
Abstract
The European Miocene records a wide diversity of hyaenid ecomorphotypes represented by multiple genera. Among these, Hyaenictis Gaudry, 1861, is one of the least known. This genus includes four species from the late Miocene and Pliocene of the Old World, but in Europe Hyaenictis is only represented by two species, recorded by scarce and fragmentary remains: Hyaenictis graeca Gaudry, 1861, from Pikermi (MN12; Greece) and Hyaenictis almerai Villalta Comella and Crusafont Pairó, 1948, from Sant Miquel de Toudell (MN10; Vallès-Penedès Basin, NE Iberia). Here, we describe a new skull of Hyaenictis aff. almerai from the Vallès-Penedès site of Ronda Oest Sabadell Sector D (MN10), representing the most complete European specimen of the genus. In the presence of m2 and virtual lack of m1 metaconid, the described cranium more closely resembles Hyaenictis rather than any other medium- to large-sized European hyaenid. However, the new skull does not fit well with previously known Hyaenictis species, more closely resembling the bone-cracking Adcrocuta Kretzoi, 1938, in the development of premolar accessory cuspids and the possession of relatively broad cheek teeth. These and other features (strong mandibular muscular insertions and enamel microstructure) denote more durophagous adaptations than previously documented in Hyaenictis (considered a cursorial/dog-like hyaena), and favor the inclusion of H. aff. almerai in the transitional bone-cracking hyaenid ecomorphotype.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Genus Hyaenictis
Hyaenictis Gaudry, 1861, and its type species, Hyaenictis graeca Gaudry, 1861, were erected on the basis of scarce dentognathic remains from the Greek locality of Pikermi (MN12, middle Turolian, late Miocene). The holotype is a juvenile hemimandible with dc, p1 alveolus, dp2–dp4, erupting m1 and m2, and exposed p2-p4 inside their crypts. A few additional dentognathic remains, including two juvenile maxillary fragments, have been further attributed to the same taxon (see also Howell and Petter 1985; Werdelin et al. 1994; Koufos 2011). This species is characterized by a mixture of primitive features (retention of p1 and m2) and several derived traits (such as vestigial m1 metaconid; Gaudry 1861; Howell and Petter 1985; Werdelin et al. 1994). An adult partial hemimandible of this genus was found in 1920 in the Iberian locality of Sant Miquel de Toudell (SMT; formerly spelled ‘Taudell’) in the area of Viladecavalls (Vallès-Penedès Basin; MN10, late Vallesian, late Miocene). This specimen was initially attributed to H. graeca by Bataller (1921; see also Villalta Comella and Crusafont Pairó 1941, 1943), but later designated as the holotype of another species of the genus, Hyaenictis almerai Villalta Comella and Crusafont Pairó, 1948. According to Villalta Comella and Crusafont Pairó (1948), H. almerai resembles H. graeca in the retention of m2 and the lack of m1 metaconid, but differs from it in several other features (shorter and stouter mandibular corpus, shorter diastema, and apparent lack of p1).
Solounias (1981) synonymized Adcrocuta Kretzoi, 1938, with Hyaenictis based on similarities in the m1 and the lack of additional remains of H. graeca among the vast hyaenid sample from Pikermi. However, this taxonomic opinion was discounted by subsequent authors (Howell and Petter 1985; Werdelin and Solounias 1990, 1991). Werdelin and Solounias (1991) noted that only a few similarities justified the attribution of the hyaenid from Sant Miquel de Toudell to Hyaenictis, but tentatively retained this generic assignment, in spite of recognizing that the erection of a new genus might eventually be justified. Later, Hyaenictis hendeyi Werdelin et al. 1994, was erected based on early Pliocene (ca. 5 Ma) material from Langebaanweg in South Africa, previously attributed to indeterminate species of Euryboas Schaub, 1941 (Hendey 1976, 1978). Some additional middle Miocene to early Pliocene hyaenid remains from elsewhere in Africa have been subsequently attributed to Hyaenictis. These specimens are mostly isolated teeth (Ginsburg 1977; Turner 1988; Morales et al. 2005; Werdelin and Peigné 2010). The exception is Hyaenictis wehaietu Haile-Selassie and Howell, 2009, from the late Miocene and early Pliocene of the Middle Awash in Ethiopia (5.8–5.2 Ma), which is represented by more abundant material (Haile-Selassie and Howell 2009). Besides its smaller size, this species differs from H. graeca by the lack of p1 and m2, and the presence of a well-developed m1 metaconid (Haile-Selassie and Howell 2009). Given the lack of these diagnostic features of the type species of the genus, the inclusion of this species in Hyaenictis is questionable (being here referred to as ?Hyaenictis wehaietu).
Hyaenictis has been classically included in the cursorial hyaenid ecomorphotype (Werdelin and Solounias 1996; Turner et al. 2008), given the lack of the typical durophagous adaptations of Adcrocuta. In this paper, we describe a new skull with associated mandible and atlas from the late Vallesian of the Vallès-Penedès Basin that, based on the retention of p1 and m2, as well as the reduced m1 metaconid, is attributed to Hyaenictis. This skull, which represents the most complete specimen of Hyaenictis known so far from the European Miocene, does not fit well with previously known species of this genus, from which it differs in several morphological and metric features that more closely resemble those of the bone-cracking hyaena Adcrocuta. The taxonomic assignment of the described specimen is discussed here, together with its implications for the evolution of durophagous adaptations in hyaenids during the late Miocene.
Age and Geological Background
The described material comes from sector ROS-D within the set of paleontological sites of Ronda Oest Sabadell (ROS), which were discovered between 2009 and 2011 during paleontological surveillance of the construction of a road near the city of Sabadell (Catalonia, Spain; Madurell-Malapeira et al. 2014). From a geological viewpoint, ROS is located within the Vallès-Penedès Basin (Fig. 1), a small and elongated half-graben oriented parallel to the Mediterranean coastline in the NE Iberian Peninsula and bounded by the Catalan (Prelittoral and Littoral) Coastal Ranges (Cabrera et al. 2004; de Gibert and Casanovas-Vilar 2011). This basin has been intensively sampled for many decades, resulting in a particularly well-known late Aragonian and Vallesian record of terrestrial mammals (Casanovas-Vilar et al. 2011, 2015). Based on biostratigraphic data, ROS localities cover a time span ranging from the late Vallesian (MN10) to the early Turolian (MN11; Madurell-Malapeira et al. 2014). The remains described here were found stratigraphically 1.5 m below locality ROS-D6, which on the basis of micromammals is correlated to the upper part of MN10 (Madurell-Malapeira et al. 2014; Casanovas-Vilar et al. 2015). More precisely, ROS-D6 is tentatively correlated to the Rotundomys bressanus local subzone of the Vallès-Penedès Basin (Casanovas-Vilar et al. 2015), with an estimated age of ca. 9.4–9.1 Ma, although alternatively it might, like SMT, belong to the older Rotundomys montisrotundi local subzone, which has an estimated age of 9.6–9.4 Ma (Casanovas-Vilar et al. 2015).

