
Abstract
Vineyard inter-rows are important biodiversity hotspots within agricultural landscapes, especially when they are covered with vegetation. However, little is known on the effects to management intensity on a broad range of surface-dwelling invertebrates and their interaction with vegetation. We assessed the diversity and activity density of ants, beetles, millipedes, mites, spiders, springtails and woodlice using pitfall traps in vineyards with either high management intensity (HI) consisting of frequently tilled inter-rows or low management intensity (LO) with alternating tillage in every second inter-row. The study was performed in the Târnave wine region in Central Romania. We wanted to know whether, (i) vineyard management intensity affects the diversity of plants and invertebrates, and (ii) local habitat characteristics affect species richness of different functional guilds and taxa. Species richness of some invertebrate taxa (Coleoptera, Araneae, Formicidae) did significantly differ between HI and LO vineyards. Only phytophages (some Coleoptera) increased in species richness and activity density with vegetation cover. Vineyard soil properties (organic matter content, pH, P, and K) did not significantly differ between HI and LO vineyards. We conclude that vineyard inter-row management can affect both the conservation of biodiversity and the provision of biodiversity-driven ecosystem services.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Grapevine (Vitis vinifera L.) is among the oldest perennial crops and well adapted to (summer-) dry climates. Vineyard inter-rows provide habitats for a range of plant and above- and belowground animal species, especially when covered with vegetation (Kehinde and Samways 2014; Kratschmer et al. 2018). Organisms colonizing these inter-rows provide various ecosystem services (e.g., primary production, pest control, pollination, erosion mitigation and soil nutrient cycling), while their occurrence and abundance are influenced by a range of factors, including tillage practices (Faber et al. 2017), weeds and cover crops, surrounding landscape structures and applications of agrochemicals for pest management (Sharley et al. 2008; Thomson and Hoffmann 2007).
Inter-row management practices are used for weed control and water conservation and can include intensive tillage (resulting in bare soil), alternating tillage, where only every second inter-row is tilled or no tillage with permanent green cover. Instead of or in addition to tillage, the application of herbicides is another common practice to control the inter-row vegetation (Bauer et al. 2004). The respective plant communities determine the physical structure, climatic conditions, food and nesting resources in most habitats and therefore exert considerable influence on the distribution and interactions of animal species (Lawton 1983; McCoy and Bell 1991) and ecosystem services (Winter et al. 2018). Spontaneous vegetation and/or seeded cover crops have a great impact on abundance and diversity of beneficial arthropods in vineyards (Buchholz et al. 2017; Franin et al. 2016). Flowering plants provide nectar and pollen resources to flower-visiting insects (Ambrosino et al. 2006; Kratschmer et al. 2019). Vegetation cover plays an important role in enhancing the abundance and diversity of arthropod predators and consequently reduces pest densities (Rusch et al. 2017). Management of vineyards may affect local biodiversity of plants and invertebrates (Balog and Markó 2007; Caprio et al. 2015; Hadjicharalampous et al. 2002; Simoni and Castagnoli 2007). Vineyard inter-row management has been shown to affect the number and activity of spiders and ground beetles (Norris and Kogan 2000) and alter interactions between collembolans and spiders (Pfingstmann et al. 2019).
In an agricultural landscape including arable crops, vineyards can be important for erosion control, improvement of soil structure and organic matter content and providing suitable habitat for beneficial arthropods (Altieri 2012; Bommarco et al. 2013). Carabid beetles (Coleoptera: Carabidae) are important components of vineyards (Caprio et al. 2015) and they are potentially important natural agents of pest control as they are polyphagous predators, thus they maintain ecosystem functions and services and promote the sustainability of the agroecosystem (Kromp 1999). Spiders (Arachnida: Araneae) also play an important role in biological control of vineyard pests (Bolduc et al. 2005; Bruggisser et al. 2010; Caprio et al. 2015; Costello and Daane 1998; Gaigher and Samways 2014; Isaia et al. 2006). In vineyards spiders can contribute to control of some economically important pests such as grape berry moths (Addante et al. 2003), mealybugs, leafhoppers and planthoppers (Daane et al. 2008). Springtails and mites are also inhabitants of vineyards (Favretto et al. 1992; Nash et al. 2010) and have a wide range of feeding strategies that contribute to soil organic matter decomposition and thus influence the amount of living and dead organic material and nutrient transfers. In vineyards, ants are used as indicators of ecosystem functioning (Chong et al. 2010; De Bruyn 1999). However, some species of ants, spiders and millipedes are also vectors of trunk disease pathogens such as Petri disease and esca which result in reduced grape yield and quality and ultimately significant financial losses (Moyo et al. 2014). Finally, as important decomposers woodlice are also known to be largely affected by local management and associated habitat characteristics, such as soil humidity, pesticide application or tillage operations (Dauber et al. 2005; Paoletti and Hassall 1999).
