
Abstract
Diatoms are universally recognized as good bio-indicators due to their high diversity and rapid response to environmental and climate changes. This paper explores whether the sedimentary diatom record from Sanjiaolongwan Maar Lake (SJML), northeastern China, reveals variations in seasonal pattern over the past 1000 years. The observed succession in diatom assemblages reveals that between 980 and 1310 CE the small-celled planktonic diatom Discostella pseudostelligera was abundant. Its relative abundance peaked during 1020–1060 CE, indicating that the duration of summer conditions over this interval was the longest within the last millennium. By contrast, the interval between 1470 and 1850 CE was marked by an increase in benthic diatoms pointing to a shortened duration of the ice-free season, and generally cold conditions. During the twentieth century warm period, the succession of eutrophic diatoms and changes in diatom cell size also reveal temperature-controlled seasonal variations in limnological conditions. The differences observed between the diatoms assemblages associated with the two warm periods indicate that the summers of the warmest part of the Medieval Climate Anomaly were longer than that of the current warm period. This result is supported by the diatom record from Erlongwan, another maar lake in the region that differs from SJML by its topographic features and in the level of human impact on its catchment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climate change is causing change in the distribution and duration of seasons. However, our understanding of variations in the relative length and amplitude of seasons is mainly derived from the meteorological monitoring data of the last ~ 50 years. Orlowsky and Seneviratne (2012) found that in the Northern Hemisphere beyond 50° north, the largest seasonal warming amplitude occurs during winter and autumn, while for the mid to low latitudes of Americas and in the Mediterranean region, the largest seasonal warming amplitude occurs in summer. In China, Yan et al. (2011) analyzed the daily temperature data from 541 meteorological stations for the period 1963–2011 CE and showed that the duration of winter decreased by 2–6 days every decade. The shortening of winter was caused by the early ending of the season, while the extension of summer was caused by the delaying of autumn, which leads to the shortening of autumn as well (Yan et al. 2011).
So far, there are very few investigations about the variations in the relative length and amplitude of seasons over the past 1000 years. Despite using a wide range of proxy records in different types of ecosystem, most studies on seasonal variations in the last millennium have focused on single-season changes. For example, carbonates in marine sediments of the Mediterranean Sea record temperature and precipitation changes in summer and winter (Goudeau et al. 2015); high latitude tree-ring records reveal summer temperature changes (Jan et al. 2002; Cook et al. 2004, 2013; D' Arrigo et al. 2006) while tree-ring records from the middle and low latitudes are mainly used to reconstruct rainfall change during the wet season (Zhang et al. 2011). Reconstructions based on alkenones (Liu et al. 2006; Chu et al. 2012) and chironomid assemblages (Larocque et al. 2010; Chang et al. 2018) in lakes in the Northern Hemisphere often reflect the changes in summer temperature. Similarly, the high-resolution stalagmite laminae of Shihua Cave near Beijing, China, also record variations in summer temperature (Tan et al. 2003). None of these studies, however, discusses the variations in the relative length of the seasons.
Lakes provide good sites for the study of seasonal variation over thousands of years. They can be sensitive to regional climate and environmental changes, and therefore act as recorders of climate change. In addition, lacustrine sediments can record the interaction between climate change and human activities (Williamson et al. 2009). The physical and chemical shifts associated with seasonal variations are often recorded in lacustrine sediments (Phlips et al. 2007; Srifa et al. 2016). Among all types of lakes, maar lakes, which originate from phreato-magmatic explosions, are particularly sensitive to seasonal variations in climate because they usually have a small catchment and limited inflow or outflow and are mostly located in remote areas with fewer disturbances. Moreover, these lakes generally have a flat bottom, usually have a very large initial depth and a high sedimentation rate (Liu et al. 2000), characteristics that are all conducive to the formation and preservation of laminae, enabling them to provide high-resolution paleoclimate records (Zolitschka et al. 2000).
In the study of lake sediments, diatoms are universally recognized as good indicators of environmental change due to their high diversity, rapid response to changes, and high preservation potential, and they are widely used to investigate changes in temperature (Smol et al. 2005; Smol and Douglas 2007; Bopp et al. 2005; Rühland et al. 2008, 2010, 2013; Li et al. 2021), salinity (Fritz 1990; Fritz et al. 1999; Yang et al. 2003), snowfall (Wang et al. 2018), as well as environmental changes caused by human activities that havean effect on lake trophic level (Yang et al. 2006, 2008; Dong et al. 2006, 2008; Chen et al. 2011; Liu et al. 2016), and lake acidification (Davis 1987; DeNicola 2000). Numerous studies have shown that seasonal variations in the composition of diatom assemblages are related to the seasonal variations in the physical characteristics of the water column, such as the duration of lake ice-cover, thermal stratification, and mixing of the water column (Lotter and Bigler 2000; Smol and Douglas 2007; Tolotti et al. 2007; Rühland et al. 2008, 2010, 2013; Winder and Sommer 2012). Thus, diatoms have the potential to be used to assess how the seasons, and the specific lake conditions associated with them, are changing in conjunction with the changes in climate.
Therefore, seasonal diatom succession has been used to decipher past seasonal variations in climate, such as temperature and monsoon. For example, in the study of Erlongwan Maar Lake (EML) in northeastern China, Wang et al. (2012a) used the composition of diatom assemblages to detect signals of climate change, despite the fact that this lake had been affected by human activities in the twentieth century. Specifically, Wang et al. (2012a) found that summers during the Medieval Climate Anomaly (MCA) was significantly longer than those in the twentieth century, and spring and fall were shorter. Seasonal succession of diatoms may also be related to wind conditions, such as in the East Asian Monsoon region, where heavy diatoms with higher nutritional requirements dominate during the period with strong winter monsoon (strong mixing), while smaller diatoms with lower nutritional requirements dominate during the period with weak winter monsoon (weak mixing) (Wang et al. 2012b). However, there are still few studies on seasonal variations over long time scales.
It should be pointed out that the seasonal variation in this study refers to the relative length of seasonal climatic conditions, which in temperate lakes are best represented by the change in relative length of the lake stratification period in summer and that of the mixing period in spring and autumn. In different climatic periods, the relative length of seasons will change. In a region such as northeastern China, when the climate becomes colder (e.g. during the Little Ice Age), winter conditions and in particular the duration of ice-cover will be prolonged. Inversely, during warm periods (e.g. the Medieval Warm Period) the duration of ice-cover will be shortened while the length of thermal stratification in summer will be prolonged (Wang et al. 2012a).
Sanjiaolongwan Maar Lake (SJML), the site of our study, is located in the same temperate monsoon-controlled region as EML, whose climate has an obvious seasonality. This site is therefore suitable to study the variations in relative length of seasons. The morphological characteristics of SJML, the hydrological parameters of its water column and sediment composition have already been studied (Mingram et al. 2004), as well as the regional vegetation changes of the last millennium (Zhang et al. 2019) but no paleolimnological investigation has been carried out yet. Thus, this paper aims to discuss the paleolimnological history of this lake from the perspective of diatoms. Historical documents testify that very few humans lived in this area of northeastern China before 1860 CE (Editorial Board for flora of China 1995), until the Qing government abolished the residence ban, after which the human population rapidly increased in the region (Ma and Ma, 1990). Therefore, human activities started to increase after 1860 CE. In addition, by comparison with EML, SJML is located closer to a population center, and pollen records (Zhang et al. 2019) from this lake show that the impact of human activities in the late twentieth century has been greater than those recorded in the sediments of EML. In that context, with stronger influence of human activities, can the sedimentary record from SJML also be used to identify signals of climate change? It is also worth discussing whether if the different diatom assemblages in SJML and EML can reflect the same pattern of seasonal variation.
Thus, this study will analyze diatom assemblages in SJML with the aim to: (1) generate a diatom record for SJML that covers the last millennium; (2) test the hypothesis of Wang et al. (2012a) that concludes that MCA summers were longer than those in the twentieth century and spring and fall were shorter; and (3) determine whether proximity to a center of human occupation has an impact on that record.
Materials and methods
Study site
SJML (42°21.57′N, 126°25.36′E, and 730 m.a.s.l.) is located in the Longgang volcanic field (LGVF) in Jilin Province, in the western part of Changbai Mountains in northeastern China (Fig. 1). The lake is dumbbell-shaped, with its major axis extending in the NW–SE direction, for about 1100 m, and its minor axis extending in the NE-SW direction, for about 550 m (Liu et al. 2000). The catchment area is 0.9 km2 and the lake surface area is 0.7 km2. SJML was formed by two phreatomagmatic episodes (Liu et al. 2000) and is divided into two basins: the main basin is 0.45 km2 in area, with a maximum water depth of 76 m; the smaller basins is 0.25 km2 in area and has a maximum water depth of 39 m. The lake is replenished by summer precipitation and groundwater inflow, without natural surface inflows or outflows. Forest vegetation in the study area consists of temperate coniferous and broad-leaved mixed forest (Editorial Committee of Vegetation Map of China 2007). Due to selective felling of Pinus koraiensis within the regional forest, following the Japanese invasion of northeastern China in the 1930s–1940s, the original Pinus koraiensis-dominated coniferous and broadleaved mixed forest was replaced by broad leaved secondary forest (Liu 1989).

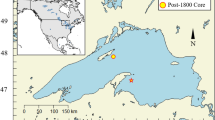
Location of Sanjiaolongwan Maar Lake in northeastern China (a). The satellite photo shows the study site, the Sanjiaolongwan Maar Lake, and the location of the coring site within the smaller basin of the lake (b). Note the strong asymmetry of the lake basin
From the analyses of five surface water samples collected between 2005 and 2010 the chemical characteristics of SJML can be summarized as follows. SJML is slightly alkaline, with pH ranging from 6.6 to 8.3, and conductivity from 71 to 96 μs⋅cm−1. Its trophic status is in the oligotrophic range, with concentrations of total phosphorus (TP) and total nitrogen (TN) from 3 to 8 μg L−1 and 200 to 585 μg L−1, respectively. The concentration of dissolved organic carbon (DOC) is low and is in the range 0.2–2.5 mg L−1. The concentrations (in mg L−1) of the main chemical elements are relatively low and vary in the following ranges: Si4+ (0.1–0.5), Na+ (5.3–6.4), Ca2+ (5.9–7.9), K+ (2.6–3.5), Mg2+ (2.6–3.2), Cl− (1.3–2.5), NO3− (0.1–1.5) and SO42− (3.9–5.0) (Table 1).