Detailed geological map of the western (Vallès) sector of the Vallès-Penedès Basin (within the Iberian Peninsula), showing the position of the referred late Vallesian mammal sites (modified after Casanovas-Vilar et al. 2015). Locality acronyms: ROS-D, Ronda Oest Sabadell Sector D; SMT, Sant Miquel de Toudell
Materials and Methods
Studied Material and Comparative Sample
The hyaenid remains described in this paper are housed in the ICP (see institutional abbreviations below). Anatomical terminology used in the descriptions follows Evans and de Lahunta (2013). The examined fossil comparative sample includes the holotype (MGB16051) of Hyaenictis almerai from SMT (MN10), housed in the MCNB; remains of Adrocuta eximia Roth and Wagner, 1854, from Torrentet de Traginers (Piera, MN12), housed in the ICP; specimens of Chasmaporthetes lunensis (Del Campana, 1914) from La Puebla de Valverde (Teruel, MN17), housed in the MNCN; and topotypic specimens of Hyaenictis hendeyi housed in SAM. Additional data from other Miocene and Pliocene hyaenids included in the comparative sample were taken from the literature (Gaudry 1861; Villalta Comella and Crusafont Pairó 1948; Hendey 1978; Howell and Petter 1985; Qiu 1987; Kurtén and Werdelin 1988; Werdelin 1988, 1999; Koufos 2011; Tseng et al. 2013). They include: Adcrocuta (MN10–MN13, Eurasia), including Adcrocuta eximia; Lycyaena Hensel, 1863 (MN9–MN12, Eurasia and North Africa), including Lycyaena chaeretis (Gaudry, 1861), Lycyaena dubia Zdansky, 1924, and Lycyaena crusafonti Kurtén, 1976; Chasmaporthetes Hay, 1921 (MN12–early Pleistocene, Eurasia, Africa, and North America), including Chasmaporthetes gangsriensis Tseng et al., 2013, Chasmaporthetes lunensis, Chasmaporthetes ossifragus Hay, 1921, Chasmaporthetes borissiaki (Khomenko, 1931), Chasmaporthetes australis (Hendey, 1974), and Chasmaporthetes nitidula (Ewer, 1955); Lycyaenops Kretzoi, 1938 (MN14–early Pleistocene, Europe and Africa), including Lycyaenops silberbergi (Broom in Broom and Schepers, 1946) and Lycyaenops rhomboideae Kretzoi, 1938; and Hyaenictis (MN10–MN14, Europe and Africa), including H. graeca, H. almerai, H. hendeyi, and ?H. wehaietu. The material of “Chasmaporthetes” bonisi Koufos, 1987, has been reassigned to A. eximia following Werdelin and Solounias (1991). The genera included in the comparative sample have been selected based on their age, geographic distribution, and morphological similarities with the ROS-D specimen.
Tooth Enamel Microstructure
Enamel microstructure (Hunter-Shreger Bands, HSB; Stefen 1997; Ferretti 1999, 2007) in the cheek teeth of the described material was described and photographed with a Leica MZ-16 light stereomicroscope with a Leica IC 3D digital camera attached. The resulting images were modified with the HDR (high dynamic range) toning function in Adobe Photoshop CS6 to enhance the contrast of the HSB.
Morphometric Comparisons
Craniomandibular and dental measurements of the studied specimens were taken with digital calipers to the nearest 0.1 mm (see measurement abbreviations below). Dental size and proportions were assessed using bivariate plots of buccolingual breadth (W) vs. mesiodistal length (L) for the lower cheek teeth (p2, p3, p4, and m1). A breadth/length index (BLI) for these teeth was computed as BLI = log (B/L), to assess differences in dental shape among the studied genera. These differences were inspected by means of boxplots and analysis of variance (ANOVA), including pairwise post-hoc comparisons (Bonferroni method), using IBM SPSS Statistics v. 19. A principal component analysis (PCA) based on the BLI of individuals preserving the p2–m1 series was performed with PAST v. 2.15 (Hammer et al. 2001). Note that the PCA (like ANOVA) was run on log-transformed data, so as to mini-mized the statistical problems associated with the usually non-normal distribution of ratios (although PCA makes no assumptions in this regard, it usually yields better results with normally distributed data; Hammer and Harper 2006). No statistical analyses were undertaken on the basis of the upper dentition, because of the scarcity of sufficiently complete cranial remains for most of the taxa included in the comparative sample (including Hyaenictis spp.). ?Hyaenictis wehaietu was not included in the morphometric comparisons (boxplots, ANOVA, and PCA), because as noted above this species is unlikely to belong to the genus Hyaenictis.
Abbreviations
Institutional Abbreviations
ICP: Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona (Spain); IPS: acronym of the collections of the ICP (formerly Institut de Paleontologia de Sabadell). MCNB, Museu de Ciències Naturals de Barcelona (Spain); MGB: acronym of the geology collections of the MCNB (including the former Museu de Geologia de Barcelona); MNCN: Museo Nacional de Ciencias Naturales–CSIC, Madrid (Spain); MNHN: Muséum National d’Histoire Naturelle, Paris (France); SAM: Iziko South African Museum, Cape Town (South Africa).
Locality Abbreviations
PIK: Pikermi; PQ-L: Pelletal Phosphorite and Quartzose Sand Members undifferentiated, Varswater Formation, “E” Quarry, Langebaanweg; ROS: Ronda Oest Sabadell; SMT: San Miquel de Toudell.
Measurement Abbreviations
B: buccolingual breadth; BLI: breadth/length index; FA: frontal-acrocranion length; LP1–P4: length of the upper premolar series; PA: prosthion-acrocranion length; Hoc: occipital height; PB: prosthion-basion length; L: mesiodistal length; LP4m: mesiodistal length of P4 metastyle; LP4pa: mesiodistal length of fourth upper premolar paracone; LP4pas: mesiodistal length of fourth upper premolar parastyle; Ltm1: mesiodistal length of the m1 talonid; PF: prosthion-frontal length; PN: prosthion-nasion length; PS, prosthion-synsphenion length; PST: prosthion-staphylion length; SB: synsphenion-basion length; SKL: prosthion-occipital condyle length; WC: palatal width at C1; WiP4: palatal width at P4; Woc: width of occipital condyles.
