropología Surandinas, 55, 5–34.
Loyola, R., Cartajena, I., Núñez, L., & López, M. P. (2018). Moving into an arid landscape: Lithic technologies of the Pleistocene–Holocene transition in the high-altitude basins of Imilac and Punta Negra, Atacama Desert. Quaternary International, 473, 206–224.
Lupo, K. D., & Schmitt, D. N. (2002). Upper Paleolithic net-hunting, small prey exploitation, and women’s work effort: A view from the ethnographic and ethnoarchaeological record of the Congo basin. Journal of Archaeological Method and Theory, 9(2), 147–179.
Lynch, T. F., & Pollock, S. (1981). Chobshi cave and its place in Andean and Ecuadorian archaeology. In L. B. Harten, C. N. Warren, & D. R. Tuohy (Eds.), Anthropological papers in memory of Earl H. Swanson (pp. 19–40). Idaho State Museum of Natural History.
MacDonald, K., Roebroeks, W., & Verpoorte, A. (2009). An energetics perspective on the Neandertal record. In J.-J. Hublin & M. P. Richards (Eds.), The evolution of hominin diets (pp. 211–220). Springer.
Manríquez, G., Moraga, M., Santoro, C. M., Aspillaga, E., Arriaza, B. T., & Rothhammer, F. (2011). Morphometric and mtDNA analyses of archaic skeletal remains from southwestern South America. Chungara Revista de Antropología Chilena, 43(2), 283–292.
Martínez, J. G. (2014). Contributions to the knowledge of natural history and archaeology of hunter-gatherers of Antofagasta de la Sierra (Argentine south puna): The case of Peñas de las Trampas 1.1. In E. Pintar (Ed.), Hunter-gatherers from a high-elevation desert: People of the salt puna (northwest Argentina). BAR International Series, 2641 (pp. 71–93). Archaeopress.
Martínez, J. G., Mauri, E., Mercuri, C., Caria, M., & Oliszewski, N. (2013). Mid-Holocene human occupations in Tucumán (Northwest of Argentina). Quaternary International, 307, 86–95.
Martinez, J. G., Powell, J. E., & Rodríguez, M. F. (2010). Dung analysis and its correlation with three different species of extinct megafauna in the southern Argentinian puna. Current Research in the Pleistocene, 27, 176–179.
Meltzer, D. J., Todd, L. C., & Holliday, V. T. (2002). The Folsom (Paleoindian) type site: Past investigations, current studies. American Antiquity, 67(1), 5–36.
Mondini, M., & Elkin, D. (2006). Cazadores-recolectores de la cuenca de Antofagasta de la Sierra (Puna Meridional Argentina): Una perspectiva zooarqueológica y tafonómica. Cazadores y recolectores del cono sur. Revista De Arqueología, 1, 67–79.
Moreno, A., Santoro, C. M., & Latorre, C. (2009). Climate change and human occupation in the northernmost Chilean Altiplano over the last ca. 11500 cal BP. Journal of Quaternary Science, 24, 373–382.
Muñoz, I., Arriaza, B. T., & Aufderheide, A. C. (Eds.) (1993). Acha-2 y los orígenes del poblamiento humano en Arica. Universidad de Tarapacá.
Musters, G. Ch. (1964 (1871). Vida entre los Patagones. Buenos Aires: Solar/Hachette.
Nicholson, S. L., Hosfield, R., Groucutt, H. S., Pike, A. W. G., & Fleitmann, D. (2021). Beyond arrows on a map: The dynamics of Homo sapiens dispersal and occupation of Arabia during Marine Isotope Stage 5. Journal of Anthropological Archaeology, 62, 101269.
Núñez, L. (1992). Ocupación arcaica en la Puna de Atacama: Secuencia, movilidad y cambio. In B. J. Meggers (Ed.), Prehistoria sudamericana: Nuevas perspectivas (pp. 283–307). Washington D.C.: Taraxacum.