Regional climate
The region is characterized by a temperate continental climate and is influenced by the East Asian Summer Monsoon and the Siberian High pressure system. Therefore, summers are humid and rainy, while springs and autumns are dry and windy. The annual average temperature is 3.9 °C with a sharp contrast between the warmest and coldest months. The average temperature in the warmest month (July) is 21 °C, while the winter is very cold with an average temperature in the coldest month (January) of − 17 °C. The freezing season is long, lasting for mid-October to late-March of the following year. The average annual rainfall is 780 mm, 60% of which falls from June through August (Fig. 2). These climatic data (compiled for the period from 1981 to 2010 CE) were recorded at the Jing Yu meteorological station (Meteorological data of China 1981–2010), located about 50 km to the east of SJML at 550 m.a.s.l.

Average monthly meteorological data from 1981 to 2010 CE, recorded at Jing Yu Station
Laboratory methods
In August 2007, a 59-cm-long and 60-mm-diameter sediment core was retrieved from the central, deepest region of the smaller basin using a UWITEC gravity corer, and was subsequently sliced into 0.5-cm intervals for analysis. The core is mainly composed of homogeneous, layered or finely laminated diatomaceous gyttja. Its composition is therefore similar to that of another core retrieved from the deep basin in a previous study (Mingram et al. 2004). The 0.5-cm thick sediment intervals is thicker than the laminations and therefore do not correspond to the annual seasonal variation. However, this does not affect the aim of this project, which aims to assess the variation in the average length of a certain season within a certain period of time (e.g. 20–30 years).
An age model was developed using radiometric 210Pb, 137Cs and Accelerator Mass Spectrometry (AMS) 14C analyses. The activities of 137Cs and 210Pb of 30 samples (0–15-cm core depth) were measured by gamma spectrometry using a low-background well-type germanium detector (EGPC 100P-15R) at the Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing. Radiometric dates were calculated using the constant rate of supply (CRS) 210Pb dating model (Appleby and Oldfield 1978; Appleby et al. 1986), and verified by using the 137Cs stratigraphic record, especially the peak value corresponding to the maximum radiation amount of nuclear weapon atmospheric test in ~ 1963 CE (Appleby 2001; Jha et al. 2003). AMS 14C Analytic (Beta Analytic Incorporated USA) was used to date terrestrial leaf macrofossils at the core depths of 21 cm, 34 cm, and 46 cm and bulk organic sediments at the depth of 58 cm. The radiocarbon dates were calibrated using the IntCal 13 radiocarbon calibration curve (Table 2).
Diatom laboratory preparation consisted in placing a weighed amount (around 0.02 g) of dry sediment sample into a conical test tube, then adding H2O2 (30%) first and then one drop of HCl (36%) after H2O2 reacted completely while heating in a water bath in order to remove organic matter and carbonate, respectively (Battarbee et al. 2001). Diatom concentrations (valves g−1) were calculated by the addition of external markers consisting of a suspension of divinyl benzene microspheres with a known concentration (Battarbee and Kneen 1982). Diatom slides were prepared using the mountant Naphrax® (Refractive Index = 1.7) on a hot plate. For each sample at least 500–600 valves were counted using oil immersion phase-contrast light microscopy at × 1000 magnification. Diatom identification was mainly based on the Krammer and Lange-Bertalot’s book series (1986–1991) but more recent and local floras were also used and the nomenclature was updated using online databases such as Algaebase (Guiry and Guiry 2021) and Diatoms of North America (https://diatoms.org).
Diatom data are displayed as relative abundance and flux. Diatom percentage and flux diagrams were created using the C2 software, version 1.5 (Juggins 2007). Principal component analysis (PCA) was used to summarize the diatom data using CANOCO version 5 (ter Braak and Smilauer 2012). Only 11 taxa were included in the analysis (those with > 1% relative abundance in any one sample). Cluster analysis was conducted using the PAST software (Hammer et al. 2001), based on Gower distance using the un-weighted pair-group method with the arithmetic mean (UPGMA).
Results
Chronology
Based on the results of 210Pbex (ex = excess) and AMS 14 C dating, a chronology was established using the Bacon age-depth model (Blaauw and Christen 2011) (Fig. 3). The three dates obtained from terrestrial leaf macrofossils (Table 2) indicate that the sedimentation rate varied little between the core depths 46–21 cm. If we extrapolate this sedimentation rate to the bottom of the core, the sediments at 58 cm were deposited between 750 and 1170 CE. However, the actual AMS 14C dates for that sample at 58 cm was 652–722 CE (Table 2). This large offset from the Bacon model (orange mark in Fig. 3) may be due to a radiocarbon reservoir effect at the bottom of the core, due to “old carbon” dust in the sample, which was a bulk organic sample instead of plant macrofossil. Therefore, we decided to exclude this bulk organic sediment sample at 58 cm from the age model and extrapolated the Bacon age-depth model from 46 to 59 cm to obtain the final age model (Fig. 3). Based on this model the core spans the interval from 980 to 2004 CE. Although the extrapolation error is rather large, it has little influence on the diatom-related discussion on variations in seasonality during the cold and warm intervals. In addition, this time resolution is sufficient to clearly distinguish the three cold and warm intervals so that we can compare and analyze the differences of average seasonal conditions between these intervals.

Records of 210Pbex, and 137Cs activities and AMS 14C dates and a combined age model for the sediment core from the Sanjiaolongwan Maar Lake
Variation of diatom relative percentages and fluxes
A total of 31 genera and 131 species of diatoms were identified in the 118 samples analyzed from the SJML sediment core. Since the species belonging to the genera Achnanthes (sensu lato), Amphora (sensu lato), and Navicula (sensu lato) are all benthic species with very low percentages in the assemblages, they were combined at these broad generic levels. The planktonic species Lindavia balatonis dominated almost the whole record and only decreased in recent decades. The relative abundance of two other planktonic species, Discostella pseudostelligera and Fragilaria tenera, were very high in 980–1310 CE and 1850–1965 CE, but decreased notably in 1470–1850 CE and in the most recent decades. The abundances of the remaining planktonic diatoms such as Asterionella formosa and Stephanodiscus parvus, have increased significantly in recent decades. Benthic diatoms such as Tabellaria flocculosa strain IV, Fragilaria capucina, Achnanthes s.l., Amphora s.l., and Navicula s.l. are generally low in abundance, and only increase in 1310–1850 CE.
Using cluster analysis base on abundance data, diatom percentages (Fig. 4) and fluxes (Fig. 5) were divided into four zones that are described below. In addition, we also distinguished subzones.

The Sanjiaolongwan Maar Lake diatom percentage diagram for the last millennium. Only major species and genera are plotted. The result of the cluster analysis used to define the assemblage zones is plotted on the right hand side

The Sanjiaolongwan Maar Lake diatom fluxes for the last millennium. Only major species and genera are plotted
Zone SJML-1 (59.0–42.0 cm, 980–1310 CE). This zone is dominated by planktonic species (75–92%, mean abundance of 84%). According to the changes in relative abundance of the two dominant planktonic species, L. balatonis and D. pseudostelligera, this zone can be divided into three subzones.
Zone SJML-1a (59.0–55.5 cm, 980–1050 CE). The relative abundance of L. balatonis decreased rapidly from a high value (65%) to a relative low value (33%). At the same time, D. pseudostelligera increased rapidly until it reached a peak (58%) around 1040 CE. Among the other planktonic species, the mean abundance of F. tenera is only 3%. The abundances of benthic diatoms such as Amphora s.l. (mean abundance of 1%), Navicula s.l. (mean abundance of 1%), F. capucina (mean abundance of 1%), and Achnanthes s.l. (mean abundance of 5%) were constantly low with no large variations. Total diatom flux relatively high (mean 17.6 × 106 valves cm−2 yr−1), and the main contributors are L. balatonis and D. pseudostelligera.
Zone SJML-1b (55.5–49.0 cm, 1050–1180 CE). The relative abundance of L. balatonis increased rapidly (49–74%), while D. pseudostelligera decreased sharply (33–6%). F. tenera is relatively stable, with no large variations (mean abundance of 3%). In the benthic species, only the relative abundance of T. flocculosa strain IV (mean abundance of 3%) and F. capucina (mean abundance of 2%) increased slightly. The flux of D. pseudostelligera decreased markedly, but the total flux was still very high (mean 18.7 × 106 valves cm−2 yr−1) due to the contribution of L. balatonis.
Zone SJML-1c (49.0–42.0 cm, 1180–1310 CE). Planktonic species in this subzone gradually decreased (87–75%), but were still dominant in the assemblages (mean abundance of 82%). L. balatonis decreased briefly (74–48%), then returned to a high value (mean abundance of 68%), and finally decreased again (mean abundance of 60%). Fluctuations in D. pseudostelligera are just the opposite, with a first rapid increase (20–33%), then a decrease to a low value (9%), and recovery (mean abundance of 21%). F. tenera briefly increased (8%) and then gradually disappeared. The relative abundance of the benthic diatom T. flocculosa strain IV decreased slightly (mean abundance of 1%), while F. capucina continued to increase (4–10%, mean abundance of 7%). The flux of planktonic diatoms did not change markedly, and the total flux of diatoms was still high (mean 22.2 × 106 valves cm−2 yr−1).
Zone SJML-2 (42.0–17.0 cm, 1310–1850 CE). The percentage of planktonic species continue to decline slowly, while the percentages of benthic species increased. According to the variability in the percentages of planktonic diatoms, the zone was divided into two subzones.