Systematic Paleontology
Order CARNIVORA Bowdich, 1821
Suborder FELIFORMIA Kretzoi, 1945
Family HYAENIDAE Gray, 1821
Subfamily HYAENINAE Gray, 1821
Genus HYAENICTIS Gaudry, 1861
Type species: Hyaenictis graeca Gaudry, 1861.
Other included species: Hyaenictis almerai Villalta Comella and Crusafont Pairó, 1948; Hyaenictis hendeyi Werdelin et al., 1994; ?Hyaenictis wehaietu Haile-Selassie and Howell, 2009.
Hyaenictis aff. almerai Footnote 1 Villalta Comella and Crusafont Pairó, 1948

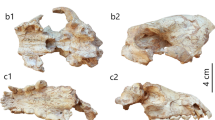
Hyaenictis aff. almerai IPS62078 from ROS-D. a–c, cranium, in ventral (a), left lateral (b) and dorsal (c) views. d–f, right P4, in occlusal (d), labial (e) and lingual (f) views. g–h, atlas, in dorsal (g) and ventral (h) views. i–k, left hemimandible, in labial (i), lingual (j) and occlusal (k) views. l–n, right hemimandible, in labial (l), lingual (m) and occlusal (n) views. o–q, right m2 in occlusal (o), lingual (p) and labial (q) views. r–t, right M1 in occlusal (r), distal (s) and mesial (t) views. o–t are depicted at a different scale

Photographs of the HSB pattern on the outer enamel surface of the bottom crown portion of P3 (a) and the middle crown portion of p3 (b) in Hyaenictis aff. almerai IPS62078 from ROS-D. Apical is towards the top of the image. Note that the zig-zag HSB pattern of the middle crown portion progressively transforms into an acute-angled undulating HSB pattern (Tseng 2012) closer to the crown base of the P3 (a)

Schematic drawings of the right postcanine toothrow of the ROS-D specimen and three species of Hyaenictis. a–c, Hyaenictis aff. almerai (IPS62070), in labial (a), lingual (b) and occlusal (c) views. d–f, Hyaenictis hendeyi (based on PQ-L21792 and the holotype PQ-L21000), in labial (d), lingual (e) and occlusal (f) views. g–i, Hyaenictis almerai (holotype, MGB16051), in labial (g), lingual (h) and occlusal (i) views. j–k, Hyaenictis graeca (holotype, MNHN-PIK-3002), in lingual (j) and occlusal (k) views. In the occlusal views, crests and cingulids are represented by thin lines, cuspids are denoted by black dots, and discontinuous lines represent inferred morphology (covered, missing and worn areas)
Holotype: MGB1605, partial right hemimandible from SMT (MN10, Spain).
Original description: “Relatively evolved Hyaenictis, having lost the p1, diastema very reduced; relatively robust mandible; shortened face, and canine in a more vertical position. m1 without metaconid. p2 long, slightly pointed and with a developed anterior cusp” (Villalta Comella and Crusafont Pairó 1948: 117, our translation from the Spanish original).
Tentatively referred material: IPS62078 from ROS-D, including: a partial cranium with left and right C1-M1; associated left and right hemimandibles with canines, left p3–m1, and right p2–m1 and m2; and associated atlas. See measurements in Table 1.
Description
Cranium
The cranium preserves most of the palate, the snout and the left side of the neurocranium (including the zygomatic arch), as well as the occipital condyles and the pterygoids (Fig. 2a–c). Most of the basicranium (basisphenoid and basioccipital) and the right side of the braincase (sphenoid complex, caudal end of the frontal, temporal, and parietal) are missing (Fig. 2a–c). The nasals are triangular, long and narrow, extending posteriorly nearly to the level of the postorbital processes (Fig. 2c). The infraorbital foramen is oval and in lateral view is located at the level of P3 (Fig. 2b). The zygomaticomaxillary suture is just above the P4, running from the anterior end of the orbit towards the distal root of P4 (Fig. 2b). The premaxillary-maxillary suture runs from the I3 root towards the mid-point of the nasals without reaching the frontal (Fig. 2c). The nasal aperture is wide and oval. The palate is wide at the level of P4 and narrows mesialwards to the canines (Fig. 2a). There is a slight postcanine constriction (Fig. 2c). The transverse palatine suture reaches the distal end of M1, as can be observed on the left side of the palate despite the presence of multiple cracks on this area (Fig. 2a). The palatine fissures are present at the level of the canine and are oval in shape (Fig. 2a). On the basicranium, the retroarticular processes are transversely wide and robust (Fig. 2a).
Mandible
The two hemimandibles are almost complete, except for a small fragment of the right coronoid process and the distalmost part of the left angular process (Fig. 2i–n). The corpus is straight and slender, and has a single mental foramen below the p2 at about corpus midheight (Fig. 2i, l). The ramus is low, with a rounded coronoid process slightly expanded backwards and a condylar process that is located at about the same level as the occlusal plane (Fig. 2i, j, l, m). The masseteric fossa is deep and extends anteriorly until the level of m2 (Fig. 2i, l). The angular process is well developed and protrudes posteriorly beyond the condylar process (Fig. 2l, m).
Dentition
The dentition is quite worn, with dentine exposure at the tips of the main and most of the accessory cusps. The upper dentition is more complete on the left side, where the I3–M1 series is preserved, whereas on the right side the canine is broken, the P1 is missing, and the P4 and M1 are detached (Fig. 2a, d–f, r–t). The two I3s and the left canine are very worn. The P1 is buccolingually wider than mesiodistally long and displays a distal cingulum. The P2 has a blunt crest running from the protocone to the mesiolingual margin and a small distal accessory cusp just mesial to the distal cingulum. The P3 has two accessory cusps: the mesial one is rather small, located very close to the protocone and on the lingual half of the crown; the distal accessory cusp is broad but low, and located just mesial to the small cingulum present on the distal crown margin. The two P4s (Fig. 2a, d-f) are quite worn, so that only a few occlusal characters can be observed, including the presence of a well-developed protocone and of a parastyle that is mesiodistally shorter than the paracone. The M1 is present but similarly worn, and displays a subrectangular occlusal contour with no visible cusp features (Fig. 2a, r–t).
Most of the lower dentition is preserved, except for the incisors, the p1s, and the left p2 and m2 (Fig. 2i–q). The lower canines, like their upper counterparts, are quite worn. The diastema between the canine and p2 is moderately short and displays, closer to the p2, an alveolus for a single-rooted p1. All the remaining premolars are longer than wide but moderately stout, with a subrectangular (rather than oval) occlusal outline. The p2 has a small distal accessory cusp that is separated from the protoconid and from the distal cingulid by small notches. The p3 and p4 have better developed protoconids and two accessory cusps, the mesial one located towards the lingual side of the crown and the distal one situated on the marked distal cingulid (more buccally on p3 and more centrally on p4). The m1 paraconid and protoconid are too worn to observe their morphology, but it can be ascertained that the metaconid, located distolingually from the protoconid, is only slightly developed and not completely isolated from the former cusp. The m1 talonid is much lower and shorter than the trigonid, and displays three main cuspids, of which the hypoconid is larger than the entoconid and hypoconulid. The m2 (Fig. 2o-q) has a round occlusal outline, a single trigonid cuspid (the mesially located paraconid), and two smaller talonid cuspids (entoconid and hypoconid) that are more distally situated.
