In this contribution we describe a partial dentary with teeth and an astragalus referred to the ancient megatheriine Megathericulus patagonicus Ameghino, 1904 (Xenarthra, Tardigrada) recovered from the Collón Curá Formation (Miocene) at Chubut Province, Argentina. Referral of the new specimens is based on metric and morphological similarities with the holotype and specimens that have been referred to this species. Shared features include: 1) overall size, smaller than other Megatheriinae such Eomegatherium Kraglievich, 1926, Pliomegatherium Kraglievich, 1930, and Pyramiodontherium (Rovereto, 1914); 2) shape of the molariforms, molariforms mesiodistally compressed and rectangular in outline; 3) Hypsodonty Index close to 100; 4) position of the navicular facet, two-thirds of the navicular facet projects dorsal to the discoid facet; 5); shape of the ventral portion of the fibular facet, the ventral portion has subtriangular outline and contacts the ectal facet; 6) in the astragalus, presence of a sesamoid facet. Considering the large intraspecific differences exhibited in Megatheriinae, the similarities and few differences between Megathericulus primaevus Cabrera, 1939, and M. patagonicus allow us to consider M. primaevus as a junior synonym of M. patagonicus. The presence of Megathericulus and ‘Xyophorus’ Ameghino, 1887 (Mammalia, Xenarthra, Nothrotheriinae) in some distant paleofauna associations (Cerro Zeballos, Argentina, and Quebrada Honda, Bolivia) would suggest a similar path of dispersion and paleoenvironment requirements of both taxa. The presence of these two genera in mentioned localities supports that the Quebrada Honda fauna as a whole is more similar to the asynchronous high-latitude faunas from Patagonia than to contemporary La Venta Fauna (Colombia).
During the middle Miocene of Patagonia (southernmost of South America) several main successive mammal faunas have been traditionally identified (Friasian, Colloncuran, Mayoan; see Flynn and Swisher 1995). The Friasian faunas are recorded from the Río Frías Formation, Chile (Marshall and Salinas 1990; Bostelmann et al. 2012), the Colloncuran fauna from the Collón Curá Formation, Argentina (Bondesio et al. 1980; Vucetich et al. 1993), and the Mayoan fauna from the El Portezuelo, Pedregoso, and Río Mayo formations, Argentina (Scillato-Yané and Carlini 1998; De Iuliis et al. 2008; González Ruiz et al. 2017). The operational use of these Miocene faunas as ages, South American Land Mammal Ages (SALMAS) or local faunas and the relationships between them is still unclear, and has been debated by several authors (see Marshall and Salinas 1990; Vucetich et al. 1993; Flynn and Swisher 1995; Madden et al. 1997; Croft et al. 2009, 2016).
We focus here on fossils from the Collón Curá Formation (15.8 Ma to ca. 11.2 Ma, Burdigalian-Tortonian) (see Brandoni et al. 2018: table 2), which overlaps temporally, at least partially, with the Río Frías Formation (ca. 16.5 Ma to ca. 14.8 Ma, Burdigalian-Langhian) (Flynn and Swisher 1995; de la Cruz and Cortés 2011; Encinas et al. 2016), and with the El Portezuelo, Pedregoso, and Río Mayo formations (ca. 13.5 Ma to ca.11.8 Ma, Serravallian-Tortonian) (Dal Molín and Franchi 1996; Madden et al. 1997; De Iuliis et al. 2008; Dunn et al. 2015; Folguera et al. 2018). Several mammalian genera (e.g., the marsupial Abderites Ameghino, 1887, the xenarthra Eonaucum Scillato-Yané and Carlini, 1998, the notoungulate Protypotherium Ameghino, 1885) were recorded from Collón Curá Formation localities in Neuquén and Río Negro provinces, Argentina (e.g., Collón Curá River, Paso Flores, and Pilcaniyeu), although some of them without precise geographic and stratigraphic information (see Pascual et al. 1984; Bondesio et al. 1980; Kramarz et al. 2011; Vera et al. 2017). Other records (e.g., the marsupial sparassodont Pseudonotictis Marshall, 1981, the glyptodontid Paraeucinepeltus González Ruiz et al., 2011, Protypotherium) from the Collón Curá Formation at their southernmost outcrops correspond to those from the locality of Cerro Zeballos (Chubut Province) (Martin and Tejedor 2007; González Ruiz et al. 2011; Brandoni et al. 2017; Vera et al. 2018).
Among the Tardigrada (=Folivora, =Phyllophaga), the records of the Megatheriinae Megathericulus Ameghino, 1904, for the Collón Curá Formation correspond to M. primaevus Cabrera, 1939, at Cañadón Ftamichi (near Paso Flores, Río Negro Province) (Cabrera 1939), and Megathericulus sp. for Neneo Rucá (near Comallo, Río Negro Province) (Casamiquela 1969) and at Collón Curá River (type locality of Collón Curá Formation, Neuquén Province) (Bondesio et al. 1980).
Megathericulus, described as M. patagonicus Ameghino, 1904, was first recorded from ‘Tehuelche antiguo del Chubut (Laguna Blanca)’ (Ameghino 1904: 282). Several other remains referred to M. patagonicus have been recovered from the Río Mayo Formation of Chubut and Santa Cruz provinces, Argentina (Brandoni 2006a; De Iuliis et al. 2008), and Quebrada Honda, Bolivia (Brandoni et al. 2018). Other remains of Megathericulus include Megathericulus sp. recorded from the middle Miocene of Peruvian Amazonia (Pujos et al. 2013). Among Megatheriinae, M. patagonicus is characterized by a long predental portion of the palate, the presence of molariforms with rectangular outline, humerus with a developed deltopectoral crest, navicular facet of the astragalus dorsally located with respect to the discoid facet, among others (see De Iuliis et al. 2008; Brandoni et al. 2018). The shape of the molariforms of M. patagonicus was described by Brandoni et al. (2018) for some molariforms of the upper series; De Iuliis et al. (2008) and Pujos et al. (2013) have inferred the shape of the lower molariforms from the shape of the alveoli of two edentulous dentaries.
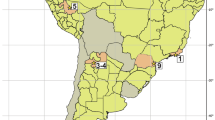
The aim of this contribution is to describe a partial dentary with teeth and an astragalus referred to M. patagonicus recovered from the Collón Curá Formation at the southernmost localities of La Gloria and Cerro Zeballos, respectively, Chubut Province, Argentina (Fig. 1). In addition, we discuss the systematic, chronologic, biogeographic, and evolutionary importance of this finding.
Fig. 1
Location map of Collón Curá Formation with indication of the localities of La Gloria and Cerro Zeballos (black triangles) and the dated localities. Modified from Brandoni et al. (2017)
LIEB-PV, Laboratorio de Investigaciones en Evolución y Biodiversidad (PV, Paleovertebrados), Esquel, Argentina; MACN, Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’, Buenos Aires, Argentina; MLP, Museo de La Plata, La Plata, Argentina; MUSM, Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Lima, Perú; PVL, Paleontología Vertebrados Lillo, Facultad de Ciencias e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Argentina; UATF-V, Universidad Autónoma Tomás Frías-Vertebrate Paleontology Collection, Potosí, Bolivia.
Material and Methods
The following species and specimens were used for comparison: Megathericulus patagonicus, MACN A 11151 (Laguna Blanca, Ameghino 1904), MLP 91-IX-7-18 (Río Mayo Formation; Santa Cruz Province; De Iuliis et al. 2008), MLP 92-XI-15-2 (Río Mayo Formation; Chubut Province; De Iuliis et al. 2008); Megathericulus primaevus, MLP 39-VI-24-1 (Collón Curá Formation, Río Negro Province; Cabrera 1939); Eomegatherium nanum Kraglievich, 1926, MACN PV 4992 (‘Mesopotamiense’, Lower Member of Ituzaingó Formation, Entre Ríos Province; Brandoni 2013; Brunetto et al. 2013); ‘Eomegatherium’ cabrerai Kraglievich, 1930, MLP 2–206 (‘Yacimiento de Laguna Blanca y río Fénix’, Kraglievich 1930); Pliomegatherium lelongi Kraglievich, 1930, MACN PV-13213, MLP 99-XI-1-1 (‘Mesopotamiense’, Lower Member of Ituzaingó Formation, Entre Ríos Province; Brandoni 2006b, 2013; Brunetto et al. 2013); Pyramiodontherium bergi (Moreno and Mercerat, 1891), MLP 2–66 (‘Araucanense’, Catamarca Province; Carlini et al. 2002); Pyramiodontherium brevirostrum Carlini et al., 2002, MLP 31-XI-12-25 (‘Araucanense’, Catamarca Province; Carlini et al. 2002); Pyramiodontherium scillatoyanei De Iuliis et al., 2004, MLP 68-III-14-1 (upper levels of the Lower Member of Toro Negro Formation; La Rioja Province; De Iuliis et al. 2004); Anisodontherium sp. Brandoni and De Iuliis, 2007, PVL 6425 (Saladillo Formation, Tucumán Province; Brandoni et al. 2012); A. halmyronomum (Cabrera, 1928), MLP-30-XII-10-21 (Arroyo Chasicó Formation, Buenos Aires Province; Cabrera 1928; Brandoni and De Iuliis 2007); Megatheriops rectidens Rovereto, 1914, MACN PV 2818 (Huayquerías de San Carlos, Mendoza Province, Rovereto 1914).
