
Abstract
There are significant geographic gaps in our knowledge of marine mammal evolution because most fossils have been found and described from Northern Hemisphere localities and a few other high-latitude areas in the Southern Hemisphere. Here, we describe fossil cetacean remains from five geological units in the South American tropics (Urumaco, Codore, Castilletes, Cantaure, and Querales formations) generally representing marginal marine depositional environments (estuaries, deltas, and tidal flats). While fossil cetaceans from Venezuelan Neogene localities have been previously studied, this paper includes the first descriptions of fossil cetaceans from Colombia, including a diverse assemblage of mysticetes and odontocetes. We identified and provisionally referred fragmentary remains to the iniid Ischyrorhynchus vanbenedeni and to the platanistid Zarhachis flagellator. The latter suggests the presence of Platanistidae in the eastern coast of South America during the early-middle Miocene, representing the second record of Platanistidae in South America and the first record of Platanistidae in eastern South America. Other less-diagnostic specimens are characterized by features commonly seen in longirostrine odontocetes such as Iniidae, Platanistidae, Pontoporiidae, Lipotidae, Eoplatanistidae, and Squalodelphinidae.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
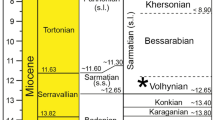
There is a significant Northern-Hemisphere bias in our understanding of whale evolution and more exploration and research should be directed to other regions of the world, including South America (Uhen 2010). Within the Southern Hemisphere, the best-sampled fossil cetacean localities are in high latitudes in Argentina (Oligocene–Miocene), Australia (Oligocene–Neogene), New Zealand (particularly Oligocene), and Peru (Miocene–Pliocene) (Cozzuol 1996; Fordyce 2009). This uneven sampling reflects a well-known bias of the fossil record in the Americas (Carrillo et al. 2015). Whereas tropical areas of the world are infamous in paleontology because of their high weathering rates and limited fossil outcrop areas, the arid climate of northernmost South America provides excellent rock exposure and a rich Neogene fossil record. Thus, research in South America offers a unique opportunity for discoveries (Sánchez‐Villagra and Aguilera 2006; Jaramillo et al. 2015). The fossil cetaceans described herein were found in two neighboring basins now exposed in northern South America: the Falcón Basin (northwest Venezuela) and the Cocinetas Basin (northern Colombia) (Fig. 1). The described specimens are dated from early Miocene to early Pliocene (Fig. 2). Previous studies reporting other fossil cetaceans from Venezuela are summarized in Table 1. Fossil cetacean remains in Colombian deposits of the Castilletes Formation (Cocinetas Basin) were for the first time mentioned in Moreno et al. (2015: table 5) and are here described.

Map of NW Venezuela (Falcón State) and N Colombia (Guajira Peninsula) showing cetacean fossil localities mentioned in the text

The Urumaco sedimentary sequence of the Falcón Basin preserves some of the best documented Miocene tropical vertebrate assemblages in the world including continental (savannas), freshwater (swamps and rivers), estuarine (brackish), and marine environments (Sánchez‐Villagra and Aguilera 2006). In contrast, the Cocinetas Basin has only recently been explored (Jaramillo et al. 2015).
The stratigraphic information (Fig. 2) and paleoenvironmental interpretations of the Castilletes Formation are based on Hendy et al. (2015) and Moreno et al. (2015); those of the Cantaure, Querales, Urumaco, and Codore formations (Falcón Basin) are based on Quiroz and Jaramillo (2010).
These new descriptions add to recent efforts to better understand the paleobiodiversity of cetaceans in South America. Apart from the ‘classic’ localities in Argentina (e.g., Moreno 1892; Lydekker 1894; Cozzuol 1996; Buono and Cozzuol 2013, Buono et al. 2016) and Peru (e.g., Muizon 1984, Muizon 1988a; Bianucci et al. 2016; Lambert et al. 2015), fossil cetaceans have been recently reported from Ecuador (Bianucci et al. 2005), Costa Rica (Valerio and Laurito 2012), Panama (Uhen et al. 2010; Pyenson et al. 2015; Vélez-Juarbe et al. 2015), and Chile (Pyenson et al. 2014).
Material and Methods
Institutional Acronyms
AMU-CURS Colección Paleontológica de la Alcaldía Bolivariana de Urumaco, Urumaco, Venezuela
MUN-STRI Museo Mapuka, Universidad del Norte, Barranquilla, Colombia
NMB Naturhistorisches Museum Basel, Basel, Switzerland
PIMUZ Paläontologisches Institut und Museum, Universität Zürich, Zürich, Switzerland
Terminological Conventions
The anatomical interpretation and terminology of the skull follow the lexicon of Mead and Fordyce (2009); terminology on teeth follows the recommendations of Smith and Dodson (2003); the interpretation and terminology of the vertebrae are based on Rommel and Reynolds (2008); finally, we follow the recommendations of Bengtson (1988) in the use of open nomenclature.
Systematic Paleontology
Cetacea Brisson, 1762
Odontoceti Flower, 1864
Delphinida Muizon, 1984
Inioidea Gray, 1846
Iniidae Gray, 1846
cf. Ischyrorhynchus vanbenedeni Ameghino, 1891

Photographs of cf. Ischyrorhynchus vanbenedeni AMU-CURS-219: a dorsal view; b lateral view; c ventral view. Abbreviations: I incisive teeth; C canine tooth; PC postcanine teeth

Photographs of cf. Ischyrorhynchus vanbenedeni AMU-CURS-165: a dorsal view; b lateral view; c ventral view

Photograph and line art of cf. Ischyrorhynchus vanbenedeni AMU-CURS-165 (rostrum close-up showing teeth alveoli). Hatching indicates major fractures
Material—Two partial skulls: 1) AMU-CURS-219, a fragmentary rostrum (a plaster cast of AMU-CURS-219 has been deposited at PIMUZ under the catalog number PIMUZ-A/V-5284); 2) AMU-CURS-165 includes most of the rostrum.
Locality—1) AMU-CURS-219 was collected in “El Vijiadero, south of Llano Grande” (11° 10.309′ N, 70° 18.216′ W); 2) AMU-CURS-165 was collected in “La Meseta, east of Casa Chiguaje” (11° 16.500′ N, 70° 11.925′ W), Urumaco town, northwestern Venezuela.
Geological framework—AMU-CURS-219 comes from the Urumaco Formation (Figs. 1 and 2), which is well exposed in the Falcón State coastal plain. The unit has been informally divided in three members: 1) the lower member, dominated by dark-gray laminated mudstones and shales with interbedding fine-grained sandstone and some coquinoidal limestone layers; 2) the middle member, characterized by interbedding medium- to fine-grained sandstone, organic-rich mudstone, shales, and coquinoidal limestones; and 3) the upper member, composed of laminated, organic-rich mudstone and shales intercalated with very fine grained sandstones (Quiroz and Jaramillo 2010). According to Quiroz and Jaramillo (2010), the environment corresponds to a prograding strandplain-deltaic complex; this interpretation is further supported by Carrillo-Briceño et al. (2015).
AMU-CURS-165 was collected in sediments belonging to the Codore Formation (Figs. 1 and 2), which extends for more than 100 km from the city of Coro to the town of Dabajuro (Venezuela). The thickness of the Codore Formation at the Urumaco River is 860 m (Quiroz and Jaramillo 2010). The Codore Formation is subdivided into three formal geological members: 1) El Jebe Member, composed of grey to reddish massive mudstones interbedding with fine-grained sandstones; 2) the Chiguaje Member, composed of dark gray laminated mudstone and shale interbedded with muddy sandstone containing abundant mollusk fragments grading to limestones with oysters; and 3) the Algodones Member, characterized by fining-upward sequences of massive-bedded to cross-stratified sandstones, grading to muddy sandstones and mudstones (Quiroz and Jaramillo 2010).
AMU-CURS-165 was found in sediments near the top of the Chiguaje Member. The Chiguaje Member represents a short transgressive event between two continental members (i.e., El Jebe and Algodones, Hambalek et al. 1994). The monospecific accumulations of oysters suggest a low-energy coastal lagoon or bay with reduced salinity (Quiroz and Jaramillo 2010). The Chiguaje Member of the Codore Formation is interpreted as a marine shallow-water depositional environment (Hambalek et al. 1994; Rey 1994). This is further supported by the presence of the foraminiferan Ammonia parkinsoniana in the Chiguaje Member, which is a dominant taxon in low-salinity habitats (Smith et al. 2010).
Age—The age of the sediments where AMU-CURS-219 was found is interpreted as late Miocene (Fig. 2), probably encompassing the Chasicoan and Huayquerian South American Land Mammal Ages (ca. 10–7 Ma; Sánchez‐Villagra and Aguilera 2006). The age of the Chiguaje Member (Fig. 2) where AMU-CURS-165 was found is interpreted as late Miocene to Pliocene (ca. 11.6–2.6 Ma) based on palynology (zone of Echitricolporites spinosus; see Hambalek et al. 1994).
Description
Rostrum
Both specimens exhibit a relatively long, high, and slender rostrum (Table 2). In dorsal view (Fig. 3a), the mesorostral groove of AMU-CURS-219 is covered by the premaxillae until 55 mm from the apex. From this level forward the mesorostral groove is dorsally open. In general, there is little lateral expansion of the maxilla anteriorly, but posteriorly the maxilla can be seen in dorsal view (Figs. 3a and 4a). In AMU-CURS-165, the maxilla extends slightly laterally from the premaxilla posteriorly where the maxillary flange marks the area near the rostrum base (Fig. 4a, b). The premaxilla widens slightly posteriorly towards the base of the rostrum (Fig. 4a).