With regard to tooth enamel microstructure, HSB are observable in all the lower and upper cheek teeth (at least on the bottom crown portion, and also, when preserved, in the middle of the crown wall, as the apical parts of the crowns are in all instances worn away). These bands clearly show a zig-zag pattern (Fig. 3) like that in extant bone-cracking hyaenids (Ferretti 1999; Tseng 2011, 2012) on the middle crown portions and, to some extent, on their bottom portion (although closer to the cervix the HSB pattern progressively becomes acute-angled undulating).
Atlas
The atlas (Fig. 2g, h) is nearly complete, with only the wings broken. The vertebral arch is slender and medially displays a small craniodorsal protuberance. The lateral vertebral foramen and the transverse foramen are distinct (close to each other but not confluent), being anteriorly situated just below the alar notch, which is not very marked. The vertebral body is quite narrow and has a pointed protuberance on its caudal margin. The cranial articular fovea is wide and slightly expanded cranially on its dorsal portion.
Results
Morphological Comparisons with Other Miocene Hyaenids
Comparisons with Other Hyaenid Genera
Overall, the dentition of IPS62078 is more slender (it displays relatively narrower premolars) than Adcrocuta eximia (see Werdelin and Solounias 1990, 1991; Turner et al. 2008). IPS62078 further differs from A. eximia in the larger P4 protocone, the presence of a p3 mesial accessory cusp, the longer and tricuspid m1 talonid (instead of bicuspid, as Adcrocuta lacks the hypoconulid), and the retention of m2.
The dentition of the described specimen also does not resemble that of Chasmaporthetes, which is characterized by more slender and trenchant premolars with well-developed distal accessory cusps, as well as several more derived features that are absent in Hyaenictis spp. (Turner et al. 2008). Thus, IPS62078 differs from Chasmaporthetes (see Kurtén and Werdelin 1988; Werdelin and Solounias 1991; Werdelin and Turner 1996; Antón et al. 2006; Tseng et al. 2013) in the presence of p1, the relatively broader cheek teeth, the less developed accessory cusps in the premolars, the mesial and distal accessory cusps of the premolars aligned with the main cusp (instead of lingually tilted), the presence of metaconid (even if vestigial), the tricuspid m1 talonid (Chasmaporthetes lacks the hypoconulid and sometimes the hypoconid), and the presence of m2.
Compared to Lycyaena, the dentition of IPS62078 is larger, somewhat stouter (i.e., it displays relatively broader cheek teeth), and is further characterized by smaller and more indistinct premolar accessory cusps (Werdelin 1988; Werdelin and Solounias 1991). The specimen further differs from Lycyaena (see Werdelin 1988; Werdelin and Solounias 1991; Werdelin and Peigné 2010) in the occlusal morphology of the m1 (with a reduced instead of well-developed metaconid, and a shorter talonid with less developed cuspids), the presence of m2, and the deeper and stouter mandibular corpus.
In terms of mandibular robusticity and relative breadth of the lower premolars, IPS62078 resembles Lycyaenops, in spite of numerous differences in the occlusal morphology of the lower dentition (see Kretzoi 1938; Werdelin and Solounias 1991; Werdelin 1999; Werdelin and Peigné 2010). Thus, the described specimen mainly differs from Lycyaenops in the presence of p1, the better developed p3 accessory cusps, the less developed p4 distal accessory cusp, the presence of an m1 metaconid (even if reduced), and the longer and tricuspid (instead of bicuspid) m1 talonid.
With regard to tooth enamel microstructure, although the top portion of the cheek teeth crowns in IPS62078 are worn away, their preserved middle and bottom portions allow one to ascertain that, on the outer enamel surface, the HSB clearly display a zig-zag pattern (except closest to the cervix; Fig. 3). In this regard, the condition of the described specimen is most similar to Chasmaporthetes, in which the zig-zag pattern (unlike in extant and extinct fully-developed bone-cracking hyaenids, such as Adcrocuta) generally does not reach the base of the crown (Tseng 2012). In contrast, the condition of IPS62078 is much more similar to bone-cracking taxa than to that displayed by Lycyaena, which does not display a zig-zag pattern of HSB at any crown level (Tseng 2012).
Comparisons with Other Hyaenictis Species
The dentition of IPS62078 more closely resembles that of other species of Hyaenictis, except ?H. wehaietu from the late Miocene and early Pliocene of Ethiopia, which is smaller, lacks the p1 and m2, and further displays a well-developed and isolated m1 metaconid (Haile-Selassie and Howell 2009). In particular, IPS62078 resembles the other three species of Hyaenictis in the possession of a long and slender mandible with m2, the weak development of the mesial accessory cusp in the lower premolars, the short m1 without a well-developed metaconid, and the presence of more than a single cuspid in the m1 talonid (Werdelin et al. 1994).
Compared to the type species of the genus, H. graeca (Fig. 4j, k; Gaudry 1861; Howell and Petter 1985; Werdelin and Solounias 1990, 1991; Werdelin et al. 1994), IPS62078 (Fig. 4a–c) displays a similarly robust mandibular corpus that is only slightly curved on its inferior margin behind the m1 (although comparisons are hindered by the juvenile status of the holotype mandible of H. graeca). Both specimens also resemble one another in the retention of p1 and m2, as well as in the m1 occlusal morphology (characterized by a greatly reduced metaconid and a tricuspid talonid with a large hypoconid). However, IPS62078 differs from H. graeca in several features, namely the relatively broader lower premolars, the lower p2 protocone, the more restricted and simpler distal cingulids in the premolars, the p3 mesial accessory cusp, p3 aligned with the protoconid (instead of lingually tilted), and the smaller distal accessory cusp but larger distal crest in the p4.
IPS62078 differs even more clearly from H. hendeyi (Fig. 4d–f) from the earliest Pliocene of South Africa (Hendey 1974, 1978; Werdelin et al. 1994; Werdelin and Peigné 2010) in the relatively broader lower cheek teeth with a subrectangular (instead of elliptical) occlusal outline, the retention of the p1, the p2 with a lower protoconid and a smaller and more indistinct distal accessory cusp, and the tricuspid m1 talonid (instead of bicuspid, as H. hendeyi lacks the hypoconulid).