Anatomical Abbreviations
DD, depth of dentary; HI, Hypsodonty Index; m1–m4, molariforms1–4; po, posterolateral opening of mandibular canal; TRL, tooth row length.
For chronological purposes, we follow the International Chronostratigraphic Chart (International Commission on Stratigraphy 2018; Cohen et al. 2013).
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study. All relevant information is included in the article itself.
Geological Setting
The Miocene Collón Curá Formation was accumulated in the extra Andean Patagonian region (Fig. 1) in a broken foreland system (Patagonian Broken Foreland; Bilmes et al. 2013; Ramos et al. 2015; Bucher et al. 2018). This system is characterized by several sedimentary basins partially or totally disconnected from each other (e.g., Collón Curá, Ñirihuau, Paso del Sapo Basin, and Gastre, among others), bounded by N-S to NW-SE trending block-mountains heights (Bilmes et al. 2013; Bucher et al. 2018). The Collón Curá Formation is composed mainly of volcaniclastic limestones and sandstones accumulated in several continental environments, ranging from alluvial to lacustrine depositional systems (Ramos et al. 2011; Bilmes et al. 2014; Bucher et al. 2018). The specimens described in this contribution were collected from two different localities, La Gloria and Cerro Zeballos, both located in northwestern Chubut Province, Argentina (Fig. 1).
La Gloria Locality (42° 50′54.79”S, 69° 27′41.06”W)
La Gloria locality constitutes the southernmost exposures of the Collón Curá Formation included in the Paso del Sapo Basin (Bucher et al. 2018). This locality shows a ca. 135 m thick, continuous, and homoclinal succession of the Collón Curá Formation (Bucher et al. 2018). The deposits of Collón Curá Formation exposed in La Gloria are constituted by brownish to yellowish white volcaniclastic to epiclastic conglomerates, sandstones, and limestones, very friable and powdery with development of paleosols (Fig. 2a). The analyzed section starts with 45 m of massive and poorly sorted medium to coarse grained tuffaceous sandstones, with a dominantly volcaniclastic composition. These deposits show mainly tabular geometries with minor intercalated lenticular bodies that registered sedimentary unidirectional structures. Over these deposits, a 40 m thick volcaniclastic sandstone to limestones succession is registered. These deposits form tabular and subordinately lenticular bodies, massive or with sedimentary unidirectional structures poorly preserved. Finally, 50 m of conglomerate to coarse sandstones are disposed (Bucher et al. 2018). These sediments shows a volcaniclastic-epiclastic mixed composition and are represented by massive tabular bodies and by lenticular levels with sedimentary unidirectional structures. Throughout the section, fossil vertebrates are abundant, and specimen LIEB-PV 6152 was collected from the middle sandstone to limestone dominated succession (Fig. 2a). The presence of massive and poorly sorted tabular bodies with paleosol development combined with lenticular and sedimentological structured deposits suggest an alluvial to fluvial depositional system, dominated by alluvial to fluvial plains with minor fluvial channels development (Bucher et al. 2018).
Fig. 2
Stratigraphic section of Collón Curá Formation at Chubut Province, Argentina. a La Gloria; b Cerro Zeballos
The Collón Curá deposits in Cerro Zeballos are located very close and immediately to the east of the deposits included in the Ñirihuau Basin (Cazau 1980; Ramos et al. 2015). The continuity of the outcrops and the lithological similarities with the western-contiguous outcrops allow us to include these deposits as the easternmost exposures of the Ñirihuau Basin. The analyzed section starts with a 15 m thick of massive ocher or yellowish white tuffaceous sandstone, in general very friable and powdery. Tabular bodies with concentrations of reddish concretions composed by tuffitic material frequently appear. These levels show the presence of roots and scarce paleosol development. Fossil vertebrates are abundant throughout the section, even inside the concretions. Specimen LIEB-PV 6153 was collected in the middle of this 15 m section (Fig. 2b). The profile continues with 3 m thick of greenish tuffaceous limestones with intercalated levels of ocher tuffaceous and massive fine sandstones. Above this, the profile continues with volcaniclastic to epiclastic deposits of claystone, mudstone, and grainstones, with some pyroclastic subordinated levels intercalated (Brandoni et al. 2017). The presence of mainly massive and tabular sandstones with scarce paleosol development indicates a shallow and low energy system, probably related with alluvial to fluvial plains located close to shallow lagoons or lakes (Brandoni et al. 2017).
Type species: Megathericulus patagonicus Ameghino, 1904.
Geographic and stratigraphic distribution: Laguna Blanca, southwestern Chubut Province, Argentina; Cañadón Ftamichi, near Paso Flores, Río Negro Province, Argentina (Collón Curá Formation); Cerro Guenguel, northwestern Santa Cruz Province, Argentina (Río Mayo Formation); Arroyo El Pedregoso, southwestern Chubut Province, Argentina (Río Mayo Formation); SEP 07 locality, southeastern Peru (Ipururo Formation); Papachacra and Huayllajara local faunas, southern Bolivia (unnamed formation of the Honda Group) (see Ameghino 1904; Kraglievich 1930; Cabrera 1939; Bondesio et al. 1980; Scillato-Yané and Carlini 1998; De Iuliis et al. 2008; Pujos et al. 2013; Brandoni et al. 2018); La Gloria and Cerro Zeballos, Chubut Province (Collón Curá Formation) (this paper).
Comments: The records of Megathericulus cited by Casamiquela (1969) and Bondesio et al. (1980) for Collón Curá Formation (Neneo Rucá and Collón Curá River respectively) are not included (see discussion).
Megathericulus patagonicus Ameghino, 1904
Megathericulus primaevus Cabrera, 1939: 475–483, Figs. 1–3. New synonymy.
Fig. 3
Dentary of Megathericulus patagonicus. a–c LIEB-PV 6152 in occlusal, lateral and medial view. d–e MLP 91-IX-7-18 in occlusal and lateral view. Anatomical abbreviations: m1–m4, molariforms1–4; po, posterolateral opening of mandibular canal
Holotype: MACN A 11151, anterior portion of cranium with edentulous palate and complete right astragalus.
Geographic and stratigraphic distribution: Laguna Blanca, southwestern Chubut Province, Argentina; Cerro Guenguel, northwestern Santa Cruz Province, Argentina (Río Mayo Formation); Arroyo El Pedregoso, southwestern Chubut Province, Argentina (Río Mayo Formation); Papachacra and Huayllajara local faunas, southern Bolivia (unnamed formation of the Honda Group) (see Ameghino 1904; Kraglievich 1930; De Iuliis et al. 2008; Brandoni et al. 2018); La Gloria and Cerro Zeballos, Chubut Province (Collón Curá Formation) (this paper).
Referred material. LIEB-PV 6152, partial left dentary with molariforms (Fig. 3a-c); LIEB-PV 6153, complete right astragalus (Fig. 4a-c).
Fig. 4
Astragalus of Megathericulus patagonicus. a–c LIEB-PV 6153 in proximal, medial and distal view; d–f MACN A 11151 (type of M. patagonicus) in proximal, medial and distal view; g–i: MLP 39-VI-24-1 (type of M. primaevus) in proximal, medial and distal view
Geographic and stratigraphic provenance: LIEB-PV 6152 comes from La Gloria and LIEB-PV 6153 comes from Cerro Zeballos, Chubut Province, Argentina. Collón Curá Formation.
Description
Dentary: The dentary assigned to M. patagonicus (LIEB-PV 6152) is the central portion of the horizontal ramus of the left dentary with four molariforms, lacking the predental portion, left articular condyle, and angular process (Fig. 3a-c). The molariforms are arranged in a continuous series, without diastema between the first and the second teeth (Fig. 3a). Considering the length of the tooth row (LIEB-PV 6152: 69.2 mm), LIEB-PV 6152 is slightly larger than the specimen referred to M. patagonicus (MLP 91-IX-7-18: 62 mm), but smaller than Megathericulus sp. (MUSM 1564: 84.6 mm), Anisodontherium sp. (PVL 6425 right: 94 mm), A. halmyronomum (MLP-30-XII-10-21, right: 115 mm), Pliomegatherium lelongi (MACN PV-13213: 135 mm), and Pyramiodontherium bergi (MLP 2–66: 195 mm) (Table 1).
Table 1 Measurements (in mm) of the dentary of Megatheriinae. TRL, tooth row length; DD, depth of the dentary; HI, Hypsodonty Index
In lateral view (Fig. 3b), the convexity of the ventral margin of the dentary is marked by the margin’s ascent posterior to the base of the m4 alveolus, where it outlines a notch between the level of the alveolus and the anterior portion of the base of the angular process. This region closely resembles that in Megathericulus sp. and A. halmyronomum (see Pujos et al. 2013: fig. 3a; Brandoni and De Iuliis 2007: fig. 3a); this area is missing in M. patagonicus (MLP 91-IX-7-18; Fig. 3d, e).
