
Abstract
A multi-proxy analysis, including pollen, non-pollen palynomorphs and plant macrofossil remains was performed to reconstruct the paleoenvironmental history of the Cabeza de Buey shallow lake (36°17′S, 61°10′W) over the last 600 years. We identified the main forcing factors behind changes in the structure and dynamics of the lake communities. Given the relatively low intensity of human activity during the early period of the lake history (prior to 1880 AD), changes in dominant communities were mainly controlled by climate. Afterwards, changes probably resulted from a combination of climate and human impacts that generated an accelerated eutrophication. Four periods in the lake’s evolution were recognized based on changes in macrophyte and algae communities’ structure. Between ca. 1320 and ca. 1630 AD a shift was evidenced from an oligotrophic brackish with low nutrient content (dominated by Botryococcus braunii) to a mesotrophic fresh water body (dominated by Pediastrum and Scenedesmus), due to an increase in the water level associated with an increase in precipitation. At ca. 1840 AD a transition towards a phase dominated by Myriophyllum spicatum was noted, induced by a low water level as a consequence of low precipitation. Later, at ca. 1880 AD, the current lake conditions were established, and the increase in nutrient and organic matter supply influenced plant community structure towards organic tolerant species of submerged macrophytes (Potamogeton pectinatus). Towards the beginning of the twenty-first century, the lake turned to a more eutrophic state, which is evidenced by the dominance of the phytoplankton community. The last hundred years of the lake history were characterized by the eutrophication process related to the impact of agriculture and cattle breeding implemented in the landscape as well as to the urban settlement. This study made it possible to infer changes in the structure and dynamics of Cabeza de Buey lake and to elucidate the forcing factors that induced these changes on a high-resolution scale.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
A large number of lakes are found in the Pampa plain of Argentina, in the southeast of South America. According to Geraldi et al. (2011) there are approximately 13,000 lakes with surface areas up to 10 ha. These lakes are rich in nutrients in response to anthropogenic activities such as intensive agriculture, livestock farming and non-regulated urbanization (Quirós et al. 2006). One of the characteristics of these lakes is the alternation into two different states, often with transition periods between them. The clear state is dominated by submerged macrophytes, while phytoplankton biomass is low. The turbid state exhibits high phytoplankton biomass and scarce macrophyte development. The shift between these states can be associated with variations in nutrient loading, changes in water level, seasonality, removal of submerged macrophytes and fish introduction, among other factors (Allende et al. 2009; O’Farrell et al. 2011; Mancini et al. 2013; Sánchez et al. 2015; Cano et al. 2016). Even though these lakes provide numerous ecological services and are important reservoirs of biological diversity and fresh water, only a few studies have evaluated their current functioning and even fewer monitoring programs have accounted for their evolution. The Holocene environmental history of Pampean shallow lakes has been elucidated by some authors (Borel et al. 2003; Fontana 2005; Laprida et al. 2014; Piovano et al. 2009; Stutz et al. 2010, 2012, 2014; Vilanova et al. 2015, among others). However, high-resolution and multi-indicator analyses on decadal-centennial scale for the late-Late Holocene are still scarce. Laprida and Valero Garcés (2009) and Laprida et al. (2009) used ostracods to infer climatic events during the last 500 years in Chascomús and del Monte lakes. In del Monte and Melincué lakes, Córdoba et al. (2014) and Guerra et al. (2015) presented environmental reconstructions based on physical and geochemical proxies with a detailed high-resolution analysis for the last 100 years.
In particular, the reconstruction of lakes and wetlands during the last 600 years is crucial due to the impact of anthropogenic pressure over these ecosystems since the European arrival (ca. 1540 AD) to the Pampa plain. Vegetation has been subject to different intensities of livestock grazing over the last four centuries, while intensive farming and urbanization have taken place during the last 120 years. In addition, farming has been associated with an increase in the use of agrochemicals, which led to the consequent degradation of the aquatic environments and acceleration of eutrophication processes. Landscape development and transformation are reported in historical records and chronicles after European settlement (Vervoorst 1967; Garavaglia 1999). This information can be compared with palaeoecological reconstructions, enabling the discrimination of climatic and anthropogenic forcings (Kowalewski et al. 2016; Rull et al. 2017, among others). Therefore, it is essential to gain a better understanding of the lake community structure and dynamics and responses to short-term environmental changes, as well as to establish their reference conditions, to lead their management in the face of growing anthropic pressure. These considerations are important for setting conservation objectives, in particular where the reference conditions of a lake are in question (Bennion et al. 2017).
In order to obtain long-term and high-resolution information, multi-proxy analyses are necessary to record changes occurring over decades and centuries and to reconstruct the evolution of aquatic systems as well as to evaluate their responses to environmental changes. In order to reconstruct the environmental history of Cabeza de Buey, a Pampean shallow lake, we analyzed changes in lake community structure and dynamics on high-resolution scale in a sediment record spanning the last 600 years, since ca. 1320 AD to present. We carried out multi-proxy analysis, including pollen, non-pollen palynomorphs, plant macrofossil remains, organic matter and carbonate content and sedimentology.
Study area
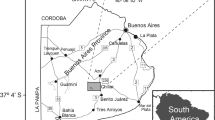
The study site (Cabeza de Buey) is located in the central Pampa plain of Argentina, near San Carlos de Bolivar city (Fig. 1). The area has a poorly integrated drainage system with two main streams, Las Flores and Vallimanca, flowing to the northeast. The region is characterized by the presence of several lakes of variable size, which have an aeolian origin formed on interdunes or on deflation basins. These lakes are associated with parabolic and blowout dunes. These paleodunes were developed by prevailing westerly-southwesterly winds during the Late Pleistocene (Zárate and Tripaldi 2012; Tripaldi and Zárate 2016). Cabeza de Buey (36°17′S, 61°10′W) is a small freshwater shallow lake (0.7 km2 surface, up to 1 m deep) and is related to the deflation hollow of a parabolic dune. Even though it is a permanent lake, it experiences water-level reductions during episodic droughts. Historical chronicles mention that in 1760 and in 1876 the water level decreased markedly, which was associated with drought episodes (Martinez 1967). During this century, after the 2008–2009 drought, a marked decrease in the water level was recorded by Sanzano et al. (2014) for Blanca Chica shallow lake, which is located 100 km from Cabeza de Buey (Fig. 1).

