
Abstract
Global climate change affects aquatic habitats in a number of ways that pose challenges for aquatic insect populations. Increasing water temperature and corresponding decreases in dissolved oxygen can impact respiratory behaviors, even in air-breathing aquatic taxa. Crawling water beetles (Coleoptera: Haliplidae) exhibit a combined respiratory strategy that employs an air store that is periodically replenished at the water surface. The frequency at which beetles surface to replenish the bubble is determined both by oxygen demand and by the capacity of the air store to uptake oxygen from the surrounding water via diffusion. However, little is known of how changes in water temperature and dissolved oxygen will affect submersion time. We investigated this question in Peltodytes callosus, a species of crawling water beetle that is widely distributed across the American West. We manipulated temperature and dissolved oxygen to mimic changes associated with global climate change and recorded the time between surfacing events. We found that beetles stayed submerged for shorter durations in response to both increasing water temperature and decreasing dissolved oxygen. Our results suggest that beetles may be able to modify their surfacing behavior to respond to climate-induced changes in water quality.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Although respiration in aquatic beetles has been widely studied (Chaui-Berlinck et al. 2001; Kehl and Dettner 2009; Calosi 2012; Madsen 2012), less is known about the environmental determinants of respiratory behaviors or their plasticity in the face of climate change (Calosi et al. 2008; Verberk and Bilton 2014). Submersion time, the interval between surfacing events, is an important component of respiratory behavior that is closely linked to environmental conditions and has broad fitness consequences for beetle populations. Feeding and reproduction occur while animals are submerged (Hickman 1931a), and time spent at the surface increases the risk of predation (Wańkowski and Thorpe 1979). Environmentally induced changes in submersion time may have widespread effects on food web structure and ecosystem functioning.
Most aquatic beetles breathe from an air store that is carried against the abdomen while diving and periodically refreshed at the water surface. In many cases, oxygen can diffuse from the water into the air store during the dive period, and submersion time is extended in highly oxygenated waters (Madsen 2012). Under ideal conditions, beetles may use this “physical gill” to remain submerged for weeks between surface visits (Hickman 1931b; Seymour and Matthews 2013). However, in nature conditions are rarely ideal, and beetles may adjust dive duration in response to changes in their environment.
One of the direct effects of global climate change on water quality is increasing water temperature (Lake 2003; Cook et al. 2014), and this warming trend may impact beetle diving behavior. Temperature affects metabolic rate and oxygen demand (Gillooly et al. 2001), as well as the solubility and diffusivity of oxygen in water (Verberk et al. 2011). Some aquatic beetles are able to modify surfacing behaviors in response to temperature (Calosi et al. 2007). However, this response could be the result of changes in metabolic activity, dissolved oxygen, or other environmental factors that change concurrently with temperature. Given that time spent at the surface is associated with predation and other species interactions, determining the drivers of surfacing behavior is important in order to understand the food web consequences of ongoing environmental changes.
Here we tested the effects of temperature and dissolved oxygen on submersion time in the crawling water beetle, Peltodytes callosus Leconte. This small (3-4 mm) beetle is abundant in ponds, marshes, and streams in the western United States (Matheson 1912; Roberts 1913; Durfee et al. 2005) and is exposed to a wide range of thermal conditions in its natural habitat. It feeds on submerged macrophytes and detritus (Merritt et al. 2008) and facilitates bioturbation of anoxic sediments in shallow pond margins. We hypothesized that increasing temperature and decreasing dissolved oxygen would result in shorter submersion time, both as independent factors and in combination.
Materials and Methods:
Study Organism
In February 2016 we collected P. callosus from macrophyte beds in Corte Madera Pond, San Diego County, California, USA. Water temperature at the time of collection was 19 °C. Beetles were maintained in the laboratory in aerated water at 18 °C, provided artificial vegetation for shelter, and fed brine shrimp until the experiment commenced.
Experimental Design
We measured the effects of temperature and dissolved oxygen on submersion time using a factorial experimental design that crossed two dissolved oxygen treatments (Constant/Variable) with three temperature treatments (Fig. 1). Temperature treatments were applied in three incubators (ThermoFisher Scientific Precision™ Low Temperature BOD Refrigerated Incubator): constant 15 °C (“Cold”), constant 25 °C (“Warm”), and ramped temperature (“Ramp”), in which we increased the temperature by 3 °C every two days, from 15 to 39 °C. Cold and Warm temperatures were selected based on median and maximum temperatures recorded at the collection site (annual 5-year median water temperature: 16 °C; maximum recorded water temperature: 26 °C). The static temperature treatments differentiated between the effects of temperature and experiment day, since these variables were correlated in the Ramp treatment. Aerators were installed in the Constant oxygen treatments to maintain constant dissolved oxygen despite changes in temperature. This experimental design yielded six treatments of 30 individuals, each placed in a separate 500 mL plastic cup.