Considering the small preserved ventral portion of the coronoid process, the m4 lies anterior to the anterior margin of the process and is entirely visible in lateral view. In M. patagonicus (MLP 91-IX-7-18), Megathericulus sp. (see Pujos et al. 2013: fig. 3a), A. halmyronomum (see Brandoni and De Iuliis 2007: fig. 3a), and Anisodontherium sp. (Brandoni et al. 2012: fig. 2a, b), the m4 lies anterior to the anterior margin of the coronoid process. In other megatheriines, the m4 is posterior with respect to the coronoid process although there is variation in the position: in Pl. lelongi the distal margin of the m4 nearly coincides with the plane of the anterior margin of the coronoid process (Brandoni 2006b: figs. 3.1–4); in Megatheriops rectidens, Py. bergi, and Py. brevirostrum (see Carlini et al. 2002: fig. 2) only the mesial half of the m4 is visible in lateral view, whereas in several specimens of Megatherium americanum Cuvier, 1796, the m4 is entirely posterior to the anterior margin of coronoid process and hidden from lateral view.
In LIEB-PV 6152 the spout of the mandible is not preserved and it is not possible to determinate the exact position of the posterior margin of the mandibular symphysis with respect to the anterior margin of the m1. Judging by the preserved portion, it seems like the margin is at least anterior to the middle portion of the m1. In M. patagonicus (MLP 91-IX-7-18, Fig. 3d) the posterior margin of the mandibular symphysis is located ventromedial and anterior to the level of m1, as also occurs in A. halmyronomum (see Brandoni and De Iuliis 2007: fig. 3b), ‘Eomegatherium’ andinum Kraglievich, 1930, and Eo. nanum (see Kraglievich 1930: fig. 5a). The position of the posterior margin of the mandibular symphysis in other megatheriines varies: in Py. bergi and Eremotherium laurillardi (Lund, 1842), it reaches approximately to a point halfway along m1. It is even farther posterior, reaching the plane of the alveolar septum between m1 and m2, in Py. brevirostrum (see Carlini et al. 2002: fig. 2), Megatherium altiplanicum Saint-André and De Iuliis, 2001 (see Saint-André and De Iuliis 2001: fig. 4b), and Megatherium tarijense Gervais and Ameghino, 1880 (see De Iuliis et al. 2009: fig. 2d). Finally, the posterior margin of the mandibular symphysis generally reaches to a point halfway along the m2 in Megatherium americanum and Megatherium gallardoi Ameghino and Kraglievich, 1921 (see Brandoni et al. 2008: fig. 2d), although in Megatherium americanum, it occasionally also lies at the level of the m1/m2 alveolar septum.
The most notable feature of the dentition of LIEB-PV 6152 is that the molariforms are mesiodistally compressed (specially the m2 and m3; Fig. 3a, Table 2) rather than isodiametric (length and width of each molariform almost equal). In Megathericulus sp. (MUSM 1564), M. patagonicus (MLP 91-IX-7-18), and ‘Eo.’ andinum teeth are unknown, but mesiodistally compressed molariforms are inferred from their alveoli (see De Iuliis et al. 2008; Pujos et al. 2013). In Py. bergi and Py. brevirostrum, Megatheriops rectidens, Pl. lelongi, and the species of Megatherium Cuvier, 1796, and Eremotherium Spillmann, 1948, the molariforms are isodiametric. In LIEB-PV 6152, the m1 is nearly trapezoidal in cross-section with a posterior half wider than the anterior one; its occlusal surface is not preserved. The m2 is rectangular to subtrapezoidal in outline; although the occlusal surface is damaged, two lophids separated by a V-shaped valley between them are observable. The m3 is complete, it is clearly rectangular in outline, and possesses two lophids separated by a V-shaped valley between them. In LIEB-PV 6152, as in A. halmyronomum and Anisodontherium sp., the V-shaped valley is deep, whereas in other taxa (e.g., Pyramiodontherium, Megatheriops, Megatherium) the valley is not markedly deep as in M. patagonicus or Anisodontherium. As in other megatheriines, the m2 and m3 have lingual and labial longitudinal grooves (Fig. 3). The m4 is nearly trapezoidal in outline with the anterior half wider than the posterior one. In A. halmyronomum and Anisodontherium sp. the outline of the molariforms is similar to that described for LIEB-PV 6152, whereas in Py. bergi, the anterior and posterior crests of the m1 and m2 are inclined with respect the anteroposterior axis, being more trapezoidal or parallelogram-shaped in cross-section. In Megatherium and Eremotherium each molariform is nearly squared in cross-section.
Table 2 Measurements (in mm) of lower molariforms of Megathericulus patagonicus. m1–m4, molariforms 1–4
The Hypsodonty Index (HI), calculated as greatest depth of the dentary / length of the molariform tooth row × 100, has been used as an indicator of hypsodonty of the molariforms in sloths (see Zetti 1964; Bargo et al. 2006; Brandoni and De Iuliis 2007; Fields 2009). For LIEB-PV 6152, the HI is 102, 92 in Megathericulus sp., 99 in Anisodontherium sp. (taking into account an average of the measurements of both dentaries), and 107 for A. halmyronomum. The HI for Py. bergi is 77, being 85 in Py. brevirostrum, 88 in Megatheriops rectidens, and 75 in Pl. lelongi (Table 1). Among Quaternary megatheriines, HI is greatest in Megatherium americanum, in which it varies between 92 and 112 (Saint-André and De Iuliis 2001). Considering these values, the HI for M. patagonicus is more similar to that of Megatherium americanum and Anisodontherium, than to the values of other Neogene megatheriines (e.g., Pliomegatherium, Pyramiodontherium, Megatheriops).
Astragalus: Although smaller (Table 3), the astragalus of M. patagonicus (Fig. 4) is similar in form to that of other megatheriines (see Brandoni et al. 2004; De Iuliis et al. 2008). The fibular facet is divided into two portions, an anteroposteriorly elongate dorsal division that joins the discoid facet, and a ventral portion with a rather subtriangular surface that is extended ventrally and contacts the ectal facet (Fig. 4a–g). In the rest of the Megatheriinae the ectal facet does not contact the ventral portion of the fibular facet, and the outline is different: in Pl. lelongi, Py. bergi, and Py. scillatoyanei the ventral portion is subcircular to oval in outline, whereas in ‘Eo.’ cabrerai it is subquadrangular. The posterior separation between the ectal and discoid facets resembles that in Py. scillatoyanei and Py. bergi. The posterior part of the discoid facet is abbreviated and at an obtuse angle relative to the odontoid facet in M. patagonicus (Fig. 4a–g), whereas the angle is less obtuse in Pl. lelongi and Eo. nanum, and it is nearly straight in Py. bergy and Py. scillatoyanei.
Table 3 Measurements (in mm) of the astragalus of Megatheriinae. Length is equal to the distance between the navicular facet and the posterior border of the discoid facet; width is equal to the distance between the lateral border of the fibular facet and the medial border of the odontoid facet
The discoid facet is an approximately semicircular surface, slightly convex in both its major axes, particularly near its perimeter. The odontoid facet is semicylindrical in outline and lies on the dorsolateral half of the tall, peg-shaped odontoid process (located at the center of the discoid facet; Fig. 4b–h). At the plane passing throw the center of the discoid and odontoid facets, the angle between both facets is nearly 115° in M. patagonicus; it is 110° in ‘Eo.’ cabrerai, and it is nearly straight in Pl. lelongi, Py. bergi, and Py. scillatoyanei. In M. patagonicus a large sesamoid facet forms the dorsomedial end of a relatively wide groove adjacent to the posterior margin of the odontoid facet (Fig. 4b–h); this facet is absent in other megatheriines (e.g., ‘Eo.’ cabrerai, Py. bergi).
In M. patagonicus, near two-thirds of the navicular facet projects dorsal to the discoid facet when the long axis of the odontoid process is vertically oriented (Fig. 4c,f,i). In ‘Eo.’ cabrerai, Eo. nanum, Pl. lelongi, Py. bergi, and Py. scillatoyanei, the navicular facet is more ventral, with the plane of the discoid facet intersecting approximately at the center of the circular depression of the navicular facet.
There are two facets for articulation with the calcaneum: the sustentacular and the ectal facets. As in most megatheriines, these two facets are separated by a deep and wide non-articular bony canal called the sulcus tali. The sustentacular facet is continuous with the cuboid facet but is separated from the ventral portion of the navicular facet; it is almost planar or flat and subtriangular in shape. The ectal facet is oval in shape, with the main axis anterolateral to posteromedial, and parallel to the sulcus tali; it is concave along the main axis, and convex along the other axis.
Discussion
After the description of M. patagonicus by Ameghino (1904), two other species belonging to Megathericulus were described for the Miocene of Patagonia, M. friasensis Kraglievich, 1930, which probably corresponds to a Scelidotheriinae (see Kraglievich 1930; Brandoni 2006a; De Iuliis et al. 2008), and M. primaevus based on a few postcranial bones (e.g., astragalus, portion of tibia). De Iuliis et al. (2008) added a diagnosis for M. patagonicus, described two new specimens from the Miocene of Patagonia, and increased the knowledge on anatomy and intraspecific variation of the species. Pujos et al. (2013) referred to Megathericulus sp. a specimen from the middle Miocene of Peru, transferred the megatheriines Eo. andinum and Eo. cabrerai to Megathericulus (as M. andinum and M. cabrerai), and considered M. primaevus a valid species, although they indicated that synomymy between M. patagonicus and M. primaevus cannot be discarded. Finally, Brandoni et al. (2018) described a new specimen of M. patagonicus from the middle Miocene of Bolivia, and considered that it is not possible to strongly establish whether Eo. cabrerai and Eo. andinum should be referred to Eomegatherium or Megathericulus, refering these species instead as ‘Eomegatherium’ andinum and ‘Eomegatherium’ cabrerai.
