
Abstract
The redbay ambrosia beetle, Xyleborus glabratus, is an important pest of redbay (Persea borbonia) and swamp bay (P. palustris) trees in forests of the southeastern USA. It is also a threat to commercially grown avocado. The beetle is attracted to host wood volatiles, particularly sesquiterpenes. Contrary to other ambrosia beetles that attack stressed, possibly pathogen-infected, and dying trees, X. glabratus readily attacks healthy trees. To date little is known about the role of leaf volatiles in the host selection behavior and ecology of X. glabratus. To address this question, an olfactometer bioassay was developed to test the behavioral response of X. glabratus to plant leaf volatiles. We found that X. glabratus was attracted to the leaf odors of their hosts, redbay and swamp bay, with no attraction to a non-host tree tested (live oak, Quercus virginiana), which served as a negative control. Gas chromatography–mass spectrometry (GS/MS) analysis of leaves revealed the absence of sesquiterpenes known to be attractive to X. glabratus and present in host wood, suggesting that additional leaf-derived semiochemicals may serve as attractants for this beetle. An artificial blend of chemicals was developed based on GC/MS analyses of leaf volatiles and behavioral assays. This blend was attractive to X. glabratus at a level that rivaled currently used lures for practical monitoring of this pest. This synthetic redbay leaf blend also was tested in the field. Baited traps captured more X. glabratus than unbaited controls and equivalently to manuka oil lures. We hypothesize that leaf volatiles may be used by X. glabratus as an additional cue for host location.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The exotic redbay ambrosia beetle, Xyleborus glabratus Eichhoff (Coleoptera: Curculionidae: Scolytinae), is an invasive species, native to Asia and established in the southeastern United States (Fraedrich et al. 2008; Rabaglia et al. 2006). Xyleborus glabratus is the vector of the fungal pathogen Raffaelea lauricola T.C. Harr., Fraedrich and Aghayeva, that causes laurel wilt, a highly lethal disease of the Lauraceae (Fraedrich et al. 2008; Harrington et al. 2008). Boring by X. glabratus inoculates host trees with R. lauricola, which, in susceptible hosts, is followed by branch wilt that progresses throughout the entire canopy, ultimately leading to tree death (Fraedrich et al. 2008; Mayfield et al. 2008). Wild and urban populations of redbay (Persea borbonia [L.] Spreng.) and swamp bay (Persea palustris [Raf.] Sarg.) have been killed by laurel wilt (Evans et al. 2013; Fraedrich et al. 2008; Shields et al. 2011; Spiegel and Leege 2013), and practical concerns are significant, given that the full impacts of this disease upon the Florida Everglades and commercial avocado (Persea americana Mill.) growing regions of south Florida have not yet been realized (Ploetz et al. 2013; Rodgers et al. 2014).
Xyleborus glabaratus is restricted to lauraceous hosts within the US (Hanula et al. 2008; Kendra et al. 2014a; Mayfield et al. 2013; Peña et al. 2012); these also are preferred in its native host range (Hulcr and Lou 2013). Contrary to most ambrosia beetles that attack weakened, damaged or recently dead trees (Hulcr et al. 2007; Lindgren 1990), X. glabratus can attack live and apparently healthy trees within its introduced range in the US (Fraedrich et al. 2008; Mayfield et al. 2008). Additionally, X. glabratus is not attracted to ethanol (Hanula and Sullivan 2008; Johnson et al. 2014), a semiochemical indicative of tree stress and decay (Kelsey et al. 2014; Kimmerer and Kozlowski 1982) that is used as an attractant for monitoring of various ambrosia beetles and wood borers of the Xyleborini tribe (Miller and Rabaglia 2009; Montgomery and Wargo 1983; Ranger et al. 2010). Extensive research has shown that X. glabratus is attracted to the sequiterpenes found within the cambium of their lauraceous hosts (Niogret et al. 2011); primarily α-copaene, but also to α-cubebene, α-humulene, and calamenene (Hanula and Sullivan 2008; Kendra et al. 2012, 2014a; Niogret et al. 2011). This led to the widespread use of manuka oil, an essential oil containing high concentration of α-copaene as well as cubeb oil as the primary attractants for trapping and surveying X. glabratus for quarantine and management purposes (Hughes et al. 2015; Johnson et al. 2014).
Thus far, the response of X. glabratus to chemical host cues has been tested with the use of cut tree bolts, synthetic terpenoids, and essential oil lures (Hanula and Sullivan 2008l; Kendra et al. 2014a,b; Kuhns et al. 2014a; Mayfield et al. 2013). Leaves represent a major tissue source of volatile emission from plants (Baldwin 2010), yet little has been done to explore what role (if any) host leaves and their volatiles may play in the ecology of X. glabratus. The purpose of this investigation was to determine the possible attractiveness of host and non-host leaf volatiles to X. glabratus. Behavioral assays were conducted by exposing X. glabratus to leaf odors in laboratory olfactometers, which indicated beetle attraction to host leaf volatiles. In addition, the volatile profile of a host plant (redbay) leaf headspace was analyzed by GC/MS and compared to redbay wood volatiles. The leaf chemical profile was reconstituted in vitro, creating an artificial ‘redbay leaf blend’ that was later tested in laboratory and field experiments. This synthetic blend, based on identified leaf volatiles, proved to be attractive to X. glabratus in the laboratory and field.
