
Abstract
Epidermal growth factor receptor (EGFR) dysregulation is observed in many human cancers and is both a cause of oncogenesis and a target for chemotherapy. We previously showed that partial charge neutralization of the juxtamembrane (JX) region of EGFR via the EGFR R1–6 mutant construct induces constitutive receptor activation and transformation of NIH 3T3 cells, both from the plasma membrane and from the ER when combined with the ER-retaining L417H mutation (Bryant et al. in J Biol Chem 288:34930–34942, 2013). Here, we use chemical crosslinking and immunoblotting to show that these mutant constructs form constitutive, phosphorylated dimers in both the plasma membrane and the ER. Furthermore, we combine this electrostatic perturbation with conformationally-restricted receptor mutants to provide evidence that activation of EGFR R1–6 dimers requires functional coupling both between the EGFR extracellular dimerization arms and between intracellular tyrosine kinase domains. These findings provide evidence that the electrostatic charge of the JX region normally serves as a negative regulator of functional dimerization of EGFR.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
The epidermal growth factor receptor (EGFR or ErbB1) has been extensively examined as a receptor tyrosine kinase that controls a system of tightly regulated pro-survival signaling pathways within the cell [1,2,3]. Binding of ligand EGF causes dimerization of monomeric receptors and concomitant conversion to an active state, or possibly converts inactive, weakly dimerized receptors into an active state. Numerous mutations in EGFR cause spontaneous receptor activation and have been observed in human cancers [4]. As exemplified by the L834R mutant, oncogenic mutations of EGFR may work by stabilizing ligand-independent dimerization and activation, rather than directly enhancing the kinase activity of monomeric receptors [5]. In contrast, the EGFRvIII mutant does not appear to form dimers and may signal from an intracellular location [6]. Despite great progress in elucidating the structural mechanisms whereby EGFR becomes activated and the regulation—or dysregulation—of these structure–function relationships, key questions about these mechanisms in live cells remain. An ultimate goal is overcoming dysregulatory factors in treatments of cancer and other pathologies, and this will require continuing efforts to determine how different kinds of mutations drive EGFR activation.
Structurally, each EGFR monomer contains an extracellular ligand binding domain (ECD), a single transmembrane helix, and intracellular portions composed of the juxtamembrane (JX) semi-helical domain, the catalytic tyrosine kinase domain (TKD), and a C-terminal tail [1, 2]. In its basal monomeric state, hydrophobic and electrostatic interactions between the plasma membrane’s inner leaflet with the JX and TKD domains keep the receptor in a tightly constrained, inactive conformation [7] (Fig. 1A.a). Ligand binding to the ECD (Fig. 1A.b) initiates a conformational change that exposes the ECD’s dimerization arm and enables the formation of stable EGFR homodimers, or heterodimers with other members of the ErbB family [2, 8] (Fig. 1A.c). Intracellularly, ligand-bound dimers release their JX and TKD domains from association with the inner leaflet of the plasma membrane, allowing them to rearrange to form an activated, asymmetric TKD dimer, in which the C lobe of the activating receptor TKD contacts the N lobe of a receiving receptor TKD [9] (Fig. 1A.d). These allosteric rearrangements result in activation of the receiving TKD. The receiving receptor then primarily trans-phosphorylates tyrosine residues in the C-terminal tail of the activating receptor (Fig. 1A.e), providing binding sites for downstream signaling proteins [2, 3, 10,11,12]. There is also evidence for a lesser degree of cis-autophosphorylation of the receiving kinase in EGFR heterodimers [13].

Schematic diagrams of EGFR signaling and variant structures. A Canonical EGFR activation pathway. Ligand binding (a) exposes the dimerization arm (b) and allows formation of EGFR dimers (c). These dimers rearrange into an active, asymmetric conformation (d) characterized by dimerized extracellular and intracellular domains with anti-parallel arrangement of the JX domains and N–C lobe contact between the TKDs of the receiving receptor (blue TKD) and the activating receptor (red TKD). The TKD of the receiving receptor phosphorylates the C-terminal tail of the activating receptor (e). Adapted from Ref. [43]. B Schematic representation of EGFR with mutagenesis sites and R1–6 sequence. Mutated residues are underlined. The 246–253* mutant (Y246E, N247A, T249D, Y251E, Q252A, and M253D) blocks extracellular dimerization; the L417H mutant blocks exit from the ER; R1–6 reduces the basic charge of the JX domain; V948R prevents allosteric activation by preventing the N–C lobe contact of the TKDs
The JX domain is conserved across human RTKs [14] and is an essential regulator of EGFR function, with some established roles in activation and signal transduction [15]. Replacement of the JX domain with an unstructured peptide sequence results in phosphorylation-incompetent receptors [16], indicating that the normal sequence plays an essential role in activation. Activating oncogenic mutations in the JX domain of HER2 (ErbB2) have been identified, suggesting this structure normally suppresses activation [17]. The JX domain may also participate in receptor multimerization and consequent phosphorylation [18, 19]. Recently identified binding partners of the JX domain suggest additional routes for participating in signaling beyond regulating EGFR conformational states [20, 21]. Several recent therapeutics and EGFR inhibitors have been developed to target the JX domain [22, 23]. Together, these findings point to significant roles for the JX domain in EGFR activation and regulation as well as a potential therapeutic target.
Despite increasing appreciation of the JX domain, the relationship between its structure and function in cells remains poorly understood. Structurally, the JX domain is composed of two conjoined, electrostatically-charged, amphipathic semi-helices. It begins as a continuation of the single transmembrane helix and is located proximal to the inner leaflet of the plasma membrane (Fig. 1A.a). In inactive receptors, the JX domain interacts with the plasma membrane through electrostatic interactions between its polybasic residues (646–656; Fig. 1B) and negatively charged membrane phospholipids [24,25,26] as well as through hydrophobic residue insertions [7]. During activation, both the TKD and JX domains are released from the inner leaflet and the JX domains in EGFR homodimers form anti-parallel helices that are stabilized by mutual interactions of their hydrophobic residues [7, 27] (Fig. 1A.d). Additionally, the JX polybasic region has been shown to interact with an acidic region within the TKD to stabilize the active dimer conformation [28] (Fig. 1A.d). Thus, the JX polybasic region appears not only to prevent activation through membrane interactions in the basal state, but also to promote dimer stability in activated EGFR.