a. Location map of study area in Pampean Grasslands. b. Location of cores extraction in Cabeza de Buey lake. c. Image of Cabeza de Buey lake during core extraction (2013)
The potential vegetation of the Pampa plain is a grassland subdivided in different vegetation units (Fig. 1). Cabeza de Buey lake is located in the Inland Pampa, where the vegetation is dominated by grasses such as Andropogon pellitus (Hack.) Herter, Elionurus muticus (Spreng.) Kuntze and several species of the genus Stipa, accompanied by species of the genus Aristida, Bothriochloa, Chloris, Eragrostis, Poa, Bromus, Digitaria and Piptochaetium. There are also associated annual herbs corresponding to the Asteraceae family (León 1991). The lake plant community is characterized by scarce Schoenoplectus californicus (C.A. Mey.) Soják patches around the lakeshore. Among the submerged vegetation, Potamogeton pectinatus L. develops both in the deepest part of the lake as well as in the shoreline areas. Myriophyllum sp. is found in low abundance. At the study site, several Eucalyptus sp. trees grow on the parabolic dune next to the lake (Fig. 1).
The climate is temperate with annual mean temperature and precipitation of 15.5 °C and 1061 mm, respectively. Mean temperature varies from 22.7 °C in January to 8.4 °C in July. Rainfall occurs mainly from spring (September) to autumn (March), with a maximum of 161.8 mm in March and a minimum of 32.5 mm in July (Estación Meteorológica Aeródromo de S.C. de Bolivar, 36°12S, 61°00 W, Servicio Meteorológico Nacional). Summer rains are related to a movement of the Intertropical Convergence Zone and the semipermanent South Atlantic and South Pacific anticyclones to the south, and to an intensified low-level jet in the Chaco region. The convective activity related to frontal systems prevails during winter, being the South Atlantic anticyclone the main source of moisture (Labraga et al. 2002). In addition to this general pattern of precipitation, another source of humidity is associated to the frontal polar irruptions from the south that oppose to the displacement of humid air masses generating frontal and pre-frontal rains. According to San Carlos de Bolivar instrumental precipitation record (Instituto Nacional de Tecnología Agropecuaria, INTA), the area was characterized by low rainfall and dry periods from 1901 to 1970. In the last 50 years, there have been important changes in the rainfall spatial distribution due to an increase in precipitation values. This change to more humid conditions known as Salto Climático (Agosta and Compagnucci 2008), has affected the Pampa plain (Piovano et al. 2002; Córdoba et al. 2014).
Methods and materials
Two sediment cores, Cabeza de Buey 1, CB1 (36°17′18″S; 61°10′ 57″W), and Cabeza de Buey 3, CB3 (36°17′17″S; 61°10′ 54″W), were recovered in May and December 2013 respectively, with a vibracorer equipped with a 5-cm inner diameter tube. The vibracorer was operated from a platform with a lake depth of 0.85 m in both cases, and core lengths were 113 cm for CB1 and 215 cm for CB3. Chronology was determined using radiocarbon dating measured on bulk sediment by means of an AMS spectrometer at DirectAMS (Radiocarbon Dating Service) and at Accelerator Mass Spectrometry Laboratory (University of Arizona). The age-depth model was constructed based on six radiocarbon dates on CB1 and CB3 cores (Table 1) and additional information related to known events was obtained using CLAM 2.2 (Blaauw 2010), a code written in R language (R CoreTeam 2014). Radiocarbon ages were calibrated with the Southern Hemisphere calibration curve (SHCal13, Hogg et al. 2013) and smooth spline regressions with smoothing level of 0.3 were applied. In addition, ages associated to known events were incorporated to the age-depth model to refine the chronology. The first appearance of Eucalyptus pollen at 28-cm depth was assigned to an age of 1880 AD, following the introduction of Eucalyptus seeds into the country by 1858 AD. An age of 1880 AD was assumed as the time of pollen deposition based on the time of growth, flowering and dispersion of the tree, as well as the founding and demographic growth of towns of the Pampa plain. This inference was based on the historical chronicles, the autoecology of the genus and the presence of this pollen grain in several lacustrine records of the Pampa plain (Veervorst 1967; Sanchez Vuichard 2019). The tephra layer deposited at 109–118-cm depth corresponded to the Alpehue pumice eruption of the Sollipulli volcano (Southern Volcanic Zone of the Andes), dated to 2951 cal years BP (Fontijn et al. 2016). The chemical composition of this tephra was analyzed on two samples by Dr. Fontijn at the Department of Earth Sciences, University of Oxford (United Kingdom). The top sample of the core was assigned to the year of core extraction, 2013. No reservoir effect was considered for this study since the vast majority of shallow Pampean lakes possess soft water, meaning that contributions of ancient dissolved carbonate to the water and sediments are likely to be low (Plastani et al. 2019). For the present work the uppermost 70 cm of core CB1 are considered, which, according to the age-depth model, span the time between ca.1320 and 2013 years AD (Fig. 2).

a. Age-depth model for Cabeza de Buey lake, based on radiocarbon dating. The blue dots are the radiocarbon dates and the green dots represent the additional information dates used in the construction of the model. The gray horizontal stripe represents the ash level. The red box indicates the time interval included in this study. Figure presented in Messineo et al. (2019). b. Detail of the age-depth model for the first 70 cm
The stratigraphy of the cores was characterized by granulometry, color and sedimentary structure descriptions (Messineo et al. 2019). Organic matter content was estimated on CB1 and CB3 cores and carbonates content on CB1 every 2 cm by weight loss-on-ignition (LOI). Samples were dried at 105 °C for 24 h, and then ignited at 550 C for 4 h and finally at 900 °C for 2 h. The weight loss between each step measures the water, the organic matter and carbonates content of the sediment. Results are expressed as a percentage of weight loss of the sediment related to the dry weight of the samples at 105 °C before combustion (Heiri et al. 2001). The correlation of the cores was based on the described stratigraphic units, the organic matter content, and the occurrence of the tephra layer. Three apparent features in both cores were used as specific tie points: first, the minimum values of organic matter (≥ 5%) that coincided with the change from U7 to U6; second, the rapid increase in organic matter values (from 6 to 18%) coincident with the transition from U6 to U5; and third, the drop of organic matter values which in turn coincided with the change from U4 to U3 (Fig. 3). This correlation allowed unifying the radiocarbon dates obtained from both cores in a single temporal profile, and thus establishing a reliable age-depth model.

Modified from Messineo et al. (2019)
a. Sedimentological units and descriptions for CB1 and CB3. Red stars represent radiocarbon dates on both cores. b. Organic matter profiles for CB1 and CB3 and carbonate content profile. Red circles represent tie points for cores correlation.