Methods and Material
Insect and Plants
Xyleborus glabratus beetles were reared and emerged from infested swamp bay logs collected in Wekiwa, FL, USA. The logs were stored at 23 ° C in large plastic containers with humidified Kimwipe® papers (Kimberly-Clark, Roswell, GA, USA) that were replaced every 2 weeks. Beetles were collected, observed under a dissection microscope to ensure mobility (no missing legs and able to walk) 1–2 hr prior to olfactometer assays. Plant material consisted of clonally propagated redbays planted in 57 L containers (Hughes and Smith 2014). Containerized (12.5 L) nursery grown swamp bays and live oak branches (Quercus virginiana Mill.) collected near Lake Alfred, FL, USA also were investigated.
Chemicals
Dichloromethane (99.8 % purity), nonyl acetate (99 %), β Caryophyllene (80 %), β-pinene (99 %), camphor, p-cymene (99 %), α-pinene (98 %), limonene (90 %), sabinene (75 %), eucalyptol (99 %), borneol, terpinen-4-ol (95 %), α-terpineol (96 %), bornyl acetate (98 %), and α-terpinyl acetate (90 %) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Manuka oil was purchased from East Cape Manuka oil (Meridian, ID, USA).
Olfactometer System
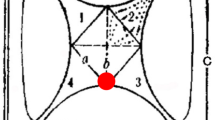
A four-choice olfactometer (Vet et al. 1983) (Analytical Research System, Gainesville, FL, USA) was used to evaluate the behavioral response of X. glabratus. The olfactometer consisted of a four-armed star-shape of four crescents within a 30 × 30 cm Teflon square (Vet et al. 1983). Each arm of the star runs into a 15 mm (internal diam [ID]) Teflon tube (Fig. 1). Four odor fields were created in the chamber by a constant airflow of 0.1 L/min pushed through each arm of the olfactometer and by pulling air (0.4 L/min) out through the floor’s central air evacuation hole (Fig. 1). The olfactometer floor and arms were covered with filter paper (25 cm diam laboratory filter paper, Curtin Matheson Scientific, Houston, TX, USA) to improve beetle traction and movement. The air evacuation hole was covered with Teflon fabric to prevent beetles from entering.

Graphic representation of a 4-way olfactometer. a glass capture trap, b vacuum air evacuation port, c odor fields, with different colors/shades representing different treatments. Arrows indicate direction of airflow
Between each bioassay, the filter paper was changed, and the olfactometer was washed with Sparkleen® detergent (Fisherbrand, Pittsburgh, PA, USA) and acetone. Each arm of the olfactometer was connected to the air delivery system through a two-way opened 350 ml glass vial that served as a collection trap for beetles choosing an arm/odor (Fig. 1). To ensure a chemical-free ambient air supply, arms of the olfactometer received charcoal purified air from a custom made air delivery system (ARS, Gainesville, FL, USA). The airflow was measured with a flowmeter (Varian, Walnut Creek, CA, USA) to ensure equivalent velocity within each arm. The olfactometer was positioned under a 150 W high-pressure sodium grow light (Hydrofarm, Petaluma, CA, USA). Twenty-five X. glabratus adult females were released into the center of the olfactometer, which was covered with a Plexiglas sheet and black filter paper so that only the glass traps were illuminated. Beetles were introduced into the olfactometer between 16:00 and 17:00 hr, and the number of beetles that entered the arms and fell into each trap was counted 16 hr later. This bioassay was performed overnight given that peak activity for X. glabratus is between 17:00 and 19:00 hr (Brar et al. 2012; Kendra et al. 2012). Beetles that did not leave the olfactometer arena were designated as non-responders (NR). Preliminary negative control tests were conducted by running bioassays without odors (humidified air only), which supported the assumption that beetles distributed randomly among the four traps.
Xyleborus Glabratus Response to Leaf Volatiles
For this experiment, beetles were tested in the 4-choice olfactometer as described above. Identical odor sources were randomly assigned to two opposing arms of the olfactometer for each treatment and, therefore, only two treatments were compared simultaneously. Odor sources consisted of undamaged redbay leaves that were enclosed within two-port glass domes (38 cm height, 14.4 cm ID). Each plant was inserted into a 1 cm diam hole within a 2.5 cm width polytetrafluoroethylene (PTFE) board (thereafter referred as ‘guillotine’). The guillotine can be opened so that the plant can be introduced within the hole without damage. The guillotine was used to separate the upper and lower portions of the tree canopy. With this procedure, a known number of leaves were present within each glass dome. Clean air was pushed through a water-filled bubbler, to humidify it, into the glass dome (with leaves enclosed within), and finally into the olfactometer at 0.1 L/min. Air was pulled from the olfactometer’s central evacuation port by a vacuum pump at 0.4 L/min to maintain a constant air stream. Laboratory conditions were maintained at 23 ± 1 C°, 49 % RH and a L14:D10 photoperiod. The response of X. glabratus to redbay, swamp bay, and live oak leaf volatiles (plants described above) was tested. Redbay and swamp bay are considered optimal hosts for X. glabratus; whereas, live oak has already been described as a non-host to X. glabratus (Kendra et al. 2014a; Mayfield et al. 2008) and, therefore, was used as a negative control. Each leaf volatile treatment was tested with three replicate trees and up to 25 beetles per assay (≈75 beetles tested in total).