Differences are less marked when IPS62078 is compared with the holotype of H. almerai (Fig. 4g–i), from the similarly-aged Vallès-Penedès locality of SMT (Villalta Comella and Crusafont Pairó 1948; Werdelin and Solounias 1991; Werdelin et al. 1994), whose lower cheek teeth are somewhat more elongated (relatively narrower), but which nevertheless display a less oval occlusal outline than in H. graeca and H. hendeyi (thus more closely resembling IPS62078). Although the latter specimen resembles H. almerai (as well as H. graeca and H. hendeyi) in the retention of m2, it differs from H. almerai (and more closely resembles H. graeca) in the longer mandibular diastema and the presence of a p1 alveolus. Given the very small available sample sizes for European Hyaenictis, it cannot be discounted that the presence/absence of p1 was variable, as in some other hyaenids (e.g., Lycyaena; Werdelin 1988; Werdelin and Solounias 1991; Werdelin and Peigné 2010). In addition, in H. hendeyi the p1 is absent (Werdelin et al. 1994) suggesting that the presence of this tooth is variable in the genus as a whole. Nevertheless, IPS62078 further differs from H. almerai in several other occlusal features, including the smaller distal accessory cusp surrounded by a low distal cingulid in the p2, the mesial accessory cusps of the p3 and p4 aligned with the protoconid (instead of lingually tilted), and the more developed distal cingulid in the p4.
Morphometric Comparisons
Comparisons of dental size for the lower cheek teeth are reported by means of bivariate plots of buccolingual breadth (B) vs. mesiodistal length (L) in Fig. 5, whereas dental proportions are assessed by means of boxplots (Fig. 6) and ANOVA comparisons of log-transformed breadth/length index (BLI). Among the analyzed genera (Adcrocuta, Chasmaporthetes, Lycyaena, and Hyaenictis for p2–m1, and Lycyaenops for p2–p3), there are significant differences (p < 0.001) in dental proportions for all the analyzed lower cheek teeth (F = 107.22 for p2, F = 79.16 for p3, F = 64.68 for p4, and F = 6.68 for m1). Among other differences (see below), the bone-cracking Adcrocuta differs from the remaining genera (except for Lycyaenops) by displaying in all instances significantly broader premolars, with p < 0.001 (see also Figs. 5a–c and 6a–c). In the case of the lower carnassial, there are no significant differences between Hyaenictis and Adcrocuta, which tend to display a broader m1 than Chasmaporthetes and Lycyaena (although differences between Hyaenictis and Chasmaporthetes are not significant at p < 0.05; see also Figs. 5d and 6d). The lower carnassial is thus the least useful cheek tooth to discriminate between cursorial and bone-cracking hyaenids, and displays considerable overlap between all the analyzed genera. The proportions of the p2 (Fig. 6a) in Hyaenictis most closely resemble those of Chasmaporthetes, which are somewhat intermediate between the broader proportions of Adcrocuta and the narrower proportions (p < 0.001) of Lycyaena, although Hyaenictis does not significantly differ in this regard from any of the studied genera other than Adcrocuta. For both p3 (Figs. 5b and 6b) and p4 (Figs. 5c and 6c), all the cursorial hyaenids overlap and do not display significant differences among them, showing narrower proportions than Adcrocuta (although with some overlap, more marked in the p3 of Lycyaenops).

Bivariate plots of buccolingual breadth vs. mesiodistal length in the cheek teeth of Hyanictis aff. almerai, compared with those of other selected Miocene hyaenids. a, p2. b, p3. c, p4. d, m1. Convex hulls encompass the range of variation in our sample of the bone-cracking Acrocuta

Boxplots of breadth/length index (BLI) in the cheek teeth of Hyaenictis aff. almerai, compared with those of other selected Miocene hyaenids. a, p2. b, p3. c, p4. d, m1
With regard to IPS62078, its p2 most closely resembles in both size (Fig. 5a) and proportions (Fig. 6a) those of Chasmaporthetes and H. hendeyi, thus being intermediate between Lycyaena and Adcrocuta. In contrast, for the remaining premolars IPS62078 most clearly overlaps with Adcrocuta (Figs. 5b, c and 6b, c), generally displaying broader proportions than all the remaining analyzed hyaenids (including Hyaenictis spp.). Finally, for the m1 (Figs. 5d and 6d), IPS62078 most extensively overlaps with Hyaenictis and Adcrocuta, although as stated above all the studied genera overlap to a large extent. Overall, based on dental size and proportions, IPS62078 only most closely resembles species of Hyaenictis and other cursorial hyaenids regarding the p2 and the m1, whereas the p3 and p4 display a relative breadth closer to that of the bone-cracking Adcrocuta.
The PCA results (Fig. 7; Table 2) further confirm that the lower cheek tooth proportions of IPS62078 are intermediate between those of Hyaenictis, on the one hand, and those of Adcrocuta, on the other. A bivariate plot of PC2 vs. PC1 (Fig. 7) shows that Adcrocuta does not overlap with the cursorial hyaenids, and that IPS62078 falls close to (but outside) the convex hull depicting the variability of Adcrocuta, overlapping with the latter in the PC1 and with Hyaenictis (which more closely resembles Chasmaporthetes and Lycyaena) in the PC2. The investigated bone-cracking and cursorial hyaenids mostly differ in PC1, which explains most of the variance (85 %) and is largely driven by the buccolingual breadth of the premolars (mostly p2, but also p3 and p4). For this component, IPS62078 overlaps with Adcrocuta and falls far from the remaining Hyaenictis species and other cursorial hyaenids, as a result of the relatively broader p3 and p4 (which more closely resemble the condition of Adcrocuta). The PC2 explains 7 % of variance and is mostly positively driven by the breadth of the m1 and, to a lesser extent, the p4. On this axis, which segregates Hyaenictis spp. from both the cursorial hyaenids and Adcrocuta (with only minimal overlap), IPS62078 only overlaps with the former. Overall, the results of the PCA highlight that IPS62078 clearly differs from the analyzed cursorial hyaenids other than Hyaenictis, although it does not comfortably fit into the latter or Adcrocuta. Given that only three specimens of European Hyaenictis are available, it is not possible to confidently discount that their variability in dental proportions can be accommodated within a single genus. However, IPS62078 is clearly in this regard much more similar to (albeit less extreme than) the bone-cracking condition of Adcrocuta.