Previously, we demonstrated that the conserved polybasic region within the JX domain is involved in maintaining EGFR in an inactive basal state. Charge neutralizing mutagenesis of this region, referred to as EGFR R1–6 (Fig. 1B), results in constitutive activation of EGFR and induces cellular transformation as determined by anchorage-independent growth in vitro. We further determined that this mutant preferentially signals via phosphatidylinositol 3-kinase (PI3K) [29]. Due to the varied regulatory roles of the JX region, the question remains that we address herein: how does electrostatic silencing lead to constitutive activation of EGFR? We previously found that EGFR R1–6 retains its constitutive activity when confined to the endoplasmic reticulum (ER) via the novel L417H mutation [29]. This mutation introduces a histidine residue into the hydrophobic core of the leucine-rich repeat of domain III in the ECD [30], which may cause this domain to misfold and prevent its successful trafficking out of the ER following translation. If this misfolding also prevents the ECD from dimerizing, then EGFR R1–6 may not require ECD dimerization for its constitutive activation.
Our studies address mechanisms underlying the regulatory roles of the JX polybasic region and provide new evidence that this region normally prevents the formation of active dimers in the absence of ligand. We employ conformationally-restricted mutants to examine both intra- and extracellular dimerization motifs of EGFR variants. We use chemical crosslinking and western blotting in conjunction with mutagenesis to provide further evidence that the electrostatic charges in this region play a dominant role in inhibiting spontaneous EGFR activation, which involves dimerization of both intra- and extracellular segments.
2 Methods
2.1 Materials
All cell culture reagents, EGF, and precast gels for blotting were from Invitrogen unless indicated otherwise. Gentamicin sulfate was from VWR. FuGENE HD was from Roche Applied Sciences. Mirus TransIT 2020 was from Mirus Bio. The chemical crosslinkers BS3 (bis(sulfosuccinimidyl)suberate) and DSS (disuccinimidyl suberate) were from CovaChem. Kaleidoscope molecular weight marker protein ladder was from BioRad. PVDF membranes for blotting were from Millipore. For immunoblotting, the primary rabbit antibodies specific for phospho-Y1173 EGFR and for total EGFR were from Cell Signaling Technology. The primary mouse antibody used to blot β-actin was from LabVision/Thermo. HRP-conjugated secondary antibodies were from GE Healthcare. Restore™ PLUS Western Blot Stripping Buffer was from ThermoFisher. Phusion High-Fidelity DNA Polymerase, restriction enzymes (Xba1 and EcoN1), and other cloning materials were from New England Biolabs. All chemicals were from Sigma-Aldrich unless otherwise stated.
2.2 Expression Plasmids
The human EGFR and EGR R1–6 constructs in pkH3 vectors were described previously [29]. The EGFR 246–253* construct was a gift from Dr. Mark Lemmon (Yale University) in a pAc5.1/V5-HisA vector [31]. We cloned into our pkH3 vector by cutting out the native extracellular domain using Xba1 and EcoN1 restriction sites and then cloning in the EGFR 246–253* extracellular domain using In-Fusion® HD Cloning (Clontech® Laboratories, Inc.) with GCAGGTCGACTCTAGATGCGACCCTCCGGGACG and TGTTCATGGCCTGAGGCAGG as primers.
The EGFR V948R and EGFR R1–6 V948R constructs were generated by site directed mutagenesis from their parent plasmids using Phusion High-Fidelity DNA polymerase and a forward primer of TGTACCATCGATGTCTACATGATCATGCGCAAGTGC and a reverse primer of CTATCTGCGTCTATCATCCAGCACTTGCGCATGATC.
2.3 Cell Culture
NIH 3T3 cells for transient transfections were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 5% (v/v) FBS and 5% NBCS (Atlanta Biologicals) and 1 U/mL Pen/Strep. NIH 3T3 cells were transfected with Mirus TransIT 2020 as per the manufacturer’s directions and allowed to express for 24 h prior to experiments. NIH 3T3 cells stably expressing EGFR, EGFR R1–6, and EGFR R1–6 L417H [29] were cultured in DMEM containing 5% FBS and 5% NBCS, 1 U/mL Pen/Strep, and 2 µg/mL puromycin.
RBL-2H3 cells were cultured in Modified Eagle’s media containing 20% FBS and 10 µg/mL gentamicin sulfate as described previously [32]. RBL 2H3 cells were transiently transfected using FuGENE HD, OptiMEM, and phorbol dibutyrate as previously described and allowed to express for 24 h prior to experiments [33].
2.4 Chemical Crosslinking
5 × 105 cells were plated and transfected, if necessary, in 6-well, 35 mm plates. All cells expressing EGFR constructs were starved in their respective serum-free media for 12–16 h prior to chemical crosslinking. In many procedures we used a phosphate buffered saline (PBS, pH 7.5) solution supplemented with 1 mM Mg2+. Prior to the experiment, cells were washed once with PBS/Mg2+ and then treated with 1 mL of 100 ng/mL EGF in PBS/Mg2+ for 5 min. 1 mL of PBS/Mg2+ containing 2 mM of either BS3 or DSS crosslinker or 10 µL of DMSO vehicle (control) was then added to the 1 mL of PBS/Mg2+ already in cell-containing wells (1 mM final crosslinker concentration). The plates containing the cells were cooled to 4 °C and allowed to incubate for 30 min. Unreacted crosslinker was then quenched with 40 µL 1 M glycine pH 7.2 in PBS for 15 min. Cells were washed once with PBS/Mg2+ and then incubated with cold lysis buffer (25 mM Tris, pH 7.4, 100 mM NaCl, 1 mM EDTA, 1% (v/v) Triton, 1 mM sodium orthovanadate, 1 mM β-glycerol phosphate, 1 µg/mL leupeptin, and 1 µg/mL aprotinin). Supernatants were retained following 10 min of microfuge centrifugation at 4 °C. Protein concentrations of whole cell lysates were measured using the Bio-Rad Protein Assay.