Referral of the new specimens described here (LIEB-PV 6153 and LIEB-PV 6152) to M. patagonicus is based on metric and morphological similarities with the holotype (MACN A 11151) and specimens that have been referred to this species (MLP 91-IX-7-18, MLP 92-XI-15-2; see De Iuliis et al. 2008). Shared features include: 1) overall size, smaller than other Neogene Megatheriinae such as Eomegatherium, Pliomegatherium, and Pyramiodontherium (Tables 1, 3); 2) shape of the molariforms, molariforms mesiodistally compressed and rectangular in outline; 3) Hypsodonty Index close to 100; 4) position of the navicular facet, two-thirds of the navicular facet projects dorsal to the discoid facet; 5); in the astragalus, shape of the ventral portion of the fibular facet, the ventral portion is subtriangular outline and contacts the ectal facet; 6) presence of a sesamoid facet in the astragalus.
Regarding the validity of M. primaevus, the astragalus (MLP 39-VI-24-1) is the only well-preserved bone of the species that can be compared with M. patagonicus (MACN A 11151, MLP 91-IX-7-18, MLP 92-XI-15-2). The astragalus of both species is similar in: 1) overall size; 2) the shape of the ventral portion of the fibular facet; 3) position of the navicular facet respect de plane of the discoid facet, and 4) the presence of a sesamoid facet. Cabrera (1939) considered that the general shape of MLP 39-VI-24-1 resembles that of the astragalus of M. patagonicus (MACN A 11151), but it differs in size, development of the odontoid process, and width of sulcus tali. With the inclusion of MLP 91-IX-7-18 and MLP 92-XI-15-2 in M. patagonicus (see De Iuliis et al. 2008), the intraspecific variation of M. patagonicus increased. Intraspecific variation, including sexual dimorphism, of ground sloths were discussed in recent contributions that dealt with systematics of Megatheriinae (e.g., Faure et al. 2014; Cartelle et al. 2014; Carlini et al. 2018), Megalonychidae (e.g., Brandoni 2010, 2011), and Nothrotheriidae (e.g., Amson et al. 2015). Considering the large intraspecific differences exhibited in some ground sloth species (see Cartelle et al. 2014, for the case of Eremotherium laurillardi; Brandoni 2010, 2011, for Neogene megalonychids; Amson et al. 2015, for Thalassocnus Muizon and McDonald, 1995), the similarities and few differences between M. primaevus and M. patagonicus allow us to consider M. primaevus as a junior synonym of M. patagonicus. In sum, M. patagonicus is the only diagnosed and well-characterized species of the genus.
The most peculiar feature of the dentition of M. patagonicus is the shape of the molariforms, which was described by Brandoni et al. (2018) for some molariforms of the upper series. De Iuliis et al. (2008) and Pujos et al. (2013) have already described the shape of the alveoli of the dentary; however, the descriptions were based on edentulous dentaries, inferring the molariform outlines from the shape of the preserved alveoli. The description herein presented confirms the aspects that were described or inferred by De Iuliis et al. (2008), Pujos et al. (2013), and Brandoni et al. (2018) (e.g., rectangular shape of the molariforms), but also adds information on the precise shape of the lower molariforms (presence of two lophids separated by a deep V-shaped valley, presence of lingual and labial longitudinal grooves).
During the course of evolution, the body size of megatherines and, as a result, the size of the molariforms generally increased (Brandoni et al. 2012). The size increase did not occur evenly in the molariforms, with the anteroposterior length of these teeth generally increasing more than their labiolingual width (Kraglievich 1930; Brandoni et al. 2012; Pujos et al. 2013). Thus, while the molariforms of megatheriines from the Serravallian-early Tortonian (e.g., Megathericulus, Anisodontherium) are usually rectangular in outline, those from the late Tortonian-Quaternary megatheriines are subtrapezoidal to squared (i.e., more isodiametric) (Fig. 5). This increase in the anteroposterior length of each molariform resulted in an increase in the relative length of the molariform tooth row, which, in turn, resulted in a relatively longer tooth row in Quaternary megatheriines (e.g., Megatherium, Eremotherium) than taxa from the Serravallian-early Tortonian (e.g., Megathericulus, Anisodontherium).
Fig. 5
Cladogram showing relationships among Megatheriinae (modified from Varela et al. 2019). Chronological distribution of mentioned genera and evolution of the morphology of lower molariform and astragalus is shown
Zetti (1964) made the first attempt to quantify the depth of the dentary in megatheriines and developed the Hypsodonty Index, where a higher HI reflects a great depth of the dentary (=increased hypsodonty). The HI has been used in Megatheriinae systematics, phylogenetic analysis, and evolutionary aspects of the group (see Saint-André and De Iuliis 2001; Pujos 2006; Brandoni and De Iuliis 2007; Brandoni et al. 2008), as well as paleobiological studies of ground sloths (see Bargo et al. 2006; Fields 2009). In particular, it is one of the characters proposed to separate Megatherium from Eremotherium (see Saint-André and De Iuliis 2001), as well as Megatherium americanum from Megatherium gallardoi (see Brandoni et al. 2008). In addition, the differences between the HI of Megatherium americanum and Er. laurillardi have been related to paleoenvironmental aspects, with Megatherium americanum (high HI) linked to open grassland, and Er. laurillardi (low HI) to closed forest habitats (Bargo et al. 2006).
As was mentioned, the HI is calculated as greatest depth of dentary / length of the molariform tooth row × 100; the HI values for Megathericulus (HI = 102) fit in the variation of Megatherium americanum (HI = 92–112); however, the relatively shorter molariform tooth row in Megathericulus exaggerates its hypsodonty. In other words, in the case of Megatherium a higher HI reflects a great depth of the dentary, whereas in M. patagonicus HI does not necessarily reflect a great depth of the dentary given that in this case the low value of the molariform tooth row length collaborates to obtain a high HI. Thus, and although the HI is commonly used for different purposes in the study of Megatheriinae, its use is problematic when taxa with different outline of molariforms are compared. In addition, this high value of HI of M. patagonicus may also reflect Patagonian paleoenvironmental conditions during the late middle Miocene (see Ortiz-Jaureguizar and Cladera 2006).
Other features, considered in the evolution of Megatheriinae, closely related to the length of the lower molariform tooth row are the position of the anterior margin of the coronoid process with respect to the m4, and the position of the posterior margin of the mandibular symphysis with respect to the m1 (see Brandoni et al. 2012). As in Anisodontherium, in M. patagonicus the molariform tooth row lies between the posterior margin of the mandibular symphysis and the anterior margin of the coronoid process, whereas the molariform tooth row extends beyond those mandibular features in Quaternary Megatheriinae, owing to a relatively longer tooth row. However, as with the case of HI, given that these characters seem closely related to the length of the molariform tooth row, and thus the shape of the molariforms, it seems likely that the latter might have an influence on the way these features are scored and interpreted in cladistic analyses (Brandoni et al. 2012).
Regarding the astragalus, in M. patagonicus, near two-thirds of the navicular facet projects dorsal to the discoid facet, whereas in other Neogene megatheriines (e.g., Pyramiodontherium) the navicular facet is more ventral, with the plane of the discoid facet intersecting approximately at the center of the circular depression of the navicular facet (Fig. 5). In Quaternary megatheriines (i.e., Eremotherium, Megatherium), the navicular facet lies farther ventrally, with the plane of the discoid facet intersecting the dorsal part of the navicular facet. In addition, in M. patagonicus the ventral portion of the fibular facet contacts the ectal facet, whereas in the rest of the Megatheriinae the ectal facet does not contact.
Megathericulus patagonicus shows characters that are plesiomorphic for Megatheriinae (e.g., molariforms that are mesiodistally compressed and rectangular in outline, very long rostrum, navicular facet of the astragalus dorsally located with respect to the discoid facet), suggesting it is the earliest diverging taxon with respect to other members of the subfamily (Brandoni 2006a; Pujos 2006; De Iuliis et al. 2008; Brandoni et al. 2018; Varela et al. 2019). The condition of earliest diverging genus of Megatheriinae coincides with the fact that M. patagonicus is probably the geologically oldest species for the subfamily (Figs. 5,6a).
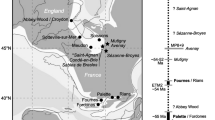
Fig. 6
Chronological and geographical distribution of taxa mentioned in Discussion.a Chronological distribution of M. patagonicus, Anisodontherium (Megatheriinae), ‘Xyophorus’ (Nothrotheriinae), and Planops, Prepotherium, Prepoplanops (Planopinae). b geographical distribution of M. patagonicus, Anisodontherium (Megatheriinae), ‘Xyophorus’ (Nothrotheriinae) (black circles), and Planopinae (grey circles); Megatheriinae and Nothrotheriinae may have originated and/or had their center of dispersion in low or middle latitudes of South America and later dispersed into higher and lower latitudes (grey arrows)
The precise geographic and stratigraphic provenance of the holotype MACN A 11151 on which M. patagonicus is based is unknown. According to Ameghino (1904: 282), it is ‘Tehuelche antiguo del Chubut (Laguna Blanca)’ but the exact geographic location where C. Ameghino collected the fossils is unknown; Ameghino (1906) mentioned that all the fossils come from the lower and middle section of the outcrops of bluish grey sandstone from the region of Fénix River and Laguna Blanca. According to the geologic charts of the region the only continental formation with bluish grey sandstones that crops out close to the Fénix River, Lago Blanco town and Lago Blanco (=Laguna Blanca) lagoon belongs to Río Mayo Formation (Dal Molín 1998; Escosteguy et al. 2003). However, below the Río Mayo Formation in the area there are also two continental formations characterized by grey sandstontes (El Portezuelo and Pedregoso formations) with records of fossil mammals (Scillato-Yané and Carlini 1998) where C. Ameghino could also collected the holotype of M. patagonicus.