In lateral view, the position of the premaxillary-maxillary boundary can be traced by the appearance of a lateral groove running along and deepening posteriorly in the rostrum (Figs. 3b and 4b). The premaxilla is about as high as the maxilla in the anterior part of the rostrum, but posteriorly becomes shorter and the maxilla higher in proportion to each other (Figs. 3b and 4b).
In ventral view (Fig. 3c), the anteriormost portion (4.5 cm) of AMU-CURS-219 is inferred as premaxilla, based on tooth morphology (see teeth description below) and the remaining area is formed by the maxilla, which widens posteriorly. There are a total of 19 pairs of dental alveoli preserved in AMU-CURS-219 (Fig. 3c). The alveoli become larger and more circular posteriorly, near the maxillary flange in AMU-CURS-165 (Figs. 4c and 5).
In both AMU-CURS-219 and AMU-CURS-165, the empty alveoli are relatively shallow and have an oval outline (long axis is mesiodistal) (Figs. 3c, 4c and 5). The spacing between teeth (diastemata) is greater in mesiodistal length at mid-rostrum and becomes shorter in both mesial and distal directions (best seen in AMU-CURS-219; Fig. 3c). The lengths of the diastemata on the left side of the rostrum of AMU-CURS-219 are as follows (in mm), in sequential order starting from the most mesial diastema (between I1 and I2): 7; 6.9; 10.3; 10.3; 13.7; 18.2; 23.2; 25.6; 25.6; 22.6; 17.8; 21.2; 12.6; 9.4; 7.5; 7.4; 3.9 and 3.9. The alveoli are placed asymmetrically (left alveolus more anterior than right) at mid-rostrum and more distally (Figs. 3c, 4c and 5).
Teeth
The anterior teeth are preserved only in AMU-CURS-219. The teeth missing their crowns (posterior rostrum) display clear and irregular distal surfaces, suggesting that the crowns were not worn, but probably broken during burial. The fourth pair of teeth are slightly longer and more rectilinear than the other teeth, indicating they may be canines (based on comparisons with Isthminia); if this is correct, the preserved rostrum of AMU-CURS-219 is composed of three pairs of incisors, a pair of canines, and 15 pairs of postcanine teeth (Fig. 3b). The crowns of the anterior teeth are simple (no cusps or accessory denticles) and slightly rugose in texture. The teeth are generally slightly curved with the apex pointing distoapically. It is evident in the anterior teeth that the roots are larger than the crowns and particularly expanded along the mediodistal axis.
Remarks
The feeding apparatus of cetaceans has been considered as strongly “evolutionarily plastic” and prone to homoplasy (Leduc et al. 1999; Bianucci 2005). These interpretations were made based on the finding of the highest intraspecific variation on those characters linked to the feeding apparatus in the spotted porpoise (i.e., Stenella; Perrin 1975). Despite this, aspects of the rostrum and teeth are informative interspecifically and are commonly used when identifying living (Leduc et al. 1999) and fossil cetaceans (Fordyce and Muizon 2001), but such characters may be less reliable when diagnosing deeper relationships, such as those above the species level (Leduc et al. 1999).
Long rostra are relatively common in Miocene cetaceans (e.g., eurhinodelphinids, platanistids, and dalpiazinids), and longirostrine forms have already been reported from Venezuelan Miocene localities (e.g., Squalodelphinidae, Table 1). The strong lateral compression of the rostrum of AMU-CURS-219 and AMU-CURS-165 (height/width ratio (H/W) of 1.6 and 1.8, respectively) is found in the rostrum of Platanista gangetica (H/W = 1.5, Pilleri and Gihr 1979) and in the holotype of Ischyrorhynchus vanbenedeni (H/W = 1.2, Pilleri and Gihr 1979). The rostrum of Inia geoffrensis is wider than high (H/W = 0.8, Pilleri and Gihr 1979). It should be noted that Barnes (2006: fig. 7) also mentioned and illustrated a mandibular symphysis referred to Platanistinae (LACM 131112) from the early Miocene Nye Formation (Oregon, USA) that exhibits strong lateral compression, but there is no overlap of features to be compared with AMU-CURS-219 and AMU-CURS-165.
The rostral groove present along the premaxilla-maxilla suture in AMU-CURS-219 and AMU-CURS-165 is also seen in Ischyrorhynchus, Pontoporia, and young individuals of Inia and Lipotes (Pyenson et al. 2015). This character is probably homoplastic, as it is also present in other longirostrine forms such as Platanista, Eurhinodelphis, Ziphius, Schizodelphis, Ziphiodelphis (Lambert 2005), Prepomatodelphis, Zarhachis (Barnes 2006), and Eoplatanista (Muizon 1988b).
Other key characters of AMU-CURS-219 and AMU-CURS-165 are the lengthening of the diastemata at mid-length rostrum and the bilateral asymmetry in the longitudinal position of the dental alveoli (Fig. 3c). Both characters are probably linked to the extreme lateral compression of the rostrum and have also been reported for Ischyrorhynchus and Platanista (Pilleri and Gihr 1979). Interestingly, Geisler et al. (2012) recovered a sister-group relationship for Ischyrorhynchus and Platanista. In contrast, Pyenson et al. (2015) rescored several characters of Ischyrorhynchus after direct observation of the holotype and referred specimens, and recovered Ischyrorhynchus in a clade that also includes Isthminia and Inia, while Platanista gangetica was found in a clade shared with Lipotes and Parpontoporia. The position of Platanista in the cladogram of Pyenson et al. (2015) was interpreted as a spurious effect possibly attributed to long-branch attraction because both exclusively-molecular and combined molecular and morphological analyses consistently recover Platanista as a separate, basal branching clade from Lipotes and Inioidea (Geisler et al. 2011: figs. 1–2, and references therein).
To conclude, the remarkable similarities of AMU-CURS-219 and AMU-CURS-165 with both Ischyrorhynchus and Platanista represent, in at least one case, the product of homoplasy. Based on the available evidence, we provisionally place AMU-CURS-219 and AMU-CURS-165 in Ischyrorhynchus based on the morphological similarities described above and the known biogeographic record of Iniidae in both the North and South Atlantic. The paleobiogeography of Iniidae is briefly addressed in the discussion section.
?Iniidae
gen. et sp. indet.
Figs. 6, 7 and 8, Tables 3 and 4

Photographs and line art of the skull of Iniidae gen. et sp. indet. MUN-STRI-37698: a dorsal view; b lateral view (flipped view of right side of skull); c ventral view. Bones in ventral view (c) are undifferentiated and therefore not shown in line art. Stippling indicates areas covered by matrix

Photographs of the mandible and posterior tooth of Iniidae gen. et sp. indet. MUN-STRI-37698: a dorsal view; b tooth in labial (left) and lingual (right) views; c ventral view. Stippling indicates areas covered by matrix

Photographs of the isolated tooth of Iniidae gen. et sp. indet. MUN-STRI-37698: a labial view; b lingual view; c line drawing of the lingual view. Hatching indicates area of occlusal tooth wear
Material—MUN-STRI-37698 (skull) and MUN-STRI-37699 (five vertebrae), both belong to the same specimen.
Locality—“Yotojoro” (STRI-490006; 11° 56.537′ N, 71° 18.913′ W), Guajira Peninsula, northern Colombia (see also Hendy et al. 2015: table 10; Moreno et al. 2015: fig. 3J, table 4). The specimen was found in two strongly weathered concretions with some isolated elements (including teeth and vertebrae) scattered around.
Geological framework—Castilletes Formation (Figs. 1 and 2). The Castilletes Formation is composed of gray massive mudstone and siltstone, fossiliferous wackestone to packstone, and medium-grained to conglomeratic fossiliferous lithic to quartz sandstone (Moreno et al. 2015). The paleoenvironment of the Castilletes Formation is generally interpreted by Hendy et al. (2015) as shallow marine with sea-level oscillations including non-marine fossil invertebrate assemblages and continental vertebrates in some areas.
Age—A late Burdigalian–Langhian (ca. 16.7–14.2 Ma) was assigned to this formation (Moreno et al. 2015: fig. 2). Based on a calibrated age model obtained from 87Sr/86Sr isotope dates, the extrapolated age of locality STRI-490006, is reported as 16 Ma (Hendy et al. 2015: table 10).
Description
Skull
The skull bones are very crumbly and softer than the matrix, making preparation difficult. The bone was heavily consolidated with Acryloyd B-72 and sutures are consequently difficult to observe.
In dorsal view (Figs. 6a), most of the rostrum is composed of both premaxillae. Each premaxilla is curved dorsally, forming a hemi-cylindrical structure. Medially, the open mesorostral canal is filled with sediment. A small (1–2 cm) portion of the maxilla flanges laterally the premaxilla on the better-preserved right side of the skull. In the neurocranium, the ascending process of the maxilla widens posteriorly. It is inferred that post-mortem compression caused the ascending process and its maxillary crest to collapse and appear in a more ventral position than would have been in life. The maxillary crest is inferred to have been similar to the longitudinal structure seen on the supraorbital process of the maxilla of Pontoporia, Inia, and Isthminia. The ascending process of the right premaxilla ends posteriorly in a knob-like shape as in Inia (Fig. 6a, b), but reaches a position near the posterior end of the naris as in Pontoporia. The relatively large nares (Table 3) are asymmetrically positioned (left naris ca. 2.5 cm more anteriorly positioned than the right). Asymmetry in the position of the nares is uncommon in inioids (although there is slight asymmetry in Brachydelphis), and interpreted for MUN-STRI-37698 as the product of post-mortem deformation. The posterior part of the cranium possibly holds small undifferentiated fragments of frontal, nasal, maxilla, and premaxilla.