Ecological studies in vineyards are of particular interest because vineyards are not only agronomically important but also for the conservation of biodiversity (Márquez-García et al. 2019). Vineyards may host many rare and endangered species and biodiversity in general can be high (Costello and Daane 1998; Isaia et al. 2006). For example, the spider species Erigonoplus globipes (L. Koch, 1872), which is generally rare in Central Europe (Hänggi et al. 1995), can be commonly found in Romanian vineyards. Therefore, vineyards have a special conservation value and an expanded knowledge on the effects of different management practices and the protection of such habitats is of great importance (Winter et al. 2018).
In the present study, we investigated the effect of different inter-row vegetation management practices on biodiversity in central Romanian vineyards. We studied the cover and diversity of plants and diversity and activity densities of surface-dwelling ants, beetles, millipedes, mites, spiders, springtails and woodlice. We compared effects on these taxa in vineyards with high management intensity (HI) with frequently tilled inter-rows and vineyards with low management intensity (LO) with vegetation cover in every second inter-row and herbicide application in the rows. Our aim was to assess whether (i) vineyard management intensity affects the diversity of plants and invertebrates, and (ii) local habitat characteristics affect species richness of different functional guilds and taxa.
Materials and methods
Study area
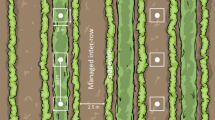
The present study was carried out in 16 Romanian vineyards. The study sites were located in the Târnave wine region, a traditional viticulture region in Transylvania, Romania (46.15971° N/23.92991° E; Fig. 1). Most vineyards are under conventional cultivation (> 90%) and are not irrigated. The trellis system consisted of within-row grapevine distances of 1.0 m and inter-row distances varying between 2.15 and 3.00 m. The regional climate is classified as “Dfb” (D: snow, f: fully humid, b: warm summer) after the Köppen–Geiger Climate Classification (Kottek et al. 2006). The dominant soil type is deep brown soil (Marginean et al. 2013). Average annual temperature is 10.8 °C and average annual precipitation is 544.6 mm (https://www.meteoromania.ro/servicii/date-meteorologice/). The vegetation period starts from the beginning of April and lasts until the end of October.

Location of study area in Romanian vineyards. Main map with location of the studied vineyards including respective tillage regime, HI- high and LO-low
Interviews with wine growers provided information about management practices, such as duration of current management, type of cover crops (spontaneous vs. seed mixture), previous land use type, frequency of herbicide and fungicide application as well as frequency and date of tillage. The raw data used for the present study are given in Table A1 of the electronic supplement S1. Vineyards with high intensive management included different vegetation management practices (tillage frequency inter-row and in-row), fertilization, and weed and fungal control by pesticide application.
Invertebrate sampling
Invertebrates were sampled between 10 and 22 May 2015. In each vineyard, four pitfall traps (diameter 17 mm, depth 60 mm) were used to collect epigaeic invertebrates (ants, beetles, millipedes, mites, spiders, springtails and woodlice) at each sampling site. Traps were filled with ethylene glycol and a drop of odourless detergent and placed about 6 m distance to each other along a transect in the middle of inter-rows. These traps were left in the field for 12 days. Collected invertebrates were stored in 80% ethanol for further analysis (see Buchholz et al. 2017 for details of the sampling method). Pitfall catches reflect the activity and density of species and results should be presented as activity density (Topping and Sunderland 1992). Almost all sampled specimens were identified to species level (Table B1—of the electronic supplement S2) with the use of taxonomic keys (a list is presented in the electronic supplement S3). Species were assigned to four trophic guilds in accordance with (Gaigher and Samways 2010): phytophages, saprophages, omnivores and predators.
Soil analysis
We collected soil samples (5.5 cm diameter, 10 cm depth) within inter-row transects in three replicates to determine soil organic matter (SOM) with the Walkley–Black method (SR ISO 14235:2000), soil carbonates with the Scheibler method (SR ISO 10693:1995), and pH potentiometry (SR ISO 10390:2005), phosphorus (P), and potassium (K) with the Egnèr–Riehm–Domingo method Element concentrations were estimated by photo- (STAS 7184/18-80) and calorimetry (STAS 7184/19-82) (Marin et al. 2017) (Table A2—electronic supplement S2).