Experimental design. Each of the three incubators held two trays and each housed 15 small cups containing a single beetle. Trays were filled with water, which was allowed to flow into the cups through small holes. Constant dissolved oxygen was maintained in one tray using an aerator (Fig. 2), and dissolved oxygen was allowed to vary naturally with temperature in the second tray
Every two days we recorded the submersion time of 8 randomly selected individuals from each treatment (n = 8). Beetles could not be observed in the incubators, so we allowed beetles to acclimate to lab conditions for five minutes before beginning behavioral observations. Although observations were made at room temperature, water temperature in the cups did not change during the acclimation period. We used hand-held stopwatches to measure the amount of time each individual remained submerged between its first and second surface visit, hereafter, “submersion time.” Note that other investigators have labeled this duration as “surfacing frequency” (Calosi et al. 2007) or “respiratory interval” (Hickman 1931b); submersion time is more appropriate in our case because we did not track surfacing intervals through time. Dissolved oxygen was measured every two days with the indigo carmine method (CHEMets water test kit). The experiment was terminated on Day 18, when the Ramp treatment reached 39 °C and all beetles in that treatment perished.
Analysis
We calculated the effects of temperature and dissolved oxygen on submersion time using mixed effects models to account for repeated measurements taken on randomly selected individuals. First we created a global model to examine the overall effects of temperature, dissolved oxygen, and experiment day on submersion time. We conducted pairwise comparisons among temperature treatments using least-squares means (LSM).
Then we examined the effect of dissolved oxygen and experiment day on submersion time within each temperature treatment. Likelihood ratio tests (LRT) were used to assess statistically significant differences among treatments with an alpha of 0.05.
We compared survival among treatments using Kaplan-Meier survival estimates and Mantel-Haenszel tests. Cox proportional hazards (CPH) tests were used to calculate hazards ratios for each temperature-by-aerator combination.
All analyses were conducted in R version 3.1.1 (R Core Team 2014) using the packages lme4 (Bates et al. 2011), lsmeans (Lenth 2016), and survival (Therneau 2015).
Results
Dissolved Oxygen
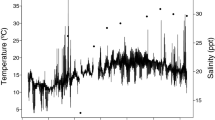
The aerator was effective in maintaining constant dissolved oxygen levels (7 ± 0.5 ppm) in the three temperature treatments (Fig. 2). Without aerators, dissolved oxygen was nearly constant in the cold (7 ± 0.5 ppm) and warm (7 ± 1 ppm) treatments, and decreased in the ramp treatment from 7 ppm to 4 ppm.

Effect of aerator presence/absence on dissolved oxygen. Solid lines: Aerator absent, Dashed lines: Aerator present
Submersion Time
Submersion time ranged from 105 to 1451 s. As hypothesized, surfacing behavior was affected by both dissolved oxygen and temperature (Fig. 3). In the global model, aerator, temperature, and experiment day were all significant predictors of submersion time (Aerator: LRT, X21 = 102, P < 0.001, Temperature: LRT, X22 = 144, P < 0.001, Day: LRT, X28 = 71.9, P < 0.001). Specifically, beetles remained submerged longer in the Cold treatment than in Warm or Ramp treatments (Cold vs Warm: LSM, P < 0.001; Cold vs Ramp: LSM, P < 0.001; Warm vs Ramp: P = 0.0542). The interaction of Temperature and Aerator was also a significant determinant of submersion time (LRT, X21 = 150, P < 0.001); the presence of an Aerator had a larger effect on submersion time in the Cold treatment than in the Ramp or Warm treatments. To examine the role of dissolved oxygen within temperature regime, we conducted the remainder of the analyses on each temperature treatment separately.