In lateral view (Fig. 6b), the rostrum is relatively high, with the upper half composed of premaxilla and the lower half composed of maxilla. The bones along the ventral surface (Fig. 6c) are poorly preserved as to be described in any detail and are therefore not reproduced as line art.
Mandible
The anterior half of the mandible and several isolated teeth (probably mandibular) were found lying on the surface near the concretion containing the skull. The posterior half of the mandible was prepared and removed from the concretion. The isolated mandibular fragments are highly eroded and lack direct contact, but exhibit more detail than the posterior part of the mandible that was removed from the concretion (Fig. 7a–c). The preserved length of the mandible is ca. 94 cm (Table 3), but the complete length would have been slightly greater, given that both mandibular condyles and anterior fragments of the mandible are lost. Both mandibles are fused anteriorly, forming an osseous symphyseal articulation of more than a half of the total length of the mandible (Table 3).
The five fragments of the body of the mandible (anterior half) exhibit large oval (longer than wide) dental alveolar outlines in dorsal view (Fig. 7a). The mandibular fragments at the symphysis have a width of approximately 4 cm. There are at least nine dental alveoli on each symphyseal dental row (Fig. 7a). The dental alveoli are deeper and narrower posteriorly, and shallower but wider anteriorly. The size of the alveoli varies slightly but in general is about 2 cm long and 1.2 cm wide. The diastemata are also relatively long (1.2 cm). Posteriorly, the mandibular fossa is more than 18 cm long and is 9 cm high (Table 3). The number of mental foramina is unknown but one of the mandible fragments preserves two mental foramina on the ventral face of each component of the symphysis. A second fragment of the mandible exhibits longitudinal sulci that are probably associated to other mental foramina (Fig. 7c).
Teeth
A total of 20 teeth or teeth fragments assumed to belong to MUN-STRI-37698 were found. Most teeth fragments are roots and were found lying on the ground near the anterior half of the mandible. Among them was found a complete tooth (Fig. 8) that is described in more detail below. A second tooth was found attached to the mandible and extracted for further study (Fig. 7a, b).
Both teeth preserving the crown are denticulate (Figs. 7b and 8) and exhibit one accessory denticle placed on the distal face (Figs. 7b and 8) and a lingual flange (sensu Best and da Silva 1993) is located at mid-height of the crown. The crowns of both teeth are slightly curved distally and have a rugose texture, with striae running along the vertical (apex-base) axis. Both teeth have a constriction at the cervix and become more voluminous in the root just basal to the cervix.
The root of the complete tooth (Fig. 8) and other isolated teeth roots (not pictured) taper and curve markedly towards the apex of the root. The isolated tooth exhibits prominent sub-vertical occlusion tooth wear on its lingual face (Fig. 8).
Vertebrae
The five vertebrae catalogued as MUN-STRI-37699 were found in a second concretion near (few centimeters) MUN-STRI-37698 (Online Resource 1, Fig. S4). They preserve mainly the centra; the transverse and spinous processes are broken. The vertebrae are similar in dimension (Table 4) and inferred to be from roughly the same region. The proportion and convexity of the articular faces and the presence of vertebrarterial canals in MUN-STRI-37699-1 (Online Resource 1, Fig. S4a) indicate that these vertebrae are from the caudal region.
Remarks
MUN-STRI-37698 is strongly weathered and details on bones are missing. The combination of features such as the long and fused mandibular symphysis, the elevated vertex, the knob-like ascending process of the premaxilla, the presence of a maxillary crest (similar to those of Inia, Isthminia, and Pontoporia), and the presence of a lingual flange on the teeth (similar to that of the teeth of Isthminia and Inia) suggests MUN-STRI-37698 is likely an iniid with close affinities to Inia and Isthminia. The roots of the teeth of MUN-STRI-37698 differ from those of Isthminia in not being as voluminous. The mental foramina of MUN-STRI-37698 are probably fewer in number than in Isthminia (seven mental foramina, Pyenson et al. 2015) and more similar to Pontoporia and Brachydelphis (one to two pairs of mental foramina).
Platanistida Fordyce and Muizon, 2001
Platanistoidea Gray, 1863
Platanistidae Gray, 1863
cf. Zarhachis flagellator Cope, 1868

Photographs of cf. Zarhachis flagellator: a NMB-S.A.1537, dorsal view; b NMB-S.A.1537, ventral view; c NMB-S.A.1538, dorsal view; d NMB-S.A.1538, ventral view
Material—NMB-S.A.1537 and NMB-S.A.1538, two vertebrae of the same individual collected by Jack and Winifred Gibson-Smith in 1975 and donated to NMB in 1998. The vertebrae were identified by Bryan Patterson (Harvard University) as belonging to a cetacean (NMB, in litt).
Locality—The exact locality is unknown, but labeled as “GS-103-PGNA, Northern scarp of Cerro San Carlos, near San José del Cocodite, Paraguaná Peninsula, NW Venezuela, Fm ? early or middle Miocene” in the notes left by Jack and Winifred Gibson-Smith in the collection data. The associated invertebrate fauna collected in the same locality by J. and W. Gibson-Smith includes Turritela (Bractospira) altilira (the most abundant, 32%), Pecten (Aequipecten) “sp. A” (24%), Glycymeris (Tucetona) cf. secticostata schenki, and Terebra (Strioterebrum) spirifera (3%), as labeled in the collections of the NMB. Faunas of other localities in the Paraguaná Peninsula were published by the Gibsons (e.g., Gibson-Smith 1979; Gibson-Smith and Gibson-Smith 1983), but not GS-103-PGNA.
Two of the authors (RS and GAF) visited Cerro San Carlos on 07.02.2015 and located a fossil-vertebrate-bearing locality mentioned in the literature and known as “El Yacural,” regarded by Aguilera et al. (2008) as containing sediments of the Paraguaná Formation (Pliocene, Fig. 1), but the locality is stratigraphically below the limestone cap that marks the upper limit of the Cantaure Formation as defined in Jung (1965) and Hunter and Bartok (1974), and therefore belongs to the Cantaure Formation. A cetacean vertebra with an embedded tooth of a Charcharocles was reported from this locality (Aguilera et al. 2008) and more cetacean material (a vertebra and an unrelated skull fragment; AMU-CURS- 709 and 710, respectively) and a crocodilian vertebra (AMU-CURS-708) were found during our reconnaissance trip at the following geographic coordinates: 11° 55.803′ N and 70° 00.679′ W. El Yacural may be the source of NMB-S.A.1537 and NMB-S.A.1538 and comparison of the associated invertebrate fauna may help to solve the issue.
Geological framework—The formation is marked as unknown in the notes of J. and W. Gibson-Smith deposited at NMB, but regarded in their notes as early or middle Miocene in age. There are two recognized formations exposed in the Paraguaná Peninsula: the Cantaure Formation (Burdigalian), and the Paraguaná Formation (Pliocene) (Figs. 1 and 2).
Here, we regard the specimens NMB-S.A.1537 and NMB-S.A.1538 as belonging to the Cantaure Formation, which is restricted to a small area near Pueblo Nuevo in the central Paraguaná Peninsula of Venezuela. The lithology consists of silty shales interbedded with algal limestones and shell beds (Aguilera et al. 2013). Based on the representative fauna (mollusks, crustaceans, and fish), the environment is interpreted as a shallow-water marine setting (Aguilera et al. 2013), which is further supported by the highly diverse elasmobranch faunal assemblage (Carrillo-Briceño et al. 2016).
Age—The collection card at NMB notes “early or middle Miocene” but this statement could not be verified. A sample of the sediment matrix encrusted in NMB-S.A.1537 was removed and prepared for micropaleontological analysis. The sample did not yield any foraminifera, coccolithophores, or any other forms of nannoplankton. As mentioned above, given the present knowledge of the geology of the Paraguaná Pensinsula and the data available in the collection record, we assume that specimens NMB-S.A.1537 and NMB-S.A.1538 belong to sediments of the Cantaure Formation. The Cantaure Formation is interpreted as early Miocene in age (Díaz de Gamero 1974; Rey 1996). A more precise dating of 16.5 ± 0.4 Ma was obtained by Griffiths et al. (2013) based on 87Sr/86Sr geochronology on corals from a site nearby. The age of the Cantaure Formation is here regarded as Burdigalian (20–16 Ma).
Description
NMB-S.A.1537 and NMB-S.A.1538 are two vertebrae of one individual (Fig. 9, Table 5). Both vertebrae are probably from the anterior lumbar region, according to the ventral position of the base of each transverse process. The dimensions (Table 5) closely resemble those provided for the vertebrae of the pomatodelphinine Zarhachis flagellator by Kellogg (1924: 33, pls. 10 and 15). Both NMB-S.A.1537 and NMB-S.A.1538 preserve the complete length of their centrum, but the transverse process, the neural arch, and the neural spine are broken away (Fig. 9).
Both vertebrae are cylindrical and slightly compressed dorsoventrally, the base of the transverse process is very broad (ca. 70% the centrum length). Ventrally (Fig. 9b, d), a partially-eroded, but well-developed hypophysis (“median keel” sensu Kellogg 1924) runs along the midline. The pair of “grooves” meeting medially that “characterize the fourth lumbar” of Zarhachis according to Kellogg (1924:32) are present in both NMB-S.A.1537 and NMB-S.A.1538 and here termed “hypovertebral grooves” (new term, Fig. 9d).