Vegetation survey
Vegetation surveys were performed in 2016 in spring (22 April–11 May) and summer (6–15 July) in 12 of the 16 vineyards where invertebrates were sampled 1 year before, because two vineyards were unavailable for sampling and two sites were unexpectedly tilled before vegetation surveys took place. In each vineyard, vegetation surveys were conducted in four 1 × 1 m plots, which were established in the non-tilled inter-rows. Plant species names follow the International Plant Names Index (IPNI). A list of all sampled species can be found in Table B2—electronic supplement S2. Plant trait data for functional diversity indices were obtained from the TRY Database (see Appendix S4 in the online supplementary material for full list of references; Kattge et al. 2011; Kattge et al. 2011) and covered about 66% of species that have been found in the field. For this study, leaf area [mm2], dry mass [g/g], specific leaf area [mm2], leaf nitrogen content [mg/g], plant height [m] and seed mass [mg] (Table B2—electronic supplement S2) were selected due to their availability and application in other related and recent research projects (Ma and Herzon 2014; Negoita et al. 2016; Ma and Herzon 2014; Negoita et al. 2016; Kazakou et al. 2016; Hall et al. 2020).
Data analysis
We used ordinary least squares regression for bivariate comparisons of variables, one-way ANOVA to infer differences in diversity, abundances, and plant functional traits between the two management categories, and PERMANOVA (Bray Curtis dissimilarity, Anderson 2001) to assess respective differences in community composition. We used numbers of traps individuals as a proxy for animal abundances. All analyses were performed with Statistica 12.0 (StatSoft, Hamburg, Germany) and Primer 7.0 (PRIMER-e, Auckland, New Zealand).
The indices of functional diversity (richness, evenness, divergence, dispersion (FDis), Rao’s quadratic entropy) were computed in the R environment (R Core Team 2019) using the FD package with plant trait data obtained from the TRY database (Kattge et al. 2011) (Table B2—electronic supplement S2). Functional richness represents the amount of niche space occupied by a community, whereas functional divergence represents how abundance of species is distributed along a trait axis occupied by the community (Mason et al. 2005). β-diversity was calculated from β = γ/α, where γ and α denote total observed richness of a given taxon and the average of local richness, respectively (Whittaker 1972). We excluded millipedes and woodlice species from statistical analysis due to insufficient field data.
Results
Characterization of the epigeic invertebrate and plants assemblages
In total, 8728 invertebrate specimens were counted, resulting in 149 operational taxonomic units (OTUs); mainly species (spp.) except for 15 at genus level and some mites at family level: 55 spp. of beetles, 31 spp. of spiders, 24 spp. of springtails, 16 spp. of mites, 15 spp. of ants, 5 spp. of woodlice and 3 spp. of millipedes (Table B1—electronic supplement S2). In total, 2237 individuals from 101 spp. were collected in the LO vineyards and 6491 individuals from 106 spp. in the HI vineyards. Mean activity density of epigeic dwelling invertebrates was 86.13 ± 98.37 individuals per LO site and 31.25 ± 45.84 individuals per HI site. Among beetles, the families Carabidae (10 spp.) and Staphylinidae (9 spp.) were the most diverse families. Collembola were the most abundant (53.7% of the material collected), followed by ants (31.5%).
Phytophages were represented by beetles of the families Tenebrionidae, Anthicidae, Cerambycidae, Cetoniidae, Curculionidae and Elateridae. Formicidae were omnivores. Predators were mainly represented by Arachnida, including Acari (1.2%) and Araneae (31.5%); Coleoptera, with: Carabidae (1.2%), Staphylinidae, Cantharidae, Coccinellidae and Monotomidae less than 1%). Saprophages were comprised mostly of Collembola (53.7%), Oniscoidea (6.3%), Diplopoda (1%) and mites (0.7%) in the families: Achipteriidae, Ceratozetidae, Phenopelopidae, Liacaridae, Scheloribatidae and Tectocepheidae. The invertebrate assemblages were mainly dominated by saprophages (mainly Collembola) and omnivores (ants), with these two trophic groups representing 85.2% of the total animals collected. Predators were the third most abundant group, representing 4.13% of the total catches. They were primarily comprised of generalist predators such as Araneae, Coleoptera (mainly Carabidae and Staphylinidae) and some groups of mites. In Araneae, the dominant family was Lycosidae (wolf spiders). Within the predator trophic group, we also noted the presence of Erigonoplus jarmilae (Miller, 1943), which is a new record for the Romanian spider fauna. This species was known before only from Austria, Czech Republic, Slovakia, Albania and Russia (Nentwig et al. 2019). This rare species prefers the ground layer or vertical surfaces of the very dry, warm, open habitats, between 200 and 500 m elevation (Buchar and Růžička 2002). The intensively managed vineyards (HI vineyards) harbored more species of animals and larger populations (except Coleoptera) than the vineyards covered with vegetation (LO vineyards) (Table 1). Relative species overlap was only 59% (Table 1).