The core CB1 was subsampled at 1-cm intervals, the analysis of pollen and non-pollen palynomorphs (NPP), and plant macrofossil remains were performed at 2-cm intervals. Samples for pollen and non-pollen palynomorphs (NPP) analysis were prepared following standard techniques, using warm KOH, HCl, heavy-liquid separation with ZnCl2, HF and acetolysis (Bennett and Willis 2001). Two Lycopodium clavatum L. tablets were added before treatment. Pollen sums varied between 500 and 1000 pollen grains. Each pollen type was calculated as a percentage of the total pollen sum (excluding exotic and long distance tree pollen). NPP, pollen of exotic trees as well as long distance tree pollen were calculated as a percentage of the pollen sum plus the sum of each group of taxa. Percentages of Azolla filiculoides Lam., Ricciocarpos natans (L.) Corda and Bryophyta spores were calculated as a percentage of the pollen sum plus the spores sum. Exotic trees include Eucalyptus, Casuarina, Betula, Cupressus, Corylus and Juglans and genera of Pinaceae family (Pinus, Cedrus, Abies). Long distance trees include Celtis ehrenbergiana (Klotzsch) Liebm, Schinus, Alnus, Podocarpus and Nothofagus dombeyi (Mirb.) Oerst. type. Other herbs include Chrysanthemum, Rubiaceae, Lamiaceae, Papilionoideae, Solanaceae, Verbenaceae, Rutaceae, Euphorbiaceae, Caryophyllaceae, Ephedra, Rosaceae, Erodium, Onagraceae, Malvaceae, Geraniaceae, Scrophullariaceae and Monocotyledoneae. The identification of pollen grains, spores and NPP were made with reference to atlases, published keys and the reference collection of the Laboratory of Paleoecology and Palynology, IIMyC CONICET-UNMdP.
Samples for plant macrofossil analysis were treated with 10% KOH for 12 h and sieved through 200 and 100 μm meshes. The volume of each sample was calculated by water displacement. Sample volume varied between 9 and 27 cm3. The residue was examined under a stereo-microscope at 50 × magnification. Recognizable plant macro remains were isolated, identified and counted. Results were expressed in number of remains per 10 cm3 of sediment. According to Sayer et al. (2010), an alternative way of gaining information about past plant abundances is to use persistent fossils of animals that have a close connection with submerged macrophytes, such as the plant-attached egg cocoons of fish leech and mollusks shells, which are closely associated to macrophytes for habitat and grazing periphyton. As such, fossil cocoons of fish leech Piscicola geometra (Linnaeus 1761) (Fam: Piscicolidae, Phylum: Annelida) and carbonatic shells of mollusk Biomphalaria peregrina (d`Orbigny 1835) (Fam: Planorbidae, Phylum: Mollusca) were counted.
Stratigraphic diagrams were plotted with Tilia 2.0.41 (Grimm 2015). The samples were classified using a restricted clustering analysis using the Edwards and Cavalli-Sforza chord distance as a measure of dissimilarity. Cluster analysis was performed with CONISS software included in the Tilia-Graph package (Grimm 2015). All taxa were considered for the palynomorphs (pollen and NPP) zonation, and for macrofossil remains plus mollusk and annelid zonation. CONISS cluster dendrograms are displayed on the right side of the diagrams, showing the zones (Figs. 4, 5).

Pollen and NPPs diagram for CB1. Organic matter content and Cluster Analysis are plotted. Bold horizontal lines correspond to the pollen zone divisions. Zones were defined using CONISS

Plant macrofossil remains and associated fauna diagram. Organic matter content and Cluster Analysis are plotted. Bold horizontal lines correspond to the plant macrofossil zone divisions. Zones were defined using CONISS
Vascular plant nomenclature follows the database Flora del Conosur, Catálogo de las Plantas Vasculares published online by the Instituto de Botánica Darwinion (http://www.darwin.edu.ar/Proyectos/FloraArgentina/fa.htm).
Results
Organic matter and carbonate content
The organic matter content increased gradually towards the top of the sedimentary record, with higher values among the modern samples (Fig. 3). Between 70–50 cm the lowest values (≤ 7%) were observed, between 50–20 cm the values increased slightly (≤ 9%), and from 20 cm to the top the highest values were found (≤ 15%). The carbonate content showed a contrary trend to the organic matter content. Between 70–50 cm values decreased from ≤ 7.5% to 3.5%, between 50–20 cm values ranged between 4.5%-7.5%, and from 20 cm to the top values decreased to 5.5%.
Pollen, NPP and plant macrofossil remains
According to the cluster analysis, four pollen and NPP assemblage zones (PAZ 1–4) (Fig. 4) and two plant macrofossil assemblage zones (MAZ 1–2) (Fig. 5) were established. The three oldest pollen and NPP assemblage zones, PAZ 1–3, corresponded to plant macrofossil MAZ 1 zone, and the pollen and NPP assemblage PAZ 4 zone corresponded to the plant macrofossil assemblage MAZ 2 zone.
PAZ 1 (ca. 1320 to ca. 1630 AD; ca. 600 to ca. 320 cal years BP). This zone was characterized by Poaceae and accompanied by Cyperaceae and Chenopodioideae, while Asteroideae and Ambrosia were present with low values (≤ 15%). Among the NPP Botryococcus braunii Kützing dominated the zone, followed by Pediastrum and Scenedesmus which showed increased values towards the top. The Zygnemataceae Spirogyra and the cyanobacteria Gloeotrichia also showed low values. Diverse emergent macrophytes such as Bacopa, Ranunculus, Alternanthera, Polygonum, Phyla nodiflora (L.) Greene, Triglochin, Typha and Apiaceae were present with values of up to 10%. In this zone, the highest values (≤ 20%) of Bryophyta were noted.
PAZ 2 (ca. 1630 to ca. 1840 AD; ca. 320 to ca. 110 cal years BP). This zone was characterized by maximum values of Poaceae and Chenopodioideae (≤ 70% and ≤ 40%, respectively) accompanied by Asteroideae and Ambrosia. Cyperaceae declined to minimum values (≤ 10%). Botryococcus braunii showed an important drop and was replaced by Pediastrum, which achieved its highest values (≤ 40%). The emergent macrophytes were present with values similar to those of the previous zone. Bryophyta displayed a slight decrease compared to PAZ 1 but with stable values along the zone.
PAZ 3 (ca. 1840 to ca. 1880 AD; ca. 110 to ca. 70 cal years BP). Myriophyllum dominated this zone, accompanied by Poaceae and Cyperaceae. Chenopodioideae, Asteroideae and Ambrosia decreased in value. Pediastrum was present with low values (≤ 20%), followed by Scenedesmus and Botrycoccus braunii. The Zygnematales (Zygnema and Spirogyra) showed slightly increased values. Bryophyta showed a tendency towards a decline.
MAZ 1 (ca. 1320 to ca. 1865 AD; ca. 600 to ca. 85 cal years BP). Myriophyllum spicatum L. mericarps presents the highest concentration values (63 units 10 cm−3) towards the top of the zone, followed by Tolypella oospores (2 units per 10 cm3). Close to the upper limit of the zone, a tendency towards an increase in Piscicola geometra and Biomphalaria peregrina was noted, while Myriophyllum spicatum remains decreased.