Remarks
The vertebrae NMB-S.A.1537 and NMB-S.A.1538 are remarkably long, with a centrum length to width ratio (CL/CW) of 1.8 and 1.9, respectively; the vertebrae are similar in dimensions, but slightly longer than those of Zarhachis (CL/CW = 1.4–1.9, Table 5). Other cetaceans with remarkably long vertebrae include archaeocetes, ziphiids, and kentriodontids. For example, Basilosaurus cetoides has a CL/CW = 1.9–2 according to Buchholtz (2001), but the vertebrae of Basilosaurus were much larger of those reported here for cf. Zarhachis. Additionally, the fossil record of Basilosauridae lasts until the end of the Eocene (Fordyce and Muizon 2001). The last two lumbar and first two caudal vertebrae measured in Omura (1972: appendix) of Ziphius cavirostris (the longest vertebrae along the vertebral column) have an average CL/CW of 1.4, which is significantly less than in NMB-S.A.1537 and NMB-S.A.1538. The ziphiid Ninoziphius platyrostris from the Sud-Sacaco vertebrate level of the Pisco Formation (Peru) includes one lumbar vertebra preserved and the CL/CW is 1.3 (Muizon 1984: 160); this taxon seems to be the only fossil ziphiid with available information on the dimensions of similar (lumbar or first caudal) vertebrae. The vertebrae of Kentriodon pernix are also relatively long, with a maximum CL/CW of 1.4 (Kellogg 1927).
It is important to remark that the holotype of Zarhachis flagellator consists of four vertebrae assumed to belong to three individuals (Cope 1868), and only a subsequent description of a skeleton including the skull and ear bones (Kellogg 1924) rendered its taxonomy into stability and therefore is used here as a basis for comparison. Zarhachis tysonii and Z. velox, also described by Cope (1869) based on isolated vertebrae, have a debated taxonomic status and are here regarded as nomina dubia and referred to Z. flagellator. Names of each species sensu Cope (1869) of Zarhachis are provided in Table 5 to aid comparisons.
Most specimens of Zarhachis flagellator have been collected in sediments of the Calvert Formation (Maryland, USA), which is regarded as late-early to middle Miocene in age and interpreted paleoenvironmentally as a shallow marine setting (Barnes 2006). Bianucci and Landini (2002) reported the presence of Zarhachis in Burdigalian sediments of southern Germany, suggesting a wider distribution. The assignation of NMB-S.A.1537 and NMB-S.A.1538 to cf. Zarhachis flagellator suggests an extension of the distribution of Zarhachis from the North Atlantic to the Caribbean Sea (12° N). Pending the discovery of more diagnostic material, NMB-S.A.1537 and NMB-S.A.1538 support the presence of Platanistidae in South America as suggested by Bianucci et al. (2013) based on a highly diagnostic periotic ear bone from sediments of the Peruvian Amazonia (western South America; late middle Miocene, ca. 13.2–12.5 Ma). The putative fossil platanistid Tachyrgrhynchus mentioned in the faunal list of Odreman-Rivas (1997) has not been described nor illustrated or referred to any collection and was not recorded in the extensive systematic catalogue of Mones (1986). Tachyrgrhynchus is here considered as nomen nudum (Table 1).
Odontoceti indet.
gen. et sp. indet. 1

Photographs of Odontoceti indet., gen. et sp. indet. 1 AMU-CURS-1021: a dorsal view; b ventral view
Material—Specimen AMU-CURS-1021, fragment of mandible with fused symphysis.
Locality—“Quebrada Chiquita” (11° 14.867′ N, 70° 11.756′ W). The specimen was found as a float (ex situ) in a small ravine near the town of Urumaco, northwestern Venezuela.
Geological framework—Urumaco Formation (Figs. 1 and 2), the specimen probably derives from the upper member. A summary of the stratigraphy of the Urumaco Formation is provided above and further information is detailed in Quiroz and Jaramillo (2010).
Age—The age is interpreted as late Miocene (Fig. 2), probably encompassing the Chasicoan and Huayquerian South American Land Mammal Ages (ca. 10–7 Ma according to Sánchez‐Villagra and Aguilera 2006).
Description
Both the teeth and external bone of AMU-CURS-1021 are strongly weathered. The mandible is small (ca. 1.5 cm wide at the anterior portion of symphysis; Fig. 10a, b; Table 6). There are seven comparatively large dental alveoli on each side of the mandible. The alveoli have subcircular outlines at the margins and are spaced uniformly along the tooth rows (Fig. 10a; Table 6); the diastemata are small (Fig. 10a).
Remarks
AMU-CURS-1021 strongly resembles previously-illustrated AMU-CURS-48 (Aguilera 2004) in dimensions of the mandible and alveoli (photos of AMU-CURS-48 are supplied in the Online Resource 1, Fig. S7). AMU-CURS-48 (Corralito locality, upper member of Urumaco Formation) was identified as Iniidae by Aguilera (2004), but both AMU-CURS-1021 and AMU-CURS-48 are poorly preserved and lack definite characters for a confident designation to Iniidae. The fused mandibular symphysis is present in all living ‘river dolphins’ (Iniidae, Pontoporiidae, Lipotidae, Platanistidae) and some fossil genera within those families (e.g., Ischyrorhynchus, Brachydelphis, Parapontoporia, Zarhachis, respectively) (Pilleri and Gihr 1979; Barnes 1985; Gutstein et al. 2009; Geisler et al. 2011), and also in: Eurhinodelphinidae (e.g., Xiphiacetus) and Ziphiidae (e.g., Ninoziphius, Tasmacetus) according to Geisler et al. (2011), Allodelphinidae (e.g., Allodelphis) and Squalodelphinidae (e.g., Notocetus) according to Barnes (2006), Eoplatanistidae (e.g., Eoplatanista) and Dalpiaziniidae (e.g., Dalpiazina) according to Muizon (1988b), Kentriodontidae (e.g., Kentriodon) according to Kellogg (1927), and Physeteridae (e.g., Physeter) and Squalodontidae (e.g., Squalodon) according to Fordyce (1994).
gen. et sp. indet. 2

Photograph of Odontoceti gen. et sp. indet. 2 AMU-CURS-588, ventral view. Stippling indicates areas covered by matrix
Material—AMU-CURS-588, skull embedded in a hard matrix.
Locality—“Quebrada Becerra or Cascajal” (11° 06.774′ N, 70° 12.888′ W), near the town of Urumaco, northwestern Venezuela. The specimen was found lying at the top of a highly bioturbated, hard, fine sandstone with the ichnofossils Thalassinoides (cylindrical to elliptical burrows forming Y- or T-shaped branching polygonal networks) and Planolites (unlined, rarely branched, generally straight and smooth burrows). No other macrofossils have been collected at this locality, with the exception of a Myliobatidae tooth found while extracting the cetacean remains. The environment of the sandstone beds has been interpreted as offshore marine (Quiroz and Jaramillo 2010). The Quebrada Becerra locality is about 800 m away from the Quebrada Corralito locality, detailed in Pérez et al. (2016). The sandstones where AMU-CURS-588 was collected are very similar to those described by Pérez et al. (2016) at the base of their section.
Geological framework—The Querales Formation (Figs. 1 and 2) was originally described in Hodson (1926). Sediments of the Querales Formation can be found south of the city of Coro and the town of Urumaco (Quiroz and Jaramillo 2010). The lithology is composed by dark gray laminated mudstone shales interbedded with fine to very fine sandstone layers, which represent repeated series of transgressive-regressive phases in a deltaic front (Pérez et al. 2016). Bivalves are common in the shales and the ichnofossils Thalassinoides and Planolites are found in the sandstone layers (Quiroz and Jaramillo 2010; Pérez et al. 2016).
Age—Based on the presence of the planktonic foraminiferans Praeorbulina glomerosa and Fohsella (Globorotalia) peripheroronda, Díaz de Gamero (1989) suggested an age at the boundary of early and middle Miocene for the Querales Formation (Fig. 2). Similarly, the planktonic foraminiferans and calcareous nannofossils found at Quebrada Corralito suggest a late early Miocene to early middle Miocene age (Pérez et al. 2016). The Querales Formation is contemporaneous with the Cantaure Formation described above (Moreno et al. 2015).
Description
Skull of longirostrine odontocete (Fig. 11), embedded in a hard, calcareous matrix. The ventral surface of the rostrum was partially exposed by erosion and further prepared. A CT-scan was performed in Coro, Venezuela, but the contrast was poor so that bone and matrix could not be differentiated. There are about 50 teeth on each dental row (Table 6) and all teeth appear to be single-rooted.
Remarks
Despite that different bones are preserved in AMU-CURS-588 (a partial skull) and AMU-CURS-1021 (a mandible, described above), the rostrum and mandible allow comparisons because there is some degree of conservatism in their relative anatomical dimension. The rostrum of AMU-CURS-588 (3 cm width at mid length, Table 6) is too wide to belong to the same specimen as the mandible of AMU-CURS-1021 (1.5 cm wide, Table 6).