GC/MS Analysis of Leaf Volatiles
A volatile collection system was used to identify the profile of redbay leaf volatile odors. It consisted of four parallel glass domes (38 cm height, 23 cm ID) each with two 3 cm outlets, one at the top, connecting to the incoming airflow, and the other at the bottom, connecting to the vacuum. Each glass dome was positioned onto a 5 cm PTFE guillotine so that each plant was separated into two parts in terms of headspace collection. A volatile collection trap (7.5 cm long) with 30 mg of HayeSep Q adsorbent (Volatile Assay Systems, Rensselaer, NY, USA) was connected to the bottom outlet with a PTFE fitting. Volatiles emitted from the upper portion of each plant enclosed within each glass chamber were swept downward by the incoming humidified and charcoa1 filter purified air at a rate of 1.0 L/min. The volatiles were forced to the bottom of the chamber by pulling air at 0.6 L/min through volatile collection traps with a controlled vacuum from the automated volatile collection system.
Volatiles also were collected from the undamaged trunks of redbay trees in a separate experiment. Ten cm of the main stem (2.5 cm diam) was enclosed within an oven bag (Reynolds, Lake Forest, IL, USA) and tied at the top and bottom with rubber bands. Air was pulled at a rate of 1.0 L/min and sampled at the bottom of the bag by pulling it at a rate of 0.6 L/min through volatile collection traps during a 24 hr collection period.
Finally, redbay wood volatiles also were collected by rasping 2 g of stem tissue (bark, cambium, and sapwood) and placing this material within a 20 cm glass tube. Air was pushed at a rate of 1.0 L/min and pulled at 0.6 L/min through volatile collection traps for a 15 m collection period.
Volatiles were extracted from the collection traps by washing with 150 μl of dichloromethane. Nonyl acetate (1080 ng) was added as an interna1 standard to the extracts. For each collection sample, 1 μl was manually injected into a Clarus 500 GC/MS (PerkinElmer, Shelton, CT, USA). The gas chromatograph was equipped with a column capillary injector system and flame ionization detector. Data collection, storage, and subsequent analysis were performed on Perkin Elmer chromatographic data system TurboMass™. Helium at a linear flow velocity of 2 ml/min was used as the carrier gas. All samples were analyzed on a fused silica RTX-5 capillary column (Restek Corporation, Bellefonte, PA, USA), 60 min × 0.25 mm ID. The temperature of the column oven was maintained at 40 °C for 1 min and then increased at a rate of 7 °C/min to a final temperature of 300 °C and maintained at 300 °C for 6 min. The injector temperature was set at 270 °C with the detector set at 200 °C. Quantitations were based on GC/MS profiles and were assigned by comparing peak areas of known amounts of nonyl acetate (1080 ng) with the peak areas of compounds extracted from the leaves. Constituents of the plant volatile emissions were identified by comparison of mass spectra with spectra in the National Institute of Standards and Technology database, and the spectra obtained from authentic reference compounds, when available. Additionally, GC retention times of plant volatiles were compared with those of authentic compounds on the RTX-5 column, when available.
Xyleborus Glabratus Response to Synthetic Volatiles
The behavioral response of X. glabratus to synthetic volatiles was tested based on the above GC/MS analyses. Test compounds were dissolved in 100 μl of dichloromethane at a 0.1 μg/μl dosage rate and pipetted onto 2 cm Richmond cotton wicks (Petty John Packaging, Inc. Concord, NC, USA). These release devices were placed into two opposing glass olfactometer traps, as described above. Manuka oil was chosen as a positive control due to its known attractiveness to X. glabratus (Hanula and Sullivan 2008). Five replicate bioassays of up to 25 beetles (total of = 125 beetles) were performed for each treatment. The response of X. glabratus was tested in the four-choice olfactometer system to: (1) manuka oil vs. solvent (dichloromethane), (2) redbay leaf blend (a mix of synthetic volatiles representing 94.10 % of the redbay leaf volatiles found after GC/MS analysis [Table 1]) vs. solvent, and (3) the redbay leaf blend vs. manuka oil.