2.5 Immunoblot Analysis
Whole cell lysates (20 µg/lane) were resolved by SDS/PAGE on a 4–12% gradient gel, and Kaleidoscope molecular weight markers were run in a separate lane on the same gel. The proteins were transferred to PVDF membranes as described previously [29]. The membranes were blocked in 5% BSA diluted in 20 mM Tris, 135 mM NaCl, and 0.02% Tween 20, and incubated with primary antibodies in the same buffer overnight. Primary antibodies were detected with HRP-conjugated secondary antibodies and a Bio-Rad VersaDoc MP 5000 imager. Molecular weights were determined by overlaying the colorimetric blot image of the Kaleidoscope molecular weight marker with the densitometry image and fitting the lanes using Image Lab’s Molecular Weight Analysis tool. Blots were initially probed for pEGFR Y1173 and β-actin prior to stripping with Restore PLUS western blot stripping buffer as per the manufacturer's instructions and then reprobed for total EGFR. The colorimetric images of molecular weight markers verified running positions of monomeric (170 kD) and dimeric (340 kD) EGFR and β-actin (42 kD).
2.6 Densitometry Quantification
Immunoblots were analyzed using Bio-Rad’s Image Lab with a rolling disc background subtraction method to obtain density values for each band. pY1173 EGFR bands were normalized over multiple lanes in the same gel by dividing through by the total amount of EGFR present (monomer + dimer bands) in respective lane. Relative intensities of pY1173 dimers for EGFR variants under set conditions in multiple experiments were compared by normalizing each experiment to a specified EGFR sample in the presence of EGF and chemical crosslinker. These normalized values were then compared using a Tukey’s comparison of means in conjunction with ANOVA using JMP.
3 Results
3.1 Partial Charge-Silencing of the JX Domain Leads to Constitutive Dimerization and Phosphorylation of EGFR Constructs Expressed in the Plasma Membrane or Retained in the ER
We investigated whether an EGF receptor with a partially charge-silenced JX domain (EGFR R1–6; Fig. 1B) signals as a constitutive dimer, as is observed for many [34,35,36,37,38,39] but not all [6] constitutively active EGFR mutants. We used covalent chemical crosslinking and immunoblotting of cell lysates to detect higher molecular weight species corresponding to EGFR homodimers [40]. We evaluated NIH 3T3 cells stably expressing EGFR or EGFR R1–6, and these were stimulated (or not) with EGF and chemically crosslinked (or not) with the membrane nonpermeable crosslinker, bis-sulfosuccinimidyl-suberate (BS3), which is 11.4 Å in length and crosslinks primary amines. The cells were then lysed and immunoblotted to detect the phosphorylated tyrosine residue 1173 (EGFR pY1173), which is present in active, but not inactive, EGFR. Immunoblots of total EGFR and β-actin, as identified by their specific antibodies, provided measures of EGFR expression and relative cell-equivalent loading.
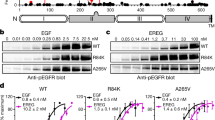
In immunoblots, monomeric EGFR (M) migrates as a 170 kDa band while dimeric EGFR (D) migrates as a 340 kDa band, as determined by standard molecular weight markers (Fig. 2A). Cells expressing wt EGFR yield an EGFR pY1173 monomer only in the presence of EFG (Fig. 2A, lane 2) and a dimer that is detectable only with both EGF stimulation and BS3 crosslinking (Fig. 2A, lane 4). In addition to demonstrating activation by EGF, these results indicate that the active, phosphorylated dimer does not withstand cell lysis and denaturation without being trapped by chemical crosslinking and that the efficiency of crosslinking is considerably less than 100%. The immunoblot also shows that the sensitivity of detection is greater for EGFR pY1173 than for total EGFR. Consistent with our previous findings [29], cells expressing EGFR R1–6 exhibit basal EGFR phosphorylation in the absence of EGF (Fig. 2A, lanes 5 and 7). We further show here that EGFR R1–6 forms a phosphorylated dimer in the absence (or presence) of EGF stimulation, and this can be detected with addition of chemical crosslinker (Fig. 2A, lanes 7 and 8). This representative immunoblot demonstrates that EGFR R1–6 forms a constitutively phosphorylated dimer when stably expressed in NIH 3T3 cells, whereas wt EGFR does not (Fig. 2A, compare lanes 3 and 7).

EGFR R1–6 forms phosphorylated dimers in the absence of ligand. A Whole cell lysates from NIH 3T3 cells stably expressing either EGFR or EGFR R1–6 and stimulated (or not) with EGF and chemically crosslinked (or not) with BS3 prior to lysis and immunoblotting with the indicated antibodies. M and D indicate monomeric and dimeric EGFR, respectively, as determined by molecular weight markers run on the same gel. As described in the text, the probing antibody specific for pY1173 is more sensitive than that for total EGFR, such that dimer bands may have differing intensities between antibodies. In these stably expressing cells, either EGFR or β-actin can serve as a loading control; incomplete transfer for blotting can occur in gel corners, which would differentially affect the intensity of the β-actin band. The appearance of a double band for β-actin is probably due to nonspecific binding to another cell component of slightly higher molecular weight. B Densitometry quantification of multiple experiments as in A (SD shown, Tukey’s HSD, n = 9, ***, p < 0.001). Data for each experiment were normalized to the total EGFR present (sum of M and D bands) and then compared over multiple experiments using stimulated and crosslinked wt-EGFR as a reference
Densitometry of multiple experiments (normalized to stimulated, crosslinked wt EGFR) indicates that the relative amount of phosphorylated EGFR R1–6 dimers in the absence of EGF is similar to phosphorylated EGFR R1–6 and wt EGFR dimers after stimulation with EGF (Fig. 2B). That EGFR R1–6 activation is not substantially enhanced by EGF indicates that mutation of the positively charged residues in the JX domain allows a transition similar to that caused by EGF and does not affect the asymmetrical TKD dimers in their capacity to trans-phosphorylate.