The rostrum of AMU-CURS-588 differs from Platanista, Inia, and Ischyrorhynchus in being broader and having symmetrically-spaced teeth alveoli (Pilleri and Gihr 1979). The ratio of rostral length to condylobasal length in AMU-CURS-588 (ca. 64%; Table 6) is similar to that reported for Notocetus vanbenedeni (Squalodelphinidae) from Argentina and Peru (67–68%; True 1910; Bianucci et al. 2015) and Tagicetus joneti (Kentriodontidae) from Portugal (71%; Lambert et al. 2005). Other comparable Miocene longirostrine forms include: Eurhinodelphis cocheteuxi (Eurhinodelphinidae; >75%) (Lambert 2005), but AMU-CURS-588 lacks the diagnostic apical portion of edentulous premaxilla that characterizes this family (Lambert 2005); Messapicetus longirotris (Ziphiidae), but the rostral contribution to the total length of the skull in AMU-CURS-588 is significantly less than in Messapicetus spp. (75%; Bianucci et al. 2010); AMU-CURS-588 is proportionally similar to Chilcacetus cavirhinus (Odontoceti incertae sedis; 72%; Lambert et al. 2015).
Mysticeti Gray, 1864
Mysticeti indet.
Fig. 12

Photographs of the rostrum of Mysticeti indet. MUN-STRI-38236: a dorsal view; b ascending process of maxilla, in lateral view; c ventral view
Material—MUN-STRI-38236, incomplete mysticete rostrum.
Locality—“Yotojoro” (STRI-340072; 11° 56.478′ N, 71° 18.966′ W), Guajira Peninsula, northern Colombia. The specimen was found in a discrete limestone layer within a mudstone that underlies stratigraphically the sandstone of locality STRI-490006 (also known as Yotojoro, described above; see also Moreno et al. 2015: fig. 3J).
Geological framework—Castilletes Formation (Figs. 1 and 2). A brief description of this formation has been detailed above and further information is available in Hendy et al. (2015) and Moreno et al. (2015).
Age—A late Burdigalian–Langhian (ca. 16.7–14.2 Ma) age is assigned to this formation (Moreno et al. 2015). The Yojotorro locality is chronologically slightly older than the locality STRI-490006, which was extrapolated as 16 Ma (Hendy et al. 2015: table 10).
Description
MUN-STRI-38236 is an incomplete mysticete rostrum preserved to the level of the antorbital process; it includes mostly the maxillae and premaxillae held together in a jacket (Fig. 12a–c), and other broken and displaced small fragments of the rostrum (premaxilla, maxilla, vomer; not figured). The degree of dorsal arching of the rostrum is unknown because of potential deformation caused during burial. The skull was found ventral-side-up and the bones were colonized by barnacles in some areas. The maximum length of the preserved skull is 108 cm, the rostral length as preserved is 90 cm, and the distance between the lateral margins of both antorbital processes is 25 cm.
In dorsal view (Fig. 12a), the lateral outline of the rostrum of MUN-STRI-38236 is relatively linear. The maxilla is relatively long and slender, while the premaxilla widens anteriorly. The posterior end of the ascending process of the maxilla that reaches the vertex is very short (Fig. 12b), and the exact position of the ascending process of the maxilla compared to the rest of the skull is unknown.
In ventral view (Fig. 12c), the premaxilla is inferred to form part of the anterior rostrum, and the ventral face of the maxilla (that overlies the premaxilla) is exposed, showing the spongy texture of the trabecular bone.
Remarks
The age range of MUN-STRI-38236 coincides with a peak in taxonomic diversity of mysticetes at 15 Ma that reflects the relative early diversification of balaenopterids (Marx and Fordyce 2015).
The epifauna attached to the mysticete skull MUN-STRI-38236 further indicates that the locality Yotojorro (Castilletes Formation) contains sediments representing a shallow marine environment as suggested in Hendy et al. (2015) because barnacles live either in the intertidal (26%) or sublittoral areas of ≤100 m (73%) (Doyle et al. 1996).
Cetacea indet.

Photographs of the partial skull of Cetacea indet. MUN-STRI-16869: a dorsal view; b ventral view. Stippling indicates areas covered by matrix
Material—MUN-STRI-16869, an incomplete skull. One rib was collected near the skull and assigned to the same field collection number (herein named MUN-STRI-16869-B; Online Resource 1, Fig. S11). The rib may not be associated with the skull because: 1) the distal end of the body is stout and tubular, while in cetaceans the distal end tends to be longitudinally compressed; 2) the size of MUN-STRI-16869-B possibly matches the range of an anterior rib of MUN-STRI-16869, but the anterior ribs of cetaceans frequently exhibit two articulation facets (Rommel 1990), while MUN-STRI-16869-B only presents one facet.
Locality—“La Tienda” (STRI-290423; 11° 55.560′ N, 71° 21.564′ W), Guajira Peninsula, northern Colombia (see also Hendy et al. 2015: table 6; Moreno et al. 2015: table 3). Common invertebrates found in this locality include Anadara sp. cf. A. zuliana and Turritella matarucana, which indicate a shallow marine (5–10 m) setting (Hendy et al. 2015).
Geological framework—Castilletes Formation (Figs. 1 and 2). MUN-STRI-16869 was found at the eastern margin of the Cocinetas Basin. A description of this formation has been detailed above and further information is available in Hendy et al. (2015) and Moreno et al. (2015).
Age—A late Burdigalian–Langhian range (16.7–14.2 Ma; Fig. 2) was assigned to this formation based on 87Sr/86Sr isotope geochronology (Moreno et al. 2015). Mollusk shells collected in locality STRI-290423 yielded an age range of 16.32–14.94 Ma (mean = 15.56 Ma; Hendy et al. 2015: table 6).
Description
MUN-STRI-16869 is a skull fragment that includes the posterior basicranium and occipital condyles. The basioccipital crest of MUN-STRI-16869 is a clear cetacean feature. In cetaceans, the basioccipital crest isolates the tympano-periotic region from the dorsal pharyngeal region (Mead and Fordyce 2009). The preserved bones include incomplete portions of the supraoccipital, exoccipital, basioccipital, basisphenoid, vomer, and squamosal. Anteriorly, a small portion interpreted as the basisphenoid is visible in dorsal view (Fig. 13a). The squamosal forms the lateral walls of the cranium. Posteriorly, the occipital condyles (exoccipital) are prominent. The foramen magnum is oval (wider than high, Table 7). The small preserved area of supraoccipital is relatively steep. In posterior view (not figured), the medial edge of the occipital condyles diverges dorsolaterally 20° from the midline.
In ventral view (Fig. 13b), the most anterior part of the preserved cranium is interpreted as the posterior part of a thin layer of vomer underlying the basisphenoid. Posteriorly, the basioccipital crests widen, and the maximum width across them is found near the jugular notches (Fig. 13b, Table 7); the angle between the two basioccipital crests is approximately 25°. The paroccipital process of the exoccipital continues a crest posterodorsal to the jugular notch. A small fragment of smooth bone on the right side is probably the tympanosquamosal recess.
Remarks
The small size of the skull (bicondylar width 73 mm) of MUN-STRI-16869 is more distinctive for an odontocete because mysticetes are generally larger. In living cetaceans, a similar bicondylar width is seen, for example, in Neophocaena phocaeinoides (75 mm), Stenella attenuata (78 mm), and Inia geoffrensis (79 mm) (Nummela et al. 2007). Notably, some Oligocene aetiocetid mysticetes such Fucaia (estimated bicondylar width 60 mm) and Chonecetus were the size of a small odontocete (Marx et al. 2015).
Discussion
Paleoenvironment and Paleoecology
The Querales and the Cantaure formations (Falcón Basin, Venezuela) are contemporaneous with the Castilletes Formation (Cocinetas Basin, Colombia) (Moreno et al. 2015: fig. 2). The interpretations on depositional environments are laterally and temporally variable, but are generally regarded as representing a proximal marine setting. The Cantaure and Castilletes formations are faunistically close, sharing 31% of the mollusk taxa (Hendy et al. 2015).
Farther up the sequence in the Falcón Basin, the Urumaco Formation (Fig. 2) is interpreted as a prograding strandplain-delatic complex (Quiroz and Jaramillo 2010) with different successive faunal assemblages representing different environments: a shallow marine environment is represented by fish and mollusks; brackish environment is represented by marine catfish; swampy environments are represented by crocodilians, turtles, and catfish of mixed origin (marine and freshwater; Sánchez‐Villagra and Aguilera 2006). The diverse vertebrate fauna suggests a paleoenvironment associated with coastal lagoons and estuaries, especially the latter (Sánchez-Villagra et al. 2010; Scheyer et al. 2013; Carrillo-Briceño et al. 2015). Our interpretation of AMU-CURS-219 and AMU-CURS-165 as Ischyrorhynchus vanbenedeni combined with the environmental interpretation of the Urumaco Formation as estuarine are in agreement with the interpretation of the Argentinian localities (e.g., Paraná, La Plata, la Curtiembre) where several specimens of Ischyrorhynchus have been previously collected (Ameghino 1891; Cozzuol 1996).
Cozzuol (2006) compared the Acre (Southwestern Amazonia) amniote faunal assemblage with other well-known localities in South America (La Venta, Colombia; Urumaco, Venezuela; Mesopotamian from Argentina and Uruguay) and found the strongest similarity among Acre and Urumaco and this relationship was further supported by comparisons of terrestrial mammal associations across South America (Carrillo et al. 2015). The aquatic and terrestrial connections along these distant localities are evidenced by the faunal overlap in both realms (Cozzuol 2006). In regards to cetaceans, care should be taken because the occurrences in Acre are reduced to a list without descriptions or illustrations of the material referred (Cozzuol 2006: appendix B). Likewise, the presence of Ischyrorhynchus and Saurocetes in Urumaco were arguable because the occurrence data stem from a publication by Linares (2004) in which those specimens are not formally described nor illustrated. Further, we briefly discuss below whether Ischyrorhynchus and Saurocetes should be considered separate species.