Núñez, L., & Briones, L. (2022). Intersite locations of prehistoric caravan traffic in the core of the Atacama Desert, northern Chile. In P. B. Clarkson & C. M. Santoro (Eds.), Caravans in socio-cultural perspective: Past and present (pp. 212–235). Routledge.
Núñez, L., Cartajena, I., & Grosjean, M. (2013). Archaeological silence and ecorefuges: Arid events in the Puna of Atacama during the middle Holocene. Quaternary International, 307, 5–13.
Núñez, L., & Dillehay, T. D. (1979). Movilidad giratoria, armonía social y desarrollo en los Andes meridionales: Patrones de tráfico e interacción económica. Universidad del Norte.
Núñez, L., Grosjean, M., & Cartajena, I. (2002). Human occupations and climate change in the puna de Atacama, Chile. Science, 298, 821–824.
Núñez, L., Grosjean, M., & Cartajena, I. (2005). Ocupaciones humanas y paleoambientes en la puna de Atacama. Instituto de Investigaciones Arqueológicas y Museo, Universidad Católica del Norte, Taraxacum.
Núñez, L., Grosjean, M., & Cartajena, I. (2010). Sequential analysis of human occupation patterns and resource use in the Atacama Desert. Chungara Revista de Antropología Chilena, 42(2), 363–391.
Núñez, L., Jackson, D., Dillehay, T. D., Santoro, C. M., & Méndez, C. (2016). Cazadores-recolectores tempranos y los primeros poblamientos en Chile hacia finales del Pleistoceno (ca.13.000–10.000 años antes del presente). In F. Falabella, M. Uribe, L. Sanhueza, C. Aldunate & J. Hidalgo (Eds.), Prehistoria en Chile desde sus primeros habitantes hasta los Incas (pp. 71–116). Santiago: Editorial Universitaria, and Sociedad Chilena de Arqueología.
Núñez, L., & Santoro, C. M. (1990). Primeros poblamientos en el cono sur de América (XII–IX milenio A.P.). Revista de Arqueología Americana, 1, 91–139.
Oliszewski, N., Martínez, J. G., Arreguez, G. A., Gramajo Bühler, C. M., & Naharro, M. E. (2017). La transición vista desde los valles intermontanos del noroeste argentino: Nuevos datos de la quebrada de los Corrales (El Infiernillo, Tucumán, Argentina). Chungara Revista de Antropología Chilena, 50(1), 71–86.
Ortega, D., Ibañez, J. J., Khalidi, L., Méndez, V., Campos, D., & Teira, L. (2014). Towards a multi-agent-based modelling of obsidian exchange in the neolithic Near East. Journal of Archaeological Method and Theory, 21, 461–485.
Osorio, D., Capriles, J. M., Ugalde, P. C., Herrera, K. A., Sepúlveda, M., Gayó, E. M., et al. (2017). Hunter-gatherer mobility strategies in the high Andes of northern Chile during the late Pleistocene–early Holocene transition (ca. 11,500–9,500 cal B.P.). Journal of Field Archaeology, 17(3), 228–240.
Osorio, D., Steele, J., Sepúlveda, M., Gayo, E. M., Capriles, J. M., Herrera, K., et al. (2017). The dry puna as an ecological megapatch and the peopling of South America: Technology, mobility, and the development of a late Pleistocene/early Holocene Andean hunter-gatherer tradition in northern Chile. Quaternary International, 461, 41–53.
Pearsall, D. (2008). Plant domestication and the shift to agriculture in the Andes. In H. Silverman & W. H. Isbell (Eds.), Handbook of South American archaeology (pp. 105–120). Springer.
Pestle, W. J., Torres-Rouff, C., Hubbe, M., Santana, F., Pimentel, G., Gallardo, F., et al. (2015). Explorando la diversidad dietética en la prehistoria del Desierto de Atacama: Un acercamiento a los patrones regionales. Chungara Revista de Antropología Chilena, 47(2), 201–209.