PAZ 4 (ca. 1880 to 2013 AD). This zone was characterized by Poaceae and Cyperaceae (maximum values), accompanied by Chenopodioideae, Asteroideae and Ambrosia which presented values higher than in PAZ 3. Exotic tree taxa such as Eucalyptus, Pinaceae, Casuarina, Corylus and Cupressus increased together with exotic herbs like Centaurea and Carduus. Other pollen types that include exotic species (Cichorioideae, Plantago, Rumex and Brassicaceae) were also recorded. Furthermore, Myriophyllum was replaced by Potamogeton pectinatus. Among the Other Emergent taxa, Typha showed the maximum values. A trend towards a more abundant and diverse NPP community was identified. Zygnema, Spirogyra, Desmidiaceae and the cyanobacteria Gloeotrichia exhibited their maximum values at the beginning of this zone (≤ 20%).
MAZ 2 (ca. 1865 to 2013 AD). A shift towards a more diverse and abundant community was detected in this zone. Macrophytes values increased: Potamogeton pectinatus and Nitella dominated this section, accompanied by Cyperaceae and Tolypella. Biomphalaria peregrina were more abundant in this zone.
Discussion
The biological record of Cabeza de Buey lake indicated the presence of a freshwater body since ca. 1320 AD until present. Simultaneous changes in relative frequencies of plant remains (pollen and other macrostructures), algae, some invertebrate remains and organic and carbonate content allowed reconstructing the lake history, structure and dynamics, including variations in trophic status, nutrient content and water level. Since ca. 1320 AD, four periods were identified according to characteristic communities, which define different lake structure and functioning. The lake evolved from an algal-dominated community towards a much diverse system conformed by angiosperms, charophytes and other algae through time. The community was initially dominated (ca. 1320–1630 AD) by the algae Botryococcus braunii and later by Pediastrum (ca. 1630–1840 AD), which was replaced by the submerged plant Myriophyllum at ca.1840 AD. At ca.1880 AD, the aquatic community became much diverse, integrated by Potamogeton pectinatus, accompanied by the charophytes Nitella and Tolypella, along with Cyperaceae and a heterogeneous algae community. A summary of the lake evolution, the changes in the structure and in the dominant communities, variations in the trophic state and in the content of nutrients as well as the forcing factors are presented in Fig. 6.

Lake evolution, including dominant communities, trophic status, nutrient content and water level for each period. Blue and yellow ellipses indicate the forcings that triggered the change
A horizontal vegetation structure as a continuous thick belt around shallow lakes of southeastern Pampa plain (Stutz et al. 2014) was interpreted based on similar percentages of other emergent macrophytes and higher percentages of Cyperaceae. The lower values of Cyperaceae in Cabeza de Buey lake allowed interpreting a discontinuous spatial structure of littoral vegetation around the lake during ca. 1320–1840 AD.
Turbid state
Between ca. 1320 and ca. 1840 AD, an enlarged, oligotrophic brackish to mesotrophic freshwater body occurred, with low to medium nutrient content and low energy. This period was dominated by phytoplankton taxa such as Pediastrum, Botryococcus braunii, Scenedesmus, Tetraedron and Tetrastrum. These taxa were consistent with those currently found in turbid shallow Pampean lakes (Allende et al. 2009; Izaguirre et al. 2012; Sanzano et al. 2014).
Initially, from ca. 1320 to ca. 1630 AD, the dominance of Botryococcus braunii indicated an oligotrophic brackish environment since this alga is more widespread in brackish water bodies than any other Chlorophyceae. It usually develops most abundantly in shallow water bodies that are subject to relatively low precipitation, and it tolerates low-nutrient conditions (Batten and Grenfell 1996; Reynolds 2006; Sanzano et al. 2014). In addition, brackish conditions were indicated by the highest carbonate content. In agreement, Plastani (2016) inferred for this period an oligo-mesohaline lake based on the dominance of the ostracod Limnocythere cf. solum in Cabeza de Buey lake. Furthermore, the presence of Spirogyra reinforced the inference of a shallow brackish-water lake, since this genus of freshwater macroalgae produces zygospores in water depths generally shallower than 50 cm, and some species tolerate slightly brackish conditions (Van Geel 2001; Borel et al. 2003). The presence of Gloeotrichia indicated low N content, because these cyanobacteria are capable of fixing N in conditions when this element is a limiting resource (Van Geel 2001). At the same time, the lowest organic matter content indicated a low nutrient content in the lake and supports the oligotrophic inference. In concordance, the sedimentology suggested calm waters with low energy that allowed a diffuse lamination of the fine sediments in a small water body, similar to what was reported by Guerra et al. (2015) for Melincué lake. In other Pampean shallow lakes very shallow and ephemeral saline lake environments were inferred. These were subject to desiccation-flood cycles based on massive deposits with low organic matter content and high carbonate content (Cordoba et al. 2014; Guerra et al. 2015).
Afterwards, between ca. 1630 and ca. 1840 AD, the dominance of Pediastrum together with Scenedesmus and the decline in Botryococcus braunii abundance indicated a freshwater environment and a small to medium size water body, enriched in nutrients (Reynolds 2006). The increase in the organic matter content supported the increased lake nutrient content. Borel et al. (2003) inferred high nutrient content at Hinojales shallow lake, in the southeastern Pampa plain, based on Pediastrum and Scenedesmus dominance. Cony et al. (2014) evaluated the trophic state and the characteristics of the phytoplankton community during one year cycle of a shallow lake in the southwestern Pampa plain, and reported species of Pediastrum and Scenedesmus as indicators of high nutrient concentrations. The change in abundance of Botryococcus and Pediastrum could indicate an increase in the input of water to the system, and hence an increase in precipitation. Medeanic (2006) inferred that high Botryococcus concentrations may be attributed to dry periods and increases in Pediastrum may indicate wet periods in Dos Patos Lagoon (Brazil). In addition, the change in granulometry and composition of the sediments, the massive accumulation and intermediate content of carbonates together with the increase in the organic matter indicated an enlarging environment with more energy in accordance with the entrance of water and sand to the lake from the surrounding area. According to Guerra et al. (2015), massive organic matter-rich levels were associated with higher lake productivity and wet phases for Melincué lake.
The increase in Chenopodioideae around 1800 AD suggested a dry period with reduction of the lake size. Pampean shallow lakes decreased in size during dry periods, and halophytic plants quickly colonized the extensive surrounding area that became free of water and consequently the surface was characterized by brackish conditions. Tonello and Prieto (2008) based on a dataset of surface pollen samples and proposed Chenopodioideae to be a good indicator of low precipitation values. Moreover, the peak in the carbonate content before 1800 AD suggested a shrinking lake. Thus, Cabeza de Buey lake could indicate a pulse of precipitation values lower than the modern ones at this moment at central Pampa plain. Historical chronicles mentioned a much lower water level for the lake in 1760 AD (Martinez 1967).