Bivariate plot of the two first axes (PC2 vs. PC1) of the principal component analysis based on breadth/length index of the lower cheek teeth of Hyaenictis aff. almerai, compared with those of other selected Miocene hyaenids. The variability of each genus is denoted by convex hulls
Discussion
Taxonomic Attribution
The skull with mandible from ROS-D described in this paper (IPS62078) differs from the bone-cracking Miocene hyena Adcrocuta in several features, such as the presence of m2, the smaller premolar accessory cusps, the larger P4 protocone, the reduced m1 metaconid, and the tricuspid and longer m1 talonid. The former further differs from cursorial Miocene hyenas, such as Chasmaporthetes, Lycyaena, and Lycyaenops in several occlusal features, namely: the presence of m2, the presence of p1 and the tricuspid m1 talonid (only compared to Chasmaporthetes and Lycyaenops), the shorter m1 talonid with smaller cuspids (compared to Lycyaena), the reduced m1 metaconid (instead of well developed, as in Lycyaena, or entirely absent, as in Chasmaporthetes and Lycyaenops), the smaller premolar accessory cusps (compared to Chasmaporthetes and Lycyaena, and to Lycyaenops in the case of the p4), and the lingually placed premolar accessory cusps (compared to Chasmaporthetes).
In the above-mentioned features and other characters, the described specimen from ROS-D (IPS62078) more closely resembles species of Hyaenictis other than ?H. wehaietu, and especially the Miocene European species H. graeca and H. almerai. Similarities include the presence of m2, the weakly developed anterior accessory cusps of the premolars, the P4 metastyle larger than the paracone, and the mesiodistally short m1 (with absent or greatly reduced metaconid, and a bicuspid or tricuspid talonid). In the retention of p1, IPS62078 more closely resembles H. graeca, although we do not consider this feature to be taxonomically relevant, as the small available samples preclude ascertaining whether this feature might have been variable in European Hyaenictis. In fact, the described specimen does not comfortably fit into any of these species, from which it differs in the stouter (relatively broader) lower premolars (especially p3 and p4), resulting in dental proportions that are intermediate between those of previously known Hyaenictis (plus other cursorial hyaenids such as Chasmaporthetes) and those of the bone-cracking Adcrocuta. IPS62078 further differs from other species of Hyaenictis (especially H. graeca and H. hendeyi) in some details of dental morphology, such as the more rectangular (less elliptical) occlusal outline of the premolars, the presence of a distal cingulid (behind the distal accessory cusp) in p2, and the lingually tilted mesial accessory cusps of the premolars (instead of aligned with the protoconid).
The above-mentioned differences preclude attribution of the described specimen to any of the previously known species of Hyaenictis. However, given that only a single individual is available from ROS-D and that the samples for both H. graeca and H. almerai are also very small, the possibility cannot be conclusively discounted that the above-mentioned differences in dental proportions and minor occlusal details merely reflect intraspecific variation. Specimen IPS62078 from ROS-D and the holotype of H. almerai have a similar geographic provenance and are roughly coeval (the former is no more than 0.5 million years younger, probably less). Taking all this evidence together, we choose to attribute IPS62078 to Hyaenictis aff. almerai. This assignment recognizes the closer morphological similarities with H. almerai and further takes into account the similar and age and geographic provenance, while at the same time it does not discount the possibility that IPS62078 represents a previously unknown species. Additional specimens from the late Miocene of the Vallès-Penedès Basin and elsewhere would be required to further ascertain the taxonomic allocation of European specimens currently attributed to Hyaenictis spp.
Implications for Feeding Behavior
European Hyaenictis have generally been included in the hyaenid ecomorphotype 4 (cursorial meat- and bone-eating hyenas, including among others Chasmaporthetes and Lycyaena), instead of ecomorphotype 5 (transitional bone-cracking hyenas, like Belbus Werdelin and Solounias, 1991, and Metahyaena Viranta and Werdelin, 2003) or 6 (fullydeveloped bone-cracking hyenas such as Adcrocuta; Werdelin 1996; Werdelin and Solounias 1996; Turner et al. 2008). However, some previous studies have favored some degree of durophagy in cursorial Miocene hyaenids such as Chasmaporthetes, in spite of noting their lesser bone-cracking abilities compared to fully developed bone-crackers (Kurtén and Werdelin 1988; Werdelin et al. 1994; Ferretti 1999; Antón et al. 2006). This was further confirmed by a finite elements analysis of the skull of Chasmaporthetes (Tseng et al. 2011), according to which this taxon would have been able to the resist the masticatory stresses generated by a bone-cracking diet (although less efficiently than Crocuta). In agreement with this, Tseng (2011, 2012) further showed that Chasmaporthetes displays a zig-zag pattern of HSB on the outer enamel surface of at least the top and middle crown portions of the premolars, and even on the bottom portion of p4, as is characteristic of all the cheek teeth of extant durophagous hyaenids and some extinct bone-cracking hyaenas (such as Adcrocuta, Pliocrocuta Kretzoi, 1938, and Pachycrocuta Kretzoi, 1938; Stefen 1997; Ferretti 1999, 2007; Stefen 1999; Stefen and Rensberger 1999; Tseng 2011, 2012; Vinuesa et al. 2014). While enamel microstructure is quite plastic from an evolutionary perspective (Tseng 2012), morphological changes in craniodental features most clearly evidence that some Miocene cursorial hyaenids such as Chasmaporthetes were, to some extent, already adapted to (albeit not specialized for) bone-cracking (Tseng et al. 2011). Based on the development of the zig-zag pattern of the HSB of IPS62078, Hyaenictis aff. almerai would have been at least as durophagous as Chasmaporthetes, although less so than Adcrocuta, which is entirely comparable to extant bone-cracking hyaenids and in some respects more derived than they are.
Furthermore, the premolar proportions of Hyaenictis aff. almerai from specimen IPS62078 (especially those of p3 and p4) are intermediate between those of Chasmaporthetes and previously known Hyaenictis spp., on the one hand, and those of Adcrocuta, on the other. This is shown not only by the breadth/length proportions of p3 and p4 separately, but also a multivariate analysis that takes into account all the lower cheek teeth (except p1) simultaneously. Our results therefore suggest for IPS62078 from ROS-D a higher commitment to durophagy than in other hyaenids customarily included in ecomorphotype 4 (including other Hyaenictis), thus being best qualified as a transitional bone-cracking hyena (ecomorphotype 5). This is further supported by other features of the described specimen, such as the deep masseteric fossa and the large insertions for the pterygoids, which are shown (although to a greater extent) by extant bone-cracking hyaenas as well as Adcrocuta (Werdelin and Solounias 1991, 1996; Turner et al. 2008). Unfortunately, the small available samples of European Hyaenictis preclude discerning whether IPS62078 differs from the latter in displaying greater durophagous adaptations (as it seems), or whether European Hyaenictis as a whole were somewhat intermediate between cursorial hyaenids such as Chasmaporthetes and fully-developed bone-crackers like Adcrocuta. In any case, the presence of some durophagous adaptations in the studied material, coupled with the fully-developed bone-cracking adaptations of the roughly coeval Adcrocuta eximia (which is first recorded in MN10 of Ravin de la Pluie, Greece; Bonis and Koufos 1981; Turner et al. 2008), support the contention that the early evolution of the bone-cracking ecomorphotype (the most common among extant hyaenids) in Europe was not related to competition with canids of the genus Eucyon Tedford and Qiu, 1996, which did not disperse from America into Eurasia until the Turolian (MN12; Sotnikova and Rook 2010).