Accurate geographic (Fig. 6b) and stratigraphic data are available for several specimens referred to M. patagonicus (i.e., MLP 91-IX-7-18, MLP 92-XI-15-2, UATF-V-001414, UATF-V-001163 and LIEB-PV 6153, LIEB-PV 6152) and Megathericulus sp. (MUSM 1564). MLP 91-IX-7–18 was recovered from Río Mayo Formation at Cerro Guenguel locality, where a level of this formation below the fossil horizon has been dated preliminarily at ca.11.8 Ma (40Ar/39Ar; Madden et al. 1997; De Iuliis et al. 2008). MLP 92-XI-15-2 was recovered also from the Río Mayo Formation but at Arroyo Pedregoso locality. At Cerro Guenguel and Arroyo Pedregoso, the Río Mayo Formation overlies the El Portezuelo Formation and Pedregoso Formation, respectively, which could be chronologically equivalents (Escosteguy et al. 2003; Rivas et al. 2015). The Pedregoso Formation at Arroyo Pedregoso has been dated at ca. 12.18 Ma (40Ar/39Ar; Dal Molín and Franchi 1996). UATF-V-001414 was recovered from the Quebrada Honda beds (Bolivia) and is closely associated with a tuff dated at 12.5–12.2 Ma, and UATF-V-001163 from the Huayllajara Local Fauna may date to 13.2–12.8 Ma (see Brandoni et al. 2018).
The two specimens presented herein (LIEB-PV 6153, LIEB-PV 6152) come from the southern outcrops of the Collón Curá Formation (i.e., Cerro Zeballos; La Gloria), where a guide level for the Collón Curá Formation (i.e., Pilcaniyeu Ignimbrite Member) is not deposited (Mazzoni 1993), and in part because of this, correlations with the fossil levels associated with the Ignimbrite, where the Colloncuran fauna sensu stricto was defined (see Vucetich et al. 1993) could not be identified. In this area where southern croppings of Collón Curá Formation at Chico River (south of Río Negro Province) are exposed, the absolute 40Ar/39Ar ages obtained spans from 14.8 Ma to 13.7 Ma (Bilmes et al. 2013; Dunn et al. 2015), while at Cushamen River (northwest Chubut Province), near Cerro Zeballos and La Gloria, there is one U-Pb absolute age of ca. 11.2 Ma (Ramos et al. 2015). According to that, and because there are no absolute dates for Cerro Zeballos, the chronological location of this locality in Fig. 6a is relative and based on the study of Brandoni et al. (2017) that indicated an early Tortonian age (its position could change if absolute dates appear). The chronological location of La Gloria in Fig. 6a is based on Bucher (2018).
The specimen MUSM 1564 (Megathericulus sp.) was recovered from the SEP-007 locality in southeastern Peru (Pujos et al. 2013) and corresponds to the Fitzcarrald local fauna, which has been referred to the Laventan South American Land-Mammal Age (SALMA), considered as Serravallian in age (13.5–11.8 Ma) (see Madden et al. 1997). Finally, we could not confirm two records of Megathericulus sp. for Collón Curá Formation, one for Neneo Rucá (Río Negro Province; Casamiquela 1969; Rabassa 1975; Pascual et al. 1984) and one for Collón Curá River (Neuquén Province; Bondesio et al. 1980), because they were published without collection numbers, descriptions, or figures of the specimens, and we found no specimens that match with this provenance at the MLP or MACN, where they were probably deposited.
The type specimen of M. primaevus (MLP 39-VI-24-1) was collected by J. Frenguelli 5 km from Paso Flores (Neuquén Province) at cañadón Ftamichi from Miocene tuffs that Cabrera (1939) considered Santacrucian in age. According to Frenguelli (1939) cañadón Ftamichi ends at the right margin of Limay River in front of the Paso Flores balse (Río Negro Province). Pascual et al. (1978) and Scillato-Yané and Carlini (1998) assigned this locality to Collón Curá Formation and to the Colloncuran age, respectively, without any details. In the geologic charts (Nullo 1979; Escosteguy et al. 2013) there is a ‘cañadón’ although without name, that ends in the right margin of Limay River in front of Paso Flores balse. This ‘cañadón’ cerro Flamiche runs just in between extensive Neogene tuffaceous sediments (ca. 5 km from Paso Flores) mostly assigned to Collón Curá Formation, from where the specimen MLP 39-VI-24-I could come (ca. 40°37′57.34’S, 70°38′22.20’W).
The other Megatheriinae species recorded from the middle Miocene are ‘Eo.’ cabrerai (with quotation marks following Brandoni et al. 2018), which was recorded from Laguna Blanca and Fenix River, and ‘Eo.’ andinum from Huemúles River (see Kraglievich 1930). However, as the case of the old records of Megathericulus from these localities, there is not much precise geochronological information (see González Ruiz et al. 2017). Brandoni and Carlini (2004) mentioned Megatheriinae remains from the Cerro Guenguel locality (ca. 11.8 Ma, Río Mayo Formation) that the authors considered probably correspond to Eomegatherium. In addition, an undetermined Megatheriinae, based on a posterior fragment of mandible, was recorded at the locality of Cerdas, Bolivia (ca. 16.3–15.1 Ma, early Miocene; Croft et al. 2016). Taking into account the absolute ages considered to the levels bearing specimens of M. patagonicus and Eomegatherium, M. patagonicus is the oldest nominated Megatheriinae ranging from nearly 13.2 Ma (Huayllajara Local Fauna, Bolivia) to nearly 11.2 Ma (at Cushamen River, near Cerro Zeballos and La Gloria, Chubut Province, Argentina) (Fig. 6a).
Regarding the phylogenetic relationships of Megatheriinae with other clades of Tardigrada, the subfamily Megatheriinae has been considered to be the sister group of Planopinae (e.g., Planops Ameghino, 1891, Prepotherium Ameghino, 1891; see Ameghino 1891a, 1891b), with both included in Megatheriidae (De Iuliis 1994; Gaudin 2004; Amson et al. 2016; Varela et al. 2019). However, Pujos et al. (2007) indicated that the relationships of Planops martini Hoffstetter, 1961, to other Tardigrada are uncertain, and that Megatheriidae are related to Nothrotheriidae; and Carlini and Scillato-Yané (2004) indicated that Megatheriinae is the sister group of Nothrotheriinae.
Most of the accurate records of Planopinae (Prepotherium, Planops, Prepoplanops Carlini et al., 2013) correspond to those from the early Miocene of Santa Cruz Province (Santa Cruz and Cerro Boleadoras formations) (see Scott 1903–04; Carlini et al. 2013) where no remains of Megatheriinae or Nothrotheriinae were collected (Fig. 6). Remains of Prepotherium and Planops were recorded from the Santa Cruz Formation, which, at the Atlantic coast of Santa Cruz Province, spans the interval of 18–16 Ma (Perkins et al. 2012). Prepoplanops was recorded from Cerro Boleadoras Formation (western Santa Cruz Province; see Carlini et al. 2013), which was considered as early Miocene in age (Charrier et al. 1979; Vucetich 1994).
Regarding the relationship between Megatheriinae and Nothrotheriinae, it is interesting to note that an undetermined Megatheriinae (see above) and the Nothrotheriinae ‘Xyophorus’ cf. ‘X’ bondesioi Scillato-Yané 1979 (see Brandoni 2014, for a discussion on the differences between ‘Xyophorus’ and Xyophorus Ameghino, 1887) at the locality of Cerdas (Bolivia) represent the oldest records of the subfamilies Megatheriinae and Nothrotheriinae (see Croft et al. 2009, 2016; Brandoni 2014). This may suggest that both groups may have originated and/or had their center of dispersion in low or middle latitudes of South America and later dispersed into higher and lower latitudes (Pujos et al. 2013; Brandoni et al. 2018) (Fig. 6b). In this sense, Megatheriinae, represented by Megathericulus, and Nothrotheriinae, by ‘Xyophorus’, were recorded together from Quebrada Honda (Bolivia) and Cerro Zeballos (Chubut Province) (Croft et al. 2016; Brandoni et al. 2017). These records, the earlier and not overlapping record of Planopinae, and the cladistics results reached by Carlini and Scillato-Yané (2004) and Pujos et al. (2007), would suggest a close relationship between Megatheriinae and Nothrotheriinae. In addition, the presence of Megathericulus and ‘Xyophorus’ in some distant paleofauna associations (ca. 2300 km) would suggest a similar path of dispersion and paleoenvironment requirements of both taxa. The presence of these two genera in Cerro Zeballos (Collón Curá Formation, Argentina) and Quebrada Honda (Honda Group, Bolivia) supports what was indicated by Croft (2007) that the Quebrada Honda Fauna as a whole is more similar to the asynchronous high-latitude faunas from Patagonia than to contemporary La Venta Fauna (Colombia).