Test of Redbay Leaf Blend Under Field Conditions
The field trapping site consisted of a hardwood hammock bordering the Kanapaha Botanical Garden (29°36′41.0″N 82°24′35.8″W) in Gainesville, Florida. This site was chosen because X. glabratus was known to be abundant, and infested redbays occurred in various stages of laurel wilt decline, as well as, apparently healthy (asymptomatic) trees. Other abundant tree species within the area included: live oak, bluff oak (Quercus astrina Small), sweet gum (Liquidambar styraciflua L.), and American holly (Ilex opaca Aiton). Laurel wilt was present on the site for approximately 2 years (Adam Black, personal communication) when this field experiment was initiated (August 2014). Traps were constructed from a 1.5 m wooden post with a 30 cm Plexiglas® square affixed to the upper portion. An Elm Bark Beetle trap (46 × 64 cm) (Great Lakes IPM Inc.) then was folded over the Plexiglas® square and held with binder clips, resulting in a double-sided sticky-panel (46 × 32 cm). Traps were deployed 10 m from the closest redbay tree, at minimum, to avoid bias due to potential mass emergence of X. glabratus. Lures consisted of 7 ml BEEM vials (Thermo Fisher Scientific, Waltham, MA, USA) filled with 1 ml of the redbay leaf odor blend without solvent affixed with twist-ties to the upper-center of the sticky, trapping panel. Immediately prior to deployment, four holes were poked into the vials with a pushpin to facilitate release of volatiles. Traps were baited with one of three treatments: water (negative control), 1 ml of manuka oil without solvent (positive control), and 1 ml of the redbay leaf blend lure. Three posts/traps, baited with their respective treatment lures were spaced 10 m apart, representing a block. Each block was separated by at least 50 m, and 3–5 blocks were deployed per trapping period. The total number of X. glabratus captured was recorded during 7 days trapping periods. Three trials were conducted: August 25 through September 1, 2014 (N = 4); September 8 through September 15, 2014 (N = 5), and October 6 through October 13, 2014 (N = 3). Four blocks were similar during the three trials, and the treatments were rotated within these blocks for each trial. In one case, an entire block was removed from the analysis to maintain a balanced design because a single trap was found on the ground.
Statistical Analysis
To analyze olfactometer data, a chi-squared test on the pooled values of the different replicates was performed. Beforehand, a heterogeneity chi-squared test was conducted to ensure that data from each replicate were homogenous (Zar 2009). Xyleborus glabratus response data obtained per replicate were found to be homogenous if the sum of the individual chi-squares for each replicate was not significantly different (α > 0.05) from the overall chi-squared of the pooled data (Zar 2009). For the field trapping experiments, capture data were log transformed to account for a non-normal distribution and were analyzed with a linear mixed model with Gaussian distribution. The fixed variable was the treatment lure and the random variable was the block number. Pairwise paired t-test with Bonferonni correction was used to determine differences among treatments.
Results
Xyleborus glabratus Response to Leaf Volatiles. Preliminary experiments indicated no bias in the response of X. glabratus among the four unbaited olfactometer arms receiving humidified air (χ 2 = 2.03, df = 3, P > 0.05). Xyleborus glabratus was significantly attracted to the leaf volatiles from its redbay and swamp bay host plants (Fig. 2). The heterogeneity test performed on redbay was significant (χ 2 = 17.94, df = 2, P < 0.001) indicating that the three replicates for this treatment were not homogeneous. Xyleborus glabratus were highly attracted toward redbay leaf volatiles on two replicates (P < 0.001), but there was more variation on the third one (P > 0.05). However, the overall chi-squared tests performed on pooled data revealed that redbay (χ 2 = 12.65, df = 1, P < 0.001) and swamp bay leaf volatiles (χ 2 = 22.44, df = 1, P < 0.001) were more attractive to X. glabratus than clean air. In contrast, leaf volatiles from the non-host tree, live oak, were not attractive to X. glabratus, as compared with the clean air control (Fig. 2) (χ 2 = 0.25, df = 1, P > 0.05).

Percentage of Xyleborus glabratus responding to natural or synthetic odorants vs. humidified air or solvent (dichloromethane) negative controls, within a four-choice olfactometer. N total number of X. glabratus used during the experiments, NR Percent of non-responders. Asterisks indicate significant differences between the two treatments (*** = P < 0.001)
GC/MS Analysis of Leaf Volatiles
The leaf volatiles of eight potted redbay trees were examined to identify compounds that may be attractive to X. glabratus (Fig. 3a). Most notably, the sesquiterpenes found in redbay wood and that are known to be attractive to X. glabratus were absent from leaf volatile emission profiles (Table 1). To ensure that our collection and extraction method allowed for the detection of those compounds, a GC/MS analysis of rasped wood samples of the same redbay individuals was performed using HayeSep Q according to Niogret et al. (2011), but with dichloromethane as a solvent. Our tests were able to detect attractive sesquiterpenes (α-copaene and calamenene) from the rasped wood, thus validating that they were undetectable or absent from the redbay leaf emissions (Fig. 3c). The volatile profiles obtained from the bagging of undamaged redbay stems was similar to that of an empty oven bag (blank control), indicating that volatiles emitted from the stem tissue were undetectable by our method (data not shown) and that rasping/shaving of the tissue was necessary to trap wood volatiles.

GC/MS Profiles. a Representative GC/MS profile from redbay leaves after 24 hr of collection. b redbay leaf blend that consists of a reconstitution of the redbay leaf volatile profile with a mixture of synthetic chemicals. c redbay cambium/wood volatiles after 15 min of collection. a: α-Pinene, b: Sabinene, c: β-Pinene, d: Myrcene, e: Cymene, f: Limonene, g: Eucalyptol, h: Camphor, i: Borneol, j: Terpinen-4-ol, k; α-Terpineol, l: Bornyl acetate, m: α-Terpinyl acetate, n: α-Copaene*, o: α-Bergamotene*, p: β Caryophyllene, q: unidentified sesquiterpene, r: Calamenene*. * Determined with NIST database only
Xyleborus Glabratus Response to Synthetic Volatiles
The olfactometer bioassays revealed that diluted manuka oil was attractive to X. glabratus when compared to the dichloromethane solvent (χ 2 = 18.89, df = 1, P < 0.001) (Fig. 2), validating the effectiveness of the bioassay. Xyleborus glabratus also preferentially chose the redbay leaf blend (χ 2 = 10.18, df = 1, P = 0.001) as compared to the solvent negative control (Fig. 2), with 65 % of beetles migrating to traps with the redbay leaf blend. Finally, there was no statistical preference between the redbay leaf blend and manuka oil (χ 2 = 1.13, df = 1, P > 0.05) at the dosage tested, although manuka oil captured slightly more beetles (Fig. 2).
