
Abstract
Interest in forest regeneration has increased as secondary forests in regeneration process are cited as the forests of the future. However, remaining forests are subjected to chronic anthropogenic disturbances, which may reorganize tropical biodiversity. This paper investigates the recovery of fruit-feeding butterfly assemblages following slash-and-burn agriculture in the Caatinga dry forest, assessing the roles that chronic disturbances, tree species richness and biomass play in their taxonomic and functional recovery during secondary succession. Butterflies were recorded monthly across old-growth forest stands and areas previously subjected to slash-and-burn agriculture, and with differing times since abandonment. While there was a high degree of similarity in community-level attributes among regenerating forests of different ages, secondary and old-growth forests supported relatively distinct assemblages, both taxonomically and functionally. As expected, chronic disturbance, forest biomass and trees species richness were excellent predictors of taxonomic and functional changes. Specifically, large-bodied and canopy-inhabiting butterflies reached higher abundances in more disturbed habitats, while those ocellus-bearing species that fly in the understory and feed on monocot as larvae responded negatively to disturbance, but benefited from increased forest biomass and tree species richness. Our findings suggest that although regenerating secondary forests can recover values of community-level attributes of fruit-feeding butterflies similar to those of old-growth forests, chronic anthropogenic disturbances across the landscape may be selecting only specific ecological groups that are tolerant to adverse conditions.
Implications for insect conservation
Chronic disturbance can taxonomically and functionally alter butterfly assemblages across forest regeneration and consequently impact their ecological functions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tropical forests harbor significant amounts of global biodiversity and are responsible for providing essential ecosystem services for human well-being, such as climate regulation and the provision of forest products (Ferraz et al. 2014). However, human disturbances are reorganizing tropical biodiversity across all levels of ecological organization, from population to ecosystem level, with tangible impacts on the provision of ecosystem services (Filgueiras et al. 2021). Local disturbances are directly related to the conversion of old-growth forests into human-modified landscapes, through which a myriad of small forest remnants and secondary forest stands of varying ages (regenerating forests) remain embedded within open-habitat matrix, as agricultural lands continue to be abandoned (Melo et al. 2013). This is particularly the case for human-modified landscapes devoted to subsistence agriculture via slash-and-burn practices, the predominant land use across several tropical regions (Singh 1998; Laurance and Peres 2006). In this context, secondary forests have been frequently referred to as potential providers of ecosystem services and refugia of biodiversity (Chazdon et al. 2009), which has led to renewed research on forest regeneration and its drivers (see Arroyo-Rodriguez et al. 2015).
The role played by secondary forests in terms of biodiversity retention and provision of ecosystem services depends on the rate at which secondary forest stands approach the conditions found in old-growth stands; i.e. recovery rate (Norden et al. 2009; Poorter et al. 2016). In this perspective, forest resilience and successional trajectory have been proposed to respond to a myriad of forces operating at multiple scales; from local (e.g. soil degradation) to regional scale (e.g. available species pool) (Arroyo-Rodriguez et al. 2015). Overall, in the absence of severe degradation imposed by intensive land use (e.g. commercial agriculture and pastures), forest can recovery relatively fast (Guariguata and Ostertag 2001; Dunn 2004; Martin et al. 2013), with the regeneration marked by directional species replacements (Lohbeck et al. 2013). This is true for plant, amphibian and reptile assemblages (Hernández‐Ordóñez et al. 2015), as well as for bats (Avila-Cabadilla et al. 2009), while changes in insect communities are less predictable (but see Neves et al. 2010).
Dry forests exposed to slash-and-burn agriculture and chronic disturbances (extractivism of forest products) during regeneration may exhibit more simple or complex regeneration patterns, including site-dependent resilience and lack of directional species replacement as chronic disturbance favours resprouting species (Ceccon et al. 2006; Barros et al. 2021). Such a resprouting-driven resilience (Quesada et al. 2009; Álvarez-Yépiz et al. 2018) probably leads to the occurrence of a common set of host plants across forest regeneration with impacts for the organization of herbivore assemblages. Additionally, the continuous extraction of forest resources (e.g. firewood, fodder) during forest regeneration may result in more disturbed forest stands (i.e. higher light incidence and desiccation) and thus benefit disturbance-adapted or generalist plant and insect species (Ribeiro-Neto et al. 2016). Both the presence of resprouting species and chronic disturbances may interfere with patterns of forest resilience and successional trajectories, as has previously been documented for plants (Barros et al. 2021). Butterflies constitute a candidate group to examine how insect assemblages recover, or change throughout forest regeneration, since they comprise ecologically sensitive insects (Freitas et al. 2003, 2006), with a diverse taxa depending on plants for larval development and adult feeding.
The Caatinga, a dry forest in northeast Brazil, has been converted into human-modified landscapes by a combination of slash-and-burn and free-ranging livestock farming (Silva et al. 2017). Vegetation mosaics consisting of (1) old-growth forest, (2) regenerating forest stands of varying ages and (3) agricultural fields comprise most landscapes, and such landscapes are fundamental for the forest products based livelihoods of rural populations (Specht et al. 2019). Recent findings have described the Caatinga forest regeneration and dynamics as largely influenced by resprouting species (Barros et al. 2021) with plant assemblages dominated by disturbance-adapted plant species (Ribeiro and Freitas 2012; Rito et al. 2017a).
Here, we address the recovery rate and the successional trajectory exhibited by fruit-feeding butterfly assemblages as forest regenerate following slash-and-burn agriculture in the Caatinga dry forest. Additionally, we assess the role that chronic disturbances, tree species richness and biomass may play in their recovery during forest regeneration. To address these questions, we contrast a large set of community-level attributes, including ecological composition relative to microhabitat use, predation avoidance and feeding behaviour. Our findings are examined in the light of both patterns and drivers of forest regeneration/resilience in human-modified landscapes marked by intensive use of forest resources.