A total of 99 taxa of plants were identified to species level, which belong to 27 families and five were identified at genus level (Table B2—electronic supplement S2). Most recorded taxa belonged to the Asteraceae family (17 species) followed by the Poaceae family (13 species) with the highest average relative cover of 42.7 ± 20.5% in LO and 33.4 ± 18.5% in HI managed vineyards. Lamiaceae (10 species) and Scrophulariaceae (8 species) were also frequently found in HI and LO vineyards, whereas Fabaceae (8 species) were more frequent in LO vineyards.
Furthermore, we found significant differences in plant community composition and vegetation cover between HI and LO managed vineyards (Table 2). We did not find significant effects of management intensity on plant species richness, abundance, and important species traits (Table 2), as well as on functional diversity (richness, evenness, divergence, dispersion, Rao’s quadratic entropy, not shown). Community composition of species plants between vineyards varied more than between less intensively managed vineyards when using β-diversity for comparison (Table 1). Only two intensively managed vineyards had significantly less vegetation cover than the LO-vineyards (Table B2—electronic supplement S2).
Species richness of some invertebrate taxa (Coleoptera, Araneae, Formicidae) did significantly differ between HI and LO vineyards (Table 3). Activity densities of Collembola and Araneae did significantly differ between HI and LO vineyards (Table 3).
Local habitat characteristics on epigeic invertebrates
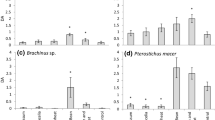
We did not find a consistent influence of vegetation cover on invertebrate species richness and activity density (Fig. 2). Coleoptera seemed to prefer intermediate plant cover (Table 2). Except for phytophages that increased in species richness and activity density with plant cover (Fig. 3), species richness and abundance of the other animal trophic groups did not significantly increase with vegetation cover (Table 2).

Species richness (a) and activity density of (b) Coleoptera (green), Formicidae (blue), Araneae (yellow), and Collembola (red) in response to mean vegetation cover. Ordinary least squares linear (a) and exponential (b) regression lines are not significant at the 5% error level. The bold green line in a refers to a second order polynomial regression where the quadratic term is significant at P < 0.05. The grey rectangles mark the four sites of intense management. (Color figure online)

Species richness (a) and activity density of (b) omnivores (blue), saprophages (yellow), predators (red) and phytophages (green) in dependence on mean annual vegetation cover. Ordinary least squares linear (a) and exponential (b) regression lines are not significant at the 5% error level. The bold green lines in a and b refer to significant regressions at P < 0.05. The grey rectangles mark the four sites of intense management. (Color figure online)
Vineyard soil properties (organic matter content, pH, P, and K) did not significantly differ between HI and LO vineyards (one-way ANOVA: all P(F1,13) > 0.05—Table A2, electronic supplement S1). Vegetation cover did correlate positively with soil organic matter content (r = 0.51, permutation P < 0.05). Except for mites where we found strong positive correlations of species richness (r = 0.85, permutation P < 0.01) and activity density (r = 0.88, permutation P < 0.01) with soil organic matter content, invertebrate species richness and abundances were not significantly linked to soil properties.
Discussion
Different effects of vineyard tillage intensity on the diversity of plants and invertebrates
The results of this study show that vineyard management intensity affect the diversity of plants and some invertebrate groups. Lower tillage intensity resulted in higher plant diversity and plant abundance (Table 1), which also increased above-ground biomass in a French study (Kazakou et al. 2016). Vegetation cover differed significantly between vineyards of different management intensity. These findings are relevant for the provision of ecosystem services such as erosion mitigation or carbon sequestration via enhanced higher vegetation cover (Biddoccu et al. 2016; Guzmán et al. 2019; Ruiz-Colmenero et al. 2013). Relative coverage of annual and perennial species also showed a significant response to management intensity, with greater coverage by annuals in vineyards with high management intensity. Tillage destroys the current vegetation cover but it also creates beneficial conditions for seedling establishment of annual species (Gago et al. 2007) such as Amaranthus powelii S. Wats., Chenopodium album L. agg. and Portulaca oleracea L. Perennial plants characteristic of grasslands and therefore less tolerant to tillage such as Lolium perenne L., Plantago lanceolata L. and Trifolium repens L., benefit from mulching in low management intensity vineyards. Consequently, plant community composition of the two inter-row management intensities was significantly different (Table 2). Species diversity of plants and consequently functional diversity clearly increased with less frequent tillage (Tables 1, 2). Functional diversity of plants was also highest in vineyards with lower tillage frequency and spontaneous vegetation in French vineyards (Kazakou et al. 2016).