Test of Redbay Leaf Blend Under Field Conditions
The redbay leaf blend was similar to the leaf volatile odors collected from redbay trees (Figs. 3a and b). There were differences in X. glabratus captures between the three treatment lures (χ 2 = 19.98, df = 2, P < 0.001) (Fig. 4). Traps baited with manuka oil or the redbay leaf blend captured more X. glabratus than the negative controls (P < 0.001 and P = 0.032, respectively). Although traps baited with manuka oil caught the most beetles, captures were statistically similar to those obtained with the redbay leaf blend (P > 0.05) (Fig. 4).

Log number of Xyleborus glabratus captured on sticky traps baited with various lures during field trapping trials. Different letters indicate significant differences between treatments at α < 0.05
Discussion
The olfactometer bioassays and field trapping experiments demonstrated that X. glabratus is attracted to redbay and swamp bay leaf volatiles, indicating that they may act as an additional cue for locating hosts by X. glabratus. The lack of attraction to live oak samples suggests that X. glabratus can distinguish between host and non-host leaf volatiles within our olfactometer bioassays. Avoidance of non-host volatiles is known to occur among some scolytid bark beetles, and synthetic blends derived from these non-hosts also can inhibit response to aggregation pheromones, resulting in possible management options for the protection of forest trees (Byers et al. 2000; Unelius et al. 2014; Zhang and Schlyter 2004). In this case, we did not observe avoidance of the non-host tested, but an absence of response (i.e., humidified air was not preferred over live oak).
Chemical analysis of attached and undamaged leaves revealed the presence of eleven compounds, which were mostly monoterpenes. The vast majority of the identified compounds in leaf and wood redbay volatiles were chiral, however, the enantiomeric composition of these volatiles was not determined in this study. Given that stereochemical properties of enantiomers impacts odor and other biological activities, such as insect behavior (Brenna et al. 2003; Mori 2014), enantiomeric compositions of wood and leaf volatiles of Laureacea coupled with behavioral bioassays should be conducted to determine if specific enantiomers are more effective attractants for X. glabratus. Nonetheless, our field trapping data proved that the blend of available synthetic chemicals tested here was attractive to X. glabratus. The volatile profile of leaves differed from that derived from freshly rasped redbay wood and lacked sesquiterpenes known to be attractive to X. glabratus such as α- copaene and calamenene. The attractiveness of the leaf volatiles in the absence of wood-derived terpenes was confirmed by in vitro reconstitution of the volatile blend, which was found to be attractive both in laboratory olfactometer and field tests. To assess the proximo-distal distributions of avocado (Persea americana Mill.) terpenes, Niogret et al. (2013) sampled leaf, branch and trunk tissues. In contrast to our results, the authors detected α-copaene and several other sesquiterpenes among all tissue types sampled, including leaves. They also noted a distinct gradient in the abundances of these sesquiterpenes from trunk (highest) to leaves (lowest), while the opposite relationship was seen for monoterpenes. Similarly, in our work, redbay leaves contained mostly monoterpenes but no sesquiterpenes, while the wood tissue also had higher sesquiterpene content. The chemical differences between our results and that of Niogret et al. (2013) may be related to the phylogenetic separation between the two plant species tested. Redbay is within the subgenus Eriodaphne and avocado in Persea (Scora and Bergh 1992). Additionally, the size of the plants or solvents utilized could have contributed to the differences found. Major differences also existed in the condition of the tissue used; while we collected leaf volatiles from undamaged leaves, Niogret et al. (2013) sectioned and cut the avocado leaves and rasped the petioles, a procedure that may have released terpenes bound within the plant’s cells, in a similar manner to herbivory (Baldwin 2010). Interestingly, all the chemicals that we found in redbay leaf volatiles also were found in the redbay wood volatiles (Fig. 3), suggesting that the leaf volatile profile represents the monoterpene fraction of the wood volatile headspace.