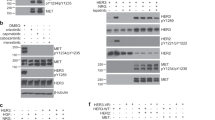
We previously identified a point mutation, L417H, that causes EGFR to be retained in the ER rather than being expressed on the cell surface [29]. We tested whether this ER-retained L417H mutation, in conjunction with wt EGFR or with EGFR R1–6, also forms dimers within the ER membrane. For this purpose, we used disuccinimidyl suberate (DSS), which is a membrane-permeable analog of BS3. NIH 3T3 cells stably expressing EGFR L417H and EGFR R1–6 L417H were stimulated, (or not), chemically crosslinked (or not), and immunoblotted (Fig. 3). As we reported previously for non-crosslinked cells, EGFR R1–6 L417H (Fig. 3A, lane 2), but not EGFR L417H (lane 1) is phosphorylated in the absence of EGF. In these cells, neither EGFR L417H nor EGFR R1–6 L417H show increased phosphorylation in the presence of EGF (Fig. 3A, compare lanes 5 and 6 with lanes 1 and 2). These results are expected because EGFR R1–6 auto-phosphorylates and because added EGF cannot access ER-retained EGFR. Use of the chemical crosslinker DSS revealed the presence of pEGFR R1–6 L417H dimers (Fig. 3A, lane 4), but not EGFR L417H (lane 3). Similar results are observed in the presence of EGF, as expected (Fig. 3A, lanes 7 and 8). Densitometry of multiple experiments of the same type (normalized to EGFR R1–6 L417H dimers; Fig. 3B) give consistent results that L417H-mediated retention in the ER affects the capacity of EGFR but not EGFR R1–6 to be activated as phosphorylated dimers. Combined, these results show that the R1–6 mutation stabilizes the active dimer conformation, regardless of EGFR R1–6 expression in the plasma membrane or in the ER.

EGFR R1–6 forms phosphorylated dimers when combined with the ER-localizing EGFR L417H mutant. A Whole cell lysates from NIH 3T3 cells stably expressing either EGFR L417H (LH) or EGFR R1–6 L417H (RLH) were stimulated (or not) with EGF and then chemically crosslinked (or not) with DSS prior to lysis and western blotting with the indicated antibodies. M and D indicate monomeric and dimeric EGFR respectively, as determined by molecular weight markers run on the same gel. See legend to Fig. 2 for description of loading controls and appearance of β-actin band for these stably expressing cells. B Densitometry quantification of multiple experiments as in A (SD shown, Tukey’s HSD, n = 4, *, p < 0.05). Data for each experiment were normalized by the total EGFR present (sum of M and D bands) in the same lane, and then compared over multiple experiments using stimulated and crosslinked R1–6-EGFR as a reference
3.2 Inhibiting the Asymmetric TKD Dimer Conformation Blocks EGFR R1–6 Constitutive Activation
We utilized the EGFR V948R mutant [13, 36, 41] (Fig. 1B) to investigate whether phosphorylation of EGFR variants requires functional dimerization, or whether these receptors can autophosphorylate as monomers prior to forming stable dimers. The V948R mutation inserts a positively charged residue in the C-terminal lobe of the TKD and prevents the N–C lobe contact required for the active, asymmetric, “CDK/Cyclin-like” kinase conformation (Fig. 1A.d). Instead, receptor dimers assume an inactive, symmetric, “Src/CDK-like” conformation [9]. A monomeric EGFR R1–6 V948R autophosphorylation event is not expected to be blocked by this mutant, but a dimer-dependent event would be.
For this experiment we used NIH 3T3 cells, transiently expressing either EGFR V948R or EGFR R1–6 V948R. These were stimulated with EGF (or not) and chemically crosslinked (or not) with BS3 as described for the experiments with stable cell lines. We found that, unlike their parental counterparts (Fig. 4, lanes 2–4), both EGFR V948R and EGFR R1–6 V948R fail to phosphorylate regardless of EGF stimulation or crosslinking (Fig. 4, lanes 5–12), indicating that both wt EGFR and EGFR R1–6 require the asymmetric kinase conformation for phosphorylation. In this experiment with transiently transfected NIH-3T3 cells we found that wt-EGFR is phosphorylated in unstimulated samples (Fig. 4, lane 2; Online Resource 1), unlike wt-EGFR as expressed in a stably transfected cell line (Fig. 2, lanes 1 and 3). This result is consistent with previous reports that over-expression and mass action leads to spontaneous dimerization and activation of EGFR [42]. However, comparison of the parallel samples in Fig. 4 underscores our primary result that the V948R mutation prevents phosphorylation in both wt EGFR and EGFR R1–6 in the absence and presence of EGF. We conclude that functional TKD dimerization is required for activating phosphorylation in both cases.