Despite the similarity of invertebrate assemblages between the two vineyard management systems (HI and LO), we found differences in terms of species diversity and density. Vineyards with high tillage intensity supported higher levels of diversity of invertebrates (with one exception: Coleoptera) than did LO vineyards. Contrary to our study, carabid species (brachypterous group of Coleoptera) were negatively correlated with grass cover and grass height (Caprio et al. 2015). In general, evidence of the effects of management practices on vineyard biodiversity is ambiguous. Several studies described a decline in biological diversity, measured as species richness, the abundance of species or other measures of community structure in high managed vineyards compared to less managed ones: carabid insects and spiders (Caprio et al. 2015). Springtails appeared to be more sensitive to tillage intensity (Buchholz et al. 2017) than to either residue management or N fertilization (Coulibaly et al. 2017). Also, oribatid mites were sensitive to mechanical cultivation of soil (Seniczak et al. 2018). Ant assemblages were negatively affected by soil tillage in vineyards (Sharley et al. 2008).
In some South African vineyards, saprophage communities were most probably favoured by the enhanced detritus-based food webs resulting from increased organic amendments and reduced fungicides (Gaigher and Samways 2010). Although we did not explicitly sample soil arthropods, our findings are in line with others suggesting that soil arthropod biodiversity and functioning is often dependent on agricultural management (Diekötter et al. 2010). Therefore, soil arthropods should be considered as a valuable resource that requires adequate habitat management so that the ecosystem services they supply are enhanced (Gonçalves et al. 2018).
Local habitat characteristics affect species richness of different functional guilds and taxa
In this study, we found that reduced tillage intensity in the LO vineyards contributed to higher diversity of plants. The roots and litter provided by cover crops can supply the appropriate microhabitats for soil phytophages, thus benefiting their populations, which in turn can promote the increase of predators, also resulting in an increase of soil biological diversity (Komatsuzaki 2008). The review of Puig-Montserrat et al. (2017) highlighted the importance of vegetation diversity for enhancing populations of beneficial arthropods in vineyards. Woodcock et al. (2008) showed the positive effects of composition and diversity of plants around the field margins on ground beetle diversity. Our study also showed that phytophages increased in species richness and activity density with plant cover. Fields with dense vegetation cover and high plant diversity usually have more predaceous and parasitic arthropods than weed-free fields (Speight and Lawton 1976). Indeed, higher arthropod diversity was associated with increased compositional and structural diversity of the vegetation in vineyards where vegetation cover was present (Gaigher and Samways 2010). Vineyards that have ground vegetation cover with high plant species diversity have higher activity densities and richness of phytophages (Gonçalves et al. 2018). A diverse plant community can influence beneficial arthropod populations by providing food or habitat resources that might not be found in a simple plant community (Costello and Daane 1998). An increase in plant species richness can potentially support increasing numbers of specialized consumers (Siemann 1998), which in turn can encourage a greater diversity of predators through cascade effects (Hunter and Price 1992).
In a study in Australian vineyards, carabids were positively affected by ground cover consisting of compost and straw, whereas staphylinids were not affected (Thomson and Hoffmann 2007). Carabids seem to be negatively affected by deep ploughing and enhanced by reduced tillage frequency (Kromp 1999). Spider abundance might be affected by small-scale habitat structure for web building (Alaruikka et al. 2002) as some Linyphiidae utilize sheet webs located on or near the ground (Thornhill 1983). Also, the proportion of non-crop land in an agricultural landscape has been shown to influence the abundance of spiders (Schmidt et al. 2008). Predators in vineyards benefit from reduced management intensity and increased heterogeneity (Isaia et al. 2006; Sharley et al. 2008). Agricultural practices have a greater effect on spiders than other invertebrate groups (Isaia et al. 2006; Jeanneret et al. 2003).
The negative correlation between omnivore richness and the percentage of ground cover suggested that vineyards with a higher percentage of ground cover were expected to have lower omnivore richness (Gonçalves et al. 2018). In other studies, ground cover positively influenced the activity of carabids (Cole et al. 2005; Saska et al. 2014), as did the maintenance of mowing residues at the surface (Shearin et al. 2008). Surface mulches may also influence the abundance of a range of invertebrates. Abundance of ground beetles, parasitoid Hymenoptera and spiders collected with pitfall traps were increased by the addition of mulches (Thomson and Hoffmann 2007). Ground cover (weeds and mowing residue) have been shown to be important in the enhancement of ant populations (White et al. 2011). Tests of different mulch material in vineyards show that springtails and ants were most affected (Addison et al. 2013).
Microarthropods, in particular, mites and springtails, are a major component of soil biota and are known to be important contributors to soil formation, organic matter transformation (mites—this study), nutrient cycling, C accumulation and plant and microbial diversity (Costantini et al. 2015). Besides, organisms which are directly associated with vineyard crops (e.g., mites living on grape leaves) presumably are exposed to a generally higher disturbance level (Peverieri et al. 2009) than the taxa we studied in this study. Different taxa react differently not only to an increase in disturbance, but also to different types of intensities (Abensperg-Traun et al. 1996; Zulka et al. 1997).