Some monoterpenes are known to be attractive to bark and ambrosia beetles. α-Pinene, for instance, is widely used alone or as part of blends to attract a wide range of bark beetles feeding on gymnosperms (Duduman 2014; Miller et al. 2013; Miller and Rabaglia 2009). An increase of monoterpene concentrations such as limonene, α-pinene, β-pinene, or myrcene in the phloem and the sapwood of the Aleppo pine (Pinus halepensis Mill.) has been correlated with higher infestation by the Mediterranean pine shoot beetle (Tomicus destruens Wollaston) (Kelsey et al. 2014). The attractiveness of redbay leaves and the synthetic blend developed here may be due to the presence of eucalyptol, which is a major component of the wood volatile profile of redbay and California bay laurel (Umbellularia californica [Hook. & Arn.] Nutt.). (Kendra et al. 2014a; Kuhns et al. 2014a). Eucalyptol is attractive to X. glabratus and in electroantennographic tests elicited a strong response from X. glabratus (Kendra et al. 2014a). In large quantities, this single chemical attracted X. glabratus in the field (Kuhns et al. 2014a). Another major peak that we found in undamaged redbay leaf volatiles was camphor. Camphor is a significant volatile constituent of camphortree (Cinnamomum camphora L.) wood (Li et al. 2014), which has been found to be more attractive to X. glabratus than swamp bay, redbay, and avocado bolts in field tests (Kendra et al. 2014a; Mayfield et al. 2013). The positive control used in this study was manuka oil. Manuka lures have been used since 2008 as a tool for X. glabratus detection (Hughes et al. 2015). However, commercially available manuka lures are short-lived in activity as attractants for X. glabratus compared to newly developed cubeb oil lures (the efficiency of Manuka lures declines after 3 weeks of activity) (Hanula et al. 2013. Kendra et al. 2015). Therefore, further investigation of cubeb oil as a positive control on a longer experimental period should be conducted to evaluate the potential of our redbay leaf volatile blend for monitoring of X. glabratus.
Xyleborus Glabratus
potentially uses leaf volatile cues after emergence, when the adult females attempt to locate host trees during their dispersal flights (Hughes et al. 2015; Maner et al. 2013). Leaf volatiles potentially could be used by X. glabratus as a long-range cue, or a cue indicating the presence of redbay in the foraging area. Following this long-range attraction through leaf volatiles, tree selection might be mediated through visual rather than olfactory cues, as X. glabratus are known to be attracted by artificial stems of larger diameter (Mayfield and Brownie 2013). Additionally, bark requires damage for volatile release, and once on the tree, the beetles will subsequently determine if the host is suitable for colonization and reproduction, guiding the decision to bore into the wood (Kendra et al. 2014a; Kuhns et al. 2014a). Additional experiments are needed to investigate the potential interactions between leaf volatiles and other known attractant cues for X. glabratus such as the odors from its symbiotic fungus (Hulcr et al. 2011; Kuhns et al. 2014b), as well as, other possible microorganisms.
It is well established that leaf volatiles change qualitatively and quantitatively in response to pathogen infection or herbivory (McLeod et al. 2005; Ponzio et al. 2013; Turlings and Wäckers 2004), and observations suggest that more attacks occur on moribund laurel wilt affected trees by X. glabratus than on uninfected counterparts (Hughes et al. 2015). An intriguing question is whether X. glabratus responds to changes in host volatile composition as a result of pathogen infection in the same manner as has been observed for the Dutch elm disease pathosystem (McLeod et al. 2005). In this case, native elm bark beetles (Hylurgopinus rufipes Eichhoff) are significantly more attracted to infected than uninfected trees. It is possible that leaf volatile profiles may be modified by beetle infestation or R. lauricola infection and that these changes may affect host plant selection preferences by mobile X. glabratus females.
References
Baldwin IT (2010) Plant volatiles. Curr Biol 20:R392–R397
Brar GS, Capinera JL, McLean S, Kendra PE, Ploetz RC, Peña JE (2012) Effect of trap size, trap height and age of lure on sampling xyleborus glabratus (coleoptera: curculionidae: scolytinae), and its flight periodicity and seasonality. Fla Entomol 95:1003–1011
Brenna E, Fuganti C, Stefano S (2003) Enantioselective perception of chiral odorants. Tetrahedron Asymmetry 14:1–42
Byers JA, Zhang QH, Birgersson G (2000) Strategies of a bark beetle, pityogenes bidentatus, in an olfactory landscape. Naturwissenschaften 87:503–507
Duduman ML (2014) Field response of the northern spruce bark beetle Ips duplicatus (sahlberg) (coleoptera: curculionidae, scolytinae) to different combinations of synthetic pheromone with (−) ‐α‐pinene and (+)-limonene. Agric For Entomol 16:102–109
Evans JP, Scheffers BR, Hess M (2013) Effect of laurel wilt invasion on redbay populations in a maritime forest community. Biol Invasions 16:1581–1588