Material and methods
Study area
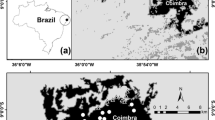
The study was carried out in Parque Nacional do Catimbau (Catimbau National Park), a protected area of 607 km2 located in the central region of the State of Pernambuco, northeastern Brazil (8° 24′ 00″ and 8° 36′ 35″ S; 37° 09′ 30″ and 37° 14′ 40″ W) (Fig. S1). The predominant climate in the region is semi-arid (Bsh), with transitions to rainy tropical (As), according to the Köppen scale. The annual rainfall varies from 480 to 1100 mm, with great irregularity in the interannual regime (Rito et al. 2017b). Generally, about 60–75% of annual rainfall occurs in the period from March/April to June/July (Rito et al. 2017b). The annual average temperature is about 23 °C, with July being the coldest month (average temperature of 21 °C), and December being the hottest month (average temperature of 25 °C) (Rito et al. 2017b). The vegetation consists of a mosaic of arboreal and shrub Caatinga (Sociedade Nordestina de Ecologia 2002).
Fruit-feeding butterfly surveys
The research was carried out in 10 plots of 0.1-ha, previously established in areas that had been used for slash-and-burn agriculture, with different ages since abandonment (forming a chronosequence of plots from 4 to 50 years). In addition, five 0.1-ha plots of old-growth forest with low levels of anthropogenic disturbance were selected to serve as controls for the regeneration plots (Fig. S1). In order to maintain spatial independence, all plots were selected so that the minimum distance between them was 1 km. The ages of forest stands were established by interviews with local residents. Plots without a record of shifting cultivation by locals for the last 150 years were considered old growth forest. All plots were established in areas sharing the same soil type (sandy soils) and similar slope (flat terrain), which indicates a similar history of land use across the landscape.
Each plot hosted one sampling unit composed of five portable Van Someren-Rydon traps (following DeVries et al. 1997), with a total of 75 traps across the study area. These traps consisted of cylindrical tubes made with netting (110 cm high × 35 cm in diameter) and an internal cone (30 cm high and 22 cm wide at the opening) at the bottom to prevent butterflies from escaping. The lower part was open and attached to a plywood platform with a distance of 4 cm to allow butterflies to enter the trap. A mixture of banana and sugar cane juice fermented for 48 h was used as attractive bait. Baits were placed inside traps in plastic pots with a perforated cover to prevent butterflies from drowning in the liquid, to avoid feeding by other insects, and to reduce evaporation (Hughes et al. 1998; Freitas et al. 2014).
Sampling was carried out monthly from May 2018 to April 2019, and during each visit, all captured individuals were identified, marked and then released. The traps were suspended between 1 and 1.5 m above soil height and separated from each other by a distance of 30 m. Monthly traps remained open for seven consecutive days and were checked every 48 h, when baits were replaced. As a measure of sampling effort we calculated the number of traps-days, i.e., the total number of traps used in the field (75 traps) multiplied by the total days studied (84 days), giving a total of 6300 traps-days. Voucher specimens of all recorded species (up to 10 vouchers by species—about 1% of the total sampled) were deposited in the entomological collections of the Universidade Federal de Pernambuco, Brazil. The identification of the butterflies was carried out at the species level and the classification follows Lamas (2004), modified following Wahlberg et al. (2009).
Functional traits
Fruit-feeding butterflies were characterized into four functional groups based on morphological and ecological attributes related to microhabitat use, predator avoidance and feeding behavior as follow: (1) wing size, (2) preferred strata (canopy or understory), (3) presence of wing ocelli, and (4) larval host plant (monocots or dicots) (Table S1). Size of the anterior wing (hereafter termed Size) was measured as the distance between the thorax joint and apex (average of up to five individuals per species). Size is directly linked to flight capacity and resistance to desiccation (i.e. larger butterflies are more tolerant to desiccation and can move longer distances through the landscape). Similarly to a large body size, the preference for the canopy (hereafter termed Canopy), an open sunny environment, is also related to resistance to high temperatures and desiccation. In our study, we classified butterflies in terms of presence and absence of wing ocelli (hereafter termed Ocellus), a trait related to deflection of predator attacks. Finally, the use of monocots as larval food (hereafter termed Larval monocots) is associated with species that live in the shaded (and more humid) understory, where grasses are more frequent and persist during the long dry periods of the Caatinga. For more details about these functional traits, see Iserhard et al. (2019).
Explanatory variables
A metric of chronic anthropogenic disturbance was obtained based on the three most important sources of disturbance: (1) disturbance by livestock (livestock pressure index; LPI): animal dung density (goats, sheep, horses and cattle), consumption of vegetation and trampling caused by goats and cattle, (2) descriptors related to firewood and timber harvesting (wood extraction index; WEI): the extraction of dead and live wood for fuel, fence construction, and artisanal production, and (3) descriptors associated with exploitation of non-timber forest products (i.e. medicinal plants, food stuffs for humans, hunting and livestock fodder) (Non-timber forest products index; NTFPI). For NTFPI, we used geographic distances based on remote sensing (e.g. distance to the farms’ headquarters, distance to paved roads (BR-232 or PE-270, two highways at the region), distance to vicinal unpaved roads that give access to the farms and distance to the nearest town) (Ribeiro and Freitas 2012). These metrics were integrated into a single chronic disturbance index (i.e. global chronic disturbance index; GI) using a principal component analysis (PCA) in which the first axis explained 60% of data variance. For more details about GI, see Arnan et al. (2018).