Conclusions
Our results demonstrate that inter-row management practices affect vegetation cover, diversity of plants and some invertebrates. This can increase the provision of ecosystem services in Romanian vineyards, such as the conservation of species, soil erosion mitigation and potentially natural pest control. In agricultural landscapes, vineyards can harbor otherwise rare species (Altieri and Nicholls 2002; Schmitt et al. 2008) as also shown in this study where the spider Erigonoplus jarmilae (Miller, 1943) was found as a new record for Romania. An identification of practices that encourage the persistence of rare species and the maintenance of natural enemy abundance may therefore help to reduce the necessity for insecticide applications.
References
Abensperg-Traun M, Smith GT, Arnold GW, Steven DE (1996) The effect of habitat fragmentation and livestock grazing on animal communities in remnants of gimlet Eucalyptus salubris woodland in the Western Australia wheatbelt. I. Arthropods. J Appl Ecol 33:1281–1301. https://doi.org/10.2307/2404771
Addante R, Moleas T, Ranieri G (2003) Preliminary investigations on the interaction between spiders (Araneae) and grapevine moth (Lobesia botrana (Denis et Schiffermuller)) populations in Apulian vineyards. IOBC WPRS Bull 26:111–116
Addison P, Baauw AH, Groenewald GA (2013) An initial investigation of the effects of mulch layers on soil-dwelling arthropod assemblages in vineyards. S Afr J Enol Vitic 34:266–271. https://doi.org/10.21548/34-2-1104
Alaruikka D, Kotze JD, Matveinen K, Niemela J (2002) Carabid beetle assemblages along a forested urban-rural gradient in southern Finland. J Insect Conserv 6:195–206. https://doi.org/10.1023/A:1024432830064
Altieri MA (2012) Insect pest management in the agroecosystems of the future. Atti Accademia Nazionale Italiana di Entomologia 60:137–144
Altieri MA, Nicholls CI (2002) The simplification of traditional vineyard based agroforests in northwestern Portugal: some ecological implications. Agrofor Syst 56:185–191
Ambrosino MD, Luna JM, Jepson PC, Wratten SD (2006) Relative frequencies of visits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae), other beneficial insects, and herbivores. Environm Entomol 35:394–400
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Balog A, Markó V (2007) Species composition and community structure of the rove beetles (Coleoptera: Staphylinidae) in an experimental vineyard under different vineyard management systems. Acta Phytopathol Entomol Hung 42:367–376. https://doi.org/10.1556/APhyt.42.2007.2.19
Bauer K, Fox R, Ziegler B (2004) Modern soil management in viticulture. (Moderne Bodenpflege im Weinbau). 1st Editio edn. Österreichischer Agrarverlag und Eugen Ulmer Verlag, Leopoldsdorf, Austria and Stuttgart, Germany
Biddoccu M, Ferraris S, Opsi F, Cavallo E (2016) Long-term monitoring of soil management effects on runoff and soil erosion in sloping vineyards in Alto Monferrato (North–West Italy). Soil Tillage Res 155:176–189. https://doi.org/10.1016/j.still.2015.07.005
Bolduc E, Buddle CM, Bostanian NJ, Vincent C (2005) Ground-dwelling spider fauna (Araneae) of two vineyards in Southern Quebec. Environm Entomol 34:635–645
Bommarco R, Kleijn D, Potts SG (2013) Ecological intensification: harnessing ecosystem services for food security. Trends Ecol Evol 28:230–238. https://doi.org/10.1016/j.tree.2012.10.012
Bruggisser OT, Schmidt-Entling MH, Bacher S (2010) Effects of vineyard management on biodiversity at three trophic levels. Biol Conserv 143:1521–1528
Buchar J, Růžička V (2002) Catalogue of spiders of the Czech Republic. Peres, Praha, p 349
Buchholz J, Querner P, Paredes D, Bauer T, Strauss P, Guernion M, Scimia J, Cluzeau D, Burel F, Kratschmer S, Winter S, Potthoff M, Zaller JG (2017) Soil biota in vineyards are more influenced by plants and soil quality than by tillage intensity or the surrounding landscape. Sci Rep 7:17445. https://doi.org/10.1038/s41598-017-17601-w
Caprio E, Nervo B, Isaia M, Allegro G, Rolando A (2015) Organic versus conventional systems in viticulture: comparative effects on spiders and carabids in vineyards and adjacent forests. Agric Syst 136:61–69. https://doi.org/10.1016/j.agsy.2015.02.009