Submersion time by day in (a) Cold (15 °C), (b) Warm (25 °C), and (c) Ramp (15–39 °C) treatments. Solid lines: Aerator absent, Dashed lines: Aerator present
In the Cold (15°) treatment, the presence of an aerator increased mean submersion time by 272 ± 23.5 s (LRT: X21 = 48.7, P < 0.001), but day of the experiment did not have an effect (LRT: X28 = 6.88, P = 0.650). The aerator also increased submersion time in the Warm (25°) treatment, by 197 ± 21.5 s (LRT: X21 = 40.6, p < 0.001). Submersion time decreased throughout the experiment in the Warm treatment (LRT: X28 = 18.8, P = 0.0272) by an average of 51.4 s per day. There was a strong effect of both day and aerator in the Ramp (15–39 °C) treatment. Beetles with an aerator remained submerged for 342 ± 21.2 s longer than those without an aerator (LRT: X21 = 77.8, P < 0.0001). Additionally, submersion times in the Ramp treatment became sequentially shorter as the experiment progressed, by an average of 459 s per day (LRT: X28 = 200, P < 0.0001).
Survival
The presence of an aerator was associated with a decreased likelihood of mortality by a factor of 1.87 (Fig. 4; CPH, z = 2.15, P = 0.0319) across all temperature treatments. When compared with the cold treatment, the likelihood of mortality in the warm treatment increased by a factor of 3.53 (CPH, z = 3.16, P = 0.0016) and in the Ramp treatment by 7.54 (CPH, z = 4.87, P < 0.001).

Kaplan-Meier survival curves demonstrating the effect of treatment on survival in Cold (15 °C), Warm (25 °C), and Ramp (15–39 °C) treatments. Solid lines: Aerator absent, Dashed lines: Aerator present
Discussion
Here we provide evidence that temperature and dissolved oxygen affect submersion time and survival in a common and widespread crawling water beetle (P. callosus). Beetles remained submerged for shorter periods of time at higher temperatures, and submersion time decreased as temperature increased. The presence of an aerator increased submersion time under all temperature regimes. Decreasing dissolved oxygen and increasing temperature both decreased survival probability. These findings and their ecological implications are discussed below.
It is likely that submersion time in P. callosus decreased with increasing temperature due to higher oxygen demand at warmer temperatures, as has been shown in other aquatic beetles (De Ruiter et al. 1949). Temperature and metabolic rate are positively correlated (e.g., Contreras and Bradley 2011), and warmer conditions are known to cause an increase in oxygen demand in aquatic ectotherms (Verberk et al. 2011). Peltodytes callosus likely remained submerged for shorter periods of time in the Warm and Ramp treatments in order to refill their rapidly depleting air stores (Hickman 1931b).
Dissolved oxygen also affected submersion behavior, regardless of temperature treatment. The presence of an aerator increased submersion time for P. callosus, and beetles in aquaria with aerators had nearly half the risk of mortality as those without aerators. The fact that higher dissolved oxygen increased submersion times at all temperatures means that respiratory behaviors were likely determined by a combination of oxygen demand (due to metabolic rate) and the solubility of oxygen in water (Verberk et al. 2011). Our results indicate that beetles can indeed modify surfacing behavior in response to temperature and dissolved oxygen content, either in combination or as independent predictors.
The static temperature treatments (Warm and Cold) accounted for potential effects of trial duration on submersion time. We observed that submersion time decreased by almost a minute per day in the Warm treatment, suggesting that these animals may have been stressed even under a constant temperature. Indeed, beetles held at 25 °C were 3.5 times less likely to survive the 18-day experiment than those at 15 °C. The Ramp treatment increased mortality even further, by a factor of 7.5 over the Cold treatment, demonstrating the combined effect of exposure time and temperature increase. The maximum water temperature we have recorded at the collection site was 25.8 °C (August 2014), which suggests that increased beetle mortality may already be occurring during extreme summer conditions.
The combined influence of temperature and oxygen availability on respiration is further supported by a series of behavioral experiments by Hickman (1931b), in which he prevented haliplid beetles (Peltodytes edentulus and Haliplus triopsis) from surfacing and recorded survival at various temperatures. He found that beetles were able to survive without surfacing for up to seven days at 4–8 °C but that survival duration decreased as temperature increased. At 12–15 °C maximum survival was four days, and at 20 °C all beetles died within 48 h. Beetles that were allowed to surface in Hickman’s experiment survived the entire experiment (between 24 and 36 days, depending on the temperature).