Finally, tree species richness, tree density and biomass (the last considered a proxy of forest age) were used as vegetation attributes (Table S2). Data on aboveground biomass were obtained for each plot from a previous study (Souza et al. 2019). The tree database available to compute biomass was composed of all trees with diameter at soil height (DAS) ≥ 3 cm and total height ≥ 1 m. For more details about the tree database, see Rito et al. (2017a, b).
Data analysis
To estimate sample coverage, diversity estimates for rarefied and extrapolated samples with sample completeness were obtained using iNEXT (Hsieh et al. 2016).
The effective numbers of species (so-called Hill numbers, qD) of orders 0 (0D, species richness), 1 (1D, exponential Shannon entropy) and 2 (2D, inverse Simpson concentration) were used to assess changes in fruit-feeding butterfly alpha diversity (Jost 2007). The 0D order is not sensitive to abundances and so gives a disproportionate weight to rare species, while 1D weights each species according to its abundance in the community, without favoring rare or abundant species (i.e. common species) (Jost 2007). Finally, 2D can be interpreted as the number of ‘very abundant’ or ‘dominant’ species in the community (Jost 2006). Rank-abundance distribution plots were examined to compare the most abundant species of fruit-feeding butterflies. In terms of beta diversity, a species partitioning method (i.e. multiple-site dissimilarity metrics; Baselga 2010) was used to separate total Sørensen dissimilarity (beta diversity, βSØR) into components of dissimilarity due to species replacements (i.e. turnover; βSIM) and dissimilarity due to nestedness (βSNE). Analysis of variance (ANOVA) with Tukey’s post-hoc tests were used to compare abundance and taxonomic diversity of fruit-feeding butterfly assemblages in regenerating and old-growth forest stands.
Functional community composition was calculated using community-weighted means (CWM), which summarize shifts in mean trait values within communities due to environmental selection for certain traits (supporting the mass-ratio hypothesis; Grime 1998). Since different types of traits were combined (i.e. categorical and continuous), the Gower approach was adopted to calculate a species dissimilarity matrix (de Bello et al. 2013). CWMs were computed using the FD package (Laliberté et al. 2015) for R (R Development Core Team 2017).
To analyze changes in species and functional composition regarding both regenerating and old-growth forests, a non-metric multidimensional scaling (NMDS) ordination was performed, based on a Bray–Curtis dissimilarity matrix, calculated from an abundance matrix (Clarke and Gorley 2006). We used ANOSIM (Clarke and Gorley 2006) to test the significance of the relationships between habitat types and species/functional groups. We used SIMPER (PRIMER version 6, Clarke and Gorley 2006) to determine the contribution that individual species and functional groups made toward distinguishing differences in community structure between habitats. NMDS, ANOSIM and SIMPER were carried out using version 6 of PRIMER (Clarke and Gorley 2006).
Finally, generalized linear models (GLMs) were employed to analyze the effects of forest vegetation structure (tree density, tree species richness and biomass) and chronic disturbance on abundance, taxonomic diversity (alpha diversity—Hill numbers of order 0, 1 and 2; beta diversity—species turnover and nestedness) and functional community composition (CWM_Ocellus, CWM_Canopy, CWM_Larval monocots and CWM_Size) of fruit-feeding butterfly assemblages. Gaussian error distributions were assumed for all data (Crawley 2013). The residuals of all GLMs were analyzed to evaluate the suitability of the error distribution and the minimally significant model was selected. A test of the correlation between pairs of predictor variables showed that tree species richness and tree density were significantly correlated (r = 0.7; p < 0.001). Consequently, the tree density was removed from the models. In addition, collinearity among predictor variables in the models were assesses using the variance inflation factor (VIF) in the car package (Fox and Weisberg 2011) for R (R Core Team 2017). All VIF values were lower than 2.0 (ranging from 1.17 to 1.89), therefore suggesting independence (i.e. no collinearity) among predictors (Chatterjee et al. 2000). We constructed seven models, which represent all combinations of explanatory variables and the null model (which includes only the intercept).
The relative effect of each predictor variable was assessed using an information-theoretic approach and multi model inference (Burnham and Anderson 2002). The Akaike's information criterion corrected for small samples (AICc) was calculated for each model. It was considered that the models with a difference in AICc < 2 when compared to the best model (i.e. the one with lowest AICc value) have similar plausibility (Burnham and Anderson 2002). Akaike weights (wi) were used to obtain model-averaged parameter estimates. A given spatial and/or forest attribute was considered to be an important explanatory variable for a given response variable if: (i) it showed a high sum of Akaike weights (i.e. considering each candidate model in which it appeared); and (ii) the model-averaged parameter estimates did not include zero (i.e. the model-averaged unconditional variance was lower than the model averaged parameter estimate) (Burnham and Anderson 2002; Grueber et al. 2011). All models were built using the package glmulti for R version 3.0.1 (Calcagno and Mazancourt 2010). The goodness-of-fit of each model was defined as: (explained deviance by the complete model/explained deviance by the null model) × 100 (Crawley 2013). These analyses were run using R (R Development Core Team 2017).