Implications on the Fossil Record and Diversity of Iniidae
An exhaustive review of the fossil record of Iniidae is out of the scope of this paper; furthermore, the fossil record of Inioidea was recently reviewed by Cozzuol (2010). The taxonomic history of Iniidae remains unstable and the recent use of phylogenetic frameworks on inioids (Geisler et al. 2012; Pyenson et al. 2015) offers a better perspective to solve the long-standing taxonomic puzzle.
The specimens described here and interpreted as iniids lack essential information to unambiguously provide species-level identifications. In spite of this, it is worth noting that most known fossil iniid species have been described based on relatively incomplete, sometimes undiagnostic material. Isthminia panamensis, a fossil known from an incomplete skull, mandible, teeth, a partial right scapula, and two carpals is probably the most complete fossil iniid known to date (Pyenson et al. 2015). Saurocetes is currently only known from isolated teeth, and rostral and mandibular fragments. The taxonomy of Saurocetes was considered as highly unstable even at family level by Cabrera (1926) and Pilleri and Gihr (1979). Regardless, the practice of assigning new species names to relatively incomplete specimens has prevailed (e.g., Cozzuol 1988). Given the lack of informative material of Saurocetes spp. and the close similarity of some putative Saurocetes and Ischyrochynchus specimens (see Pilleri and Gihr 1979 for an overview on the taxonomic conundrum of Ischyrorhynchus-Saurocetes), we question whether Saurocetes should be considered a separate taxon, an issue already raised by Muizon (1988c: 182).
Zarhachis in South America
The postcranial skeleton of cetaceans has been generally regarded as of limited taxonomic use (Fordyce and Muizon 2001) and sometimes may not be reported in anatomical descriptions of living and fossil cetaceans. Some studies (e.g., Slijper 1936; Buchholtz 2001; Buchholtz and Schur 2004) have provided useful functional and evolutionary frameworks to be used on fragmentary material, such as fossils. The proportion of the lumbar vertebrae is indeed a character used in cladistic analyses (e.g., character 286 of Geisler et al. 2011).
Our identification of Zarhachis in Miocene sediments of Venezuela is based on two vertebrae (NMB-S.A.1537 and NMB-S.A.1538) of one individual. This identification is provisional pending more diagnostic finds and it should be treated as ambiguous or unstable, and the lack of contradictory data may represent ignorance or a true basis of relationship. Either way, this may serve as a reminder of the utility of including descriptions of postcranial material of fossil taxa when available. Our comparisons of proportions of lumbar vertebrae led to one close match being Zarhachis. The attribution to the family Platanistidae is based on the similarity of NMB-S.A.1537 and NMB-S.A.1538 to Zarhachis and not on more inclusive, family-based synapomorphies.
Conclusions
The taxa identified in this paper provide a general overview of the paleodiversity of cetaceans in northern South America during the Miocene and early Pliocene. We report the presence of iniids in the Neogene sequences, which is in agreement with previously-published mentions in which taxa were not described. The assignation of AMU-CURS-219 and AMU-CURS-165 to cf. Ichyrorhynchus and their estuarine paleoenvironmental interpretation (Urumaco Formation) supports previous similar interpretations for the Argentinian localities (Paraná, La Plata, la Curtiembre) where most specimens of Ischyrorhynchus have been collected.
The possible occurrence of cf. Zarhachis flagellator in early–middle Miocene sediments of Venezuela, if correct, represents the first potential record of a platanistid in eastern South America and the second potential record of Zarhachis outside the Chesapeake Group (North West Atlantic). Other material, such as the mysticete, and more fragmentary odontocetes indicate a greater diversity in the area that remains mostly obscure until better-preserved taxa are found and described.
References
Aguilera OA (2004) Tesoros paleontológicos de Venezuela: Urumaco, patrimonio natural de la humanidad. Editorial Arte, Venezuela
Aguilera OA, García L, Cozzuol MA (2008) Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontol Z 82:204–208. doi: 10.1007/BF02988410
Aguilera OA, Moraes-Santos H, Costa S, Ohe F, Jaramillo C, Nogueira A (2013) Ariid sea catfishes from the coeval Pirabas (northeastern Brazil), Cantaure, Castillo (Northwestern Venezuela), and Castilletes (north Colombia) formations (early Miocene), with description of three new species. Swiss J Palaeontol 132:45–68
Ameghino F (1891) Caracteres diagnósticos de cincuenta especies nuevas de mamíferos fósiles argentinos. Rev Argent Hist Nat 1:129–167
Barnes LG (1985) Fossil pontoporiid dolphins (Mammalia: Cetacea) from the Pacific coast of North America. Contrib Sci 363:1–34
Barnes LG (2006) A phylogenetic analysis of the superfamily Platanistoidea (Mammalia, Cetacea, Odontoceti). Beitr Palaeontol 30:25–42
Bengtson P (1988) Open nomenclature. Palaeontology 31:223–227
Best RC, da Silva VMF (1993) Inia geoffrensis. Mammal Species 1–8
Bianucci G (2005) Arimidelphis sorbinii a new killer whale-like dolphin from the Pliocene of the Marecchia River (central eastern Italy) and a phyogenetic analysis of the Orcininae (Cetacea: Odonticeti). Riv Ital Paleontol S 111:329–344
Bianucci G, Di Celma C, Landini W, Post K, Tinelli C, Muizon C de, Gariboldi K, Malinverno E, Cantalamessa G, Gioncada A, Collareta A, Gismondi R-S, Varas-Malca R, Urbina M, Lambert O (2016) Distribution of fossil marine vertebrates in Cerro Colorado, the type locality of the giant raptorial sperm whale Livyatan melvillei (Miocene, Pisco Formation, Peru). J Maps 12:543–557. doi: 10.1080/17445647.2015.1048315
Bianucci G, Lambert O, Post K (2010) High concentration of long-snouted beaked whales (genus Messapicetus) from the Miocene of Peru. Palaeontology 53:1077–1098. doi: 10.1111/j.1475-4983.2010.00995.x
Bianucci G, Lambert O, Salas-Gismondi R, Tejada J, Pujos F, Urbina M, Antoine P-O (2013) A Miocene relative of the Ganges River dolphin (Odontoceti, Platanistidae) from the Amazonian Basin. J Vertebr Paleontol 33:741–745. doi: 10.1080/02724634.2013.734888
Bianucci G, Landini W (2002) Change in diversity, ecological significance and biogeographical relationships of the Mediterranean Miocene toothed whale fauna. Geobios 24:19–28
Bianucci G, Landini W, Valleri G, Ragaini L, Varola A (2005) First cetacean fossil records from Ecuador, collected from the Miocene of Esmeraldas Province. Riv Ital Paleontol S 111:345–350
Bianucci G, Urbina M, Lambert O (2015) A new record of Notocetus vanbenedeni (Squalodelphinidae, Odontoceti, Cetacea) from the early Miocene of Peru. C R Palevol 14:5–13. doi: 10.1016/j.crpv.2014.08.003
Buchholtz EA (2001) Vertebral osteology and swimming style in living and fossil whales (Order: Cetacea). J Zool 253:175–190
Buchholtz EA, Schur SA (2004) Vertebral osteology in Delphinidae (Cetacea). Zool J Linn Soc 140:383–401
Buono MR, Cozzuol MA (2013) A new beaked whale (Cetacea, Odontoceti) from the late Miocene of Patagonia, Argentina. J Vertebr Paleontol 33:986–997. doi: 10.1080/02724634.2013.752377
Buono MR, Viglino M, Cozzuol MA, Lucero S (2016) Descifrando la historia evolutiva de los Neoceti (Mammalia: Cetacea): aportes del registro fósil del Neógeno del Atlántico Sudoccidental. In: Agnolin FL, Lio GL, Brissón Egli F, Chimento NR, Novas FE (ed) Historia Evolutiva y Paleobiogeográfica de los Vertebrados de América del Sur, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia,” Buenos Aires, pp 323–334
Cabrera A (1926) Cetáceos fósiles del Museo de la Plata. Rev Mus La Plata 24:363–411
Carrillo JD, Forasiepi A, Jaramillo C, Sánchez-Villagra MR (2015) Neotropical mammal diversity and the Great American Biotic Interchange: spatial and temporal variation in South America’s fossil record. Front Genet 5. doi: 10.3389/fgene.2014.00451
Carrillo-Briceño JD, Aguilera OA, De Gracia C, Aguirre-Fernández G, Kindlimann R, Sánchez-Villagra MR (2016) An Early Neogene Elasmobranch fauna from the southern Caribbean (western Venezuela). Palaeontol Electron 19.2.28A
Carrillo-Briceño JD, Maxwell E, Aguilera OA, Sánchez R, Sánchez-Villagra MR (2015) Sawfishes and other elasmobranch assemblages from the Mio-Pliocene of the south Caribbean (Urumaco sequence, northwestern Venezuela). PLoS ONE 10:e0139230. doi: 10.1371/journal.pone.0139230