Respiration has been more extensively studied in diving beetles (Dytiscidae) than in crawling water beetles (Haliplidae). Beetles in both families visit the surface to replenish an air store that is carried against the abdomen while submerged, and dytiscid beetles have also been shown to modify respiratory behavior in response to temperature. Calosi et al. (2007) quantified submersion time in dytiscid beetle Ilybius montanus at 20.5 °C and at 14.5 °C and found that these beetles also surfaced more frequently at the higher temperature. However, these investigators did not isolate the effect of dissolved oxygen from that of temperature.
The unique abdominal morphology of Haliplidae suggests that diffusion of dissolved oxygen into the air store may be particularly important for these beetles. Unlike dytiscid beetles, which exclusively carry air underneath the elytra, haliplids hold an air bubble using a combination of a subelytral chamber and extended coxal plates (Hickman 1931b). Air held under these plates connects to the subelytral air store, providing a larger respiratory surface area than in beetles relying on a subelytral bubble alone. This characteristic may make haliplids well-suited to take advantage of high dissolved oxygen conditions to prolong dive duration.
These environmentally induced behavioral changes are important in order to understand aquatic invertebrate community interactions under future global climate scenarios. Droughts are predicted to increase in frequency, severity, and duration across the western United States over the next century (Seager et al. 2007; Ruff et al. 2011; Seager et al. 2013; Cook et al. 2014), with widespread impacts on water quality and water availability for aquatic taxa (Lake 2003). Drought-stressed taxa may exhibit a range of behavioral modifications, including dietary changes (Crump 1983; Mas-Martí et al. 2010), drought-induced dispersal (Boersma and Lytle 2014), burial (Cover et al. 2015), or diapause/aestivation (Bogan 2017). These changes will alter species interactions, either directly or indirectly, and will modify the structure and functioning of aquatic food webs (Greig et al. 2013; Boersma et al. 2014).
The changes in beetle submersion times we observed in response to temperature and dissolved oxygen provide another example of an environmentally induced behavioral modification with potentially widespread ecological impacts. The capacity to regulate respiratory behavior in response to the environment may decrease species vulnerability to global climate warming (Verberk and Bilton 2014), but reduced submersion times may impart strong fitness consequences. Diving behavior is associated with feeding, reproduction, and predator evasion, and environmentally induced changes to submersion time can have important implications for aquatic communities. Warmer, less oxygenated water may reduce foraging time and increase energy expended on surface visits. Aquatic beetles are more vulnerable to fish predation at the surface because they become temporarily inactive and are particularly visible to fish. Given projections of future climate warming, cascading effects of beetle behavioral changes on aquatic food webs can be expected.
References
Bates D, Maechler M, Bolker BM (2011) lme4: linear mixed-effects models using S4 classes. In, R package version 0.999375–42
Boersma KS, Bogan MT, Henrichs BA, Lytle DA (2014) Top predator removals have consistent effects on large species despite high environmental variability. Oikos 123:807–816
Boersma KS, Lytle DA (2014) Overland dispersal and drought-escape behavior in a flightless aquatic insect, Abedus herberti (Hemiptera: Belostomatidae). Southwest Nat 59:301–302
Bogan MT (2017) Hurry up and wait: life cycle and distribution of an intermittent stream specialist (Mesocapnia arizonensis). Freshw Sci 36:805–815
Calosi P (2012) The comparative biology of diving in two genera of European Dytiscidae (Coleoptera). J Evol Biol 25:329–341
Calosi P, Bilton DT, Spicer JI (2007) The diving response of a diving beetle: effects of temperature and acidification. J Zool 273:289–297
Calosi P, Bilton DT, Spicer JI (2008) Thermal tolerance, acclimatory capacity and vulnerability to global climate change. Biol Lett 4:99–102
Chaui-Berlinck JG, Bicudo JEPW, Monteiro LHA (2001) The oxygen gain of diving insects. Respir physiol 128:229–233