Pfeiffer, M., Heimsath, A., Santoro, C. M., Latorre, C., Gayo, E., Finstad, K., et al. (2018). Chronology, stratigraphy and hydrological modelling of extensive wetlands and paleolakes in the hyperarid core of the Atacama Desert during the late Quaternary. Journal of Quaternary Science Review, 197, 224–245.
Pimentel, G., Ugarte, M., Gallardo, F., Blanco, J. F., & Montero, C. (2017). Chug-Chug en el contexto de la movilidad internodal prehispánica en el Desierto de Atacama, Chile. Chungara Revista de Antropología Chilena, 49(4), 483–510.
Pintar, E. L. (2008). Astiles, intermediarios y sistemas dearmas en la puna salada. Cazadores-Recolectores del Cono Sur. Revista de Arqueología, 3, 113–125.
Pintar, E. L. (2014). Desert hunter-gatherers: Mobility and aridity thresholds. A view from the Argentine Puna. In E. Pintar (Ed.), Hunter-gatherers from a high elevation desert: People of the salt puna (northwest Argentina). BAR International Series 2641 (pp. 97–118). Archaeopress.
Pintar, E., Martínez, J. G., Aschero, C. A., & Glascock, M. D. (2016). Obsidian use and mobility during the early and middle Holocene in the Salt Puna, NW Argentina. Quaternary International, 422, 93–108.
Pitblado, B., & Rademaker, K. (2017). The peopling of high-altitude landscapes of the Americas. Quaternary International, 461, 1–3.
Placzek, C., Quade, J., Betancourt, J. L., Patchett, P. J., Rech, J. A., Latorre, C., et al. (2009). Climate in the dry central Andes over geologic, millennial, and interannual timescales. Annals of the Missouri Botanical Garden, 96(3), 386–397.
Politis, G., Gutiérrez, M. A., Rafuse, D., & Blasi, A. (2016). The arrival of Homo sapiens into the southern cone at 14,000 years ago. PLoS ONE, 11(9), e0162870.
Quade, J., Rech, J. A., Betancourt, J. L., Latorre, C., Quade, B., Rylander, K. A., & Fisher, T. (2008). Paleowetlands and regional climate change in the central Atacama Desert, northern Chile. Quaternary Research, 69, 343–360.
Rademaker, K., & Hodgins, G. (2018). Exploring the chronology of occupations and burials at Cuncaicha rockshelter, Peru. In K. Harvati, G. Jäger, & H. Reyes-Centeno (Eds.), New perspectives on the peopling of the Americas. Words, bones, genes, tools: DFG Center for Advanced Studies (pp. 107–124). Kerns Verlag.
Rademaker, K., & Moore, K. (2019). Variation in the occupation intensity of early forager sites of the Andean puna. In A. K. Lemke (Ed.), Foraging in the past: Archaeological studies of hunter-gatherer diversity (pp. 76–118). University Press of Colorado.
Roberts, A., Pate, F. D., Petruzzelli, B., Carter, C., Westaway, M., Santoro, C. M., et al. (2013). Retention of hunter-gatherer economies among maritime foragers from Caleta Vitor, northern Chile, during the late Holocene: Evidence from stable carbon and nitrogen isotopic analysis of skeletal remains. Journal of Archaeological Sciences, 40, 2360–2372.
Rothhammer, F. (2014). Temprana complejidad funeraria en la costa norte de chile: Origen, desarrollo, funcionalidad y relevancia antropológica. Chungara Revista de Antropología Chilena, 46(1), 145–151.
Rothhammer, F., Santoro, C. M., Poulin, E., Moraga, M., & Standen, V. G. (2009). Archeological and mtDNA evidence for tropical lowland migrations and cultural change during the Late Archaic/Formative in northern Chile. Revista Chilena de Historia Natural, 82(4), 543–552.
Salinas, P., Naranjo, J. A., & Marshall, L. G. (1991). Nuevos restos del perezoso gigante (Megatheriidae, Megatherium Medinae) de la formación Chiu-Chiu, cuenca del río Loa, Calama, norte de Chile. In Actas de Resúmenes Expandidos Congreso Geológico Chileno No. 6, Vol. 1. (pp. 306–309).