Zone SJML-2a (42.0–34.0 cm, 1310–1470 CE). The abundance of L. balatonis decreased, but remains high, with a mean abundance of 55%, D. pseudostelligera decreased to 15% and F. tenera remained below 1%. The decline of planktonic diatoms (77–68%) is largely driven by a decrease in L. balatonis and D. pseudostelligera. The abundance of the benthic diatoms T. flocculosa strain IV and Achnanthes s.l., increased slightly to 6% and 8%, respectively. The total flux of diatoms decreased with the decrease of fluxes of the two dominant planktonic species (mean 21.0 × 106 valves cm−2 yr−1).
Zone SJML-2b (34.0–17.0 cm, 1470–1850 CE). This zone is mainly characterized by the large decrease of D. pseudostelligera (mean abundance of 8%), L. balatonis abundances remained high (mean abundance of 56%), while the overall percentages of planktonic diatoms are at their lowest level for the entire sequence (mean abundance of 69%). Benthic diatoms such as T. flocculosa strain IV (mean abundance of 8%), the Achnanthes s.l. (mean abundance of 9%) and the Navicula s.l. (mean abundance of 3%) have increased significantly. The total diatom flux decreased (mean 11.9 × 106 valves cm−2 yr−1), mainly driven by a large decrease in the flux of planktonic diatoms, while the flux of benthic diatom only decreased slightly.
Zone SJML-3 (17.0–7.5 cm, 1850–1965 CE). The main feature of this zone is that the abundances of D. pseudostelligera (mean abundance of 18%) and F. tenera (mean abundance of 5%) increased. L. balatonis decreased to 51% in mean abundance, but it still dominates the assemblages. T. flocculosa strain IV increased slightly (mean abundance of 11%), while other benthic diatoms such as F. capucina, the Achnanthes s.l., the Amphora s.l. and the Navicula s.l. decreased slightly. The total diatom flux increased (mean 33.2 × 106 valves cm−2 yr−1), driven by significantly increased fluxes of D. pseudostelligera and F. tenera, and also increasing flux of benthic diatoms.
Zone SJML-4 (7.5–0.5 cm, 1965–2004 CE). This zone is still dominated by planktonic species, but the assemblage is quite different from SJML-1, D. pseudostelligera and F. tenera abundances are very low and these species almost disappear from the assemblages, while S. parvus and A. formosa appear in large number. According to this species shift, the assemblages were divided into two subzones.
Zone SJML-4a (7.5–5.5 cm, 1965–1975 CE). In this zone, the abundance of D. pseudostelligera and F. tenera decreased rapidly, nearly going locally extinct, and the abundance of L. balatonis also decreased quickly (41–8%). S. parvus increased sharply, reaching a peak (66%) in 1970 CE, while formosa also increased (mean abundance of 16%). Planktonic percentages increased again to a very high level (mean abundance of 95%). The relative abundances of benthic diatoms such as T. flocculosa strain IV, F. capucina, Achnanthes s.l., Amphora s.l., and the Navicula s.l. decreased significantly or even disappeared from the assemblages. The flux of S. parvus increased rapidly but quickly declined after a short interval. The flux of A. formosa and F. tenera increased greatly, and the fluxes of the other planktonic diatoms such as D. pseudostelligera and L. balatonis decreased significantly. The total diatom flux was very high (mean 51.9 × 106 valves cm−2 yr−1).
Zone SJML-4b (5.5–0.5 cm, 1975–2004 CE). This zone is dominated by A. formosa and L. balatonis which increased significantly to mean abundances of 47% and 36%, respectively. S. parvus suddenly decreased from the previous peak to less than 1%, and then increased slightly (mean abundance of 6%). Other planktonic diatoms such as D. pseudostelligera and F. tenera reappear in the assemblage, but their increase is not large. The percentages of planktonic taxa remained at a very high level (mean abundance of 93%). The flux of A. formosa and L. balatonis decreased slightly, and the flux of D. pseudostelligera, F. tenera and S. parvus are very low. The total flux of diatoms decreases (mean 22.1 × 106 valves cm−2 yr−1).
Principal components analysis (PCA)
A PCA was carried out on the diatom data from SJML to try to identify the main environmental factors affecting the changes observed in the diatom assemblages. A preliminary analysis of the entire data set and a check on the variable loadings on the first principal component (Fig. 6a), suggested that PCA axis 1 represents a trophic gradient. The plot of the sample scores on the first two principal components (Fig. 6b) defines two distinct sample groups. Group I comprises samples from the depth interval 0–7 cm (post ~ 1965 CE), and Group II comprises samples from the depth interval 7.5–59 cm (1965–980 CE). After 1965 CE, the relative percentages of S. parvus and A. formosa which are two species with high nutritional needs (Dong et al. 2006; Bennion et al. 2011) increased rapidly and dominate the assemblages. Moreover, the pollen record indicates strong human activities in this area after 1960s (Zhang et al. 2019). The increase in the lake trophic status, associated with human disturbances the most plausible explanation for the two distinct sample groups shown in the PCA plot.

Results of principal components analysis (PCA) of the diatom percentage data from for the whole sequence (the depth interval 0–59-cm) of the Sanjiaolongwan Maar Lake. Sample scores of the most abundant and ecologically most-indicative diatom taxa on PCA axis 1 and axis 2 (a) and sample distribution with different core depths on PCA axis 1 and axis 2 (b)
In order to better identify the natural influencing factors we selected a sub-set of samples for another PCA, consisting of the depth interval of 7.5–59 cm. The results are plotted in Fig. 7. The first two principal components (PCA axis 1 and PCA axis 2) account for 61% and 27% of the total variance, respectively. On Axis-1, D. pseudostelligera and F. tenera have highly positive scores while L. balatonis has a highly negative score.

Results of principal components analysis (PCA) of the diatom percentage data from the depth intervals of 7.5–59-cm of the Sanjiaolongwan Maar Lake. Only the most abundant and ecologically most indicative diatom taxa are shown. Axis 1 is best interpreted as a gradient in water column turbulence
Discussion
Relationship between diatom assemblages and environment
Planktonic diatoms are dominant in the whole sequence, while benthic diatoms have relatively low abundance (Fig. 4). This is mainly related to the basin morphology of SJML. SJML is a deep lake with a narrow littoral zone and the rapidly increasing water depth restricts the extent of benthic diatom habitats inside the lake basin. By contrast, the open water provides a large area for planktonic diatoms to develop that is little affected by changes in lake level. Only a dramatic decrease in water level (by more than 20 m), resulting in the potential growth of benthic diatoms at the bottom of the shallower lake basin, would significantly increase the relative content of benthic diatoms in the sediment at the coring site (Wang et al. 2012a). Although evaporation during the Medieval Climate Anomaly (MCA) was surely higher than during the Little Ice Age (LIA), the amount of rainfall was also higher during the MCA (Chen et al. 2015). Therefore, it is unlikely that the water depth of SJML changed greatly in the past 1000 years. The diatom assemblages also indicate that the planktonic species were still dominant, implying that the lake remained deep throughout the record.
In addition, it should be noted that the planktonic diatom L. balatonis remained highly abundant except for two short intervals in 1020–1060 CE and 1965–1975 CE (Fig. 4). From the literature dealing with planktonic diatoms, some general characteristics of Lindavia species emerge. These diatoms have a relative heavy frustule with a fast sinking rate (Ptacnik et al. 2003). These characteristics are suitable for living in strong turbulent environments and L. balatonis are considered as indicators of strong mixing of the water column (Rimet et al. 2009). In Sihailongwan, another maar lake on the LGVF located 16 km away from SJML, analyses of sediment trap samples collected between 2003 and 2014 show that L. balatonis flourished in spring and autumn when wind caused strong mixing of the water column (Rioual et al. 2015). Lindavia is heavily silicified, implying that high levels of silicon might be required for rapid growth of this diatom (Bradbury and DieterichRurup 1993). In the European Alps and in Central Europe, L. balatonis has been found in mesotrophic to eutrophic lakes (Houk et al. 2010; Budzyńska et al. 2011), and in the sediment covering the twentieth century, it was found to be related to the beginning of oligotrophication or the start of eutrophication (Alefs and Muller 1999; Hausmann and Kienast 2006), suggesting again a preference for mesotrophic conditions. In EML, strong water mixing can promote increased concentration of nutrients, which is conducive to large populations of L. balatonis in spring (Wang et al. 2012a).
In most maar lakes of the LGVF, the supply of water mainly comes from rainfall and groundwater, and there are no inflows or outflow. Therefore, the main factor affecting the lake turbulence is wind forcing. Wind can also result in large-scale horizontal current pattern, which usually generates gyres on the lake surface, bringing surface water to the shore, while deeper water gushes out at the center of the gyres (McGinnis and Wuest 2005), thus promoting the mixing of the lake’s water-column. Gyres form mainly as a result of the Coriolis effect and non-uniform wind forcing, which is usually observed in large lakes (McGinnis and Wuest 2005). However, in small lakes such as SJML, the formation of gyres is typically due to the non-uniform wind forcing caused by asymmetric topography (Podsetchine and Schernewski 1999).
SJML is a "compound maar lake" formed by two phreatomagmatic eruptions (Liu et al. 2000), and surrounded by relatively high crater rims. The lake is divided into two basins, one large, and one small. A topographic ridge with a height of 40 m and a width of 220 m separates the two basins from the southwest, while on the northeast side the surface water connection between the two basins is 280 m wide (Fig. 1). This strongly asymmetrical lake topography may cause non-uniform wind force. A study from Lake Belau in Germany (Podsetchine and Schernewski 1999), similar in shape to SJML, found that the blocking effect of hills and vegetation around the lake causes spatial variation of wind speed. Under the action of non-uniform wind, a large gyre was formed in the larger basin while a small gyre was formed in the smaller basin (Podsetchine and Schernewski 1999). SJML’s topography may contribute to the mixing of its water column because it likely facilitates the formation of a circulation pattern similar to that prevailing in Lake Belau.