Finally, the record of M. patagonicus (Megatheriinae) adds a new taxa to the Patagonian Cerro Zeballos fauna, that, although some groups are still under study (e.g., caviomorph rodents, dasypodid xenarthrans, hegetotherids, toxodontids, astrapotherids, and litopterns ungulates), is composed of tortoises [Chelonoidis gringorum (Simpson, 1942)], marsupials (Pseudonotictis chubutensis Martin and Tejedor, 2007, Palaeothentidae sp. nov. A and B, Argyrolagydae sp. nov., Atlyiacinidae sp. nov.; Martin and Tejedor 2007; Martin et al. 2016), xenarthrans (Paraeucinepeltus raposeirasi González Ruiz et al., 2011, ‘Xyophorus’, M. patagonicus; González Ruiz et al. 2011; Brandoni et al. 2017; this paper), and ungulates (Caenophilus tripartitus Ameghino, 1903, Protypotherium colloncurensis Vera et al., 2018, Protypotherium cf. Pr. endiadys (Roth, 1899); Vera et al. 2018), and that represents one of the southern and probably younger faunas of the Collón Curá Formation.
References
Ameghino C, Kraglievich L (1921) Descripción del “Megatherium gallardoi” C. Amegh. descubierto en el Pampeano inferior de la ciudad de Buenos Aires. Anales del Museo Nacional de Historia Natural de Buenos Aires 31: 134–156.
Ameghino F (1885) Nuevos restos de mamíferos fósiles oligocenos, recogidos por el professor Pedro Scalabrini y pertenecientes al Museo Provincial de la Ciudad del Paraná. Boletín de la Academia Nacional de Ciencias de Córdoba 8: 5–207
Ameghino F (1887) Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austral y depositados en el Museo de La Plata. Boletín del Museo de La Plata 1: 1–26
Ameghino F (1891a) Nuevos restos de mamíferos fósiles descubiertos por Carlos Ameghino en el Eoceno inferior de la Patagonia austral. Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1: 289–328
Ameghino F (1891b) Caracteres diagnósticos de cincuenta especies nuevas de mamíferos fósiles argentinos. Revista Argentina de Historia Natural 1: 129–167
Ameghino F (1903) Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 56: 193–208
Ameghino F (1904) Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 58: 225–291
Ameghino F (1906) Les Formations Sedimentaires du Crétacé Supérieur et du Tertiaire de Patagonie avec un paralléle entre leurs faunes mammalogiques et celles de l'ancien continent. Anales del Museo Nacional de Buenos Aires 15 (ser. 3a, 8): 1–568
Amson E, Argot C, McDonald HG, Muizon C de (2015) Osteology and functional morphology of the hind limb of the marine sloth Thalassocnus (Mammalia, Tardigrada). J Mammal Evol 22: 355–419
Amson E, Muizon C de, Gaudin TJ (2016) A reappraisal of the phylogeny of the Megatheria (Mammalia: Tardigrada), with an emphasis on the relationships of the Thalassocninae, the marine sloths. Zool J Linn Soc. doi: https://doi.org/10.1111/zoj.12450
Bargo MS, De Iuliis G, Vizcaíno SF (2006) Hypsodonty in Pleistocene ground sloths. Acta Palaeontol Pol 51: 53–61
Bilmes A, DʼElia L, Franzese JR, Veiga GD Hernández M (2013) Miocene block uplift and basin formation in the Patagonian foreland: the Gastre Basin, Argentina. Tectonophysics 601: 98–111
Bilmes A, D’Elia, L, Veiga GD, Franzese JR (2014) Relleno intermontano en el antepaís fragmentado patagónico: evolución neógena de la cuenca de gastre. Revista de la Asociación Geológica Argentina 71: 311–330
Bondesio P, Rabassa J, Pascual R, Vucetich MG, Scillato-Yané GJ (1980) La Formación Collón-Curá de Pilcaniyeu Viejo y sus alrededores (Río Negro, República Argentina). Su antigüedad y las condiciones ambientales según su distribución, su litogénesis y sus vertebrados. Actas II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología 3: 85–99. Buenos Aires, 1978
Bostelmann E, Bobe R, Carrasco G, Alloway B, Santi-Malnis P, Mancus A, Agüero B, Alemseged Z, Godoy Y (2012) The Alto Río Cisnes Fossil Fauna (Río Frías Formation, early middle Miocene, Friasian SALMA): a keystone and paradigmatic vertebrate assemblage of the South American fossil record. III Simposio de Paleontología en Chile, Punta Arenas: 46–49
Brandoni D (2006a) Los Megatheriinae (Xenarthra, Tardigrada, Megatheriidae) Terciarios de la Argentina. Sistemática, Evolución y Biogeografía. Unpublished Doctoral Thesis, Facultad de Ciencias naturales y Museo, Universidad Nacional de La Plata, 392 pp
Brandoni D (2006b) A review of Pliomegatherium Kraglievich, 1930 (Xenarthra: Phyllophaga: Megatheriidae). Neues Jahrb Geol Paläontol – Mon 2006: 212–224
Brandoni D (2010) On the systematics of Ortotherium Ameghino (Xenarthra, Tardigrada, Megalonychidae) from the “Conglomerado osífero” (late Miocene) of Argentina. J Vertebr Paleontol 30: 975–980
Brandoni D (2011) The Megalonychidae (Xenarthra, Tardigrada) from the late Miocene of Entre Ríos Province, Argentina, with remarks on their systematics and biogeography. Geobios 44: 33–44
Brandoni D (2013) Los Tardigrada (Mammalia, Xenarthra) del Mioceno Tardío de Entre Ríos, Argentina. In: Brandoni D, Noriega JI (eds) El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14: 135–144
Brandoni D (2014) “Xyophorus” sp. en el Mioceno Medio de Chubut: implicancias sistemáticas, biogeográficas y biocronológicas del registro de un Nothrotheriinae en el Neógeno de la Argentina. Ameghiniana 51: 94–105
Brandoni D, Carlini AA (2004) Nuevos restos de Megatheriinae (Xenarthra, Phyllophaga, Megatheriidae) en el Mayoense (Mioceno Medio) de Santa Cruz. XX Jornadas Argentinas de Paleontología de Vertebrados. La Plata, 2004. Ameghiniana 41(4):Suplemento Resúmenes:37R
Brandoni D, Carlini AA, Anaya F, Gans P, Croft DA (2018) New remains of Megathericulus patagonicus Ameghino, 1904 (Xenarthra, Tardigrada) from the Serravallian (middle Miocene) of Bolivia; chronological and biogeographical implications. J Mammal Evol 25: 327–337. doi: https://doi.org/10.1007/s10914-017-9384-y
Brandoni D, Carlini AA, Pujos F, Scillato-Yané GJ (2004) The pes of Pyramiodontherium bergi (Moreno & Mercerat, 1891) (Mammalia, Xenarthra, Phyllophaga): the most complete pes of a Tertiary Megatheriinae. Geodiversitas. 26: 643–659
Brandoni D, De Iuliis G (2007) A new genus for the Megatheriinae (Xenarthra, Phyllophaga, Megatheriidae) from the Arroyo Chasicó Formation (upper Miocene) of Buenos Aires Province, Argentina. Neues Jahrb Geol Paläontol Abh 244: 53–64
Brandoni D, González Ruiz L, Reato A, Martin G (2017) Chronological implications of the nothrotheriid ‘Xyophorus’ (Mammalia, Xenarthra) from the Collón Curá Formation (Miocene of Patagonia, Argentina). Hist Biol. doi: https://doi.org/10.1080/08912963.2017.1398748
Brandoni D, Powell JE, González OE (2012) Anisodotherium from the late Miocene of north-western Argentina. Acta Palaeontol Pol 57: 241–249
Brandoni D, Soibelzon E, Scarano A (2008) On Megatherium gallardoi (Xenarthra, Megatheriidae) and the Megatheriinae from the Ensenadan (lower to middle Pleistocene) of the Pampean region, Argentina. Geodiversitas 30: 793–804
Brunetto E, Noriega JI, Brandoni D (2013) Sedimentología, estratigrafía y edad de la Formación Ituzaingó en la provincia de Entre Ríos, Argentina. In: Brandoni D, Noriega JI (eds) El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14: 13–27
Bucher J (2018) Tectónica, sedimentación y volcanismo del antepaís roto durante la inversión andina en la zona central de Chubut. Unpublished Doctoral Thesis, Facultad de Ciencias naturales y Museo, Universidad Nacional de La Plata, 191 pp
Bucher J, López M, García M, Bilmes A, D’Elia, L, Funes D, Feo R, Franzese J (2018, In Press) Estructura y estratigrafía de un bajo neógeno del antepaís norpatagónico: el depocentro Paso del Sapo, provincia de Chubut. Revista de la Asociación Geológica Argentina 75
Cabrera A (1928) Sobre algunos megaterios Pliocenos. Revista del Museo de La Plata 31: 339–352
Cabrera A (1939) Un nuevo representante Santacrucense de la Familia Megatheriidae. Notas del Museo de La Plata. Paleontología 4: 475–483