Chong CS, D'Alberto CF, Thomson LJ, Hoffmann AA (2010) Influence of native ants on arthropod communities in a vineyard. Agric For Entomol 12:223–232. https://doi.org/10.1111/j.1461-9563.2010.00472.x
Cole LJ, McCracken DI, Downie IS, Dennis P, Foster GN, Waterhouse T, Murphy KJ, Griffin AL, Kennedy MP (2005) Comparing the effects of farming practices on ground beetle (Coleoptera: Carabidae) and spider (Araneae) assemblages of Scottish farmland. Biodivers Conserv 14:441–460
Costantini EAC, Agnelli AE, Fabiani A, Gagnarli E, Mocali S, Priori S, Simoni S, Valboa G (2015) Short-term recovery of soil physical, chemical, micro- and mesobiological functions in a new vineyard under organic farming. Soil 1:443–457. https://doi.org/10.5194/soil-1-443-2015
Costello MJ, Daane KM (1998) Influence of ground cover on spider populations in a table grape vineyard. Ecol Entomol 23:33–40
Coulibaly SFM, Coudrain V, Hedde M, Brunet N, Mary B, Recous S, Chauvat M (2017) Effect of different crop management practices on soil Collembola assemblages: a 4-year follow-up. Appl Soil Ecol 119:354–366. https://doi.org/10.1016/j.apsoil.2017.06.013
Daane KM, Cooper ML, Triapitsyn SV, Walton VM, Yokota GY, Haviland DR, Bentley WJ, Godfrey KE, Wunderlich LR (2008) Vineyard managers and researches seek sustainable solutions for mealybugs, a changing pest complex. Calif Agric 62:167–176. https://doi.org/10.3733/ca.v062n04p167
Dauber J, Purtauf T, Allspach A, Frisch J, Voigtlander K, Wolters V (2005) Local vs. landscape controls on diversity: a test using surface-dwelling soil macroinvertebrates of differing mobility. Glob Ecol Biogeogr 14:213–221. https://doi.org/10.1111/j.1466-822X.2005.00150.x
De Bruyn LAL (1999) Ants as bioindicators of soil function in rural environments. Agric Ecosyst Environ 74:425–441. https://doi.org/10.1016/S0167-8809(99)00047-X
Diekötter T, Wamser S, Wolters V, Birkhofer K (2010) Landscape and management effects on structure and function of soil arthropod communities in winter wheat. Agric Ecosyst Environ 476(137):108–112. https://doi.org/10.1016/j.agee.2010.01.008
Faber F, Wachter E, Zaller JG (2017) Earthworms are little affected by reduced soil tillage methods in vineyards. Plant Soil Environ 63:257–263
Favretto MR, Paoletti MG, Caporali F, Nannipieri P, Onnis A, Tomei PE (1992) Invertebrates and nutrients in a Mediterranean vineyard mulched with subterranean clover (Trifolium subterraneum L.). Biol Fertil Soils 14:151–158. https://doi.org/10.1007/BF00346055
Franin K, Barić B, Kuštera G (2016) The role of ecological infrastructure on beneficial arthropods in vineyards. Span J Agric Res 14:1–10. https://doi.org/10.5424/sjar/2016141-7371
Gago P, Cabaleiro C, Garcia J (2007) Preliminary study of the effect of soil management systems on the adventitious flora of a vineyard in northwestern Spain. Crop Prot 26:584–591. https://doi.org/10.1016/j.cropro.2006.05.012
Gaigher R, Samways MJ (2010) Surface-active arthropods in organic vineyards, integrated vineyards and natural habitat in the Cape Floristic Region. J Insect Conserv 14:595–605. https://doi.org/10.1007/s10841-010-9286-2
Gaigher R, Samways MJ (2014) Landscape mosaic attributes for maintaining ground-living spider diversity in a biodiversity hotspot. Insect Conserv Divers 7:470–479. https://doi.org/10.1111/icad.12070
Gonçalves F, Carlos C, Aranha J, Torres L (2018) Does habitat heterogeneity affect the diversity of epigaeic arthropods in vineyards? Agric For Entomol 20:366–379. https://doi.org/10.1111/afe.12270
Guzmán G, Cabezas JM, Sánchez-Cuesta R, Lora Á, Bauer T, Strauss P, Winter S, Zaller JG, Gómez JA (2019) A field evaluation of the impact of temporary cover crops on soil properties and vegetation communities in southern Spain vineyards. Agric Ecosyst Environm 272:135–145. https://doi.org/10.1016/j.agee.2018.11.010
Hadjicharalampous E, Kalburtji KL, Mamolos AP (2002) Soil arthropods (Coleoptera, Isopoda) in organic and conventional agroecosystems (vineyards). Environ Manage 29:683–690. https://doi.org/10.1007/s00267-001-0056-5
Hall RM, Penke N, Kriechbaum M, Kratschmer S, Jung V, Chollet S, Guernion M, Nicolai A, Burel F, Fertil A, Lora A, Sánchez-Cuesta R, Guzmán G, Gómez J, Popescu D, Hoble A, Bunea C-I, Zaller JG, Winter S (2020) Vegetation management intensity and landscape diversity alter plant species richness, functional traits and community composition across European vineyards. Agric Syst 177:102706