Results
A total of 20,252 individuals of fruit-feeding butterflies from 22 species within four subfamilies of Nymphalidae was recorded across the 15 forest stands (Table S3). Biblidinae was the most species-rich and abundant subfamily (9 species and 10,613 individuals), followed by Charaxinae (7 species and 7447 individuals), Satyrinae (4 species and 2175 individuals) and Nymphalinae (2 species and 17 individuals) (Table S3). Butterfly assemblages were dominated by few species, with three of them, Hamadryas februa februa (8971 individuals), Fountainea halice moretta (3316 individuals) and Fountainea glycerium cratais (2964 individuals) accounting for near 75% of all captured individuals (Fig. S2). A total of 15,008 individuals of 19 species were recorded across regenerating forest stands, while 5244 individuals of 19 species were recorded in the old-growth forest (Table S3).
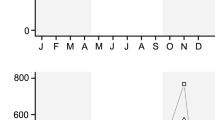
Observed species richness in both regenerating and old-growth forests comprised about 89% and 95% of the estimated richness, but coverage-based sampling curves detected no significant difference in the species richness of these forest types (Fig. S3). The abundance of fruit-feeding butterfly assemblages varied greatly across the chronosequence, with late-successional forest stands supporting more individuals (Fig. 1). Although regenerating forests supported more individuals than old-growth forests; however, this difference was not significant in terms of mean scores (ANOVA; F = 3.64; p = 0.07). Some regenerating stands exhibiting diversity scores higher than the mean score from old-growth forests (Fig. 2). Fruit-feeding butterfly beta diversity was low with most of the variation (74%) due to species replacement rather than variation in nestedness. Interestingly, the early-regenerating stands exhibited the highest values of species turnover (Fig. S4), although old-growth and regenerating forests did not differ relative to either nestedness (ANOVA; F = 2.15; p = 0.16) or species turnover (ANOVA; F = 0.02; p = 0.82) (Fig. S4).

Relative abundance of fruit feeding-butterfly species along regenerating forest stands. Subspecies names were omitted for the sake of simplicity

Relationship between forest age and community-level attributes of fruit-feeding butterflies (including fitted models ± 95% confidence intervals)
Forest habitats were slightly different relative to both taxonomic and functional composition (Fig. S5) as confirmed by ANOSIM tests (R = 0.55, p = 0.001; R = 0.34, p = 0.02; respectively) and as suggested by the changes in abundance of particular species along the forest regeneration chronosequence (see Fig. 1). More precisely, average cross-habitat dissimilarity (via SIMPER analysis) indicated a taxonomic and functional segregation of 14% and 12% between regenerating and old-growth forest stands (Tables S4, S5), respectively. Opsiphanes invirae remoliatus (12%) and Pharneuptychia sp. 1 (9%) were the two species that most contributed to habitat taxonomic differentiation (Table S4), while small-bodied butterfly species presenting wing ocelli and monocot-feeding larvae (26%), as well as large-bodied and canopy-inhabiting butterflies (17%) were the functional groups that most contributed to habitat functional differentiation (Table S5).
In addition to forest successional status (i.e. regenerating vs. old-growth forest), chronic disturbance, biomass and tree species richness also affected butterfly assemblages, as suggested by their high scores of Akaike weights (∑wi) and parameter estimators differing from zero (Fig. 3; Fig. S6; Table 1). As expected, biomass and tree species richness (positively) and chronic disturbance (negatively) were excellent predictors, significantly affecting abundance, and α diversity scores, particularly the total number of species (0D) (Fig. 3; Fig. S6; Table 1). Moreover, chronic disturbance was the most important predictor, positively affecting CWMs for microhabitat (i.e. canopy-inhabiting butterflies), while chronic disturbance had a negative, but non-significant relationship with the CWM for feeding behavior monocot-feeding butterflies (Fig. 4; Fig. S7; Table S6). On the other hand, tree species richness was the predictor that was most positively related to the CWM for ocellus-bearing species (Fig. 4; Fig. S7; Table S6).

Responses of abundance, total species richness (0D), common species (1D) and dominant species (2D) of fruit-feeding butterfly assemblages to biomass (BIO), chronic anthropogenic disturbance (CAD) and tree species richness (TR). The fitted line and shaded area are the linear trends observed ± 95% confidence intervals, respectively

Responses of community-weighted mean fruit-feeding butterfly traits to chronic anthropogenic disturbance (CAD) and tree species richness (TR). The fitted line and shaded area are the linear trends observed ± 95% confidence intervals, respectively. Butterfly traits: Size size of the anterior wing, Ocellus presence of ocellus, Canopy use of the canopy, Larval monocots larval feeding on monocotyledon plants
Discussion
Our results suggest that fruit-feeding butterflies can be abundant, but impoverished in human-modified landscapes of the Caatinga, with a few butterfly species dominating assemblages at both the local and landscape level. Moreover, regenerating and old-growth forest stands support assemblages that are very similar relative to several attributes; i.e. high resilience relative to abundance, species richness and diversity. However, regenerating and old-growth forests support relatively distinct assemblages taxonomically and functionally due to changes in species abundance. Apparently, two major groups control assemblage functional profile: large-winged and canopy-inhabiting butterflies, that reach higher abundances in more disturbed habitats (i.e. higher chronic disturbance), while those bearing wing ocelli that fly in the understory and whose larvae feed on monocots respond negatively to disturbance, but benefit from increased forest biomass and tree species richness. Therefore, chronic disturbance, forest biomass, tree species richness and forest successional status emerge as key drivers of species assembly and community organization at the landscape level, including directional taxonomic and functional changes. This implies a successional trajectory marked by predictable changes.