Therefore, the mixing of the upper water column in SJML is very strong which favors the growth of L. balatonis during a large part of the year. This phenomenon was also observed in Yunlong Lake in southern China (Zou et al. 2018). In that lake, strong winds year-round cause the epipelagic zone to be mixed for the entire year and large planktonic diatoms to dominate the assemblages.
Variations in seasonality derived from the diatom record of Lake Sanjiaolongwan
In the last millennium, the relative abundance of planktonic diatoms has changed significantly, while the abundance of benthic diatoms has changed little. However, the small change in benthic diatoms reflects the main climate change that affected the region in that timeframe. Our research area has a humid climate with lager annual variation in temperature and previous studies have shown that the vegetation composition controlled by precipitation did not change substantially in the past 1000 years (Li et al. 2013). This indicates that the biggest climate change in this area should be change in temperature rather than precipitation. Temperature controls the change of the length of ice-cover, and then controls the change of the relative proportion of benthic and planktonic diatoms (Wang et al. 2012a). Ice-cover inhibits the growth of most diatoms, resulting in a decrease in the productivity of both planktonic and benthic diatoms. However, the effect is much more severe on the planktonic taxa, as conditions of very low light and low turbulence prevent the cells from remaining suspended in the water column (Wang et al. 2012a). As observed in high-latitudes and high elevation lakes, ice starts to melt at the shorelines although a long-lasting ice platform may remain in the central part of the lake (Douglas and Smol 1999; Lotter and Bigler 2000). The existence of this ice free moat restricts the development of planktonic diatom communities, while there are still many suitable environments to allow periphyton to develop (Smol 1983). In these lakes with a very long period of ice-cover the sedimentary diatom sequences are therefore largely dominated by benthic species and it is only when the length of the ice-free season lengthens that planktonic taxa can develop larger populations. In mid-latitude lakes, the same phenomenon applies but obviously the planktonic community is less restricted. Thus, the several periods identified in the SJML record of planktonic/benthic diatom abundances most likely correspond to the MCA (SJML-1), the LIA (SJML-2) and the twentieth century warm period (SJML-3 and SJML-4).
The abundance of D. pseudostelligera displayed high frequency oscillations during the MCA (Fig. 4, 980–1310 CE), and was highest in the interval 1020–1060 CE. D. pseudostelligera is a small planktonic species with a very low sinking rate (Ptacnik et al. 2003). Compared with large diatoms affected by sinking, these small diatoms can easily remain suspended in the warm water column below the surface of the lake (Tolotti et al. 2007). Small Discostella species are widely distributed in the world, and are recorded in lake sediments from tropical to high latitudes and polar regions (Köster and Pienitz 2006; Rühland et al. 2003, 2008; Wang et al. 2012a, b; Stewart and Lamoureux 2012; Saros and Anderson 2015; Malik and Saros 2016). Due to its large surface area to volume ratio, a Discostella cell can quickly absorb nutrients, so it has strong competitiveness in oligotrophic environment (Winder and Hunter 2008). The diatom species data from Lake Tahoe in the United States (California-Nevada) that covers a 30-year monitoring period show that climate warming and enhancement of thermal stratification is to the advantage of small-cell Cyclotella [Discostella] species with relatively high surface area to volume ratio (Winder and Hunter 2008; Winder et al. 2009). In Huguangyan Maar Lake in southern China, Discostella species maintain high abundance throughout the year but dominate the assemblages in summer. The seasonal diatom succession in this lake shows that it is controlled by the strong stratification of the water column and the weakening of hydrodynamic forces in summer (Wang et al. 2008, 2012b). Similar observations were made in Lake Sihailongwan in northeastern China, where Discostella are present year-round and have the highest relative abundance in summer when the lake is thermally stratified (Rioual et al. 2015).
During the interval 1020–1060 CE, the abundance of the relatively heavy diatom L. balatonis decreased, and the abundance of the relatively light diatom D. pseudostelligera increased rapidly, indicating that the water column mixing may have weakened. This period of weakened mixing should occur in summer when the water column is thermally stratified. A seasonal survey of diatoms in Lake Huguangyan shows that when the temperature starts to rise in spring, the lake water column begins to stratify and although the wind is still strong at the end of spring, the mixing intensity is significantly reduced by the stratification of the water column, and the abundance of heavy diatoms decreases (Wang et al. 2012b). Thus, the increase in temperature is a very important reason for the decrease of lake hydrodynamic force (Wang et al. 2012b). SJML is strongly stratified only during the seasonal period of high temperature in summer (June to August), when the thermocline is about 10 m deep (Mingram et al. 2004). Outside this time window, SJML’s upper water column has strong mixing conditions, resulting in heavy diatoms like L. balatonis being able to dominate for a long part of the year. However, a rapid rise of temperature in summer undoubtedly results in thermal stratification to occur in the uppermost water column, as observed in all the lakes of the region (Mingram et al. 2004). During the interval 1020–1060 CE, the relative abundance of heavy diatoms L. balatonis decreased significantly and the relatively abundance of the lighter diatom D. pseudostelligera increased rapidly, suggesting that summer thermal stratification during this interval may have been stronger with the mixing periods in spring and autumn shortened.
In the following interval of time, the relative abundance of L. balatonis and D. pseudostelligera showed short-lived oscillations. During the interval 1060–1150 CE and 1210–1270 CE, L. balatonis increased rapidly and regained its dominant position, while D. pseudostelligera decreased correspondingly. This indicates that the mixing intensity of the water column was strengthened, while the stratification intensity was weakened, and implies that the thermal stratification period was shortened in summer and the mixing period was prolonged in spring and autumn. However, during the interval 1150–1210 CE and 1270–1310 CE, L. balatonis decreased again, while D. pseudostelligera showed an increasing trend. In these two intervals, however, the decrease of L. balatonis was not as pronounced as that of the interval 1020–1060 CE, and D. pseudostelligera did not increase so much, which indicates that although the thermal stratification in summer was very strong in these two intervals, it was not stronger than before. The higher oscillation between these two planktonic taxa is also the response to the fluctuation of climate during the MCA (Jones and Mann 2004).
During the LIA (1470–1850 CE), the relative abundance of planktonic diatoms was the lowest of the sequence, indicating that this period was the coldest in the last millennium. There is no obvious short-term oscillation between L. balatonis and D. pseudostelligera. The abundance of L. balatonis remained high, while the abundance of D. pseudostelligera remained at a low value after a large decrease, which indicates that the lake thermal stratification weakened during this period, while the lake mixing was always strong.
For the twentieth century warm period, the assemblages are still dominated by planktonic diatoms and generally indicate a warm environment (Fig. 4). Through most of the twentieth century, D. pseudostelligera increased significantly (1850–1965 CE). During this period, the relative abundance of another planktonic diatom, Fragilaria tenera, also increased significantly. F. tenera has a low sinking rate and can bloom under conditions favoring strong thermal stability (Liu et al. 2017). F. tenera is particularly effective at utilizing phosphorus, but it is less competitive for silicon, and is therefore considered adapted to high Si: P ratio environment (Sommer 1983). F. tenera is most abundant in oligotrophic and mesotrophic lakes (Almeida et al. 2016). The increase of F. tenera in the twentieth century may be due to the increase of nitrogen and phosphorus in the lake caused by human activities. Sedimentary pollen records of SJML show that the relative abundance of Pinus decreased significantly between 1900 and 1950 CE, accompanied by an increase in the abundance of Artemisia and other herbs. From 1930 to 1940 CE, Pinus decreased to the lowest value and Artemisia increased to the highest value, indicating forest degradation caused by human activities (Zhang et al. 2019). Similar pollen changes have been recorded in the sediments from Lake Sihailongwan and EML (Mingram et al. 2004; Li et al. 2013). These pollen records are consistent with the historical records of human activities in northeastern China (Li et al. 2013; Zhang et al. 2019).
It is worth noting that the oligotrophic D. pseudostelligera began to increase in 1850 CE, accompanied by a small increase of mesotrophic F. tenera. However, the relative abundance of F. tenera before 1910 CE was similar to that of the MCA. It is possible that the increase of D. pseudostelligera and F. tenera between 1850 and 1910 CE is mainly related to global warming. It should be pointed out that the diatom flux in this period (Fig. 5) is higher than that in the MCA, which may reflect an increase in nutrients. However, the increase in trophic status did not lead to a complete change in diatom assemblages. Therefore, judging from the increase in the relative percentages of D. pseudostelligera and F. tenera, the thermal stratification of the lake increased during this interval. In addition, the relative percentages of L. balatonis decreased, indicating that lake mixing also weakened.
However, from 1910 to 1965 CE, the relative percentage of oligotrophic D. pseudostelligera decreased (Fig. 4), although its flux did not change significantly (Fig. 5). The relative percentage and flux of mesotrophic F. tenera began to increase significantly, surpassing that observed during the MCA. This may be closely related to the increase in nutrients in this period. Eutrophication was more beneficial to F. tenera than to D. pseudostelligera as we would expect from their trophic requirements.
Since 1965 CE, the relative abundance of small planktonic diatoms Stephanodiscus parvus has increased rapidly, replacing F. tenera and D. pseudostelligra. The total flux of diatoms increased greatly. S. parvus has been reported in many eutrophic lakes (Bradshaw and Anderson 2001; Schönfelder et al. 2002; Dong et al. 2006; Reavie and Kireta 2015). This species prefers higher concentrations of phosphorus and is a good indicator of eutrophication in lakes (Dong et al. 2006). Pollen data from SJML showed that the abundance of Pinus, which had not been restored, decreased a little from 1965 to 1975 CE (Zhang et al. 2019), indicating that human activities and probably nutrient load increased at this period. These changes in nutrients altered the levels of competition between planktonic species, causing a sharp decrease in L. balatonis.