The asymmetric kinase conformation is required for EGFR R1–6 phosphorylation. Whole cell lysates from untransfected NIH 3T3 cells (n/a) or NIH 3T3 cells transiently expressing either EGFR, EGFR R1–6, EGFR V948R, or EGFR R1–6 V948R were stimulated (or not) with EGF and then chemically crosslinked (or not) with BS3 prior to immunoblotting with the indicated antibodies. M and D indicate monomeric and dimeric EGFR, respectively, as determined by molecular weight markers run on the same gel. Blot shown is representative of six independent experiments. For these transiently transfected cells, phosphorylated protein bands (pY1173 EGFR) can be visually normalized by comparing to total EGFR bands. The appearance of a double band for β-actin and incomplete transfer of this band in gel corners is described in the legend of Fig. 2
Using the anti-EGFR antibody to identify total EGFR, we observed non-phosphorylated receptor dimers in cells expressing EGFR V948R or EGFR R1–6 V948R following stimulation with EGF and crosslinking (Fig. 4, lanes 8 and 9). This result provides direct evidence of dimer formation for EGFR R1–6, independently of phosphorylation, which is reasonable because ligand binding facilitates dimerization through the ECD (Fig. 1A.b, c). We occasionally, but not consistently, detected EGFR V948R R1–6 dimers in the absence of EGF (Fig. 4, lane 10); this variability in detection (signal/noise) is likely due to the relatively lower sensitivity of the anti-EGFR antibody as described above. Interestingly, the stimulated EGFR V948R and EGFR R1–6 V948R dimer populations seem more stable than their non-V948R counterparts: with the anti-EGFR antibody we consistently detect stimulated, non-phosphorylated V948R mutant dimers, but detect stimulated parental EGFR and EGFR R1–6 dimers more variably. Together, our results show that ECD dimers are stabilized by EGF, but, like wt EGFR, EGFR R1–6 requires functional intracellular dimerization of the TKDs to activate.
3.3 Inhibiting ECD Dimerization Blocks EGFR R1–6 Constitutive Activation
We investigated whether the ECD dimer conformation (Fig. 1A) is required for EGFR R1–6 phosphorylation. The 246–253* mutation in EGFR alters key residues (Y246E, N247A, T249D, Y251E, Q252A, and M253D) along the domain II dimerization arm that normally stabilizes ECD dimers following ligand binding (Fig. 1A.b,c and B) [31]. EGFR 246–253* binds ligand, but does not form an ECD dimer [31]. This mutation was previously employed with clinically observed oncogenic EGFR mutants and shown to prevent their oncogenic activity [31, 39].
We found that transient expression of EGFR 246–253* in NIH 3T3 cells results in overexpression-induced, spontaneous phosphorylation, similar to wt EGFR (Online Resource 1). Unlike V948R, which structurally prevents receptor phosphorylation, the 246–253* mutation apparently does not prevent phosphorylation when EGFR is expressed at sufficiently high concentrations that overcome the inhibitory effect of the ECD. To circumvent this mass-action driven dimerization and phosphorylation, we used RBL-2H3 cells because transient transfections of these cells result in lower protein expression compared to NIH3T3 cells. We confirmed that transfected RBL 2H3 cells do not exhibit basal wt EGFR phosphorylation, and we observed the unstimulated, crosslinked EGFR R1–6 constitutive dimers, similar to those detected in stably expressing NIH 3T3 cells (Online Resource 2 and Fig. 2).
RBL 2H3 cells transiently expressing EGFR 246–253* or EGFR R1–6 246–253* were stimulated with EGF (or not) and chemically crosslinked (or not) with BS3. In these transfected cells both EGFR 246–253* and EGFR R1–6 246–253* fail to form dimers or phosphorylate, regardless of EGF stimulation and chemical crosslinking (Fig. 5, lanes 5–12). We detect phosphorylated monomers and dimers of wt EGFR in response to EGF under these conditions, as expected (Fig. 5, lane 4). These results indicate that at limited expression levels, ECD dimerization is required for phosphorylation of both wt EGFR and EGFR R1–6.

EGFR R1–6 requires ECD dimerization in order to phosphorylate. Whole cell lysates from untransfected RBL-2H3 cells (n/a) or transiently expressing EGFR, EGFR R1–6, EGFR 246–253*, or EGFR R1–6 246–253* were stimulated (or not) with EGF and then chemically crosslinked (or not) with BS3 prior to immunoblotting with the indicated antibodies. M and D indicate monomeric and dimeric EGFR, respectively, as determined by molecular weight markers run on the same gel. Blot shown is representative of four independent experiments. See legend to Fig. 4 for additional comments about gels for transiently transfected cells and β-actin band
4 Discussion
We investigated structural mechanisms whereby EGFR becomes activated in cells, with the purpose of understanding dysregulation of these mechanisms such as occurs with oncogenic mutations. We focused on the intracellular JX domain, which participates in EGFR activation in a manner that is complex and multi-faceted [15, 16]. The JX domain is both required for dimeric receptor activation [16] and necessary for maintaining autoinhibition of unliganded receptors by means of its electrostatically-charged polybasic region [29]. We showed previously that EGFR R1–6 (Fig. 1B) is constitutively activated to initiate downstream signaling by using alanine mutagenesis to reduce the overall charge of the polybasic region [29]. Somehow, this electrostatic perturbation allows EGFR to bypass the initial ligand binding step and proceed directly to spontaneous receptor activation. Our present studies show that this charge-silencing leads to asymmetric dimerization of the TKD domains that are stabilized by the dimerization arms in the ECD, in the same manner as for ligand-dependent activation of wt EGFR.
Canonical EGFR dimerization is initiated by ligand binding, which releases the dimerization arm and thereby enables an outside-in mode of dimerization in which ECD interactions drive the initial steps of EGFR activation, leading to asymmetric conjoining of TKDs and consequent transphosphorylation (Fig. 1A) [13, 31]. Activation of EGFR R1–6 occurs in the absence of ligand, and we set out to investigate the structural rearrangements involved. We hypothesized that this dysregulated mutant operates via an inside-out dimerization mechanism: charge-silencing of the JX polybasic region disrupts its association from the inner leaflet of the plasma membrane, facilitating formation of the intracellular TKD dimer, and this is stabilized by conjoining of the dimerization arms in the ECD. This view is consistent with the previous demonstration with L858R and other oncogenic mutants associated with non-small cell lung cancers, where ECD dimerization was found to be necessary for EGFR activation, and an inside-out model was suggested [39].