Although the pattern of species richness was not initially considered as our aim, the occurrence of relatively abundant but species-poor assemblages in the studied landscape deserve attention, as it probably helps to explain our uncovered patterns relative to assemblage resilience, successional trajectory and community organization. In terms of recovery/resilience of fruit-feeding butterfly assemblages, some studies indicate high similarity of community-level attributes between secondary and old-growth forests, indicating high resilience (Sáfián et al. 2011; Nyafwono et al. 2014; Sant’Anna et al. 2014). Others highlight contrasting scores, including lower (Barlow et al. 2007) or higher species richness in secondary as compared to old-growth forest (Bobo et al. 2006). Overall, fruit-feeding butterfly assemblages have been found to recover after more than 50 years of land abandonment or reforestation (Sáfián et al. 2011; Sant’Anna et al. 2014; Oloya et al. 2021). Our findings suggest the possibility of a faster recovery (i.e. a 4-year-old forest stand similar to average old-growth forest) for some attributes (e.g. species richness and density), but recovery rate or resilience is apparently site-specific (see Fig. 2). This is because variables other than forest age probably affect butterfly community-level attributes along forest regeneration.
This perspective reinforces the idea that fruit-feeding butterfly assemblages exhibit marked replacements of ecological groups along forest regeneration. However, their taxonomic/functional successional trajectories reflects not only forest age, but also other variables such as biomass, tree species richness and chronic disturbance. In this perspective, the presence of environmental filters (e.g. anthropogenic disturbance) across the landscape can delay succession processes by selecting only some ecological groups more tolerant to adverse conditions, preventing the structure of old-growth forests being reached. The impacts of these variables on butterfly community organization have been reported for other tropical ecosystems (Akite 2008; Beirão et al. 2017), but there is little or even no information on dry forests, particularly on the effects of chronic disturbances on butterfly community organization and resilience across human-modified landscapes.
Although we did not explicitly examine the underlying mechanisms for this high resilience and successional trajectory marked by taxonomic and functional replacements in our focal landscape, we can propose potential explanations. Namely, the intense degradation that Caatinga has experienced (e.g. slash-and-burn agriculture, livestock and charcoal production) since Europeans arrived in the sixteenth century (Tabarelli et al. 2018), coupled with harsh natural conditions (i.e. reduced forest biomass and chronic water deficit) may limit species and ecological groups that require more forested and mesic habitats, while favoring disturbance-adapted species. In fact, in the Caatinga, human disturbances and low precipitation levels are shown to be associated with reduced biomass and more open habitats (Souza et al. 2019; Barros et al. 2021), the proliferation of disturbance-adapted species, including those able to resprout (Barros et al. 2021; Vanderlei et al. 2021) and impoverished woody plant assemblages (Ribeiro and Freitas 2012; Ribeiro et al. 2016; Rito et al. 2017b). Thus, pioneer/colonizer species of plants (e.g. some species of Euphorbiaceae) that are more tolerant to disturbance, tend to benefit from habitats that are more open and reach exceptional levels of biomass (Rito et al. 2017a). This leads to microclimatic conditions and host plant compositions that favor disturbance-adapted butterflies (e.g. open habitat specialists) able to oviposit and feed on them; i.e. winner plants supporting winner butterfly species (see Tabarelli et al. 2012; Filgueiras et al. 2021). We suggest such assemblages will support more generalist, light-demanding and disturbed-adapted butterfly species (e.g. Charaxinae and Biblidinae). These processes result in the lower Caatinga dry forest growing on sandy soils to harbor understory butterfly assemblages that are similar to the assemblages present in the canopies of humid tropical forests, which are dominated by sun-loving species.
On the other hand, butterflies whose larvae feed on monocots (mostly grasses) and are associated with specific habitat conditions (shaded understory and sensitivity to high temperatures and desiccation) are more associated with sites with low disturbance levels and with higher forest complexity in the Caatinga. We refer to shade-demanding species associated with more structured forests (i.e. understory, canopy and emergent layers) such as the Satyrini (Nymphalidae: Satyrinae). This tribe, with hundreds of species, reaches the highest diversity scores across humid ecosystems such as tropical rain forests (Lamas 2004; Marín et al. 2011).
In summary, human-modified landscapes of the Caatinga dry forest support relatively impoverished but resilient assemblages in the context of slash-and-burn agriculture. However, secondary forests are not able to support the assemblages of old-growth forests in terms of their taxonomic and functional dimension, particularly due to patterns of species abundance in key ecological groups. Thus, the provision of ecological functions and consequently ecosystem services may be simplified across secondary forests, which in turn, increases the importance of conservation strategies that focus on tracts of old-growth tropical dry forest. Although relatively simple, recovery of butterfly assemblages is affected by a set of variables, including human chronic disturbance. Accordingly, spatio-temporal organization of butterfly assemblages is relatively predictable (i.e. structured communities) with secondary forests capturing a high proportion of butterfly fauna. As the Caatinga and other dry forests are expected to experience increasing aridity and more intense land use (Silva et al. 2017), the role played by human disturbance vs natural drivers in the occurrence of impoverished butterfly faunas (e.g. the lack of Satyrinae) deserves further investigation.