At the same time, S. parvus is also a small (average valve diameter of 5.9 µm) planktonic diatom with strong suspension capacity, so the stratification conditions are also important factors affecting growth rates. Previous studies have found that the smaller the eutrophic diatoms are, the more dominant they are in strongly stratified, high trophic status lakes (Yan et al. 2018). In other words, in mesotrophic and eutrophic lakes with strong water stratification, small-sized eutrophic diatoms predominate (Yan et al. 2018). In a study of Gonghai Lake in north China, it was found that the lake contained a small number of eutrophic species Cyclotella meneghiniana and Stephanodiscus hantzschii due to the relatively high nutrient level of that lake. C. meneghiniana is smaller, ranging from 5 to 15 μm, while S. hantzschii ranges from 15 to 40 μm. From the LIA to the twentieth century warm period, there was no significant change in the nutrient level of the lake, but the relatively large S. hantzschii was replaced by the smaller C. meneghiniana (Yan et al. 2018). In SJML, the relatively large F. tenera was replaced by the small-celled S. parvus as the aquatic ecosystem was affected by global warming. Therefore, the rapid increase of small-sized S. parvus indicates that the water stratification is further strengthened with global warming under high nutrient conditions. At the same time, the mixing of water was also strong, because A. formosa remained relatively abundant in the assemblages in that interval.
There are two main reasons why A. formosa was very abundant in the latter half of the twentieth century. The first is that this species needs high nutrient conditions. A. formosa often occurs in mesotrophic and eutrophic lakes (Hall et al. 1999; Reynolds et al. 2002; Bennion et al. 2011). In Peninsula Lake, Ontario, Canada, with the twentieth century population increase, the trophic status of the lake increased as did the abundance of A. formosa (Clerk et al. 2000). A. formosa usually blooms in spring and autumn, when the lake has high nutrient concentrations, and then its growth decreases as nutrients become limited (Morabito et al. 2002; Ptacnik et al. 2003).
Secondly, it requires high turbulence conditions. Although A. formosa has a high surface area to volume ratio and a relatively low sinking rate (Round et al. 1990), its sinking rate is still higher compared with the very small Discostella species and Stephanodiscus species, especially because it forms star-shaped colonies composed of several frustules. Therefore, A. formosa is abundant in strong water turbulence and is often described as a spring and autumn blooming species (Simola et al. 1990; Ptacnik et al. 2003). For example, in Yunlong Lake in southern China, A. formosa only appears in the cold season, although the upper layer of the lake is mixed strongly all year round. In the cold season, stronger turbulence can provide stronger mixing and more nutrients, which is beneficial to the growth of A. formosa (Zou et al. 2018).
Therefore, during the interval 1965–1975 CE, the rapid increase of the eutrophic species S. parvus indicates that the lake trophic level was relatively high at that time and the lakes had strong thermal stratification. In addition, under the background of the lake eutrophication, lake mixing was also relatively strong in the water column during spring and autumn.
After 1975 CE, A. formosa continued to increase and replaced S. parvus in becoming the dominant species, while L. balatonis also recovered to a relative high abundance. According to recent studies, the rapid increase of A. formosa in numerous lakes around the world is related to the prolonged ice-free season of lakes caused by rapid global warming in the 1980s (Solovieva et al. 2005; Hyatt et al. 2011; Rühland et al. 2010, 2013, 2015; Sivarajah et al. 2016). Although these studies report that the increasing abundance of A. formosa in these lakes is related to the increase of lake stratification caused by global warming, it should be pointed out that most of these are large lakes. Saros et al. (2012) found that with global warming, the mixing depth in large lakes will increase, as will the strength of mixing and the relative abundance of some larger planktonic diatoms. Therefore, whether the blooming of A. formosa in some lakes in the northern hemisphere is related to this mechanism requires more research on the effect of global warming on water column structure in large lakes. The detailed investigation of seasonal diatom succession proposed by Zou et al. (2018) suggests that A. formosa requires strong mixing. Therefore, we suspect that the increasing abundance of A. formosa and L. balatonis in SJML may be due to the shortening of the freezing period of the lake caused by warming, resulting in an increase in the length of spring and autumn when the water column is mixed strongly.
The rapid decrease of S. parvus and of the total diatom flux (Fig. 5), suggest that the lake trophic level decreased after 1975 CE. However, there was no obvious increase of oligotrophic species D. pseudostelligera and mesotrophic species F. tenera, indicating that the lake trophic level did not decrease too much and remained relatively high, still suitable for A. formosa to dominate the assemblages. The decrease in trophic level may be related to the environmental protection measures implemented by the Chinese government from the 1980s. From the pollen data of SJML, we can see that although there was still no obvious recovery of Pinus in the region around 1980 CE, other woody plants such as Quercus, Betula and Ulmus showed an upward trend, reflecting the gradual recovery of the natural environment in the region (Zhang et al. 2019).
Recently, the increase in A. formosa observed in many lakes worldwide has been linked to the increase in atmospheric nitrogen deposition, such as in the alpine lakes of the Rocky Mountains in the western United States (Wolfe et al. 2003; Saros et al. 2005; Hundey et al. 2014). In SJML, when atmospheric nitrogen deposition increased rapidly in the 1980s (Liu et al. 2011), A. formosa also experienced synchronous growth. Therefore, we cannot exclude that the increase of A. formosa during that period may be in part related to the increase in atmospheric nitrogen deposition.
In summary, the rapid increase in the relative abundance of A. formosa after 1975 CE is probably due to the combined effect of the prolonged mixing periods of the lake in spring and autumn and the increase in atmospheric nitrogen deposition. However, it is worth noting that the increase in atmospheric nitrogen deposition is not a pre-condition for increase in the relative abundance of A. formosa (Sivarajah et al. 2016). It only accelerates the development of A. formosa. The more important factors are the extension of the ice-free season of the lake and the change of the lake mixing regime. Hence, as the temperature continues to rise, the lake ice-free period gradually lengthens, which may cause the lake mixing period to extend further in spring and autumn, thus prolonging the growth cycle of A. formosa.
After 1995 CE, the relative abundance and flux of A. formosa and L. balatonis both showed a decreasing trend. We interpret this as a signal that the lake thermal stratification has been further strengthened with the further increase of temperature in this period. Under the conditions of higher nutrient concentrations and enhanced thermal stratification in the lake, mesotrophic species F. tenera and eutrophic species S. parvus began to bloom. At the same time, strong thermal stratification will cause the nutrients in the uppermost water column to be reduced to a certain extent, which is also conducive to the growth of D. pseudostelligera. On the other hand, under the influence of continuous warming, the length of summer thermal stratification increases, resulting in a shortening of the duration of mixing in spring and autumn, thus shortening the growth time of A. formosa and L. balatonis in spring and autumn. If the temperature continues to rise in the future and the lake trophic status remains relatively high, the mesotrophic species F. tenera and the eutrophic species S. parvus should regain their dominance in the diatom assemblages. On the other hand, if the temperature continues to rise but human interference and nutrient concentrations decrease, then the oligotrophic species D. pseudostelligera should become dominant in the assemblage.
Comparison of the relative length of season between the MCA and twentieth century warm period
As mentioned above, planktonic diatoms were more abundant during the MCA and the twentieth century warm period (Fig. 4) with high total diatom fluxes (Fig. 5). However, there are significant differences between the planktonic diatom assemblages in the MCA and the twentieth century warm period, which may be related to human activities. In the MCA, Lake SJML was undisturbed by human beings and was in an oligotrophic state, while by contrast at the beginning of the warm period in the twentieth century, the lake also was affected by human activities. Diatom species such as the mesotrophic Fragilaria tenera rapidly increased and this species shift suggests that the lake’s trophic status in twentieth century was higher than that during the MCA. Pollen records from EML and SJML show that the area was strongly disturbed by human activities in the early twentieth century and a large number of trees were cut (Li et al. 2013; Zhang et al. 2019). The reduction of forest cover likely increased the flux of nutrients to the lake. During rainfall events, nutrients in the soil around the lake could be leached into the lake, resulting in an increase in the nutrient loading in the lake. High trophic level caused diatom bloom, which made the diatom flux in twentieth century warm period higher than that in the MCA.
In addition, the patterns in terms of changes in the relative length of the seasons are also different. During the MCA, there are several short-term oscillations in the changes of thermal stratification in summer and mixing in spring and autumn. By contrast, in twentieth century warm period, the thermal stratification in summer gradually increased, while the mixing in spring and autumn first weakened and then increased. During the warmest period of the MCA, the thermal stratification in summer may have extended to the autumn, which eventually leads to the predominance of small-celled diatoms (D. pseudostelligera) in this period, while the percentages of large diatoms were very low. From this we derive that the warmest period of the MCA (1020–1060 CE) may be the hottest interval in the last millennium.
After 1975 CE, with the continuous increase of temperature, the duration of the ice-free period of the lakes was gradually extended. Although the lake thermal stratification was also strengthened in the summer, these conditions did not extend much into the autumn season, because the relative abundance of A. formosa, which represents the mixing intensity in spring and autumn, was very high in this period. This further indicates that the summer thermal stratification in the recent warm period is still not as long as that inferred for the MCA, whereas the mixing periods in the spring and autumn seasons are longer than those for the MCA. The seasonal variation in the twentieth century is therefore characterized by the autumn being prolonged, which is different from the seasonal characteristics of the MCA when the duration of summer lengthened and that of spring and autumn shortened.
Response of diatoms in Sanjiaolongwan Maar Lake and Erlongwan Maar Lake to warming
This feature of the SJML diatom sequence is remarkably different from that of EML (Wang et al. 2012a). In SJML, L. balatonis dominates the whole sequence (Fig. 4), while in neighboring EML under the same regional conditions, L. balatonis [as Puncticulata praetermissa] only flourished in the early phase of climate warming, reflecting the longer spring and autumn conditions that occurred in those periods (Wang et al. 2012a). This may be related to the different morphological characteristics of the two lakes. As mentioned earlier, the irregular lake topography of SJML can provide strong hydrodynamic forces necessary for L. balatonis. EML, however, has a regular elliptical topography, which results in that the hydrodynamic forces are not as strong as in SJML.