Figure 6 depicts an inside-out model to explain ligand-independent EGFR activation caused by charge-neutralization of the JX polybasic region as represented by EGFR R1–6. The electrostatic charge of the corresponding sequence in wt EGFR normally facilitates tight binding between the JX domain and negatively charged lipids in the plasma membrane [7, 24]. Loss of this electrostatic character decreases the affinity of the JX domain for plasma membrane lipids and thereby increases the degrees of motional freedom accessible to the protein. Under these conditions, receptors may spontaneously release their JX domains, and consequently their TKDs, from the plasma membrane (Fig. 6b). Upon contacting another receptor with similar intracellular flexibility, the JX and TKDs may progress through a series of reversible conformational changes (Fig. 6c) until they form an intracellular dimer (Fig. 6d), and this may translate a conformational change to ECDs freeing the dimerization arms for additional stabilization. The TKDs within stabilized receptor dimers are thereby asymmetrically positioned for transphosphorylation and downstream signaling (Fig. 6e).

Proposed EGFR R1–6 activation scheme. Partial charge-silencing of the JX domain (a) leads to spontaneous release of the TKD from the plasma membrane (b). This receptor can encounter another receptor and form an unstable intermediate dimer (c) with the capacity to rearrange into an active dimer (d). The active dimer can then stabilize itself through phosphorylation (e) and recruitment of other signaling proteins
Our findings provide support for components of this model for inside-out activation as follows. Employing chemical crosslinking reagents, we directly observed phosphorylated EGFR R1–6 dimers in the plasma membranes of unstimulated cells (Fig. 2), consistent with the existence of the species proposed in Fig. 6e. In parallel samples, phosphorylated dimers were observed for wt EGFR only with EGF stimulation, whereas EGF caused little or no increase in phosphorylated dimers for EGFR R1–6 (Fig. 2B). We also observed that charge-silencing the JX polybasic region results in phosphorylated dimers of the EGFR(L417H) R1–6, which is retained in the ER, indicating that this functional dimerization occurs readily, independent of membrane localization.
We considered the possibility of phosphorylation in the monomeric species of EGFR R1–6 preceding dimerization, which we could test with the V948R mutation. EGFR R1–6 V948R shows no evidence of phosphorylated monomers or dimers (Fig. 4), consistent with the view that active asymmetric TKD dimers are required for phosphorylation. Thus, phosphorylation occurs after a sequence of conformational rearrangements leading to both dimerization and correct positioning of the TKD. The 246–253* mutation allowed us to test participation of the ECD dimerization arms in receptor activation. That EGFR R1–6 246–253* does not form phosphorylated dimers (Fig. 5) is consistent with the model that the ECD dimerization arms facilitate formation of the asymmetric TKD pair or stabilize this active conformation (Fig. 6d). We conclude that the loss of electrostatic charge in the polybasic region of the JX domain allows the receptor to initiate a series of conformational changes that culminate in an ECD stabilized, TKD functional dimer and consequent trans-phosphorylation. This appears to result from spontaneous release of the TKD from the membrane and thereby release of the autoinhibitory mechanism.
Our data with EGFR R1–6 are consistent with and support previous work with EGFR homodimers and with oncogenic EGFR mutants in determining that active receptors require dimerization of both intracellular and extracellular domains. Homo- and hetero-receptor dimerization within the ErbB family of receptor tyrosine kinases has emerged as a major component of oncogenic signaling [36]. Heterodimers, in particular, have been implicated as a means of chemoresistance in many cancers as they are often able to compensate for vulnerabilities in each of the component receptors that a homodimer may not [5]. Mixed therapies, which simultaneously target multiple members of the ErbB family, have been shown to have greater efficacy than therapies targeting single receptors [34]. Interestingly, therapies targeting the ErbB family’s conserved JX domain to prevent receptor activation have begun emerging as new potential therapeutics [22, 23] with the potential to mitigate the oncogenic resilience of these heterodimers. Improving our understanding of receptor activation, structure, and function within cells is necessary to continue advancing these strategies. Our study contributes to this understanding by linking the polybasic region of the JX domain to structural requirements for EGFR activation and by providing a better understanding of the mechanism by which this segment regulates the activation process.
Data Availability
The data, expression plasmids, and cell lines used in this study are available upon reasonable request.
Code Availability
Software application or custom code: not applicable.