References
Álvarez-Yépiz JC, Martínez-Yrízar A, Fredericksen TS (2018) Special issue: resilience of tropical dry forest to extreme disturbance events. For Ecol Manag 426:1–6. https://doi.org/10.1016/j.foreco.2018.05.067
Akite P (2008) Effects of anthropogenic disturbances on the diversity and composition of the butterfly fauna of sites in the Sango Bay and Iriiri areas, Uganda: implications for conservation. Afr J Ecol 46:3–13. https://doi.org/10.1111/j.1365-2028.2008.00923.x
Arnan X, Leal IR, Tabarelli M, Andrade JF et al (2018) A framework for deriving measures of chronic anthropogenic disturbance: surrogate, direct, single and multi-metric indices in Brazilian Caatinga. Ecol Indic 94:274–282. https://doi.org/10.1016/j.ecolind.2018.07.001
Arroyo-Rodriguez V, Melo FPL, Martinez-Ramos M, Bongers F, Chazdon RL, Meave JA, Norden N, Santos BA, Leal IR, Tabarelli M (2015) Multiple successional pathways in human-modified tropical landscapes: new insights from forest succession, forest fragmentation and landscape ecology research. Biol Rev 92:326–340. https://doi.org/10.1111/brv.12231
Avila-Cabadilla LD, Stoner KE, Henry M, Álvarez-Añorve MY (2009) Composition, structure and diversity of phyllostomid bat assemblages in different successional stages of a tropical dry forest. For Ecol Manag 258:986–996. https://doi.org/10.1016/j.foreco.2008.12.011
Barlow J, Overal WL, Araujo IS, Gardner TA, Peres CA (2007) The value of primary, secondary and plantation forests for fruit-feeding butterflies in the Brazilian Amazon. J Appl Ecol 44:1001–1012. https://doi.org/10.1111/j.1365-2664.2007.01347.x
Barros MF, Ribeiro EMS, Vanderlei RS, De Paula AS, Silva AB, Wirth R, Cianciaruso MV, Tabarelli M (2021) Resprouting drives successional pathways and the resilience of Caatinga dry forest in human-modified landscapes. For Ecol Manag 482:118881. https://doi.org/10.1016/j.foreco.2020.118881
Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity. Glob Ecol Biogeogr 19:134–143. https://doi.org/10.1111/j.1466-8238.2009.00490.x
Beirão MV, Neves FS, Penz CM, DeVries PJ, Fernandes GW (2017) High butterfly beta diversity between Brazilian Cerrado and Cerrado-Caatinga transition zones. J Insect Conserv 21:849–860. https://doi.org/10.1007/s10841-017-0024-x
Bobo KS, Waltert M, Fermon H, Njokagbor J, Muhlenberg M (2006) From forest to farmland: butterfly diversity and habitat associations along a gradient of forest conversion in Southwestern Cameroon. J Insect Conserv 10:29–42. https://doi.org/10.1007/s10841-005-8564-x
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Calcagno V, Mazancourt C (2010) glmulti: a R package for easy automated model selection with (generalized) linear models. J Stat Softw 34:1–29. https://doi.org/10.18637/jss.v034.i12
Ceccon E, Huante P, Rincón E (2006) Abiotic factors influencing Tropical Dry Forests regeneration. Braz Arch Biol Technol 49:305–312
Chatterjee S, Hadi AS, Price B (2000) Regression analysis by example. Wiley, New York
Chazdon RL, Peres CA, Dent D, Sheil D, Lugo AE, Lamb D, Stork NE, Miller SE (2009) The potential for species conservation in tropical secondary forests. Conserv Biol 23:1406–1417. https://doi.org/10.1111/j.1523-1739.2009.01338.x
Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial. PRIMER-E, Plymouth
Crawley MJ (2013) The R Book, 2nd edn. Wiley, Hoboken
De Bello F, Carmona CP, Mason NWH, Sebastià M-T, Lepš J (2013) Which trait dissimilarity for functional diversity: trait means or trait overlap? J Veg Sci 24:807–819. https://doi.org/10.1111/jvs.12008
DeVries PJ, Murray D, Lande R (1997) Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian rainforest. Biol J Linn Soc 62:343–364. https://doi.org/10.1006/bijl.1997.0155
Dunn RR (2004) Recovery of faunal communities during tropical forest regeneration. Conserv Biol 18:302–309. https://doi.org/10.1111/j.1523-1739.2004.00151.x
Ferraz SFB, Ferraz KMPMB, Cassino CC, Brancalion PHS, Luz DTA, Azevedo TN, Tambosi LR, Metzger JP (2014) How good are tropical forest patches for ecosystem services provisioning? Landsc Ecol 29:187–200. https://doi.org/10.1007/s10980-014-9988-z
Filgueiras BKC, Peres CA, Melo FPL, Leal IR, Tabarelli M (2021) Winner-loser species replacements in human-modified landscapes. Trends Ecol Evol 36:545–555. https://doi.org/10.1016/j.tree.2021.02.006
Fox J, Weisberg S (2011) An {R} companion to applied regression, 2nd edn. SAGE, Thousand Oaks. http://socserv.socsci.mcmaster.ca/jfox/Books/Companion. Accessed 14 April 2020
Freitas AVL, Francini RB, Brown KS Jr (2003) Insetos como indicadores ambientais. In: Cullen JRL, Rudran R, Valladares-Pádua C (eds) Métodos de estudos em biologia da conservação e manejo da vida silvestre. Editora da UFPR. Fundação O Boticário de Proteção à Natureza, Curitiba, pp 125–151
Freitas AVL, Leal IR, Uehara-Prado M, Iannuzzi L (2006) Insetos Como Indicadores De Conservação Da Paisagem. In: Rocha CFD, Bergallo HG, Van Sluys M, Alves MAS (eds) Biologia da Conservação: Essências. RiMa Editora, São Carlos, pp 357–384