Santoro, C.M. (1989). Antiguos cazadores de la puna (9000–6000 a.C.). In J. Hidalgo, V. Schiappacasse, H. Niemeyer, C. Aldunate & I. Solimano (Eds.), Culturas de Chile: Prehistoria, desde sus orígenes hasta los albores de la conquista (pp. 33–55). Santiago: Editorial Andrés Bello.
Santoro, C. M., & Núñez, L. (1987). Hunters of the dry puna and the salt puna in northern Chile. Andean Past, 1, 57–109.
Santoro, C. M., Standen, V. G., Arriaza, B. T., & Dillehay, T. D. (2005). Andean Archaic funerary pattern or postdepositional alteration? The Patapatane burial in the highlands of South Central Andes. Latin American Antiquity, 16(3), 329–346.
Santoro, C. M., Rivadeneira, M., Latorre, C., Rothhammer, F., & Standen, V. G. (2012). Rise and decline of Chinchorro sacred landscapes along the hyperarid coast of the Atacama Desert. Chungara Revista de Antropología Chilena, 44(4), 637–653.
Santoro, C. M., Osorio, D., Ugalde, P., Sepúlveda, M., Cartajena, I., Standen, V. G., et al. (2016), Cazadores, recolectores y pescadores arcaicos del Desierto de Atacama: Entre el Pacífico y los Andes, norte de Chile (ca. 10.000 a 3.700 años a.p.). In C. Aldunate, F. Falabella, L. Sanhueza, & M. Uribe (Eds.), Prehistoria: Culturas de Chile (pp. 117–180). Santiago: Editorial Universitaria.
Santoro, C. M., Gayo, E. M., Capriles, J. M., de Porras, M. E., Maldonado, A., Standen, V. G., et al. (2017). Continuities and discontinuities in the socio-environmental systems of the Atacama Desert during the last 13,000 years. Journal of Anthropological Archaeology, 46, 28–39.
Santoro, C. M., Gayo, E. M., Capriles, J. M., Rivadeneira, M. M., Herrera, K. A., Mandakovic, V., et al. (2019). From the Pacific coast to the tropical forests: Late Pleistocene network of interaction in Pampa del Tamarugal, northern Chile Atacama Desert. Chungara Revista de Antropología Chilena, 51(1), 5–25.
Santoro, C. M., Castro, V., Carter, C., & Valenzuela, D. (2020). Marine communities in the Atacama Desert: Masters of the sub-tropical Pacific coast of South America. In G. Prieto & D. H. Sandweiss (Eds.), Maritime communities of the Andes (pp. 39–73). University Press of Florida.
Simberloff, D. (1988). The contribution of population and community biology to conservation science. Annual Review of Ecology and Systematics, 19, 473–511.
Sierralta, S. (2019). Función y selección de materias primas en la transición Pleistoceno–Holoceno: Punta Negra e Imilac, región de Antofagasta, Chile. Intersecciones En Antropología, 20(1), 11–23.
Standen, V. G., Arriaza, B. T., Santoro, C. M., & Santos, M. (2014). La práctica funeraria en el sitio Maestranza Chinchorro y el poblamiento costero durante el Arcaico medio en el extremo norte de Chile. Latin American Antiquity, 25(3), 300–321.
Standen, V. G., Santoro, C. M., Arriaza, B. T., & Coleman, D. (2018). Hunting, gathering, and fishing on the coast of the Atacama Desert: Chinchorro population mobility patterns inferred from strontium isotopes. Geoarchaeology: An International Journal, 33(2), 162–176.
Steele, J., & Rockman, M. (2003). ‘Where do we go from here?’ Modelling the decision making during exploratory dispersal. In M. Rockman & J. Steele (Eds.), Colonization of unfamiliar landscapes: The archaeology of adaptation (pp. 130–143). Routledge.
Swift, J., Cupper, M. L., Greig, A., Westaway, M. C., Carter, C., Santoro, C. M., et al. (2015). Skeletal arsenic of the pre-Columbian population of Caleta Vitor, northern Chile. Journal of Archaeological Science, 58, 31–45.