Clear state
From ca. 1840 to ca. 1880 AD, the aforementioned phytoplankton taxa decreased, coinciding with a sudden increase of submerged macrophytes and the filamentous chlorophytes of the Zygnemataceae family. According to Scheffer´s (1998) hypothesis of alternative stable states (ASS), maximum values of both pollen and plant macrofossils of submerged macrophytes suggest a clear water body with considerable light penetration. The phytoplankton community decreased and simultaneously the indicators associated with clear water conditions Spirogyra, Zygnema, Mougeotia and Debarya (the latest two not showed) increased; particularly, the Zygnemataceae inhabits shallow and oxygen-rich freshwater lakes (Van Geel 2001). Also at ca. 1840 AD the submerged macrophyte Myriophyllum increased significantly, reinforcing the transition between the previous period dominated by phytoplankton and the onset of a period with better light penetration into the lake that could be associated with clear conditions in the water body. The presence of Myriophyllum indicated an alkaline inorganic meso-eutrophic lake with a high pH (García et al. 2009). Egertson et al. (2004) also mentioned that as water level decreases, macrophytes can overcome light limitation. Light can reach the sediment and germinate seeds and permit photosynthetically active radiation to penetrate to new areas in the water column. Thus, the development of Myriophyllum might have been favored by a low lake level, probably as a result of low rainfall. According to historical chronicles, between ca. 1800 and ca.1880 AD there were important droughts in Cabeza de Buey area, and at 1876, historical chronicles mention a much lower water level of the lake (Martinez 1967). Furthermore, the organic matter content decreased towards the end of the period, suggesting a drop in the nutrient content. The organic matter drop and the diffuse lamination could indicate a smaller and calmer environment that enabled a slower sedimentation (Guerra et al. 2015). The aforementioned decrease in precipitation in Cabeza de Buey lake area with the consequent decrease of the lake level would have contributed to reduce the runoff of nutrients into the lake, reducing phytoplankton growth and turbidity, and allowing light penetration after ca.1800 AD. Clear lakes and plenty of submerged macrophytes, such as Zannichellia sp. and charophytes, were observed in some southeastern Pampean lakes after the drought of 2008-2009 when lakes became recovering (author’s personal observations). It is also evident that between ca. 1850-1880 AD plant-associated fauna Biomphalaria peregrina and Piscicola geometra began to appear at the same time that Myriophyllum developed, since both are closely associated with macrophytes for habitat and grazing periphyton. According to Tietze and De Francesco (2010), B. peregrina is associated with submerged vegetation because these plants reduce the current velocity, supply oxygen, and absorb toxic substances, as well as provide suitable substrata for oviposition and feeding due to the periphyton growing on them.
Changes in the dominant communities in shallow lakes (e.g. from a macrophyte-dominated community to a phytoplankton-dominated one and viceversa) are known to be triggered by several factors such as nutrient loading, fish introduction/removal or changes in water level (Scheffer 1998; Cano et al. 2008). We are not able to specifically indicate the factor that provoked the change in the community structure in Cabeza de Buey lake, but it is probably related to a decrease in the water level, likely caused by a decrease in precipitation. This change in the community structure caused by a decrease in the water level was inferred by Sanchez et al. (2015) when analyzing changes in phytoplankton during the transition from a clear vegetated to a turbid state from 2005 to 2013 for El Triunfo lake (northeast Pampa plain); Cano et al. (2008) showed that Lacombe lake (northeast Pampa plain) changed from a phytoplankton dominated state (turbid) to a epiphyton-dominated state (clear) in relation to increasing water level and abundance of macrophytes. Moreover, low precipitation has been mentioned as a trigger from turbid to clear state during the last 500 years in southeastern Pampa plain (Laprida et al. 2009). Probably, a period with dry conditions characterized Cabeza de Buey area close to the end of the phytoplankton dominated period (turbid state) ca. 1800 AD, as was previously described by the increase in Chenopodioideae. These climatic conditions were finally established during the macrophyte-dominated period (1840–1880), with precipitation values lower than at present.
Turbid with clear episodes
From ca.1880 AD to the present, phytoplankton coincided with the presence of Potamogeton pectinatus, Myriophyllum, Nitella and Tolypella along with the filamentous algae of the Zygnemataceae family. Therefore, and in relation to the previous period, it is proposed that the system represented an alternation between communities dominated by phytoplankton and by macrophytes suggesting a general turbid state with clear episodes. The shift towards a predominance of Potamogeton pectinatus evidenced a eutrophication process. In addition, this period corresponded to the development of a perennial, higher-level lake.
Around 1880 AD, the emergent macrophyte community continued to present a similar discontinuous spatial structure around the lake. The emergent macrophytes showed a decrease in diversity, but Cyperaceae and Typha prevailed as at present. However, a change towards a more diverse submerged macrophyte community was noted. This community was shaped by Potamogeton pectinatus, Myriophyllum spicatum and the charophytes Nitella and Tolypella, suggesting an alkaline, freshwater, high pH and nutrient-rich lake, in a clear condition (García et al. 2009), along with the increase in Zygnemataceae and decrease in the Chlorococcales. The dominance of macrophytes limited the nutrients available to Chlorococcales resulting in higher water clarity and a predominance of benthic primary production (Bennion et al. 2017). These conditions characterized the beginning of this period in Cabeza de Buey lake. Macrophytes favored clear conditions by several mechanisms, such as competing for nutrients with phytoplankton, secreting allelopathic substances and providing refuge for zooplankton grazers (Sanchez et al. 2015).
Pollen and plant-macrofossil remains indicated that the aquatic vegetation changed over the last 150 years towards species associated with eutrophic conditions, probably related to the pasture-arable conversion that started around 1880 AD. A series of processes that operated since the first European settlement (ca. 1540 AD) have modified the vegetation of the Pampa plain. The initial forcing was the introduction of large herbivores such as cows and horses around 1541 AD. Even though cattle breeding was restricted to a small area around Buenos Aires city until the end of the nineteenth century, both livestock and horses became wild and expanded into the Pampa plain, increasing the grazing pressure and favoring the seed dispersal of native and exotic weeds (Garavaglia 1999). After 1880 AD, the expansion of the colonized territory and the arrival of European immigrants, together with the advent of railroads and wire fences introduced new transformations into the landscape. During the twentieth century a period of major agricultural development arose with the introduction of cereal crops throughout Pampa plain, together with a succession of technological improvements. This period is locally known as Pampa Agricola Cerealera. Thus, the Pampa plain presented large areas with crops and cultivated pastures that were invaded by weeds impacting on the agricultural production and, because of this, pesticides were implemented. During the mid-1970s the introduction of the soybean crop generated structural and functional changes like the increase in the use of agrochemicals and the intensity of soil tillage, together with different methods of handling crop residues (Ghersa and Martinez Ghersa 1991).