Freitas AVL, Iserhard CA, Santos JP, Carreira JYO, Ribeiro DB, Melo DHA, Rosa AHB, Marini-Filho OJ, Accacio GM, Uehara-Prado M (2014) Studies with butterfly bait traps: an overview. Rev Colomb Entomol 40:209–218
Grime JP (1998) Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol 86:902–910. https://doi.org/10.1046/j.1365-2745.1998.00306.x
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 24:699–711. https://doi.org/10.1111/j.1420-9101.2010.02210.x
Guariguata MR, Ostertag R (2001) Neotropical secondary forest succession: changes in structural and functional characteristics. For Ecol Manag 148:185–206. https://doi.org/10.1016/S0378-1127(00)00535-1
Hernández-Ordóñez O, Urbina-Cardona JN, Martínez-Ramos M (2015) Recovery of amphibian and reptile assemblages during old-field succession of tropical rain forests. Biotropica 47:377–388. https://doi.org/10.1111/btp.12207
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol Evol 7:1451–1456. https://doi.org/10.1111/2041-210X.12613
Hughes JB, Daily GC, Ehrlich PR (1998) Use of fruit bait traps for monitoring of butterflies (Lepidoptera: Nymphalidae). Rev Biol Trop 46:697–704. https://doi.org/10.15517/RBT.V46I3.20199
Iserhard CA, Duarte L, Seraphim N, Freitas AVL (2019) How urbanization affects multiple dimensions of biodiversity in tropical butterfly assemblages. Biodivers Conserv 28:621–638. https://doi.org/10.1007/s10531-018-1678-8
Jost L (2006) Entropy and diversity. Oikos 113:363–375. https://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost L (2007) Partitioning diversity into independent alpha and beta components. Ecology 88:2427–2439. https://doi.org/10.1890/06-1736.1
Laliberté E, Legendre P, Shipley B (2015) Measuring functional diversity (FD) from multiple traits, and other tools for functional ecology. Package “FD” for version 1.0-12
Lamas G (2004) Checklist: Part 4A Hesperioidea–Papilionoidea. In: Heppner JB (ed) Atlas of Neotropical Lepidoptera. Association of Tropical Lepidoptera, Gainesville, p xxxiv + 428
Laurance WF, Peres CA (2006) Emerging threats to tropical forests. University of Chicago Press, Chicago
Lohbeck M, Poorter L, Lebrija-Trejos E, Martínez-Ramos M, Meave JA, Paz H, Pérez-García EA, Romero-Pérez IE, Tauro A, Bongers F (2013) Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 94:1211–1216. https://doi.org/10.1890/12-1850.1
Marín MA, Peña C, Freitas AVL, Wahlberg N, Uribe SI (2011) From the phylogeny of the Satyrinae butterflies to the systematics of Euptychiina (Lepidoptera: Nymphalidae): history, progress and prospects. Neotrop Entomol 40:1–13. https://doi.org/10.1590/S1519-566X2011000100001
Martin PA, Newton AC, Bullock JM (2013) Carbon pools recover more quickly than plant biodiversity in tropical secondary forests. Proc R Soc B 280:20132236. https://doi.org/10.1098/rspb.2013.2236
Melo FPL, Arroyo-Rodríguez V, Fahrig L, Martínez-Ramos M, Tabarelli M (2013) On the hope for biodiversity-friendly tropical landscapes. Trends Ecol Evol 28:462–468. https://doi.org/10.1016/j.tree.2013.01.001
Neves FS, Oliveira VHF, Espírito-Santo MM, Vaz-De-Mello FZ, Louzada J, Sanchez-Azofeifa A, Fernandes GW (2010) Successional and seasonal changes in a community of dung beetles (Coleoptera: Scarabaeinae) in a Brazilian Tropical Dry Forest. Nat Conserv 8:160+164. https://doi.org/10.4322/NATCON.00802009
Norden N, Chazdon RL, Chao A, Jiang Y-H, Vílchez-Alvarado B (2009) Resilience of tropical rain forests: tree community reassembly in secondary forests. Ecol Lett 12:385–394. https://doi.org/10.1111/j.1461-0248.2009.01292.x
Nyafwono M, Valtonen A, Nyeko P, Roininen H (2014) Butterfly Community composition across a successional gradient in a human-disturbed Afro-tropical rain forest. Biotropica 46:210–218. https://doi.org/10.1111/btp.12085
Oloya J, Malinga GM, Nyafwono M, Akite P, Nakadai R, Holm S, Valtonen A (2021) Recovery of fruit-feeding butterfly communities in Budongo Forest Reserve after anthropogenic disturbance. For Ecol Manag 491:119087. https://doi.org/10.1016/j.foreco.2021.119087
Poorter L, Bongers F, Aide TM et al (2016) Biomass resilience of Neotropical secondary forests. Nature 530:211–214. https://doi.org/10.1038/nature16512
Quesada M, Sanchez-Azofeifa GA, Alvarez-Añorve M et al (2009) Succession and management of tropical dry forests in the Americas: review and new perspectives. For Ecol Manag 258:1014–1024. https://doi.org/10.1016/j.foreco.2009.06.023
R Development Core Team (2017) R a language and environment for statistical computing. R Foundation for Statistical Computing. http://www.R-project.org. Accessed 21 July 2019
Ribeiro DB, Freitas AVL (2012) The effect of reduced-impact logging on fruit-feeding butterflies in Central Amazon, Brazil. J Insect Conserv 16:733–744. https://doi.org/10.1007/s10841-012-9458-3
Ribeiro EMS, Santos BA, Arroyo-Rodrigues V, Tabarelli M, Souza G, Leal IR (2016) Phylogenetic impoverishment of plant communities following chronic human disturbances in the Brazilian Caatinga. Ecology 97:1583–1592. https://doi.org/10.1890/15-1122.1