Tamayo, M., & Frassinetti, D. (1980). Catálogo de los mamíferos fósiles y vivientes de Chile. Boletín del Museo Nacional de Historia Natural, 37, 323–399.
Tully, C. D., Rech, J. A., Workman, T. C., Santoro, C. M., Capriles, J., Gayó, E. M., et al. (2019). In-stream wetland deposits, mega-droughts, and cultural change in the Atacama Desert, Chile. Quaternary Research, 91, 63–80.
Ugalde, P. C., Quade, J., Santoro, C. M., & Holliday, V. T. (2020). Processes of Paleoindian site and desert pavement formation in the Atacama Desert, Chile. Quaternary Research, 98, 58–80.
Varela, H. H., Cocilovo, J. A., Quevedo, S., & Costa-Junqueira, M. A. (2004). La estructura de la población de pescadores del período Arcaico Tardío y Formativo de Arica. Boletín del Museo Nacional de Historia Natural, 53, 149–160.
Varela, H. H., O’Brien, T. G., & Cocilovo, J. A. (2008). The genetic divergence of prehistoric populations of the South-Central Andes as established by means of craniometric traits. American Journal of Physical Anthropology, 137, 274–282.
Villavicencio, N. A., Corcoran, D., & Marquet, P. A. (2019). Assessing the causes behind the late Quaternary extinction of horses in South America using species distribution models. Frontiers in Ecology and Evolution, 7, 226.
Weinstein, K. J. (2017). Morphological signatures of high-altitude adaptations in the Andean archaeological record: Distinguishing developmental plasticity and natural selection. Quaternary International, 461, 14–24.
Whittaker, R. J. (1998). Island biogeography. Ecology, evolution, and conservation. Oxford University Press.
Wobst, M. (1974). Boundary conditions for palaeolithic social systems: A simulation approach. American Antiquity, 39(2, part 1), 147–178.
Workman, T. R., Rech, J. A., Gayo, E. M., Santoro, C. M., Ugalde, P. C., De Pol-Holz, R., et al. (2020). Landscape evolution and the environmental context of human occupation of the southern Pampa del Tamarugal, Atacama Desert, Chile. Quaternary Science Reviews, 243, 106502.
Yacobaccio, H. D. (1994). Biomasa animal y consumo en el Pleistoceno – Holoceno surandino. Arqueología, 4, 43–71.
Yacobaccio, H. D. (2017). Peopling of the high Andes of northwestern Argentina. Quaternary International, 461, 34–40.
Yacobaccio, H. D., & Madero, C. M. (1992). Zooarqueología de Huachichocana III (Prov. De Jujuy, Argentina). Arqueología, 2, 149–188.
Yacobaccio, H. D., & Morales, M. R. (2011). Ambientes pleistocénicos y ocupación humana temprana en la Puna Argentina. Boletín De Arqueología PUCP, 15, 337–356.
Yacobaccio, H. D., Morales, M. R., & Hoghin, R. (2017). Habitats of ancient hunter-gatherers in the Puna: Resilience and discontinuities during the Holocene. Journal of Anthropological Archaeology, 46, 92–100.
Acknowledgements
Fondo Nacional de Ciencia y Tecnología (FONDECYT) grant 1201786, Comisión Nacional de Investigación Científica y Tecnológica (CONICYT)/Programa Cooperación Internacional, Modalidad de Estadías Cortas (MEC) project PAI80160111, CONICYT/Programa de Investigación Asociativa (PIA) project ANILLO SOC1405, CONICYT/Programa de Cooperación Internacional (PCI), project PII20150081. We thank Paola Salgado for fixing Figure 1 and Persis Clarkson for translating Luis Advis’ poem.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Borrero, L.A., Santoro, C.M. Metapopulation Processes in the Long-Term Colonization of the Andean Highlands in South America. J World Prehist 35, 135–162 (2022). https://doi.org/10.1007/s10963-022-09167-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10963-022-09167-x