Cope ED (1868) Second contribution to the history of the Vertebrata of the Miocene period of the United States. Proc Acad Nat Sci Philadelphia 20:184–194. doi: 10.2307/4059884
Cope ED (1869) Third contribution to the fauna of the Miocene period of the United States. Proc Acad Nat Sci Philadelphia 21:6–12. doi: 10.2307/4060091
Cozzuol MA (1988) Una nueva especie de Saurodelphis Burmeister, 1891 (Cetacea: Iniidae) del “Mesopotamiense” (Mioceno tardío-Plioceno temprano) de la provincia de Entre Ríos, Argentina. Ameghiniana 25:39–45
Cozzuol MA (1996) The record of the aquatic mammals in southern South America. Münchner Geowiss Abh 30:321–342
Cozzuol MA (2006) The Acre vertebrate fauna: age, diversity, and geography. J South Am Earth Sci 21:185–203. doi: 10.1016/j.jsames.2006.03.005
Cozzuol MA (2010) Fossil record and the evolutionary history of Inioidea. In: Ruiz-García M, Shostell JM (eds) Biology, Evolution and Conservation of River Dolphins within South America and Asia. Nova Science Publishers, New York, pp 193–217
Cozzuol MA, Aguilera OA (2008) Cetacean remains from the Neogene of northwestern Venezuela. Paläontol Z 82:196–203. 10.1007/BF02988409
Díaz de Gamero ML (1974) Microfauna y edad de la Formación Cantaure, Península de Paraguaná, Venezuela. Bol Inf Asoc Venez Geol Min Petr 17:41–47
Díaz de Gamero ML (1989) El Mioceno temprano y medio de Falcón septentrional. GEOS 29:25–35
Doyle P, Mather AE, Bennett MR, Bussell MA (1996) Miocene barnacle assemblages from southern Spain and their palaeoenvironmental significance. Lethaia 29:267–274. doi: 10.1111/j.1502-3931.1996.tb01659.x
Fordyce RE (1994) Waipatia maerewhenua, new genus and new species (Waipatiidae, new family), an archaic late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand. In: Berta A, Deméré T (eds) Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore, Jr. Proc San Diego Soc of Nat Hist 29, pp 147–176
Fordyce RE (2009) Fossil sites, noted. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of Marine Mammals, 2nd ed. Academic Press, San Diego, pp 459–465
Fordyce RE, Muizon C de (2001) Evolutionary history of cetaceans: a review. In: Mazin J-M, Buffrenil V de (eds) Secondary Adaptation of Tetrapods to Life in Water. Proceedings of the International Meeting in Poitiers, 1996, Pfeil, Munich, pp 169–233
Geisler JH, Godfrey SJ, Lambert O (2012) A new genus and species of late Miocene inioid (Cetacea, Odontoceti) from the Meherrin River, North Carolina, U.S.A. J Vertebr Paleontol 32:198–211. doi: 10.1080/02724634.2012.629016
Geisler JH, McGowen MR, Yang G, Gatesy J (2011) A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evol Biol 11:1–33
Gibson-Smith W (1979) The genus Arcinella (Mollusca: Bivalvia) in Venezuela and some associated faunas. GEOS 24:11–32
Gibson-Smith J, Gibson-Smith W (1983) Neogene melogenid gastropods from the Paraguaná Peninsula, Venezuela. Eclogae geol Helv 76:719–728
Griffiths N, Müller W, Johnson KG, Aguilera OA (2013) Evaluation of the effect of diagenetic cements on element/Ca ratios in aragonitic early Miocene (~16 Ma) Caribbean corals: Implications for ‘deep-time’ palaeo-environmental reconstructions. Palaeogeogr Palaeoclimatol Palaeoecol 369:185–200. doi: 10.1016/j.palaeo.2012.10.018
Gutstein CS, Cozzuol MA, Vargas AO, Suárez ME, Schultz CL, Rubilar-Rogers D (2009) Patterns of skull variation of Brachydelphis (Cetacea, Odontoceti) from the Neogene of the southeastern Pacific. J Mammal 90:504–519
Hambalek N, Rull V, Digiacomo E de, Díaz de Gamero M (1994) Evolución paleoecológica y paleoambiental de la secuencia del Neógeno en el surco de Urumaco. Estudio palinológico y litológico. Bol Soc Venezolana Geol 191:7–19
Hendy AJW, Jones DS, Moreno F, Zapata V, Jaramillo C (2015) Neogene molluscs, shallow marine paleoenvironments, and chronostratigraphy of the Guajira Peninsula, Colombia. Swiss J Palaeontol 134:45–75. doi: 10.1007/s13358-015-0074-1
Hodson F (1926) Venezuelan and Caribbean turritellas, with a list of type stratigraphic localities. Bull Am Paleontol 11:173–220
Hunter VF, Bartok P (1974) The age and correlation of the Tertiary sediments of the Paraguaná Península, Venezuela. Bol Inf Asoc Venezolana Geol Min Petr 17:143–154
Jaramillo C, Moreno F, Hendy AJW, Sánchez-Villagra MR, Marty D (2015) Preface: La Guajira, Colombia: a new window into the Cenozoic neotropical biodiversity and the Great American Biotic Interchange. Swiss J Palaeontol 134:1–4. doi: 10.1007/s13358-015-0075-0
Jung P (1965) Miocene Mollusca from the Paraguaná Peninsula, Venezuela. Bull Am Paleontol 49:389–607
Kellogg AR (1924) A fossil porpoise from the Calvert Formation of Maryland. Proc US Natl Mus 63:1–39
Kellogg AR (1927) Kentriodon pernix, a Miocene porpoise from Maryland. Proc US Natl Mus 69:1–55
Lambert O (2005) Phylogenetic affinities of the long-snouted dolphin Eurhinodelphis (Cetacea, Odontoceti) from the Miocene of Antwerp, Belgium. Palaeontology 48:653–679
Lambert O, Estevens M, Smith R (2005) A new kentriodontine dolphin from the middle Miocene of Portugal. Acta Palaeontol Pol 50:239–248
Lambert O, Muizon C de, Bianucci G (2015) A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru. Geodiversitas 37:79–108. doi: 10.5252/g2015n1a4
Leduc RG, Perrin WF, Dizon AE (1999) Phylogenetic relationships among the delphinid cetaceans based on full cytochrome b sequences. Mar Mammal Sci 15:619–648
Linares OJ (2004) Bioestratigrafía de la fauna de mamíferos de las formaciones Socorro, Urumaco y Codore (Mioceno medio-Plioceno temprano) de la región de Urumaco, Falcón, Venezuela. Paleobiol Neotrop 1:1–26
Lydekker R (1894) Contributions to the knowledge of the fossil vertebrates of Argentina. Part II. Cetacean skulls from Patagonia. An Mus La Plata 2:1–15
Marx FG, Fordyce RE (2015) Baleen boom and bust: a synthesis of mysticete phylogeny, diversity and disparity. R Soc Open Sci 2. doi: 10.1098/rsos.140434
Marx FG, Tsai C-H, Fordyce RE (2015) A new early Oligocene toothed ‘baleen’ whale (Mysticeti: Aetiocetidae) from western North America: one of the oldest and the smallest. R Soc Open Sci 2:10.1098/rsos.150476
Mead JG, Fordyce RE (2009) The therian skull: a lexicon with emphasis on the odontocetes. Smithson Contrib Zool 627:1–248
Mones A (1986) Palaeovertebrata Sudamericana. Catálogo sistemático de los vertebrados fósiles de America del Sur, Parte I. Lista preliminar y bibliografía. Courier Forschungsinstitut Senckenberg 82:1–625
Moreno FP (1892) Noticias sobre algunos cetáceos fósiles y actuales de la República Argentina. Rev Mus La Plata 3:381–400
Moreno F, Hendy AJW, Quiroz L, Hoyos N, Jones DS, Zapata V, Zapata S, Ballen GA, Cadena E, Cárdenas AL, Carrillo-Briceño JD, Carrillo JD, Delgado-Sierra D, Escobar J, Martínez JI, Martínez C, Montes C, Moreno J, Pérez N, Sánchez R, Suárez C, Vallejo-Pareja MC, Jaramillo C (2015) Revised stratigraphy of Neogene strata in the Cocinetas Basin, La Guajira, Colombia. Swiss J Palaeontol 134:5–43. doi: 10.1007/s13358-015-0071-4
Muizon C de (1984) Les vertébrés fossiles de la Formation Pisco (Pérou). Deuxième partie: les odontocètes (Cetacea, Mammalia) du Pliocène inférieur de Sud-Sacaco. Trav Inst Fr E And 50:1–188
Muizon C de (1988a) Les vertébrés fossiles de la Formation Pisco (Pérou). Troisème partie: Les odontocètes (Cetacea, Mammalia) du Miocène. Mem Inst Fr Et And 78:1–244
Muizon C de (1988b) Le polyphylétisme des Acrodelphidae, odontocètes longisrostres du Miocène européen. Bull Mus Natl Hist Nat 10:31–88
Muizon C de (1988c) Les relations phylogénétiques des Delphinida (Cetacea, Mammalia). Ann Paleontol 74:159–227
Nummela S, Thewissen JGM, Bajpai S, Hussain T, Kumar K (2007) Sound transmission in archaic and modern whales: anatomical adaptations for underwater hearing. Anat Rec 290:716–733. doi: 10.1002/ar.20528
O’Leary MA (2004) A fragmentary odontocete cranium from the lower Miocene of Venezuela. Spec Pap Palaeontol 71:99–104
Ochsenius C (1980) Cuaternario en Venezuela. UNEFM, Coro, Venezuela
Odreman-Rivas OE (1997) Lista actualizada de los fósiles de Venezuela. In: La Marca E (ed) Vertebrados actuales y fósiles de Venezuela. Museo de ciencia y tecnología de Mérida, Mérida, Venezuela, pp 231–234
Odreman-Rivas OE, Medina CJ (1984) Vertebrados fósiles de Venezuela secuencia, relaciones con otros países de América del Sur. Cuad Geol 1:60–86