Another interesting possibility raised by the present study derives from the fact that the earliest Hyaenictis (such as the one described here) and the earliest Lycyaena (i.e., L. crusafonti) are the most potentially durophagous species of their respective lineages (Kurtén 1976; Werdelin 1988; Werdelin and Solounias 1991; Werdelin et al. 1994). Therefore, it is possible that the earliest members of lineages within the Chasmaporthetes clade were more durophagous than later representatives of these lineages. This may imply that some degree of durophagy first evolved earlier than customarily assumed, in some (currently unknown) representative of the hyaenictitheres, and that reduction of durophagy subsequently evolved in parallel along several lineages.
Summary and Conclusions
Here, we describe a nearly complete skull with associated mandible and atlas of a medium-sized hyaenid from the late Miocene (MN10) of ROS-D (Vallès-Penedès Basin, NE Iberian Peninsula). Compared to other late Miocene hyaenids from Europe, the described specimen (IPS62078) most closely resembles the European species of Hyaenictis in several features, such as the retention of m2 and the reduced m1 metaconid. The described remains represent the most complete specimen recovered so far of European Hyaenictis, previously recorded from fragmentary remains attributed to H. almerai (MN10 of the same basin) and H. graeca (MN12 of Greece). In spite of the similar chronology and geographic provenance to H. almerai, IPS62078 from ROS-D is only tentatively assigned to this taxon (Hyaenictis aff. almerai). This is because the described specimen does not fit well into any of the previously known species of Hyaenictis, from which it differs by displaying cheek tooth proportions (in particular, relatively broader premolars) and other occlusal and mandibular features more similar to Adcrocuta, the earliest European hyaena with fully developed bone-cracking adaptations. Coupled with tooth enamel microstructure, this indicates a greater degree of durophagy in H. aff. almerai than previously recognized for Hyaenictis, seemingly intermediate between cursorial (ecomorphotype 4) and fully bone-cracking (ecomorphotype 6) hyaenids. This suggests that, during the Vallesian, bone-cracking adaptations evolved in parallel among several Eurasian hyaenid taxa due to selection pressures unrelated to competition with canids, which did not disperse from America into the Old Word until much later.
Notes
For the use of open nomenclature we follow Bengtson (1988).
References
Antón M, Turner A, Salesa M, Morales J (2006) A complete skull of Chasmaporthetes lunensis (Carnivora, Hyaenidae) from the Spanish Pliocene site of La Puebla de Valverde (Teruel). Estudios Geol 62: 375–388
Bataller JR (1921) Mamífers fòssils de Catalunya. Nota paleontològica. Butll Inst Cat Hist Nat 21: 80–86
Bengtson P (1988) Open nomenclature. Palaeontology 31: 223–227
Bonis L de, Koufos G (1981) A new hyaenid (Carnivora, Mammalia) in the Vallesian (late Miocene) of northern Greece. Sci Ann Fac Phys Math Univ Thessaloniki 21: 79–94
Cabrera L, Roca E, Garcés M, de Porta J (2004) Estratigrafía y evolución tectonosedimentaria oligocena superior-neógena del sector central del margen catalán (Cadena Costero-Catalana). In: Vera JA (ed) Geología de España. SGE-IGME, Madrid, pp 569–573
Casanovas-Vilar I, Alba DM, Robles JM, Moyà-Solà S (2011) Registro paleontológico continental del Mioceno de la cuenca del Vallès-Penedès. Paleontol Evol memòria especial 6: 55–80
Casanovas-Vilar I, Madern A, Alba DM, Cabrera L, García-Paredes I, van den Hoek Ostende L, DeMiguel D, Robles JM, Furió M, van Dam J, Garcés M, Angelone C, Moyà-Solà S (2015 published online) The Miocene mammal record of the Vallès-Penedès Basin (Catalonia). C R Palevol. doi:10.1016/j.crpv.2015.07.004
de Gibert JM, Casanovas-Vilar I (2011) Contexto geológico del Mioceno de la cuenca del Vallès-Penedès. Paleontol Evol memòria especial 6: 39–45
Evans HE, de Lahunta A (2013) Miller’s Anatomy of the Dog. 4th ed. Elsevier, St. Louis
Ferretti MP (1999) Tooth enamel structure in the hyaenid Chasmaporthetes lunensis lunensis from the late Pliocene of Italy, with implications for feeding behavior. J Vertebr Paleontol 19: 767–770
Ferretti MP (2007) Evolution of bone-cracking adaptations in hyaenids (Mammalia, Carnivora). Swiss J Geosci 100: 108–128
Gaudry A (1861) Note sur les carnassiers fossiles de Pikermi (Grèce). Bull Soc Geol Fr 18: 527–538
Ginsburg L (1977) Les carnivores du Miocène de Beni Mellal (Maroc). Geol Mediterr 4: 225–240
Haile-Selassie Y, Howell FC (2009) Carnivora. In: Haile-Selassie Y, WoldeGabriel G (eds) Ardipithecus kadabba: Late Miocene Evidence from the Middle Awash, Ethiopia. University of California Press, Berkeley, pp 237–275
Hammer Ø, Harper DAT (2006) Paleontological Data Analysis. Blackwell Publishing, Malden
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1): Art. 4 [9 pp.]. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hendey QB (1974) The late Cenozoic Carnivora of the south-western Cape Province. Ann S Afr Mus 63: 1–369
Hendey QB (1976) The Pliocene fossil occurrences in ‘E’ Quarry, Langebaanweg, South Africa. Ann S Afr Mus 69: 215–247
Hendey QB (1978) Late Tertiary Hyaenidae from Langebaanweg, South Africa, and their relevance to the phylogeny of the family. Ann S Afr Mus 76: 265–297
Howell FC, Petter G (1985) Comparative observations on some middle and upper Miocene hyaenids. Genera: Percrocuta Kretzoi, Allohyaena Kretzoi, Adcrocuta Kretzoi (Mammalia, Carnivora, Hyaenidae). Geobios 18: 419–476
Koufos GD (2011) The Miocene carnivore assemblage of Greece. Estudios Geol 67: 291–320