Fraedrich SW, Harrington TC, Rabaglia RJ, Ulyshen MD, Mayfield AE, Hanula JL, Eickwort JM, Miller DR (2008) A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other lauraceae in the southeastern United States. Plant Dis 92:215–224
Hanula JL, Sullivan B (2008) Manuka oil and phoebe oil are attractive baits for xyleborus glabratus (coleoptera : scolytinae), the vector of laurel wilt. Environ Entomol 37:1403–1409
Hanula JL, Mayfield AE, Fraedrich SW, Rabaglia RJ (2008) Biology and host associations of redbay ambrosia beetle (coleoptera: curculionidae: scolytinae), exotic vector of laurel wilt killing redbay trees in the southeastern United States. J Econ Entomol 101:1276–1286
Hanula JL, Sullivan BT, Wakarchuk D (2013) Variation in manuka oil lure efficacy for capturing xyleborus glabratus (coleoptera: curculionidae: scolytinae), and cubeb oil as an alternative attractant. Environ Entomol 42:333–340
Harrington TC, Fraedrich SW, Aghayeva DN (2008) Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the lauraceae. Mycotaxon 104:399–404
Hughes MA, Smith JA (2014) Vegetative propagation of putatively laurel wilt-resistant redbay (persea borbonia). Native Plants J 15:42–50
Hughes MA, Smith J, Ploetz R, Kendra P, Mayfield AE, Hanula J, Hulcr J, Stelinski L, Cameron S, Riggins J, Carrillo D, Rabaglia B, Eickwort J, Pernas T (2015) Recovery plan for laurel wilt on redbay and other forest species caused by raffaelea lauricola and disseminated by xyleborus glabratus. National Plant Disease Recovery System, Homeland Security Presidential Directive Number 9 (HSPD-9)
Hulcr J, Lou Q-Z (2013) The redbay ambrosia beetle (coleoptera: curculionidae) prefers lauraceae in its native range: records from the Chinese national insect collection. Fla Entomol 96:1595–1596
Hulcr J, Mogia M, Isua B, Novotny V (2007) Host specificity of ambrosia and bark beetles (col: curculionidae: scolytinae and platypodinae) in a New Guinea rainforest. Ecol Entomol 32:762–772
Hulcr J, Mann R, Stelinski LL (2011) The scent of a partner: ambrosia beetles are attracted to volatiles from their fungal symbionts. J Chem Ecol 37:1374–1377
Johnson C, Cameron R, Hanula J, Bates C (2014) The attractiveness of manuka oil and ethanol, alone and in combination, to xyleborus glabratus (coleoptera: curculionidae: scolytinae) and other curculionidae. Fla Entomol 97:861–864
Kelsey RG, Gallego D, Sánchez-García FJ, Pajares JA (2014) Ethanol accumulation during severe drought may signal tree vulnerability to detection and attack by bark beetles. Can J For Res 44:554–561
Kendra PE, Montgomery WS, Niogret J, Deyrup MA, Guillén L, Epsky ND (2012) Xyleborus glabratus, X. affinis, and X. ferrugineus (coleoptera: curculionidae: scolytinae): electroantennogram responses to host-based attractants and temporal patterns in host-seeking flight. Environ Entomol 41:1597–1605
Kendra PE, Montgomery WS, Niogret J, Pruett GE, Mayfield AE, MacKenzie M, Deyrup MA, Bauchman GR, Ploetz RC, Epsky ND (2014a) North American lauraceae: terpenoid emissions, relative attraction and boring preferences of redbay ambrosia beetle, xyleborus glabratus (coleoptera: curculionidae: scolytinae). PLoS One 9, e102086
Kendra PE, Montgomery WS, Niogret J, Schnell EQ, Deyrup MA, Epsky ND (2014b) Evaluation of seven essential oils identifies cubeb oil as most effective attractant for detection of xyleborus glabratus. J Pest Sci 87:681–689
Kendra PE, Niogret J, Montgomery WS, Deyrup MA, Epsky ND (2015) Cubeb oil lures: terpenoid emissions, trapping efficacy, and longevity for attraction of redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae). J Econo Entomol. doi:10.1093/jee/tou023
Kimmerer TW, Kozlowski TT (1982) Ethylene, ethane, acetaldehyde, and ethanol production by plants under stress. Plant Physiol 69:840–847
Kuhns EH, Martini X, Tribuiani Y, Coy M, Gibbard C, Peña J, Hulcr J, Stelinski LL (2014a) Eucalyptol is an attractant of the redbay ambrosia beetle, xyleborus glabratus. J Chem Ecol 40:355–362
Kuhns EH, Tribuiani Y, Martini X, Meyer WL, Peña J, Hulcr J, Stelinski LL (2014b) Volatiles from the symbiotic fungus raffaelea lauricola are synergistic with manuka lures for increased capture of the redbay ambrosia beetle xyleborus glabratus. Agric For Entomol 16:87–94
Li Q, Wang XX, Lin JG, Liu J, Jiang MS, Mao-Sheng J, Chu LX (2014) Chemical composition and antifungal activity of extracts from the xylem of cinnamomum camphora. Bioresources 9:2560–2571
Lindgren BS (1990) Ambrosia beetles. J For 88:8–11
Maner ML, Hanula JL, Braman SK (2013) Gallery productivity, emergence, and flight activity of the redbay ambrosia beetle (coleoptera: curculionidae: scolytinae). Environ Entomol 42:642–647
Mayfield AE III, Brownie C (2013) The redbay ambrosia beetle (coleoptera: curculionidae: scolytinae) uses stem silhouette diameter as a visual host-finding cue. Environ Entomol 42:743–50
Mayfield AE III, Peña JE, Crane JH, Smith JA, Branch CL, Ottoson ED, Hughes M (2008) Ability of the redbay ambrosia beetle (coleoptera: curculionidae: scolytinae) to bore into young avocado (lauraceae) plants and transmit the laurel wilt pathogen (raffaelea lauricola). Fla Entomol 91:485–487