Abbreviations
- EGFR:
-
Epidermal growth factor receptor
- ECD:
-
Extracellular domain
- JX:
-
Juxtamembrane
- TKD:
-
Tyrosine kinase domain
- RTK:
-
Receptor tyrosine kinase
- EGFR R1–6:
-
EGFR JX charge-reduced mutant
- EGFR L417H:
-
ER-retained EGFR mutant
- EGFR 246–253*:
-
EGFR ECD domain III mutant that cannot form ECD dimers
- EGFR V948R:
-
EGFR TKD mutant that cannot form asymmetric TKD dimers
- DSS:
-
Disuccinimidyl suberate intracellular crosslinker
- BS3 :
-
Bis(sulfosuccinimidyl)suberate extracellular crosslinker
References
Lemmon MA, Schlessinger J (2010) Cell signaling by receptor tyrosine kinases. Cell 141:1117–1134. https://doi.org/10.1016/j.cell.2010.06.011
Endres NF, Barros T, Cantor AJ, Kuriyan J (2014) Emerging concepts in the regulation of the EGF receptor and other receptor tyrosine kinases. Trends Biochem Sci 39:437–446. https://doi.org/10.1016/j.tibs.2014.08.001
Lemmon M, Schlessinger J, Ferguson KM (2014) The EGFR family: not so prototypical receptor tyrosine kinases. Cold Spring Harb Perspect Biol 6:a020768. https://doi.org/10.1101/cshperspect.a020768
Zhang H, Berezov A, Wang Q et al (2007) ErbB receptors: from oncogenes to targeted cancer therapies. J Clin Invest 117:2051–2058. https://doi.org/10.1172/JCI32278
Shan Y, Eastwood MP, Zhang X et al (2012) Oncogenic mutations counteract intrinsic disorder in the EGFR kinase and promote receptor dimerization. Cell 149:860–870. https://doi.org/10.1016/j.cell.2012.02.063
Chu CT, Everiss KD, Wikstrand CJ et al (1997) Receptor dimerization is not a factor in the signalling activity of a transforming variant epidermal growth factor receptor (EGFRvIII). Biochem J 324:855–861. https://doi.org/10.1042/bj3240855
Arkhipov A, Shan Y, Das R et al (2013) Architecture and membrane interactions of the EGF receptor. Cell 152:557–569. https://doi.org/10.1016/j.cell.2012.12.030
Sliwkowski MX, Schaefer G, Akita RW et al (1994) Coexpression of erbB2 and er6B3 proteins reconstitutes a high affinity receptor for heregulin. J Biol Chem 269:14661–14665. https://doi.org/10.1002/art.30521
Zhang X, Gureasko J, Shen K et al (2006) An allosteric mechanism for activation of the kinase domain of epidermal growth factor receptor. Cell 125:1137–1149. https://doi.org/10.1016/j.cell.2006.05.013
Huse M, Kuriyan J (2002) The conformational plasticity of protein kinases. Cell 109:275–282. https://doi.org/10.1016/S0092-8674(02)00741-9
Jura N, Zhang X, Endres NF et al (2011) Catalytic control in the EGF receptor and its connection to general kinase regulatory mechanisms. Mol Cell 42:9–22. https://doi.org/10.1016/j.molcel.2011.03.004
Hubbard SR, Till JH (2000) Protein tyrosine kinase structure and function. Annu Rev Biochem 69:373–398. https://doi.org/10.1146/annurev.biochem.69.1.373
Kancha RK, von Bubnoff N, Duyster J (2013) Asymmetric kinase dimer formation is crucial for the activation of oncogenic EGFRvIII but not for ERBB3 phosphorylation. Cell Commun Signal 11:39. https://doi.org/10.1186/1478-811X-11-39
Hedger G, Sansom MSPP, Koldsø H (2015) The juxtamembrane regions of human receptor tyrosine kinases exhibit conserved interaction sites with anionic lipids. Sci Rep 5:1–10. https://doi.org/10.1038/srep09198
Doerner A, Scheck R, Schepartz A (2015) Growth factor identity is encoded by discrete coiled-coil rotamers in the EGFR juxtamembrane region. Chem Biol 22:776–784. https://doi.org/10.1016/j.chembiol.2015.05.008
He L, Hristova K (2012) Consequences of replacing EGFR juxtamembrane domain with an unstructured sequence. Sci Rep 2:854. https://doi.org/10.1038/srep00854
Pahuja KB, Nguyen TT, Jaiswal BS et al (2018) Actionable activating oncogenic ERBB2/HER2 transmembrane and juxtamembrane domain mutations. Cancer Cell 34:792-806.e5. https://doi.org/10.1016/j.ccell.2018.09.010
Huang Y, Bharill S, Karandur D et al (2016) Molecular basis for multimerization in the activation of the epidermal growth factor receptor. Elife 5:1–27. https://doi.org/10.7554/eLife.14107
Trenker R, Jura N (2020) Receptor tyrosine kinase activation: from the ligand perspective. Curr Opin Cell Biol 63:174–185. https://doi.org/10.1016/j.ceb.2020.01.016
Cai G, Zhu L, Chen X et al (2018) TRAF4 binds to the juxtamembrane region of EGFR directly and promotes kinase activation. Proc Natl Acad Sci USA 115:11531–11536. https://doi.org/10.1073/pnas.1809599115
Xue F, An C, Chen L et al (2019) 4.1B suppresses cancer cell proliferation by binding to EGFR P13 region of intracellular juxtamembrane segment. Cell Commun Signal 17:1–16. https://doi.org/10.1186/s12964-019-0431-6
Mitchell RA, Luwor RB, Burgess AW (2018) Epidermal growth factor receptor: Structure-function informing the design of anticancer therapeutics. Exp Cell Res 371:1–19. https://doi.org/10.1016/j.yexcr.2018.08.009
Gerhart J, Thévenin AF, Bloch E et al (2018) Inhibiting epidermal growth factor receptor dimerization and signaling through targeted delivery of a juxtamembrane domain peptide mimic. ACS Chem Biol 13:2623–2632. https://doi.org/10.1021/acschembio.8b00555
Abd Halim KB, Koldsø H, Sansom MSPP (2015) Interactions of the EGFR juxtamembrane domain with PIP2-containing lipid bilayers: insights from multiscale molecular dynamics simulations. Biochim Biophys Acta - Gen Subj 1850:1017–1025. https://doi.org/10.1016/j.bbagen.2014.09.006
Gambhir A, Hangyás-Mihályné G, Zaitseva I et al (2004) Electrostatic sequestration of PIP2 on phospholipid membranes by basic/aromatic regions of proteins. Biophys J 86:2188–2207. https://doi.org/10.1016/S0006-3495(04)74278-2
Maeda R, Sato T, Okamoto K et al (2018) Lipid-protein interplay in dimerization of juxtamembrane domains of epidermal growth factor receptor. Biophys J 114:893–903. https://doi.org/10.1016/j.bpj.2017.12.029