Kretzoi M (1938) Die Raubtiere von Gombaszög nebst einer Übersicht der Gesamtfauna. Ann Mus Nat Hung 31: 88–157
Kurtén B (1976) Fossil Carnivora from the late Tertiary of Bled Douarah and Cherichira, Tunisia. Notes Serv géol Tunis 42: 177–214
Kurtén B, Werdelin L (1988) A review of the genus Chasmaporthetes Hay, 1921 (Carnivora, Hyaenidae). J Vertebr Paleontol 8: 46–66
Madurell-Malapeira J, Robles JM, Casanovas-Vilar I, Abella J, Obradó P, Alba DM (2014) The scimitar-toothed cat Machairodus aphanistus (Carnivora: Felidae) in the Vallès-Penedes Basin (NE Iberian Peninsula). C R Palevol 13: 569–585
Morales J, Pickford M, Soria D (2005) Carnivores from the late Miocene and basal Pliocene of the Tugen Hills, Kenya. Rev Soc Geol Esp 18: 39–61
Qiu Z (1987) Die Hyaeniden aus dem Ruscinium und Villafranchium Chinas. Münch Geowiss Abh A 9: 1–108
Solounias N (1981) The Turolian fauna from the island of Samos, Greece, with special emphasis on the hyaenids and the bovids. Contrib Vertebr Evol 6: 1–232
Sotnikova M, Rook L (2010) Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the late Miocene to early Pleistocene. Quaternary Internatl 212: 86–97
Stefen C (1997) Differentiations in Hunter-Schreger bands of carnivores. In: Koenigswald Wv, Sander PM (eds) Tooth Enamel Microstructure. A.A. Balkema, Rotterdam, pp 123–136
Stefen C (1999) Enamel microstructure of recent and fossil Canidae (Carnivora: Mammalia). J Vertebr Paleontol 19: 576–587
Stefen C, Rensberger JM (1999) The specialized structure of hyaenid enamel: description and development within the lineage—including percrocutids. Scanning Microsc 13: 363–380
Tseng ZJ (2011) Variation and implications of intra-dentition Hunter-Schreger band pattern in fossil hyaenids and canids (Carnivora, Mammalia). J Vertebr Paleontol 31: 1163–1167
Tseng ZJ (2012) Connecting Hunter-Shreger band microstructure to enamel microwear features: new insights from durophagous carnivores. Acta Palaeontol Pol 57: 473–484
Tseng ZJ, Antón M, Salesa MJ (2011) The evolution of the bone-cracking model in carnivorans: cranial functional morphology of the Plio-Pleistocene cursorial hyaenid Chasmaporthetes lunensis (Mammalia, Carnivora). Paleobiology 37: 140–156
Tseng ZJ, Li Q, Wang X (2013) A new cursorial hyaena from Tibet, and analysis of biostratigraphy, paleozoogeography, and dental morphology of Chasmaporthetes (Mammalia, Carnivora). J Vertebr Paleontol 33: 1457–1471
Turner A (1988) On the claimed occurrence of the hyaenid genus Hyaenictis Gaudry at Swartkrans (Mammalia, Carnivora). Ann Transvaal Mus 34: 523–533
Turner A, Antón M, Werdelin L (2008) Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe. Geobios 41: 677–687
Villalta Comella JF de, Crusafont Pairó M (1941) Noticia preliminar sobre la fauna de carnívoros del Mioceno continental del Vallés-Penedés. Bol R Soc Esp Hist Nat 39: 201–208
Villalta Comella JF de, Crusafont Pairó M (1943) Los vertebrados del Mioceno continental de la cuenca del Vallés-Panadés (provincia de Barcelona). I. Insectívoros. II. Carnívoros. Bol Inst Geol Min Esp 56: 145–336
Villalta Comella JF de, Crusafont Pairó M (1948) Nuevas aportaciones al conocimiento de los carnívoros pontienses del Vallés-Penedés. In: Miscelánea Almera. Publ Inst Geol 7: 81–121
Vinuesa V, Madurell-Malapeira J, Ansón M, Alba DM (2014) New cranial remains of Pliocrocuta perrieri (Carnivora, Hyaenidae) from the Villafranchian of the Iberian Peninsula. Boll Soc Paleontol Ital 53: 39–47
Werdelin L (1988) Studes of fossil hyaenas: the genera Thalassictis Gervais ex Nordmann, Palhyaena Gervais, Hyaenictitherium Kretzoi, Lycyaena Hensel and Palinhyaena Qiu, Huang & Guo. Zool J Linn Soc 92: 211–265
Werdelin L (1996) Community-wide character displacement in Miocene hyaenas. Lethaia 29: 97–106
Werdelin L (1999) Studies of fossil hyaenas: affinities of Lycyaenops rhomboideae Kretzoi from Pestlörinc, Hungary. Zool J Linn Soc 126: 307–317
Werdelin L, Peigné WJ (2010) Carnivora. In: Werdelin L, Sanders WJ (eds) Cenozoic Mammals of Africa. University of California Press, Berkeley, pp 603–657
Werdelin L, Solounias N (1990) Studies of fossil hyaenids: the genus Adcrocuta Kretzoi and the interrelationships of some hyaenid taxa. Zool J Linn Soc 98: 363–386
Werdelin L, Solounias N (1991) The Hyaenidae: taxonomy, systematics and evolution. Fossils Strata 30: 1–104
Werdelin L, Solounias N (1996) The evolutionary history of hyaenas in Europe and western Asia during the Miocene. In: Bernor RL, Fahlbusch V, Mittmann H-W (eds) The Evolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press, New York, pp 290–306
Werdelin L, Turner A (1996) The fossil Hyaenidae of Africa: present status. In: Stewart KM, Seymour KL (eds) Palaeoecology and Palaeoenvironments of Late Cenozoic Mammals. University of Toronto Press, Toronto, pp 637–659
Werdelin L, Turner A, Solounias N (1994) Studies of fossil hyaenids: the genera Hyaenictis Gaudry and Chasmaporthetes Hay, with a reconsideration of the Hyaenidae of Langebaanweg, South Africa. Zool J Linn Soc 111: 197–217
Acknowledgments
This paper has been funded by the Spanish Ministerio de Economía y Competitividad (CGL2014-54373-P) and the Generalitat de Catalunya (2014 SGR 416 GRC, and predoctoral grant 2015FI-B00088 to VV). The work of LW on fossil carnivorans is supported by the Swedish Research Council (VR).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Vinuesa, V., Madurell-Malapeira, J., Werdelin, L. et al. A New Skull of Hyaenictis Gaudry, 1861 (Carnivora, Hyaenidae) Shows Incipient Adaptations to Durophagy. J Mammal Evol 24, 207–219 (2017). https://doi.org/10.1007/s10914-016-9334-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10914-016-9334-0