Secondly, the largest difference in diatom assemblages is observed for the later period of the sequences. During the twentieth century warm period, a large number of mesotrophic and eutrophic diatoms, such as F. tenera, S. parvus, and A. formosa, appeared in SJML. However, only the mesotrophic diatom A. formosa showed a temporary increase in EML (Wang et al. 2012a). The appearance of mesotrophic and eutrophic diatoms in the sediment core of the two lakes indicates that the nutrient conditions of the lakes have changed due to human interferences. However, pollen and diatom assemblages show that the degree of disturbance is significantly different between the two lakes: although the trees around EML were cut down, the forest cover has recovered in the latter half of the twentieth century (Li et al. 2013); while in the SJML area the Pinus-dominated forest has not recovered which may be related to the site being close to a town (Zhang et al. 2019).
In spite of the greater human interference in SJML, diatoms have also responded to changes in the natural environment. In the previous discussion, we already mentioned that with increasing trophic status, and with global warming, the dominant diatoms are all small-celled and/or eutrophic. In the diatom sequence of SJML, the change of D. pseudostelligera—F. tenera—S. parvus reflects the common response to human activities and global warming, indicating that the thermal stratification period is also gradually increasing in length while the lake becomes more eutrophic. However, judging from the rapid increase of A. formosa in the later period, global warming has also led to the extension of spring and autumn, which indicates that the thermal stratification at this time is not as strong as that in the warmest period of the MCA. Therefore, compared with the warmest period of the MCA, the seasonal variations of the twentieth century warm period in SJML is characterized by relatively shorter summer and longer spring and autumn, which is consistent with the variations in seasonality recorded in EML (Wang et al. 2012a).
At the top of both diatom records, the mesotrophic diatom A. formosa gradually decreases, while the oligotrophic diatom Discostella species began to increase. This may be related to the strengthening of management in the Longgang volcanic field and the improvement of lake water quality. For that matter, recent water chemistry data show that the nutrient conditions of the two lakes are basically the same (Table 1), and may be moving towards a lower trophic status. In response to global warming, small oligotrophic diatoms have begun to replace mesotrophic and eutrophic diatoms.
Comparison with Northern Hemisphere temperature records
As shown in Fig. 7, the small-celled D. pseudostelligera with a low sinking rate has a highly positive score on PCA axis 1, while L. balatonis with a high sinking rate has a very negative score. Therefore, the PCA axis 1 can represent the gradient of water column stratification/mixing. In addition, as the thermal stratification of the water column in this area is mainly controlled by the change in temperature, PCA axis 1 can be considered to indirectly reflect the change in temperature in the study area.
In Fig. 8, the PCA axis 1 sample scores derived from the SJML record are plotted with the temperature reconstruction curves for China (Yang et al. 2002) and the Northern Hemisphere (Crowley et al. 2000). In zones SJML-1 (a, c), high sample scores on PCA axis 1 reflect strong thermal stratification and indicate a warm period. While in SJML-1b, the scores of PCA axis 1 samples are very low, reflecting a period of cooling. This may correspond to the low temperature anomalies in the middle of the MCA (D' Arrigo et al. 2001; Jones and Mann 2004). In zones SJML-2 (a, b), low PCA axis1 scores correspond with weak thermal stratification and indicate a cold period. At the bottom part of zone SJML-3, high scores on PCA axis 1 indicate a return to warm conditions at 1850 CE. After 1910 CE, however, the PCA Axis 1 sample scores decreased and started to decouple with the temperature curve, which is due to the influence of human activities in the SJML area (Zhang et al. 2019). These three periods correspond well to the MCA, the LIA and the twentieth century warm period. The curve of PCA axis 1 scores broadly matches with the temperature reconstruction of China, and partly fits with the variation of temperature in the Northern Hemisphere.

Comparison of the PCA axis 1 samples scores derived from the diatom data of Sanjiaolongwan Maar Lake; (a) with temperature data for the whole of China (Yang et al. 2002) (b) and the Northern Hemisphere (Crowley et al. 2000) (c) during the past 1000 years. The scores on PCA axis 1 are for the samples from the depth interval 7.5–59-cm
Conclusions
In this study, we discussed the stratification and/or mixing of lake and seasonal variations of climate in the past 1000 years based on the diatom record from SJML in northeastern China. We found that the particular topographic features of SJML may cause the upper water column to remain in a strong mixed state throughout the year, resulting in the assemblage being dominated by large and heavy planktonic diatoms throughout the record. The composition and abundance of diatom assemblages, especially the changes in the relative abundance of planktonic diatoms, indicate that the period between 1020 and 1060 CE was the warmest interval over the past 1000 years. The seasonal lengths of the MCA and the twentieth century warm periods are also different: The summer duration of the warmest period of the MCA was longer than that of the twentieth century warm period, while the lengths of spring and autumn were shorter. This is consistent with seasonal variations recorded by diatoms in EML.
At the same time, seasonal variations are not simply the shortening and/or extending of a certain season. Although the seasonal characteristics of the two warm periods show the extension of summer, the pattern of the extension is different in the MCA and twentieth century. During the warmest period of the MCA, the summer was prolonged while both spring and autumn were shortened. By contrast, in the early phase of global warming at the start of the twentieth century, the duration of autumn is extended first. It is only when global temperature rises further that the length of summer is extended.
Finally, despite the fact that human activities had an impact on SJML during the twentieth century and diatom assemblages changed a lot, the diatom sequence is still recording the most recent global warming, as observed elsewhere in North America and Europe (Rühland et al. 2008). Global warming and human activities both affect the ecological environment and diatoms respond to those changes in different ways. For example, diatoms respond to warming with a shift to planktonic assemblages dominated by smaller species, while under the effect of human activities, diatoms respond to changes in nutrient load through variation in diatom assemblage composition (from oligotrophic to eutrophic diatom species). The results of this study show that the current diatom ecological community characteristics are the result of the combined effects of global warming and human activities.
References
Alefs J, Muller J (1999) Differences in the eutrophication dynamics of Ammersee and Starnberger See (Southern Germany), reflected by the diatom succession in varve-dated sediments. J Paleolimnol 21:395–407
Almeida PD, Morales EA, Wetzel CE, Ector L, Bicudo DDC (2016) Two new diatoms in the genus Fragilaria Lyngbye (Fragilariophyceae) from tropical reservoirs in Brazil and comparison with type material of F. tenera. Phytotaxa 246(3):163–183
Appleby PG (2001) Chronostratigraphic techniques in recent sediments. In: Last WM, Smol JP (eds) Tracking environmental change using lake sediments: basin analysis, coring, and chronological techniques. Kluwer Academic Publishers, Dordrecht, pp 171–203
Appleby PG, Oldfield F (1978) The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210Pb to the sediment. CATENA 5:1–8
Appleby PG, Nolan PJ, Gifford DW, Godfrey MJ, Oldfield F, Anderson NJ, Battarbee RW (1986) 210Pb dating by low background gamma counting. Hydrobiologia 143:21–27
Battarbee RW, Kneen MJ (1982) The use of electronically counted microspheres in absolute diatom analysis. Limnol Oceanogr 27:184–188
Battarbee RW, Jones VJ, Flower BP, Cameron NG, Bennion H, Carvalho L, Juggins S (2001) Diatoms. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments. Kluwer Academic Publishers, Dordrecht, pp 155–201
Bennion H, Simpson GL, Anderson NJ, Clarke G, Dong XH, Hobæk A, Guilizzoni P, Marchetto A, Sayer CD, Thies H, Tolotti M (2011) Defining ecological and chemical reference conditions and restoration targets for nine European lakes. J Paleolimnol 45:415–431
Blaauw M, Christen JA (2011) Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6:457–474
Bopp L, Aumont O, Cadule P, Alvain S, Gehlen M (2005) Response of diatoms distribution to global warming and potential implications: a global model study. Geophys Res Lett 32(19):307–323
ter Braak CJF, Smilauer P (2012) Canoco reference manual and user's guide: software for ordination, version 5.0. Microcomputer Power, Ithaca, New York.