Omura H (1972) An osteological study of the Cuvier’s beaked whale, Ziphius cavirostris, in the Northwest Pacific. Sci Rep Whales Res Inst 24:1–34
Pérez LM, Pérez Panera JP, Aguilera OA, Ronchi DI, Sánchez R, Manceñido MO, Sánchez-Villagra MR (2016) Palaeontology, sedimentology, and biostratigraphy of a fossiliferous outcrop of the early Miocene Querales Formation, Falcón Basin, Venezuela. Swiss J Palaeontol 135:187–203. doi: 10.1007/s13358-015-0105-y
Perrin WF (1975) Variation of spotted and spinner porpoise (genus Stenella) in the Eastern Pacific and Hawaii. Bull Scripps Inst Oceanogr 21:1–206
Pilleri G, Gihr M (1979) Skull, sonar field and swimming behaviour or Ischyorhynchus vanbenedeni (Ameghino, 1891) and taxonomical position of the genera Ischyorhynchus, Anisodelphis and Pontoplanodes (Cetacea). Investig Cetacea 10:17–70
Pyenson ND, Gutstein CS, Parham JF, Le Roux JP, Chavarría CC, Little H, Metallo A, Rossi V, Valenzuela-Toro AM, Velez-Juarbe J, Santelli CM, Rogers DR, Cozzuol MA, Suárez ME (2014) Repeated mass strandings of Miocene marine mammals from Atacama Region of Chile point to sudden death at sea. Proc R Soc Biol Sci Ser B 281:doi: 10.1098/rspb.2013.3316
Pyenson ND, Vélez-Juarbe J, Gutstein CS, Little H, Vigil D, O’Dea A (2015) Isthminia panamensis, a new fossil inioid (Mammalia, Cetacea) from the Chagres Formation of Panama and the evolution of ‘river dolphins’ in the Americas. PeerJ 3:e1227. doi: 10.7717/peerj.1227
Quiroz L, Jaramillo C (2010) Stratigraphy and sedimentary environments of Miocene shallow to marginal marine deposits in the Urumaco Through, Falcón Basin, western Venezuela. In: Sánchez-Villagra MR, Aguilera O, Carlini AA (eds) Urumaco and Venezuelan Paleontology: The Fossil Record of the Northern Neotropics. Indiana University Press, Bloomington, pp 153–172
Rey O (1994) Miembro Chiguaje: Formación La Vela, o Formación Codore. Boletín de la Sociedad Venezolana de Geólogos 19: 50–53
Rey O (1996) Estratigrafía de la península de Paraguaná, Venezuela. Rev Fac Ing 11:36–45
Rommel SA (1990) Osteology of the bottlenose dolphin. In: Leatherwood S, Reeves RR (eds) The Bottlenose Dolphin. Academic Press, San Diego, pp 29–49
Rommel SA, Reynolds JE, III (2008) Skeleton, postcranial. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of Marine Mammals. Academic Press, San Diego, pp 1021–1032
Sánchez‐Villagra MR, Aguilera OA (2006) Neogene vertebrates from Urumaco, Falcón State, Venezuela: diversity and significance. J Syst Palaeontol 4:213–220. doi: 10.1017/S1477201906001829
Sánchez-Villagra MR, Aguilera OA, Sánchez R, Carlini AA (2010) The fossil vertebrate record of Venezuela of the last 65 million years. In: Sánchez-Villagra MR, Aguilera O, Carlini AA (eds) Urumaco and Venezuelan Paleontology: The Fossil Record of the Northern Neotropics. Indiana University Press, Bloomington, pp 19–51
Sánchez-Villagra MR, Burnham RJ, Campbell DC, Feldmann RM, Gaffney ES, Kay RF, Lozsán R, Purdy R, Thewissen JGM (2000) A new near-shore marine fauna and flora from the early Neogene of northwestern Venezuela. J Paleontol 74:957–968
Sánchez-Villagra MR, Gasparini Z, Lozsán R, Moody JM, Uhen MD (2001) New discoveries of vertebrates from a near-shore marine fauna from the early Miocene of northwestern Venezuela. Paläontol Z 75:227–232. doi: 10.1007/BF02988016
Scheyer TM, Aguilera OA, Delfino M, Fortier DC, Carlini AA, Sánchez R, Carrillo-Briceño JD, Quiroz L, Sánchez-Villagra MR (2013) Crocodylian diversity peak and extinction in the late Cenozoic of the northern Neotropics. Nat Commun 4:1907. doi: 10.1038/ncomms2940
Slijper EJ (1936) Die Cetaceen. Vergleischend-Anatomisch und Systematisch. Ein Beitrag zur vergleichenden Anatomie des Blutgefäss-, Nerven- und Muskelsystems, sowie des Rumpfskelettes der Säugetiere, mit Studien über die Theorie des Aussterbens und der Foetalisation. A. Asher & Co. B. V., Amsterdam
Smith JB, Dodson P (2003) A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. J Vertebr Paleontol 23:1–12
Smith CJ, Collins LS, Jaramillo C, Quiroz LI (2010) Marine paleonvironments of Miocene–Pliocene formations of north-central Falcón State, Venezuela. J Foraminifer Res 40:266–282. doi: 10.2113/gsjfr.40.3.266
True FW (1910) Description of a skull and some vertebrae of the fossil cetacean Diochotichus vanbenedeni from Santa Cruz, Patagonia. Bull Am Mus Nat Hist 28: 19–32
Uhen MD (2010) The origin(s) of whales. Annu Rev Earth Planet Sci 38:189–219. doi: 10.1146/annurev-earth-040809-152453
Uhen MD, Coates AG, Jaramillo CA, Montes C, Pimiento C, Rincon A, Strong N, Velez-Juarbe J (2010) Marine mammals from the Miocene of Panama. J South Am Earth Sci 30:167–175. doi: 10.1016/j.jsames.2010.08.002
Valerio AL, Laurito CA (2012) Cetáceos fósiles (Mammalia, Odontoceti, Eurhinodelphinoidea, Inioidea, Physeterioidea) de la Formación Curré, Mioceno superior (Hemphilliano temprano tardío) de Costa Rica. Rev Geol Am Cent 46:151–160
Vélez-Juarbe J, Wood AR, De Gracia C, Hendy AJW (2015) Evolutionary patterns among living and fossil kogiid sperm whales: evidence from the Neogene of Central America. PLoS ONE 10:e0123909. doi: 10.1371/journal.pone.0123909
Acknowledgements
This work greatly benefited from the suggestions of the reviewers (Giovanni Bianucci and Olivier Lambert) and the editors (François Pujos, Pierre-Olivier Antoine, and John Wible). We thank Carlos Jaramillo (STRI), Analía Forasiepi (CCT-CONICET-Mendoza), and Yoshihiro Tanaka (Numata Fossil Museum) for reading through and commenting on earlier versions of this manuscript. Juan David Carrillo (University of Zurich), Mary Parra and Fredy Parra (CIP) are thanked for fossil preparation. Loïc Costeur (NMB), Mary Parra (STRI), and Jaime Escobar (Universidad del Norte) kindly provided access to specimens under their care. Thanks to Andrés Reyes Cespedes (Universidad Experimental Francisco de Miranda) for help in collections. Thanks to Michael Knappertbusch (NMB) for guidance with micropaleontological analysis and access to laboratory and equipment. Austin Hendy (LACM) and Gustavo Ballen (STRI) kindly provided contextual information regarding the fieldwork in which the MUN-STRI specimens were collected. Juan David Carrillo, Ashley Latimer, and Torsten Scheyer (University of Zurich) are thanked for useful discussions; Juan David Carrillo provided measurements of AMU-CURS-219. GAF wishes to thank Jairo Carrillo (Colombia)and his family for their hospitality and logistical aid. Thanks to Scott Hartman for producing the silhouette used in Fig. 1 and PhyloPic for making it available.
We thank the Instituto del Patrimonio Cultural de Venezuela and the Alcaldía Bolivariana de Urumaco, for the authorization and permissions to collect and study the fossil material from Venezuela. The Smithsonian Institution, the National Geographic Society, Universidad del Norte, the Anders Foundation, Gregory D. and Jennifer Walston Johnson, and the National Science Foundation (grant EAR 0957679) helped to support fieldwork. Thanks to Carlos Rosero for managing all the logistics in the field in Colombia, Liliana Londoño and Maria Ines Barreto for administrative and logistic support. Thanks to the Wayuú Community for allowing access to their lands and for support during fieldwork, the Colombian National Police (Castilletes base), and all the members of the field teams over the past four years.
This project was funded by Swiss National Science Foundation grant 31003A_149605 to MRS-V. GAF was supported by Fonds für Lehre und Forschung from the Naturhistorisches Museum Basel. EA was supported by a Smithsonian Tropical Research Institute postdoctoral fellowship grant.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary data
The Online Resource 1 includes unlabeled specimen photographs. Additional field data linked to MUN-STRI specimens can be downloaded from the Smithsonian Tropical Research Institute Geological Sample Database webpage (http://biogeodb.stri.si.edu/jaramillo/fossildb/index.php/) using either the locality (e.g., “490006”) or the sample (e.g., “37698”) numbers. (PDF 1656 kb)
Rights and permissions
About this article

Cite this article
Aguirre-Fernández, G., Carrillo-Briceño, J.D., Sánchez, R. et al. Fossil Cetaceans (Mammalia, Cetacea) from the Neogene of Colombia and Venezuela. J Mammal Evol 24, 71–90 (2017). https://doi.org/10.1007/s10914-016-9353-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10914-016-9353-x