The process of eutrophication was evidenced by the increase of Potamogeton pectinatus, the decline in abundance of charophytes and Myriophyllum spicatum and the increase in the organic matter content. One possible explanation of the decline of the submerged macrophytes could be the increase of nutrients that entered to the system due to the increase of agriculture and precipitation. The massive organic rich muds, along with the increased organic matter, suggested the development of a lake of intermediate to high water level and a high primary productivity (Guerra et al 2015; Plastani et al 2019). An important water level rise was inferred from organic matter increases, finer grain sized sediments and salinity drops (as noted by low carbonate amounts). Even though Myriophyllum spicatum is known to tolerate nutrient-rich waters, it is also known to decline over the mesotrophic-eutrophic transition (Sayer et al. 2010). Another plausible explanation for this context is that Myriophyllum spicatum and charophytes might have been outcompeted by Potamogeton pectinatus, which is able to form a canopy to use the light near the water surface (Van den Berg et al. 1998), overshadowing and limiting light availability. Tall canopy-forming angiosperms such as Potamogeton pectinatus have a competitive advantage over low growing charophytes, which are typically outcompeted by these nutrient-tolerant species. This pattern of charophytes still coexisting with angiosperms is indicative of a lake getting enriched with nutrients and becoming eutrophic (Blindow 1992; Bennion et al. 2017). The eutrophication process of the lake is also supported by the presence of Pediastrum, Scenedesmus, Zygnemataceae and Gloeotrichia. Borel et al. (2003) suggested nutrient increment based on the increase of Pediastrum, Scenedesmus, Zygnemataceae and Gloeotrichia in the southeastern Pampa plain. During the 20th, the Pampean lakes became more eutrophic as a consequence of increased nutrient loading connected mainly with the intensification of agriculture that occurred during the last 50 years (Quirós et al. 2006; Stutz et al. 2012; Plastani et al. 2019). The eutrophication took place in lakes highly exposed to intense land use of their drainage basins such as Cabeza de Buey lake, which is located in an area subjected to grazing and agriculture since ~ 1880 AD (Martinez 1967). According to Guerra et al. (2015), the increased productivity in Melincué lake during the last 40 years can be associated with the higher water levels as well as with the potential effect of the agricultural expansion that started in the region after the 1970s. In concordance, Plastani et al. (2019) recognized that the sustained expansion of agricultural production and the onset of severe anthropogenic landscape modifications increased the nutrient input with the consequent eutrophication of La Barrancosa shallow lake (southern Pampa plain).
During the last 700 years, the regional vegetation in Cabeza de Buey area was represented by grasses accompanied by herbs, such as Plantago, Rumex, and taxa belonging to Asteraceae, Brassicaceae, Caryophyllaceae and Solanaceae families. The presence of Plantago, Rumex, Brassicaceae and Cichorioideae has been regular since ca. 1320 AD, with a noticeable increase in the last two pollen types since 1900. These four pollen types represent exotic and native species. Therefore, their presence in the record between ca. 1320 and ca. 1900 AD could be interpreted as representative of native vegetation, while after 1900 AD the marked increase of Brassicaceae and Cichorioideae could be associated with both native and exotic species introduced with agriculture intensification throughout the Pampa plain. For example, Brassica nigra, Rumex crispus and Plantago lanceolata L. are introduced weeds associated to crops (Vervoorst 1967). Also, the presence of Carduus and Centaurea (Asteraceae subf. Asteroideae) in the pollen record after 1900 could be associated with cattle impact, since after cattle introduction several grassland areas of the Pampa plain were replaced by thistles (Vervoorst 1967). Ambrosia´s increase (Asteraceae subf. Asteroideae) after 1930 could be associated with both cattle impact and intensive agriculture, since it is a species that rapidly encroaches on overgrazed Pampean grassland (Poggio et al. 2015) and, additionally, it is an important crop weed. The presence of Eucalyptus pollen is used as chronological marker because the historical chronicle pointed out the year 1858 as the date of Eucalyptus seeds introduction in Argentina. The seeds were distributed among farmers and trees were quickly incorporated in the landscape as they provided shade for livestock. In the pollen record, Eucalyptus values increased after ca. 1920 AD. These values could be associated with the first farm settlements in the area and the foundation of San Carlos de Bolivar at 1877 AD, which had a commercial and demographic development after 1900 AD. Through the analysis of these pollen indicators it was possible to identify the occurrence and the time of the human induced perturbations in the area, despite the difficulty of taxonomic resolution in pollen identification that made it impossible to separate the natural from the introduced taxa in some cases.
Conclusions
Cabeza de Buey lake proved to be a good sentinel of environmental change, where past conditions are inferred, enabling the reconstruction of lake history based on changes in structure and dynamics, as well as variations in the trophic state, the nutrient content and its water level. It was also possible to recognize anthropogenic impact that affected the lake basin after 1880 AD. Given the relatively low intensity of human activity during the early period of the lake history changes in phytoplankton and macrophyte communities are expected to have been mainly controlled by climate during the early period of the lake history (prior to 1880 AD). After 1880 AD, the lake changes probably resulted from a combination of climate (increases and decreases of precipitation values) and human impact (intensive agriculture and urbanization), generating an accelerated eutrophication. The last hundred and fifty years of the lake history are characterized by the eutrophication related to the impact of agriculture and cattle breeding implemented in the landscape, as well as to the establishment of towns in the surroundings.
Several current ecological studies in Pampean shallow lakes are based on Quirós et al. (2002) which described a pristine clear state of these aquatic ecosystems prior to human impact. Our study agrees with the conclusion given that Cabeza de Buey lake experienced a clear state before the evidence of intensive human impact and provides evidence that frames it in time at 1850 AD. Nowadays, there is great concern in acknowledging shallow lakes reference conditions in terms of restoration and management. Therefore, it is of great importance to carry out studies that enable us to fulfill this task.