Bradbury JP, Dieterich-Rurup KV (1993) Holocene diatom palelimnology of Elk Lake, Minnesota. Geol Soc Am Spec Pap 276:215–236
Bradshaw EG, Anderson NJ (2001) Validation of a diatom-phosphorus calibration set for Sweden. Freshw Biol 46:1035–1048
Budzyńska A, Wojtal AZ (2011) The centric diatom Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska, comb. nov., in the plankton of eutrophic-hypertrophic Rusałka Lake (Western Poland). Nova Hedwigia 93:509–524
Chang J, Zhang EL, Liu EF, Sun WW, Langdon PG, Shulmeister J (2018) A 2500-year climate and environmental record inferred from subfossil chironomids from Lugu Lake, southwestern China. Hydrobiologia 811:193–206
Chen X, Yang XD, Dong XH, Liu Q (2011) Nutrient dynamics linked to hydrological condition and anthropogenic nutrient loading in Chaohu Lake (southeast China). Hydrobiologia 661:223–234
Chen JH, Chen FH, Feng S, Huang W, Liu JB, Zhou AF (2015) Hydroclimatic changes in China and surroundings during the medieval climate anomaly and little ice age: spatial patterns and possible mechanisms. Quat Sci Rev 107:98–111
Chu G, Sun Q, Wang X, Liu M, Ling Y, Xie M, Shang W, Liu J (2012) Seasonal temperature variability during the past 1600 years recorded in historical documents and varved lake sediment profiles from Northeastern China. Holocene 22:785–792
Clerk S, Hall R, Quinlan R, Smol JP (2000) Quantitative inferences of past hypolimnetic anoxia and nutrient levels from a Canadian Precambrian Shield lake. J Paleolimnol 23:319–336
Cook ER, Esper J, D’Arrigo RD (2004) Extra-tropical Northern Hemisphere land temperature variability over the past 1000 years. Quat Sci Rev 23:2063–2074
Cook ER, Krusic PJ, Anchukaitis KJ, Buckley BM, Nakatsuka T, Sano M (2013) Tree-ring reconstructed summer temperature anomalies for temperate East Asia since 800 C.E. Clim Dynam 41:2957–2972
Crowley TJ (2000) Causes of climate change over the past 1000 years. Science 289:270–277
D’Arrigo R, Jacoby G, Frank D, Pederson N, Cook E, Buckley B, Nachin B, Mijiddorj R, Dugarjav C (2001) 1738 years of Mongolian temperature variability inferred from a tree-ring width chronology of Siberian pine. Geophys Res Lett 28:543–546
D’Arrigo R, Wilson R, Jacoby G (2006) On the long-term context for late twentieth century warming. J Geophys Res 111:D03103. https://doi.org/10.1029/2005JD006352
Davis RB (1987) Paleolimnological diatom studies of acidification of lakes by acid rain: an application of quaternary science. Quat Sci Rev 6:147–163
DeNicola DM (2000) A review of diatoms found in highly acidic environments. Hydrobiologia 433:111–122
Dong XH, Yang XD, Wang R (2006) Diatom indicative species of eutrophication of the lake in the middle and lower reach regions of Yangtze River. Chin Environ Sci 26(5):570–574 (in Chinese)
Dong XH, Bennion H, Battarbee R, Yang XD, Yang HD, Liu EF (2008) Tracking eutrophication in Taihu Lake using the diatom record: potential and problems. J Paleolimnol 40:413–429
Douglas MSV, Smol JP (1999) Freshwater diatoms as indicators of environmental change in the High Artic. In: Stoermer EF, Smol JP (eds) The diatoms: application for the environmental and earth sciences. Cambridge University Press, Cambridge, pp 227–244
Editorial Board for Flora of China (1995) Flora of China, 1st edn. Science Press, Beijing (in Chinese)
Editorial Committee of Vegetation Map of China (2007) Vegetation of China and its geographic pattern-illustration of the vegetation map of the People’s Republic of China (1:1,000,000), 1st edn. Geological Publishing House, Beijing (in Chinese)
Fritz SC (1990) Twentieth-century salinity and water-level fluctuations in Devils Lake, North Dakota: Test of a diatom-based transfer function. Limnol Oceanogr 35(8):1771–1781
Fritz SC, Cumming BF, Gasse F, Laird KR (1999) Diatoms as indicators of hydrologic and climatic change in saline lakes. In: Smol JP, Stoermer EF (eds) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, Cambridge, pp 41–72
Goudeau MLS, Reichart GJ, Wit JC, de Nooijer LJ, Grauel AL, Bernasconi SM, de Lange GJ (2015) Seasonality variations in the Central Mediterranean during climate change events in the Late Holocene. Palaeogeogr Palaeoclimatol Palaeoecol 418:304–318
Guiry MD, Guiry GM (2021) AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org. Searched on 05 July 2021
Hall RI, Leavitt PR, Quinlan R, Dixit AS, Smol JP (1999) Effects of agriculture, urbanization, and climate on waterqualityinthe northern Great Plains. Limnol Oceanogr 44:739–756
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:1–9
Hausmann S, Kienast F (2006) A diatom-inference model for nutrients screened to reduce the influence of background variables: application to varved sediments of Greifensee and evaluation with measured data. Palaeogeogr Palaeoclimatol Palaeoecol 233:96–112
Houk V, Klee R, Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions Part III. Stephanodiscaceae A: Cyclotella, Tertiarus, Discostella. In: Poulíčková A (ed) Fottea 10 (Supplement):1–498
Hundey EJ, Moser KA, Longstaffe FJ, Michelutti N, Hladyniuk R (2014) Recent changes in production in oligotrophic Uinta Mountain lakes, Utah, identified using paleolimnology. Limnol Oceanogr 59:1987–2001
Hyatt CV, Paterson AM, Rühland KM, Smol JP (2011) Examining twentieth century water quality and ecological changes in the Lake of the Woods, Ontario, Canada: a paleolimnological investigation. J Gt Lakes Res 37(3):456–469
Jan E, Edward RC, Fritz HS (2002) Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 295:2250–2253
Jha SK, Chavan SB, Pandit GG, Sadasivan S (2003) Geochronology of Pb and Hg pollution in a coastal marine environment using global fallout. J Environ Radioact 69:145–157
Jones PD, Mann ME (2004) Climate over past millennia. Rev Geophys 42:1–42
Juggins S (2007) C2 Version 1.5: Software for ecological and palaeoecological data analysis and visualisation. Newcastle University, Newcastle upon Tyne, UK
Köster D, Pienitz R (2006) Seasonal diatom variability and paleolimnological inferences—a case study. J Paleolimnol 35:395–416
Krammer K, Lange-Bertalot H (1986–1991) Bacillariophyceae. Süsswasserflora von Mitteleuropa, Band 2. Spektrum Akademischer Verlag Heidelberg, Berlin
Larocque I, Grosjean M, Heiri O, Trachsel M, Kamenik C (2010) Thousand years of climate change reconstructed from chironomid subfossils preserved in varved Lake Silvaplana, Engadine, Switzerland. Quat Sci Rev 29:1940–1949
Li J, Mackay AW, Zhang Y, Li JJ (2013) A 1000-year record of vegetation change and wildfire from Maar Lake Erlongwan in northeast China. Quat Int 290–291:313–321
Li JJ, Wang L, Cao Q, Rioual P, Lei GL, Cai BG, Zhang JY, Zou YF, Yan Y, Wan XQ, Xiao JL (2021) Diatom response to global warming in Lake Douhu, Southeast China. Acta Geol Sin-Engl. https://doi.org/10.1111/1755-6724.14294
Liu MY (1989) Huinan county annals. Shenzhen Haitian Press, Shenzhen (in Chinese)
Liu JQ, Negendank JFW, Wang WY, Chu GQ, Mingram J, Guo ZF, Luo XJ, Chen R, Liu DS (2000) The distribution and geological characteristics of maar lakes in China. Quat Sci 20:78–86 (in Chinese)
Liu Z, Henderson ACG, Huang Y (2006) Alkenone-based reconstruction of late-Holocene surface temperature and salinity changes in Lake Qinghai, China. Geophys Res Lett 33:1–4
Liu X, Duan L, Mo JM, Du EZ, Shen JL, Lu XK, Zhang Y, Zhou XB, He CN, Zhang FS (2011) Nitrogen deposition and its ecological impact in China: an overview. Environ Pollut 159:2251–2264
Liu YY, Chen GJ, Shi HB, Chen XL, Lu HB, Duan LZ, Zhang HC, Zhang WX (2016) Responses of a diatom community to human activities and climate change in Xingyun Lake. Acta Geol Sin-Engl 36(10):3063–3073 (in Chinese)
Liu JB, Rühland KM, Chen JH, Xu YY, Chen SQ, Chen QM, Huang W, Xu QH, Chen FH, Smol JP (2017) Aerosol-weakened summer monsoons decrease lake fertilization on the Chinese Loess Plateau. Nat Clim Chang 7:190–194
Lotter AF, Bigler C (2000) Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat Sci 62:125–141
Ma RY, Ma DZ (1990) Research on frontier development in Qing dynasty, 1st edn. Science China Press, Beijing (in Chinese)
Malik H, Saros JE (2016) Effects of temperature, light and nutrients on five Cyclotella sensu lato taxa assessed with in situ experiments in Arctic lakes. J Plankton Res 38:431–442
McGinnis D, Wuest A (2005) Lake hydrodynamics. McGraw-Hill Companies Publisher, Mcgraw Hill Yearbook of Science and Technology
Meteorological data of China (1981–2010) Cumulative average monthly meteorological data from Jing Yu, China, 1981–2010. http://data.cma.cn/dataService/cdcindex/datacode/A.0029.0004/show_value/normal.html. Accessed 23 June 2020
Mingram J, Allen JRM, Brüchmann C, Liu J, Luo X, Negendank JFW, Nowaczyk N, Schettler G (2004) Maar—and crater lakes of the Long Gang Volcanic Field (N. E. China)—overview, laminated sediments, and vegetation history of the last 900 years. Quat Int 123–125:135–147
Morabito G, Ruggiu D, Panzani P (2002) Recent dynamics (1995–1999) of the phytoplankton assemblages in Lago Maggiore as a basic tool for defining association patterns in the Italian deep lakes. J Limnol 61:129–145
Orlowsky B, Seneviratne SI (2012) Global changes in extreme events: regional and seasonal dimension. Clim Chang 110:669–696
Phlips EJ, Hendrickson J, Quinlan EL, Cichra MF (2007) Meteorological influences on algal bloom potential in a nutrient-rich blackwater river. Freshw Biol 52:2141–2155
Podsetchine V, Schernewski G (1999) The influence of spatial wind homogeneity on flow patterns in a small lake. Water Res 33:3348–3356
Ptacnik R, Diehl S, Berger S (2003) Performance of sinking and nonsinking phytoplankton taxa in a gradient of mixing depths. Limnol Oceanogr 48:1903–1912