Endres NF, Das R, Smith AW et al (2013) Conformational coupling across the plasma membrane in activation of the EGF receptor. Cell 152:543–556. https://doi.org/10.1016/j.cell.2012.12.032
Jura N, Endres NF, Engel K et al (2009) Mechanism for activation of the EGF receptor catalytic domain by the juxtamembrane segment. Cell 137:1293–1307. https://doi.org/10.1016/j.cell.2009.04.025
Bryant KL, Antonyak MA, Cerione RA et al (2013) Mutations in the polybasic juxtamembrane sequence of both plasma membrane- and endoplasmic reticulum-localized epidermal growth factor receptors confer ligand-independent cell transformation. J Biol Chem 288:34930–34942. https://doi.org/10.1074/jbc.M113.513333
Ward CW, Garrett TPJ (2001) The relationship between the L1 and L2 domains of the insulin and epidermal growth factor receptors and leucine-rich repeat modules. BMC Bioinform 2:4. https://doi.org/10.1186/1471-2105-2-4
Dawson JP, Berger MB, Lin C et al (2005) Epidermal growth factor receptor dimerization and activation require ligand-induced conformational changes in the dimer interface. Mol Cell Biol 25:7734–7742. https://doi.org/10.1128/MCB.25.17.7734-7742.2005
Gosse JJ, Wagenknecht-Wiesner A, Holowka David et al (2005) Transmembrane sequences are determinants of immunoreceptor signaling. J Immunol 175:2123–31
Wakefield DL, Holowka D, Baird B (2017) The FcεRI signaling cascade and integrin trafficking converge at patterned ligand surfaces. Mol Biol Cell 28:3383–3396. https://doi.org/10.1091/mbc.E17-03-0208
Pines G, Köstler WJ, Yarden Y (2010) Oncogenic mutant forms of EGFR: lessons in signal transduction and targets for cancer therapy. FEBS Lett 584:2699–2706. https://doi.org/10.1016/j.febslet.2010.04.019
Chakraborty S, Li L, Puliyappadamba VT et al (2014) Constitutive and ligand-induced EGFR signalling triggers distinct and mutually exclusive downstream signalling networks. Nat Commun 5:5811. https://doi.org/10.1038/ncomms6811
Wang Z, Longo PA, Tarrant MK et al (2011) Mechanistic insights into the activation of oncogenic forms of EGF receptor. Nat Struct Mol Biol 18:1388–1393. https://doi.org/10.1038/nsmb.2168
Poger D, Mark AE (2014) Activation of the epidermal growth factor receptor: a series of twists and turns. Biochemistry 53:2710–2721. https://doi.org/10.1021/bi401632z
Valley CC, Arndt-Jovin DJ, Jovin TM et al (2015) Inside-out signaling of oncogenic EGFR mutants promotes ligand-independent dimerization. Biophys J 108:351a. https://doi.org/10.1016/j.bpj.2014.11.1921
Valley CC, Arndt-Jovin DJ, Karedla N et al (2015) Enhanced dimerization drives ligand-independent activity of mutant EGFR in lung cancer. Mol Biol Cell. https://doi.org/10.1091/mbc.E15-05-0269
Cochet C, Kashles O, Chambaz EM et al (1988) Demonstration of epidermal growth factor-induced receptor dimerization in living cells using a chemical covalent cross-linking agent. J Biol Chem 263:3290–3295
Coban O, Zanetti-Dominguez LC, Matthews DR et al (2015) Effect of phosphorylation on EGFR dimer stability probed by single-molecule dynamics and FRET/FLIM. Biophys J 108:1013–1026. https://doi.org/10.1016/j.bpj.2015.01.005
Di Fiore PP, Pierce JH, Fleming TP et al (1987) Overexpression of the human EGF receptor confers an EGF-dependent transformed phenotype to NIH 3T3 cells. Cell 51:1063–1070. https://doi.org/10.1016/0092-8674(87)90592-7
Holowka D, Baird B (2017) Mechanisms of epidermal growth factor receptor signaling as characterized by patterned ligand activation and mutational analysis. Biochim Biophys Acta - Biomembr 1859:1430–1435. https://doi.org/10.1016/j.bbamem.2016.12.015
Acknowledgements
We thank Dr. Maurine Linder (Cornell University) for the use of her VersaDoc MP 5000 imaging system. We appreciate support of these studies by the National Institutes of Health (grants cited elsewhere); the content of this paper is solely the responsibility of the authors.
Funding
This study was supported by the National Institutes of Health (Grant Nos. R01 GM117552 and R01 AI022449).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
NIH 3T3 cells transiently expressing wt-EGFR are constitutively active. NIH 3T3 cells were transiently transfected with wt-EGFR or EGFR R1–6, EGF stimulated (or not) and chemically crosslinked with BS3, lysed, and immunoblotted for EGFR pY1173, total EGFR, and β-actin. Constitutive activation in the absence of EGF (lane 1) is seen in unstimulated cells expressing wt EGFR, likely due to overexpression. The appearance of double band for β-actin is probably due to nonspecific binding to another cell component of slightly higher molecular weight. Supplementary file 1 (TIF 12872 KB)
Online Resource 2
RBL-2H3 cells transiently expressing wt EGFR at low levels do not exhibit constitutive activation. RBL 2H3 cells were transiently transfected with wt-EGFR or EGFR R1–6, EGF stimulated (or not) and chemically crosslinked or not with BS3, lysed, and immunoblotted for EGFR pY1173, total EGFR, and β-actin. There is no constitutive activation in unstimulated cells expressing wt EGFR in the absence of EGF (lane 6); after addition of EGF phosphorylated dimers are trapped by chemical crosslinking (lanes 3-5). Supplementary file 2 (TIF 18105 KB)
Rights and permissions
About this article

Cite this article
Mohr, J.D., Wagenknecht-Wiesner, A., Holowka, D.A. et al. Basic Amino Acids Within the Juxtamembrane Domain of the Epidermal Growth Factor Receptor Regulate Receptor Dimerization and Auto-phosphorylation. Protein J 39, 476–486 (2020). https://doi.org/10.1007/s10930-020-09943-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10930-020-09943-8