Ribeiro-Neto JD, Arnan X, Tabarelli M, Leal IR (2016) Chronic anthropogenic disturbance causes homogenization of plant and ant communities in the Brazilian Caatinga. Biodivers Conserv 25:943–956. https://doi.org/10.1007/s10531-016-1099-5
Rito KF, Arroyo-Rodríguez V, Queiroz RT, Leal IR, Tabarelli M (2017a) Precipitation mediates the effect of human disturbance on the Brazilian Caatinga vegetation. J Ecol 105:828–838. https://doi.org/10.1111/1365-2745.12712
Rito KF, Tabarelli M, Leal IR (2017b) Euphorbiaceae responses to chronic anthropogenic disturbances in Caatinga vegetation: from species proliferation to biotic homogenization. Plant Ecol 218:749–759. https://doi.org/10.1007/s11258-017-0726-x
Sáfián S, Csontos G, Winkler D (2011) Butterfly community recovery in degraded rainforest habitats in the Upper Guinean Forest Zone (Kakum Forest, Ghana). J Insect Conserv 15:351–359. https://doi.org/10.1007/s10841-010-9343-x
Sant’Anna CLB, Ribeiro DB, Garcia LC, Freitas AVL (2014) Fruit-feeding butterfly communities are influenced by restoration age in tropical forests. Restor Ecol 22:480–485. https://doi.org/10.1111/rec.12091
Silva JMC, Leal IR, Tabarelli M (2017) Caatinga: the largest tropical dry forest region in South America. Springer, Cham
Singh SP (1998) Chronic disturbance, a principal cause of environmental degradation in developing countries. Environ Conserv 25:1–2. https://doi.org/10.1017/S0376892998000010
Sociedade Nordestina de Ecologia (2002) Projeto Técnico para a Criação do Parque Nacional do Catimbau/PE. Secretaria de Ciência, Tecnologia e Meio Ambiente de Pernambuco – SECTMA, Recife
Souza DG, Sfair JC, De Paula AS, Barros MF, Rito KF, Tabarelli M (2019) Multiple drivers of aboveground biomass in a human-modified landscape of the Caatinga dry forest. For Ecol Manag 435:57–65. https://doi.org/10.1016/j.foreco.2018.12.042
Specht MJ, Santos BA, Marshall N, Melo FPL, Leal IR, Tabarelli M, Baldauf C (2019) Socioeconomic differences among resident, users and neighbour populations of a protected area in the Brazilian dry forest. J Environ Manag 232:607–614. https://doi.org/10.1016/j.jenvman.2018.11.101
Tabarelli M, Peres CA, Melo FPL (2012) The ‘few winners and many losers’ paradigm revisited: emerging prospects for tropical forest biodiversity. Biol Converv 155:136–140. https://doi.org/10.1016/j.biocon.2012.06.020
Tabarelli M, Leal IR, Scarano FR, Silva JMC (2018) Caatinga: legado, trajetória e desafios ruma à sustentabilidade. Ciênc Cult 70:25–29. https://doi.org/10.21800/2317-66602018000400009
Vanderlei RS, Barros MF, Domingos-Melo A, Alves GD, Silva AB, Tabarelli M (2021) Extensive clonal propagation and resprouting drive the regeneration of a Brazilian dry forest. J Trop Ecol 37:35–42. https://doi.org/10.1017/S0266467421000079
Wahlberg N, Leneveu J, Kodandaramaiah U, Peña C, Nylin S, Freitas AVL, Brower AVZ (2009) Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proc Biol Sci 276:4295–4302. https://doi.org/10.1098/rspb.2009.1303
Acknowledgements
We thank the ICMBio for allowing us to work in Catimbau National Park and landowners for giving us permission to work on their properties. This study was supported by the ‘Conselho Nacional de Desenvolvimento Científico e Tecnológico’ (CNPq, Processes PELD 403770/2012-2 and “Edital Universal” 470480/2013-0) and “Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco” (FACEPE, process APQ-0738-2.05/12, APQ 0138-2.05/14, and PRONEX APQ 0138-2.05/14). This study is part of DHAM’s PhD Thesis at the “Programa de Pós Graduação em Biologia Animal” (PPGBA-UFPE). DHAM acknowledges the “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior” (CAPES, Finance Code 001) for a PhD Scholarship. BKCF acknowledges the FACEPE for a Post-doc Grant (BFP-0141-2.05/21), and AVLF, MT and IRL for Productivity Grants (Processes 304291/2020-0, 310228/2016-6 and 305611/2014-3, respectively). AVLF also acknowledges support from the “Fundação de Amparo à Pesquisa do Estado de São Paulo” (FAPESP 2013/50297-0 and 2021/03868-8) and from the National Science Foundation (NSF DEB-1256742). This publication is part of the “RedeLep – Rede Nacional de Pesquisa e Conservação de Lepidópteros”: SISBIOTA-Brasil/CNPq (Grant 563332/2010-7).
Funding
This study was supported by the ‘Conselho Nacional de Desenvolvimento Científico e Tecnológico’ (CNPq, Processes PELD 403770/2012-2 and “Edital Universal” 470480/2013-0) and “Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco” (FACEPE, process APQ-0738-2.05/12, APQ 0138-2.05/14, and PRONEX APQ 0138-2.05/14).
Author information
Authors and Affiliations
Contributions
DHAM, BKCF and IRL originally formulated the idea, DHAM and AVLF developed methodology, DHAM and BKCF conducted fieldwork, DHAM and BKCF performed statistical analyses, and DHAM, IRL, MT, AVLF and BKCF wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Melo, D.H.A., Freitas, A.V.L., Tabarelli, M. et al. Resilient fruit-feeding butterfly assemblages across a Caatinga dry forest chronosequence submitted to chronic anthropogenic disturbance. J Insect Conserv 27, 467–477 (2023). https://doi.org/10.1007/s10841-023-00470-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-023-00470-2