Mayfield AE III, MacKenzie M, Cannon PG, Oak SW, Horn S, Hwang J, Kendra PE (2013) Suitability of California bay laurel and other species as hosts for the non-native redbay ambrosia beetle and granulate ambrosia beetle. Agric For Entomol 15:227–235
McLeod G, Gries R, von Reuß SH, Rahe JE, McIntosh R, König WA, Gries G (2005) The pathogen causing Dutch elm disease makes host trees attract insect vectors. Proc R Soc B 272:2499–2503
Miller DR, Rabaglia RJ (2009) Ethanol and (−)-alpha-pinene: attractant kairomones for bark and ambrosia beetles in the southeastern US. J Chem Ecol 35:435–48
Miller DR, Dodds KJ, Eglitis A, Fettig CJ, Hofstetter RW, Langor DW, Mayfield AE, Munson AS, Poland TM, Raffa KF (2013) Trap lure blend of pine volatiles and bark beetle pheromones for monochamus spp. (coleoptera: cerambycidae) in pine forests of canada and the United States. J Econ Entomol 106:1684–1692
Montgomery ME, Wargo PM (1983) Ethanol and other host-derived volatiles as attractants to beetles that bore into hardwoods. J Chem Ecol 9:181–90
Mori K (2014) Stereochemical studies on pheromonal communications. Proc Jpn Acad Ser B 90:373–388
Niogret J, Kendra PE, Epsky ND, Heath RR (2011) Comparative analysis of terpenoid emissions from Florida host trees of the redbay ambrosia beetle, xyleborus glabratus (coleoptera: curculionidae: scolytinae). Fla Entomol 94:1010–1017
Niogret J, Epsky ND, Schnell RJ, Boza EJ, Kendra PE, Heath RR (2013) Terpenoid variations within and among half-sibling avocado trees, persea americana mill. (lauraceae). PLoS One 8, e73601
Peña JE, Carrillo D, Duncan RE, Capinera JL, Brar G, McLean S, Arpaia ML, Focht E, Smith JA, Hughes M, Kendra PE (2012) Susceptibility of persea spp. and other lauraceae to attack by redbay ambrosia beetle, xyleborus glabratus (coleoptera : curculionidae: scolytinae). Fla Entomol 95:783–787
Ploetz RC, Hulcr J, Wingfield M, de Beer ZW (2013) Ambrosia and bark beetle-associated tree diseases: black swan events in tree pathology? Plant Dis 95:856–872
Ponzio C, Gols R, Pieterse CM, Dicke M (2013) Ecological and phytohormonal aspects of plant volatile emission in response to single and dual infestations with herbivores and phytopathogens. Funct Ecol 27:587–598
Rabaglia RJ, Dole SA, Cognato AI (2006) Review of American xyleborina (coleoptera: curculionidae: scolytinae) occurring north of mexico, with an illustrated key. Ann Entomol Soc Am 99:1034–1056
Ranger CM, Reding ME, Persad AB, Herms DA (2010) Ability of stress-related volatiles to attract and induce attacks by xylosandrus germanus and other ambrosia beetles. Agric For Entomol 12:177–185
Rodgers L, Derksen A, Pernas T (2014) Expansion and impact of laurel wilt in the Florida everglades. Fla Entomol 97:1247–1250
Scora R, Bergh B (1992) in Proceedings of Second World Avocado Congress, pp 505–514
Shields J, Jose S, Freeman J, Bunyan M, Celis G, Hagan D, Morgan M, Pieterson EC, Zak J (2011) Short-term impacts of laurel wilt on redbay (persea borbonia L. spreng.) in a mixed evergreen-deciduous forest in northern Florida. J For 109:82–88
Spiegel KS, Leege LM (2013) Impacts of laurel wilt disease on redbay (persea borbonia [L.] spreng.) population structure and forest communities in the coastal plain of Georgia, USA. Biol Invasions 15:2467–2487
Turlings TC, Wäckers F (2004) Recruitment of predators and parasitoids by herbivore-injuried plants. In: Carde RT, Millar GM (eds) Advance in insect chemical ecology. Cambridge University Press, Cambridge, UK, pp 21–75
Unelius CR, Schiebe C, Bohman B, Andersson MN, Schlyter F (2014) Non-host volatile blend optimization for forest protection against the European spruce bark beetle, Ips typographus. PLoS One 9, e85381
Vet LEM, van Lenteren JC, Hetmans M, Meelis E (1983) An airflow olfactometer for measuring olfactory responses of hymenopterous parasitoids and other small insects. Physiol Entomol 8:97–106
Zar J (2009) Biostatisical analysis, 5th edn. Prentice Hall, Upper Saddle River, NJ
Zhang Q-H, Schlyter F (2004) Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric For Entomol 6:1–20
Acknowledgments
We thank Adam Black for acquiring infested swamp bay logs for emergence; Kelsey Olson for help with Fig. 1; Angelique Hoyte, Laurie Martini, and Barry Fleming for help with behavioral assays. Funding was provided by the USDA Forest Service (Region 8) [Marc Hughes and Jason Smith] and USDA-APHIS grant (Cooperative Agreement 13-8212-0990-CA) [Lukas L. Stelinski].
Author information
Authors and Affiliations
Corresponding author
Additional information
Xavier Martini and Marc A. Hughes contributed equally to this work.
Rights and permissions
About this article

Cite this article
Martini, X., Hughes, M.A., Smith, J.A. et al. Attraction of Redbay Ambrosia Beetle, Xyleborus Glabratus, To Leaf Volatiles of its Host Plants in North America. J Chem Ecol 41, 613–621 (2015). https://doi.org/10.1007/s10886-015-0595-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-015-0595-5