References
Agosta EA, Compagnucci RH (2008) The 1976/77 austral summer climate transition effects on the atmospheric circulation and climate in southern South America. J Clim 21:4365–4383
Allende L, Tell G, Zagarese H, Torremorell A, Perez G, Bustingorry J, Escaray R, Izaguirre I (2009) Phytoplankton and primary production in clear-vegetated, inorganic-turbid, and algal-turbid shallow lakes from the pampa plain (Argentina). Hydrobiologia 624:45–60
Batten DJ, Grenfell HR (1996) Chapter 7D. Botryococcus. In: Jansonius J, McGregor DC (eds) Palynology: principles and applications. Texas, American Association of Stratigraphic Palynologist Fundation, Dallas, pp 205–214
Bennett KD, Willis KJ (2001) Pollen. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments: Terrestrial, Algal and Siliceous Indicators, 3. Kluwer, Dordrecht, pp 5–32
Bennion H, Davidson TA, Sayer CD, Simpson GL, Rose NL, Sadler JP (2015) Harnessing the potential of the multi-indicator palaeoecological approach: an assessment of the nature and causes of ecological change in a eutrophic shallow lake. Freshwater Biol 60:1423–1442
Bennion H, Sayer CD, Clarke SJ, Davidson TA, Rose NL, Goldsmith B, Rawcliffe R, Burgess A, Clarke G, Turner S, Wiik E (2017) Sedimentary macrofossil records reveal ecological change in English lakes: implications for conservation. J Paleolimnol 60(2):329–348
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518
Blindow I (1992) Decline of charophytes during eutrophication: comparison with angiosperms. Freshwater Biol 28:9–14
Borel CM, Guerstein GR, Prieto AR (2003) Palinomorfos acuáticos (algas y acritarcos) del Holoceno de la laguna Hinojales (Buenos Aires, Argentina): interpretación paleoecológica. Ameghiniana 40:531–544
Cano MG, Casco MA, Solari LC, Mac Donagh ME, Gabellone NA, Claps MC (2008) Implications of rapid changes in chlorophyll-a of plankton, epipelon, and epiphyton in a Pampean shallow lake: an interpretation in terms of a conceptual model. Hydrobiologia 614:33–45
Cano MG, Casco MA, Claps MC (2016) Epipelon dynamics in a shallow lake through a turbid-and a clear-water regime. J Limnol. https://doi.org/10.4081/jlimnol.2016.1340
Cony NL, Ferrer NC, Cáceres EJ (2014) Evolución del estado trófico y estructura del fitoplancton de un lago somero de la Región Pampeana: laguna Sauce Grande (provincia de Buenos Aires, Argentina). Biología Acuática 30:70–91
Córdoba F, Guerra L, Cuña Rodríguez C, Sylvestre F, Piovano EL (2014) Una visión paleolimnológica de la variabilidad hidroclimática reciente en el centro de Argentina: desde la Pequeña Edad de Hielo al siglo XXI. Lat Am J Sedimentol Basin Anal 21:139–163
Egertson CJ, Kopaska JA, Downing JA (2004) A century of change in macrophyte abundance and composition in response to agricultural eutrophication. Hydrobiologia 524:145–156
Fontana SL (2005) Holocene vegetation history and palaeoenvironmental conditions of the temperate Atlantic coast of Argentina, as inferred from multiple proxy lacustrine records. J Paleolimnol 34:445–469
Fontijn K, Rawson H, Van Daele M, Moernaut J, Abarzúa AM, Heirman K, Bertrand S, Pyle DM, Mather TA, De Batist M, Naranjo JA, Moreno H (2016) Synchronisation of sedimentary records using tephra: a postglacial tephrochronological model for the Chilean Lake District. Quaternary Sci Rev 137:234–254
Garavaglia JC (1999) Pastores y labradores de Buenos Aires. Una historia agraria de la campaña bonaerense 1700–1840. Buenos Aires, Ediciones de la Flor, p 408
García P, Fernández R, Cirujano S (2009) Habitantes del agua Macrófitos. Editorial Monto Cultura, 278 pp
Geraldi AM, Piccolo MC, Perillo GME (2011) El rol de lagunas bonaerenses en el paisaje pampeano. Ciencia Hoy 21:8–14
Ghersa CM, Martinez Ghersa MA (1991) Cambios Ecológicos en los Agroecosistemas de la Pampa Ondulada. Efectos de la Introducción de la Soja. Cienc Invest 5:182–188
Grimm E (2015) Tilia Software. Springfield, Illinois State Museum
Guerra L, Piovano EL, Córdoba FE, Sylvestre F, Damatto S (2015) The hydrological and environmental evolution of shallow Lake Melincué, central Argentinean Pampas, during the last millennium. J Hydrol 529:570–583
Heiri O, Lotter AF, Lemcke G (2001) Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. J Paleolimnol 25:101–110
Hogg AG, Hua Q, Blackwell PG, Niu M, Buck CE, Guilderson TP, Heaton TJ, Palmer JG, Reimer PJ, Reimer RW, Turney CS, Zimmerman SRH (2013) SHCal13 Southern Hemisphere calibration, 0–50,000 years cal BP. Radiocarbon 55:1889–1903
Izaguirre I, Allende L, Escaray R, Bustingorry J, Pérez G, Tell G (2012) Comparison of morpho-functional phytoplankton classifications in human-impacted shallow lakes with different stable states. Hydrobiologia 698:203–216
Kowalewski GA, Kornijów R, McGowan S, Kaczorowska A, Bałaga K, Namiotko T, Gąsiorowski M, Wasiłowska A (2016) Disentangling natural and anthropogenic drivers of changes in a shallow lake using palaeolimnology and historical archives. Hydrobiologia 767:301–320
Labraga JC, Scian B, Frumento O (2002) Anomalies in the atmospheric circulation associated with the rainfall excess or deficit in the Pampa Region in Argentina. J Geophys Res Atmos. https://doi.org/10.1029/2002JD002113
Laprida C, Valero-Garcés B (2009) Cambios ambientales de épocas históricas en la pampa bonaerense en base a ostrácodos: historia hidrológica de la laguna de Chascomús. Ameghiniana 46:95–111
Laprida C, Orgeira MJ, García Chapori N (2009) El registro de la Pequeña Edad de Hielo en lagunas pampeanas. Revista de la Asociación Geológica Argentina 65:603–611
Laprida C, Massaferro J, Mercau R, Josefina M, Cusminsky G (2014) Paleobioindicadores del fin del mundo: ostrácodos y quironómidos del extremo sur de Sudamérica en ambientes lacustres cuaternarios. Lat Am J Sedimentol Basin Anal 21:97–117
Leon RJC (1991) Vegetacion. In: Coupland RT (ed) Ecosystems of the World 8A. Natural Grasslands. Introduction and Western Hemisphere, Amsterdam, The Netherlands, Elsevier, pp 380–387
Mancini M, Salinas V, Biolé F, Morra G, Montenegro H (2013) Nuevo registro para la provincia de Córdoba (Argentina) y aportes a la ecología de Parapimelodus valenciennis (Pisces, Pimelodidae). Bioscriba 6:1–8
Martinez AM (1967) San Carlos de Bolivar. Impresiones Schmidel, Buenos Aires, p 506
Medeanic S (2006) Freshwater algal palynomorph records from Holocene deposits in the coastal plain of Rio Grande do Sul, Brazil. Rev Palaeobot Palyno 141:83–101
Messineo PG, Tonello MS, Stutz S, Tripaldi A, Scheifler N, Pal N, Sanchez Vuihard G, Navarro D (2019) Human occupation strategies and related environmental-climate during the middle and late Holocene in central Pampas of Argentina. Holocene 29:244–261