Carlini AA, Brandoni D, Dal Molín CN (2013) A new genus and species of Planopinae (Xenarthra: Tardigrada) from the Miocene of Santa Cruz Province, Argentina. Zootaxa 3694: 565–578
Carlini AA, Brandoni D, Sánchez R, Sánchez-Villagra MR (2018) A new Megatheriinae skull (Xenarthra, Tardigrada) from the Pliocene of northern Venezuela – implications for a giant sloth dispersal to Central and North America. Palaeontol Electron 21.2.16A 1-12. https://doi.org/10.26879/771
Carlini AA, Brandoni D, Scillato-Yané GJ, Pujos F (2002) Una nueva especie de megaterino (Xenarthra, Megatheriidae) del Mioceno Tardío-Plioceno de Catamarca, Argentina. Ameghiniana 39: 367–377
Carlini AA, Scillato-Yané GJ (2004) The oldest Megalonychidae (Xenarthra: Tardigrada); phylogenetic relationships and an emended diagnosis of the family. Neues Jahrb Geol Paläontol Abh 233:423–443
Cartelle C, De Iuliis G, Pujos F (2014) Eremotherium laurillardi (Lund, 1842) (Xenarthra, Megatheriinae) is the only valid megatheriine sloth species in the Pleistocene of intertropical Brazil: a response to Faure et al., 2014. C R Palevol 14: 15–23
Casamiquela RM (1969) Historia geológica del valle de Huahuel Niyeo área extraandina del suroeste de la provincia de Río Negro, República Argentina (con énfasis en el Pleistoceno). Revista de la Asociación Geológica Argentina 24: 287–329
Cazau LB (1980) Cuenca de Ñirihuau-Ñorquinco-Cushamen. II Simposio de Geología Regional Argentina 2: 1149–1172
Charrier R, Linares E, Niemeyer H, Skarmeta J (1979) Edades Potasio-Argón de vulcanitas mesozoicas y cenozoicas del sector chileno de la meseta Buenos Aires, Aysen, Chile, y su significado geológico. VII Congreso Geológico Argentino 2: 23–41
Cohen KM, Finney SC, Gibbard PL, Fan J-X (2013) Updated. The ICS international chronostratigraphic chart. Episodes 36: 199–204
Cope ED (1889) The Edentata of North America. Am Nat 23: 657–664
Croft DA (2007) The middle Miocene (Laventan) Quebrada Honda Fauna, southern Bolivia, and a description of its notoungulates. Palaeontology 50: 277–303
Croft DA, Anaya F, Auerbach D, Garzione C, MacFadden BJ (2009) New data on Miocene Neotropical provinciality from Cerdas, Bolivia. J Mammal Evol 16: 175–198
Croft DA, Carlini AA, Ciancio MR, Brandoni D, Drew NE, Engelman RK, Anaya F (2016) New mammal faunal data from Cerdas, Bolivia, a mid-latitude Neotropical site that chronicles the end of the middle Miocene climatic optimum in South America. J Vertebr Paleontol 36: e1163574
Cuvier G (1796) Notice sur le squelette d’une très grande espèce de quadrupède inconnue jusqu’à présent trouvé au Paraguay et déposé au cabinet d’histoire naturelle de Madrid. Magasin Encyclopédique: ou Journal des Sciences, des Lettres et des Arts 1: 303–310
Dal Molín C (1998) Hoja geológica 4572-IV, Alto Río Senguerr, escala 1:250.000, provincial del Chubut. Servicio Geológico Minero Argentino Boletín N° 255: 1–55. (Mapa y texto preliminar para consulta)
Dal Molín CN, Franchi M (1996) Reinterpretación estratigráfica de las sedimentitas terciarias del suroeste de Chubut. XIII Congreso Geológico Argentino y III Congreso de Exploración de Hidrocarburos, Actas 1: 473–478
De Iuliis G (1994) Relationships of the Megatheriinae, Nothrotheriinae, and Planopsinae: some skeletal characteristics and their importance for phylogeny. J Vertebr Paleontol 14: 577–591
De Iuliis G (1996) A systematic review of the Megatheriinae (Mammalia: Xenarthra: Megatheriidae). Unpublished Doctoral Thesis, University of Toronto, Toronto, 781 pp
De Iuliis G, Brandoni D, Scillato-Yané GJ (2008) New remains of Megathericulus patagonicus Ameghino, 1904 (Xenarthra, Megatheriidae): information on primitive features of megatheriines. J Vertebr Paleontol 28: 181–196
De Iuliis G, Pujos F, Tito G (2009) Systematic and taxonomic revision of the Pleistocene ground sloth Megatherium (Pseudomegatherium) tarijense (Xenarthra: Megatheriidae). J Vertebr Paleontol 29: 1244–1251
De Iuliis G, Ré GH, Vizcaíno SF (2004) The Toro Negro megatheriine (Mammalia: Xenarthra): a new species of Pyramiodontherium and a review of Plesiomegatherium. J Vertebr Paleontol 24: 214–227
de la Cruz R, Cortés J (2011) Geología del área oriental de la Hoja Puerto Cisnes, Región Aysén del Gral. Carlos Ibáñez del Campo. Serie Geología Básica 127: 1–70
Dunn RE, Strömberg CAE, Madden RH, Kohn MJ, Carlini AA (2015) Linked canopy, climate, and faunal change in the Cenozoic of Patagonia. Science 347: 258–261
Encinas A, Folguera A, Litvak VD, Echaurren A, Gianni G, Fernández Paz F, Bobe R, Valencia V (2016) New age constraints for the Cenozoic deposits of the Patagonian Andes and the Sierra de San Bernardo between 43° and 46°S. I Simposio de tectónica Sudamericana: 140
Escosteguy L, Dal Molín C, Franchi M, Geuna S, Lapido O (2003) Hoja Geológica 4772-II, Lago Buenos Aires, escala 1:250.000, provincia de Santa Cruz. Servicio Geológico Minero Argentino Boletín N° 339: 1–72
Escosteguy L, Geuna S, Franchi M, González Díaz E, Dal Molín C (2013) Hoja Geológica 4172-II, San Martín de los Andes, escala 1:250.000, provincias del Neuquén y de Río Negro. Servicio Geológico Minero Argentino Boletín N° 409: 1–92
Faure M, Guérin C, Parenti F (2014) Sur l’existence de deux espèces d’ Eremotherium, E. rusconii (Schaub, 1935) et E. laurillardi (Lund, 1842), dans le Pléistocène supérieur du Brésil intertropical. C R Palevol 13: 259–266
Fields EF (2009) Hypsodonty in the Pleistocene ground sloth Megalonyx: closing the “diastema” of data. Acta Palaeontol Pol 54: 155–158
Flynn JJ, Swisher CC III (1995) Cenozoic South American land mammal ages: correlation to global geochronologies. In: Berggren WA, Kent DV, Aubry M-P, Hardenbol J (eds) Geochronology, Time Scales, and Global Stratigraphic Correlation. SEPM (Society for Sedimentary Geology) Spec Publ 54: 317–333
Folguera A, Encinas A, Echaurren A, Gianni G, Orts D, Valencia V, Carrasco G (2018) Constraints on the Neogene growth of the central Patagonian Andes at the latitude of the Chile triple junction (45-47 °S) using U/Pb geochronology in synorogenic strata. Tectonophysics 744: 134–154
Frenguelli J (1939) Nidos fósiles de insectos en el terciario del Neuquén y Río Negro. Notas del Museo de La Plata. Paleontología 4: 379–402
Gaudin TJ (2004) Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zool J Linn Soc 140: 255–305
Gervais H, Ameghino F (1880). Los mamíferos fósiles de la América del Sud. F. Savy-Ignon Hermanos, Paris-Buenos Aires. 225 pp
González Ruiz LR, Reato A, Cano M, Martínez O (2017) Old and new specimens of a poorly known glyptodont from the Miocene of Patagonia and their biochronological implications. Acta Palaeontol Pol 62: 181–194
González Ruiz LR, Zurita AE, Scillato-Yané GJ, Zamorano M, Tejedor MF (2011) Un nuevo Glyptodontidae (Mammalia, Xenarthra, Cingulata) del Mioceno de Patagonia (Argentina) y comentarios acerca de la sistemática de los gliptodontes del SALMA ‘Friasense’. Revista Mexicana de Ciencias Geológicas 28: 566–579
Gray JE (1821) On the natural arrangement of vertebrose animals. London Medical Repository 5: 296–310
Hoffstetter R (1961) Description d'un squelette de Planops (Gravigrade du Miocène de Patagonie). Mammalia 25: 57–96
International Commission on Stratigraphy (2018) International Chronostratigraphic Chart. www.stratigraphy.org
Kraglievich L (1926) Notas sobre gravígrados de Sud América. Anales del Museo Nacional de Historia Natural de Buenos Aires 34: 21–36
Kraglievich L (1930) La formación friaseana del Río Frías, Río Fénix, Laguna Blanca, etc. y su fauna de mamíferos. Physis 10: 127–161
Kramarz A G, Forasiepi AM, Bond M (2011) Vertebrados Cenozoicos. In: Leanza HA, Arregui C, Carbone O, Danieli JC, Vallés JM (eds) Relatorio del XVIII Congreso Geológico Argentino, Abstracts: 557–572
Latham J, Davies H (1795) Faunula Indica, 38 Pp. Appendix to Forster JR. 2nd edition, Gebauer JJ, Halle a.S.