Contreras HL, Bradley TJ (2011) The effect of ambient humidity and metabolic rate on the gas-exchange pattern of the semi-aquatic insect Aquarius remigis. J Exp Biol 214:1086–1091
Cook BI, Smerdon JE, Seager R, Coats S (2014) Global warming and 21st century drying. Clim Dyn 43:2607–2627
Cover MR, Seo JH, Resh VH (2015) Life history, burrowing behavior, and distribution of Neohermes filicornis (Megaloptera: Corydalidae), a long-lived aquatic insect in intermittent streams. West North Am Nat 75:474–490
Crump ML (1983) Opportunistic cannibalism by amphibian larvae in temporary aquatic environments. Am Nat 121:281–289
De Ruiter L, Wolvekamp HP, van Tooren AJ (1949) Experiments on the respiration of some aquatic insects (Hydrous piceus L., Naucoris cimicoides L. and Notonecta glauca L.). Acta Physiol Pharmacol Neerlandica 1:657–659
Durfee RS, Knight Jasper S, Kondratieff BC (2005) Colorado Haliplidae (Coleoptera): biogeography and identification. J Kansas Entomol Soc 78:41–70
Gillooly JF, Brown JH, West GB, Savage VM, Charnov EL (2001) Effects of size and temperature on metabolic rate. Science 293:2248–2251
Greig HS, Wissinger SA, McIntosh AR (2013) Top-down control of prey increases with drying disturbance in ponds: a consequence of non-consumptive interactions? J Anim Ecol 82:598–607
Hickman JR (1931a) Contribution to the biology of the Haliplidae (Coleoptera). Ann Entomol Soc Am 24:129–142
Hickman JR (1931b) Respiration of the Haliplidae (Coleoptera). Papers of the Michigan Academy of Science, Arts and Letters 8:277–289
Kehl S, Dettner K (2009) Surviving submerged: setal tracheal gills for gas exchange in adult rheophilic diving beetles. J Morphol 270:1348–1355
Lake PS (2003) Ecological effects of perturbation by drought in flowing waters. Freshw Biol 48:1161–1172
Lenth RV (2016) Least-squares means: the R package lsmeans. J Stat Softw 69:1–33
Madsen BL (2012) Submersion respiration in small diving beetles (Dytiscidae). Aquat Insects 34:57–76
Mas-Martí E, García-Berthou E, Sabater S, Tomanova S, Munoz I (2010) Comparing fish assemblages and trophic ecology of permanent and intermittent reaches in a Mediterranean stream. Hydrobiologia 657:167–180
Matheson R (1912) The Haliplidæ of North America, North of Mexico. J N Y Entomol Soc 20:156–193
Merritt RW, Cummins KW, Berg MB (2008) An introduction to the aquatic Insects of North America. Kendell/Hunt, Dubuque, IA
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Roberts CH (1913) Critical notes on the species of Haliplidae of America north of Mexico with descriptions of new species. J N Y Entomol Soc 21:91–123
Ruff TW, Kushnir Y, Seager R (2011) Comparing twentieth- and twenty-first-century patterns of interannual precipitation variability over the western United States and northern Mexico. J Hydrometeorol 13:366–378
Seager R, Ting M, Held I, Kushnir Y, Lu J, Vecchi G, Huang H-P, Harnik N, Leetmaa A, Lau N-C, Li C, Velez J, Naik N (2007) Model projections of an imminent transition to a more arid climate in southwestern North America. Science 316:1181–1184
Seager R, Ting M, Li C, Naik N, Cook B, Nakamura J, Liu H (2013) Projections of declining surface-water availability for the southwestern United States. Nat Clim Chang 3:482–486
Seymour RS, Matthews PG (2013) Physical gills in diving insects and spiders: theory and experiment. J Exp Biol 216:164–170
Therneau T (2015) A package for Survival Analysis in S. Version 2.38
Verberk WCEP, Bilton DT (2014) Respiratory control in aquatic insects dictates their vulnerability to global warming. Biol Lett 9:20130473
Verberk WCEP, Bilton DT, Calosi P, Spicer JI (2011) Oxygen supply in aquatic ectotherms: partial pressure and solubility together explain biodiversity and size patterns. Ecology 92:1565–1572
Wańkowski JWJ, Thorpe JE (1979) Spatial distribution and feeding in Atlantic salmon, Salmo salar L. juveniles. J Fish Biol 14:239–247
Acknowledgements
We are grateful to S Brudzinski, L Patrick, and R Paulson for logistical support during this project and to MT Bogan for useful feedback on an earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Banks, T.B., Kincaid, R.M. & Boersma, K.S. Temperature and Dissolved Oxygen Determine Submersion Time in Aquatic Beetle Peltodytes callosus (Coleoptera: Haliplidae). J Insect Behav 31, 427–435 (2018). https://doi.org/10.1007/s10905-018-9689-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-018-9689-6