O’Farrell I, Izaguirre I, Chaparro G, Unrein F, Sinistro R, Pizarro H, Rodríguez P, Tezanos Pinto P, Lombardo R, Tell G (2011) Water level as the main driver of the alternation between a free-floating plant and a phytoplankton dominated state: a long-term study in a floodplain lake. Aquat Sci 73:275–287
Piovano EL, Ariztegui D, Moreira SD (2002) Recent environmental changes in Laguna Mar Chiquita (central Argentina): a sedimentary model for a highly variable saline lake. Sedimentology 49:1371–1384
Piovano EL, Ariztegui D, Cordoba F, Cioccale M, Sylvestre F (2009) Hydrological variability in South America below the Tropic of Capricorn (Pampas and Patagonia, Argentina) during the last 13.0. In: Vimeux F (ed) Past Climate Variability in South America and Surrounding Regions: from the Last Glacial Maximum to the Holocene. Dev Paleoenviron Res, Springer, New York, pp 323–351
Plastani MS, Laprida C, Montes de Oca F, Massaferro J, Panarello HO, Ramón Mercau J, Lami A (2019) Recent environmental changes inferred from sediments in a shallow lake of the Argentinean pampas. J Paleolimnol 61:37–52
Plastani (2016) Ostrácodos y paleolimnología del sur de la pampa bonaerense: reconstrucción de parámetros ambientales y climáticos para el Holoceno Tardío. Ph. D. Thesis. University of Buenos Aires, Argentina
Poggio SL, Perelman SB, Mollard FPO, Leon RJC (2015) Guests and gatecrashers in a New World’s banquet: Old World plant species introduced from the Mediterranean Basin enriched the flora of grasslands and croplands in the Pampas of Argentina. Flora Mediterránea 25:39–54
Prieto AR (1996) Late quaternary vegetational and climatic changes in the Pampa grassland of Argentina. Quaternary Res 45:73–88
Prieto AR (2000) Vegetational history of the Late glacial-Holocene transition in the grassland of eastern Argentina. Palaeogeogr Palaeocl 157:167–188
Quirós R, Boveri MB, Renella AM, Rosso J, Sosnovsky A, von Bernard HT (2006) Los efectos de la agriculturización del humedal pampeano sobre la eutrofización de sus lagunas. Causas, conseqüências e tecnologias de gerenciamentoe controle. Eutrofização na América do Sul: 1–16
Quirós R, Rennella AM, Boveri MB, Rosso JJ, Sosnovsky A (2002) Factores que afectan la estructura y el funcionamiento de las lagunas pampeanas. Ecología austral 12:175–185
Reynolds CS (2006) Ecology of phytoplankton. Cambridge University Press, Cambridge, UK, p 535
Rull V, Lara A, Rubio-Inglés MJ, Giralt S, Gonçalves V, Raposeiro P, Hernandez A, Sanchez-Lopez G, Vazquez-Loureiro D, Bao R, Masque P, Masqué P (2017) Vegetation and landscape dynamics under natural and anthropogenic forcing on the Azores Islands: a 700-year pollen record from the São Miguel Island. Quaternary Sci Rev 159:155–168
Sánchez ML, Lagomarsino L, Allende L, Izaguirre I (2015) Changes in the phytoplankton structure in a Pampean shallow lake in the transition from a clear to a turbid regime. Hydrobiologia 752:65–76
Sanchez Vuichard G (2019) Historia reciente (últimos 500 años) de los lagos someros de la llanura Pampeana a partir del análisis de múltiples indicadores. Ph. D. Thesis. National University of Mar del Plata, Argentina
Sanzano P, Grosman F, Colasurdo V (2014) Estudio limnologico de laguna Blanca Chica (Olavarria, Provincia de Buenos Aires) durante un periodo de sequía. Biol Acuat 30:189–202
Sayer CD, Burgess A, Kari K, Davidson TA, Peglar S, Yang H, Rose N (2010) Long-term dynamics of submerged macrophytes and algae in a small and shallow, eutrophic lake: implications for the stability of the macrophyte dominance. Freshwater Biol 55:565–583
Scheffer M (1998) Ecology of shallow lakes. Ed. Chapman & Hall. Londres, 357 pp.
Stutz S, Borel CM, Fontana SL, del Puerto L, Inda H, García-Rodriguez F, Tonello MS (2010) Late Holocene environmental evolution of Nahuel Rucá freshwater shallow lake, SE Pampa grasslands, Argentina. J Paleolimnol 44:761–775
Stutz S, Borel CM, Fontana SL, Tonello MS (2012) Holocene evolution of three shallow lakes in the SE Pampa plain (Argentina) as evidenced by analyses of pollen, non-pollen palynomorphs and plant macrofossils. Holocene 22:1215–1222
Stutz S, Tonello MS, González Sagrario MS, Navarro D, Fontana SL (2014) Historia ambiental de los lagos someros de la llanura Pampeana (Argentina) desde el Holoceno medio: inferencias paleoclimáticas. Lat Am J Sedimentol Basin Anal 21:119–138
Tietze E, De Francesco CG (2010) Environmental significance of freshwater mollusks in the Southern Pampas, Argentina: to what detail can local environments be inferred from mollusk composition? Hydrobiologia 641:133–143
Tonello MS, Prieto AR (2008) Modern vegetation-pollen-climate relationship for the Pampa grasslands of Argentina. J Biogeogr 35:926–938
Tripaldi A, Zárate MA (2016) A review of Late Quaternary inland dune systems of South America east of the Andes. Quatern Int 410:96–110
Van den Berg MS, Coops H, Meijer ML, Scheffer M, Simons J (1998) Clear water associated with a dense Chara vegetation in the shallow and turbid Lake Veluwemeer, The Netherlands. In: Jeppesen E, Sondergaard M, Sondergaard M, Christoffersen K (eds) The structuring role of submerged macrophytes in lakes. Springer-Verlag, New York, pp 339–352
Van Geel B (2001) Non-pollen palynomorphs. In: Smol JP, Birks HJB, Last WM (eds) Tracking Environmental Change using Lake Sediments: Terrestrial, Algal and Siliceous Indicators, 3. Kluwer, Dordrecht, pp 99–119
Vervoorst F (1967) La vegetación de la República Argentina. VII Las comunidades vegetales de la depresión del Salado (Provincia de Buenos Aires). INTA Serie Fitogeográfica 7 Buenos Aires, 262 pp
Vilanova I, Schittek K, Geilenkirchen M, Schäbitz F, Schulz W (2015) Last millennial environmental reconstruction based on a multi-proxy record from Laguna Nassau, Western Pampas, Argentina. Neues Jahrb Geol P-A 277:209–224
Zárate MA, Tripaldi A (2012) The aeolian system of central Argentina. Aeolian Res 3:401–417
Acknowledgements
We thank Pablo Messineo, Nahuel Scheifler and Gonzalo Sottile for their support during coring activities. Alfonsina Tripaldi provided useful comments on the manuscript. Financial support was provided by PIP CONICET 582 (2015-2017), PIP CONICET 0088 (2014-2016), National Geographic Society Grants 9773-15, EXA 913/18 (UNMdP).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sánchez Vuichard, G., Stutz, S., Tonello, M.S. et al. Structure and dynamics of a Pampa plain, (Argentina) shallow lake over the last 600 years. J Paleolimnol 66, 141–155 (2021). https://doi.org/10.1007/s10933-021-00194-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-021-00194-w