Linnaeus C (1758) Systema Naturae, Ed. 10°. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, locis. Vol 1: Regnum animale. 10 i-ii + 1–824
Lund PW (1842) Blik paa Brasiliens Dyreverden för Sidste Jordomvaeltning. Fjerde Afhandling: Fortsaettelse af Pattedyrene. Detkongelige Danske Videnscavernes Selskabs Skrifter; Naturvidenskabelige og mathematisk Afhandlinger 9: 137–208
Madden RH, Guerrero J, Kay RF, Flynn JJ, Swisher CC III, Walton AH (1997) The Laventan stage and age. In: Kay RF, Madden RH, Cifelli RL, Flynn JJ (eds) Vertebrate Paleontology in the Neotropics: The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, D.C., pp 499–519
Marshall LG (1981) Review of the Hathlyacyninae, an extinct subfamily of South American “dog-like” marsupials. Fieldiana Geol New Ser 7: 1–120
Marshall LG, Salinas P (1990) Stratigraphy of the Río Frías Formation (Miocene), along the Alto Río Cisnes, Aisen, Chile. Revista Geológica de Chile 17: 57–58
Martin GM, Tejedor MF (2007) Nueva especie de Pseudonotictis Ameghino (Metatheria, Sparassodonta, Hathliacynidae) del Mioceno medio de Chubut noroccidental, Argentina. Ameghiniana 44: 747–750
Martin GM, Tejedor MF, Novo NM, González Ruiz LR, Martínez O (2016) Nuevos metaterios (Mammalia) del Mioceno medio del noroeste del Chubut (Argentina). XI Congreso de la Asociación Paleontológica Argentina, Libro de Resúmenes 65
Mazzoni MM (1993) Sedimentación Colloncurense en las provincias de Río Negro y Neuquén. Ameghiniana 30: 351
Moreno FP, Mercerat A (1891) Exploración arqueológica de la Provincia de Catamarca: paleontología. Revista del Museo de La Plata 1: 222–236
Muizon C de, McDonald HG (1995) An aquatic sloth from the Pliocene of Peru. Nature 375: 224–227
Nullo FE (1979) Descripción Geológica de la Hoja 39c, Paso Flores, escala 1: 200.000. Servicio Geológico Nacional Boletín N° 167: 1–70
Ortiz-Jaureguizar E, Cladera GA (2006) Paleoenvironmental evolution of southern South America during the Cenozoic. J Arid Environments 66: 498–532
Pascual R, Bondesio P, Scillato-Yané GJ, Vucetich MG, Gasparini Z (1978) Vertebrados. VII Congreso Geológico Argentino. Relatorio Geología y Recursos Naturales del Neuquén: 177–185
Pascual R, Bondesio P, Vucetich MG, Scillato-Yané GJ, Bond M, Tonni EP (1984) Vertebrados fósiles Cenozoicos. In: Ramos V (ed) IX Congreso Geológico Argentino, Relatorio 2: 439–461
Perkins ME, Fleagle JG, Heizler MT, Nash B, Bown TM, Tauber AA, Dozo MT (2012) Tephrochronology of the Miocene Santa Cruz and Pinturas formations, Argentina. In: Vizcaíno SF, Kay RF, Bargo MS (eds) Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation. Cambridge University Press, Cambridge, pp 23–40
Pujos F (2006) Megatherium celendinense sp. nov. from the Pleistocene of Peruvian Andes and the megatheriine phylogenetic relationship. Palaeontology 49: 285–306
Pujos F, De Iuliis G, Argot C, Werdelin L (2007) A peculiar climbing Megalonychidae from the Pleistocene of Peru and its implication for sloth history. Zool J Linn Soc 149: 179–235
Pujos F, Salas-Gismondi R, Baby G, Baby P, Goillot C, Tejada J, Antoine P-O (2013) Implication of the presence of Megathericulus (Xenarthra: Tardigrada: Megatheriidae) in the Laventan of Peruvian Amazonia. J Syst Palaeontol 11: 973–991
Rabassa J (1975) Geología de la región de Pilcaniyeu-Comallo (Provincia de Río Negro, Argentina). Fundación Bariloche. Departamento de Recursos Naturales y Energía 17: 1–128
Ramos ME, Orts D, Calatayud F, Pazos PJ, Folguera A, Ramos VA (2011) Estructura, Estratigrafía y evolución tectónica de la cuenca de Ñirihuau en las nacientes del río Cushamen, Chubut. Revista de la Asociación Geológica Argentina 68: 210–224
Ramos ME, Tobal J, Sagripanti L, Folguera A, Orts D, Giménez M, Ramos VA (2015) The north Patagonian orogenic front and related foreland evolution during the Miocene, analyzed from synorogenic sedimentation and U/Pb dating (∼42°). J So Am Earth Sci 64: 467–485
Rivas H, Bostelmann E, Le Roux J, Ugalde R (2015) Fluvial facies and architecture of the late middle Miocene, Mayoan, deposits of Chilean Patagonia. XIV Congreso Geológico Chileno Actas 1: 812–815
Roth S (1899) Apuntes sobre la geología y la paleontología de los territorios del Río Negro y Neuquén. Revista del Museo de La Plata 9: 1–56
Rovereto C (1914) Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1–247
Saint-André P-A, De Iuliis G (2001) The smallest and most ancient representative of the genus Megatherium Cuvier, 1796 (Xenarthra, Tardigrada, Megatheriidae), from the Pliocene of the Bolivian Altiplano. Geodiversitas 23: 625–645
Scillato-Yané GJ (1979) Nuevo Nothrotheriinae (Edentata, Tardigrada) de Edad Chasiquense (Plioceno temprano) del sur de la Prov. de Buenos Aires (Argentina). Su importancia bioestratigráfica, filogenética y paleobiogeográfica. VII Congreso Geológico Argentino Actas 2: 449–457
Scillato-Yané GJ, Carlini AA (1998) Nuevos Xenarthra del Friasense (Mioceno medio) de Argentina. Stvdia Geologica Salmanticensia 34: 43–67
Scott WB (1903-1904) Mammalia of the Santa Cruz Beds. Part 1: Edentata. Reports of the Princeton University to Patagonia 1896–1899 5: 1–364
Simpson GG (1942) A Miocene tortoise from Patagonia. Am Mus Novitates 1209: 1–6
Varela L, Tambusso PS, McDonald HG, Fariña RA (2019) Phylogeny, macroevolutionary trends and historical biogeography of sloths: insights from a Bayesian morphological clock analysis. Syst Biol 68: 204–218. doi:https://doi.org/10.1093/sysbio/syy058
Vera B, González Ruiz L, Novo N, Martin G, Reato A, Tejedor MF (2018) The Interatheriinae (Mammalia, Notoungulata) of the Friasian sensu stricto and Mayoan (middle to late Miocene), and the fossils from Cerro Zeballos, Patagonia, Argentina. J Syst Palaeontol. doi: https://doi.org/10.1080/14772019.2018.1511387
Vera B, Reguero MA, González Ruiz L (2017) The Interatheriinae (Notoungulata) from the Collón Curá Formation (middle Miocene), Argentina. Acta Palaeontol Pol 62: 845–863. https://doi.org/10.4202/app.00373.2017
Vucetich MG (1994) La fauna de roedores de la formación cerro boleadoras (Mioceno inferior?) en la provincia de Santa Cruz (Argentina). Acta Geológica Leopoldensia 17: 365–374
Vucetich MG, Mazzoni MM, Pardiñas FJ (1993) Los roedores de la Formación Collón Cura (Mioceno Medio), y la Ignimbrita Pilcaniyeu. Cañadon del Tordillo, Neuquén. Ameghiniana 30: 361–381
Zetti J (1964) El hallazgo de un Megatheriidae en el "médano invasor" del SW de Toay, provincia de La Pampa. Ameghiniana 3: 257–265
The authors thank to O. Martínez, G. Martin, N. Novo, A. Reato, and M. Tejedor for field assistance. We also thank the comments and suggestions of two reviewers and two editors. Funding comes from PICT 2013-2270, PICT 2014-1818 and PICT 2016-3831.
Author information
Authors and Affiliations
Laboratorio de Paleontología de Vertebrados, Centro de Investigaciones Científicas y Transferencia de Tecnología a la Producción (CICYTTP-CONICET, Prov de Entre Ríos, UADER), Matteri y España, 3105, Diamante, Entre Ríos, Argentina
Diego Brandoni
Centro de Investigación Esquel de Montaña y Estepa Patagónica (CIEMEP) (CONICET-UNPSJB), Roca 780, 9200, Esquel, Chubut, Argentina
Laureano González Ruiz
Centro de Investigaciones Geológicas (CONICET-UNLP), Diagonal 113 y 64, 1900, La Plata, Buenos Aires, Argentina
Brandoni, D., Ruiz, L.G. & Bucher, J. Evolutive Implications of Megathericulus patagonicus (Xenarthra, Megatheriinae) from the Miocene of Patagonia Argentina.
J Mammal Evol27, 445–460 (2020). https://doi.org/10.1007/s10914-019-09469-6