Reavie ED, Kireta AR (2015) Centric, araphid and eunotioid diatoms of the coastal Laurentian great lakes. Bibl Diatomol 62:1–184
Reynolds CS, Huszar V, Kruk C, Naselli-Flores L, Melo S (2002) Towards a functional classification of the freshwater plankton. J Plankton Res 24:417–428
Rimet F, Druart JC, Anneville O (2009) Exploring the dynamics of plankton diatom communities in Lake Geneva using emergent self-organizing maps (1974–2007). Ecol Inform 4:99–110
Rioual P, Chu GQ, Mingram J, Han J, Liu J (2015) Diatom seasonality assessed from sediment traps and implications for the interpretation of an annually laminated sedimentary record from China. ASLO 2015 Aquatic Sciences Meeting, Granada, Spain. http://www.sgmeet.com/aslo/granada2015/viewabstract.asp?AbstractID=26463
Round FE, Crawford RM, Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge
Rühland K, Priesnitz A, Smol JP (2003) Paleolimnological evidence from diatoms for recent environmental changes in 50 Lakes across Canadian Arctic Treeline. Arct Antarct Alp Res 35:110–123
Rühland K, Paterson AM, Smol JP (2008) Hemispheric-scale patterns of climate related shifts in planktonic diatoms from North American and European lakes. Glob Chang Biol 14:2740–2754
Rühland KM, Paterson AM, Hargan K, Jenkin A, Clark BJ, Smol JP (2010) Reorganization of algal communities in the lake of the woods (Ontario, Canada) in response to turn-of-the-century damming and recent warming. Limnol Oceanogr 55:2433–2451
Rühland KM, Paterson AM, Keller W, Michelutti N, Smol JP (2013) Global warming triggers the loss of a key Arctic refugium. Proc R Soc B. https://doi.org/10.1098/rspb.2013.1887
Rühland KM, Paterson AM, Smol JP (2015) Lake diatom responses to warming: reviewing the evidence. J Paleolimnol 54:1–35
Saros JE, Anderson NJ (2015) The ecology of the planktonic diatom Cyclotella and its implications for global environmental change studies. Biol Rev 90:522–541
Saros JE, Michel TJ, Interlandi SJ, Wolfe AP (2005) Resource requirements of Asterionella formosa and Fragilaria crotonensis in oligotrophic alpine lakes: implications for recent phytoplankton community reorganizations. Can J Fish Aquat Sci 62(7):1681–1689
Saros JE, Stone JR, Pederson GT, Slemmons KEH, Spanbauer T, Schliep A, Cahl D, Williamson CE, Engstrom DR (2012) Climate-induced changes in lake ecosystem structure inferred from coupled neo-and paleoecologcial approaches. Ecology 93:2155–2164
Schönfelder I, Gelbrecht J, Schönfelder J, Steinberg CEW (2002) Relationships between littoral diatoms and their chemical environment in Northeastern German lakes and rivers. J Appl Phycol 38:66–82
Simola H, Hanski I, Liukkonean M (1990) Stratigraphy, species richness and seasonal dynamics of plankton diatoms during 418 years in Lake Lovojarvi, south Finland. Ann Bot Fenn 27(3):241–259
Sivarajah B, Rühland KM, Labaj AL, Paterson AM, Smol JP (2016) Why is the relative abundance of Asterionella formosa increasing in a Boreal Shield lake as nutrient levels decline? J Paleolimnol 55:357–367
Smol JP (1983) Paleophycology of a high arctic lake near Cape Herschel, Ellesmere Island. Can J Bot 61:2195–2204
Smol JP, Douglas MSV (2007) From controversy to consensus: making the case for recent climate change in the Arctic using lake sediments. Front Ecol Environ 5(9):466–474
Smol JP, Wolfe AP, Birks HJB, Douglas MS, Jones VJ, Korhola A, Pienitz R, Rühland K, Sorvari S, Antoniades D, Brooks SJ, Fallu MA, Mike H, Keatley BE, Laing TE, Michelutti N, Nazarova L, Nyman M, Paterson AM, Perren B, Quinlan R, Rautio M, Talbot ES, Siitonen S, Solovieva N, Wechström J (2005) Climate-driven regime shifts in the biological communities of arctic lakes. Proc Natl Acad Sci 102:4392–4402
Solovieva N, Jones VJ, Nazarova L, Brooks SJ, Birks HJB, Grytnes JA, Appleby PG, Kauppila T, Kondratenok B, Renberg I, Ponomarev V (2005) Palaeolimnological evidence for recent climatic change in lakes from the northern Urals, Arctic Russia. J Paleolimnol 33:463–482
Sommer U (1983) Nutrient competition between phytoplankton species in multispecies chemostat experiments. Arch Hydrobiol 96:399–416
Srifa A, Phlips EJ, Hendrickson J (2016) How many seasons are there in a sub-tropical lake? A multivariate statistical approach to determine seasonality and its application to phytoplankton dynamics. Limnologica 60:39–50
Stewart KA, Lamoureux SF (2012) Seasonal and microhabitat influences on diatom assemblages and their representation in sediment traps and surface sediments from adjacent high Arctic lakes: Cape Bounty, Melville Island, Nunavut. Hydrobiologia 683:265–286
Tan M, Liu TS, Hou JZ, Qin XG, Zhang HG, Li TY (2003) Cyclic rapid warming on centennial-scale revealed by a 2650-year stalagmite record of warm season temperature. Geophys Res Lett. https://doi.org/10.1029/2003GL017352
Tolotti M, Corradini F, Boscaini A, Calliari D (2007) Weather-driven ecology of planktonic diatoms in Lake Tovel (Trentino, Italy). Hydrobiologia 578:147–156
Wang L, Lu HY, Liu JQ, Gu ZY, Mingram J, Chu GQ, Li JJ, Rioual P, Negendank JFW, Han JT, Liu TS (2008) Diatom-based inference of variations in the strength of Asian winter monsoon winds between 17,500 and 6000 calendar years B.P. J Geophys Res 113:1–9. https://doi.org/10.1029/2008JD010145
Wang L, Rioual P, Panizzo VN, Lu HY, Gu ZY, Chu GQ, Yang DG, Han JT, Liu JQ, Mackay AW (2012a) A 1000-yr record of environmental change in NE China indicated by diatom assemblages from Maar Lake Erlongwan. Quat Res 78:24–34. https://doi.org/10.1016/j.yqres.2012.03.006
Wang L, Li JJ, Lu HY, Gu ZY, Rioual P, Hao QZ, Mackay AW, Jiang WY, Cai BG, Xu B, Han JT, Chu GQ (2012b) The East-Asian winter monsoon over the last 15,000 years: its links to high-latitudes and tropical climate systems and complex correlation to the summer monsoon. Quat Sci Rev 32:131–142. https://doi.org/10.1016/j.quascirev.2011.11.003
Wang L, Jiang WY, Jiang DB, Zou YF, Liu YY, Zhang EL, Hao QZ, Zhang DG, Zhang DT, Peng ZY, Xu B, Yang XD, Lu HY (2018) Prolonged heavy snowfall during the younger Dryas. J Geophys Res-Atmos 123(13):748–762. https://doi.org/10.1029/2018JD029271
Williamson CE, Saro JE, Vincent WF (2009) Lakes and reservoirs as sentinels integrators and regulators of climate change. Limnol Oceanogr 54(6):2273–2282
Winder M, Hunter DA (2008) Temporal organization of phytoplankton communities linked to physical forcing. Oecologia 156:179–192
Winder M, Sommer U (2012) Phytoplankton response to a changing climate. Hydrobiologia 698:5–16
Winder M, Reuter JE, Schadlow SG (2009) Lake warming favours small-sized planktonic diatom species. Proc R Soc B 276:427–435
Wolfe AP, Grop ACV, Baron JS (2003) Recent ecological and biogeochemical changes in alpine lakes of Rocky Mountain National Park (Colorado, USA): a response to anthropogenic nitrogen deposition. Geobiology 1:153–168
Yan Z, Xia J, Qian C, Zhou W (2011) Changes in seasonal cycle and extremes in China during the period 1960–2008. Adv Atmos Sci 28:269–283
Yan Y, Wang L, Li J, Li JJ, Zou YF, Zhang JY, Li P, Liu Y, Xu B, Gu ZY, Wan X (2018) Diatom response to climatic warming over the last 200 years: a record from Gonghai Lake, North China. Palaeogeogr Palaeoclimatol Palaeoecol 495:48–59
Yang B, Braeuning A, Johnson KR, Shi YF (2002) General characteristics of temperature variation in China during the last two millennia. Geophys Res Lett 29:1324
Yang XD, Kamenik C, Schmidt R, Wang SM (2003) Diatom-based conductivity and water-level inference models from eastern Tibetan (Qinghai-Xizang) Plateau lakes. J Paleolimnol 30:1–19
Yang XD, Shen J, Dong XH, Liu EF, Wang SM (2006) Historical trophic evolutions and their ecological responses from shallow lakes in the middle and lower reaches of the Yangtze River: case studies on Longgan Lake and Taibai Lake. Sci China Ser D 49(Suppl I):51–61
Yang XD, Anderson NJ, Dong XH, Shen J (2008) Surface sediment diatom assemblages and epilimnetic total phosphorus in large, shallow lakes of the Yangtze floodplain: their relationships and implications for assessing long-term eutrophication. Freshw Biol 53:1273–1290
Zhang Y, Tian Q, Gou X, Chen F, Leavitt S, Wang Y (2011) Annual precipitation reconstruction since AD 775 based on tree rings from the Qilian Mountains, northwestern China. Int J Climatol 31:371–381
Zhang JY, Li J, Yan Y, Li JJ, Wan XQ (2019) A 1000-year record of centennial-scale cyclical vegetation change from Maar Lake Sanjiaolongwan in northeastern China. J Asian Earth Sci 176:315–324
Zolitschka B, Brauer A, Negendank JFW, Stockhausen H, Lang A (2000) Annually dated late Weichselian continental paleoclimate record from the Eifel, Germany. Geology 28:783–786
Zou YF, Wang L, Zhang LZ, Liu Y, Li P, Peng ZY, Yan Y, Zhang JY, Lu HY (2018) Seasonal diatom variability of Yunlong Lake, southwest China—a case study based on sediment trap records. Diatom Res 33:381–396
Acknowledgements
This work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (Grant XDB26000000), the National Key R&D Program of China (Grant 2017YFA0603400), and the National Natural Science Foundation of China (Grants 41772379 and 41372372).
Funding
Strategic Priority Research Program of Chinese Academy of Sciences, XDB26000000, Luo Wang, National Key R&D Program of China, 2017YFA0603400, Luo Wang, National Natural Science Foundation of China, 41772379, Luo Wang, 41372372, Luo Wang
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Luo, H., Li, J., Li, P. et al. Variation in the seasonal response to climate change during the past 1000 years as inferred from a Maar Lake sediment record, northeast China. J Paleolimnol 68, 133–154 (2022). https://doi.org/10.1007/s10933-021-00228-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-021-00228-3