Hänggi A, Stöckli E, Nentwig W (1995) 4, (1995) Habitats of Central European spiders—characterisation of the habitats of the most abundant spider species of Central Europe and associated species, vol 4. CNCF, Neuchâtel
Hunter MD, Price PW (1992) Playing chutes and ladders: heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology 73:724–732
Isaia M, Bona F, Badino G (2006) Influence of landscape diversity and agricultural practices on spider assemblage in Italian vineyards of Langa Astigiana (Northwest Italy). Environm Entomol 35:297–307
Jeanneret P, Schüpbach B, Pfiffner L, Walter T (2003) Arthropod reaction to landscape and habitat features in agricultural landscapes. Landsc Ecol 18:253–263
Kattge J, Díaz S, Lavorel S, Prentice IC, Leadley P, Bönisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP et al (2011) TRY—a global database of plant traits. Global Change Biol 17:2905–2935. https://doi.org/10.1111/j.1365-2486.2011.02451.x
Kazakou E, Fried G, Richarte J, Gimenez O, Violle C, Metay A (2016) A plant trait-based response-and-effect framework to assess vineyard inter-row soil management. Bot Lett 163:373–388. https://doi.org/10.1080/23818107.2016.1232205
Kehinde T, Samways MJ (2014) Insect-flower interactions: network structure in organic versus conventional vineyards. Anim Conserv 17:401–409. https://doi.org/10.1111/acv.12118
Komatsuzaki M (2008) Ecological significance of cover crop and no tillage practices for ensuring sustainability of agriculture and ecosystem service. In: Chen J, Guô C (eds) Ecosystem ecology research trends. Nova Science Publishers, New York, pp 177–207
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World Map of the Köppen-Geiger climate classification updated. Meteorol Z 15:259–263. https://doi.org/10.1127/0941-2948/2006/0130
Kratschmer S, Pachinger B, Schwantzer M, Paredes D, Guernion M, Burel F, Nicolai A, Strauss P, Bauer T, Kriechbaum M, Zaller JG, Winter S (2018) Tillage intensity or landscape features: what matters most for wild bee diversity in vineyards? Agric Ecosyst Environ 266:142–152. https://doi.org/10.1016/j.agee.2018.07.018
Kratschmer S, Pachinger B, Schwantzer M, Paredes D, Guzmán G, Goméz JA, Entrenas JA, Guernion M, Burel F, Nicolai A, Fertil A, Popescu D, Macavei L, Hoble A, Bunea C, Kriechbaum M, Zaller JG, Winter S (2019) Response of wild bee diversity, abundance, and functional traits to vineyard inter-row management intensity and landscape diversity across Europe. Ecol Evol 2019:1–13. https://doi.org/10.1002/ece3.5039
Kromp B (1999) Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation impacts and enhancement. Agric Ecosyst Environ 74:187–228
Lawton JH (1983) Plant architecture and the diversity of phytophagous insects. Annu Rev Entomol 28:23–39. https://doi.org/10.1146/annurev.en.28.010183.000323
Ma M, Herzon I (2014) Plant functional diversity in agricultural margins and fallow fields varies with landscape complexity level: conservation implications. J Nat Conserv 22:525–531. https://doi.org/10.1016/j.jnc.2014.08.006
Marginean MC, Tana CM, Tița O (2013) Soil characteristics from Târnave vineyard. SGEM2013 conference proceedings: 663–668 doi:10.5593/SGEM2013/BC3/S13.025
Marin N, Dumitru M, Cioroianu T, Sȋrbu C (2017) Evolution of soil attributes in conservative agriculture. Ann Univ Craiova - Agric Montanol Cadastre Ser 47:330–336
Márquez-García M, Jacobson SK, Barbosa O (2019) Wine with a bouquet of biodiversity: assessing agricultural adoption of conservation practices in Chile. Environ Conserv 46:34–42. https://doi.org/10.1017/S0376892918000206
Mason NWH, Mouillot D, Lee WG, Wilson JB (2005) Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 111:112–118
McCoy ED, Bell SS (1991) Habitat structure: the evolution and diversification of a complex topic. In: Bell SS, McCoy ED, Mushinsky HR (eds) Habitat structure: the physical arrangement of objects in space. Chapman & Hall, London, pp 3–27
Moyo P, Allsopp E, Roets F, Mostert L, Halleen F (2014) Arthropods vector grapevine trunk disease pathogens. Phytopathology 104:1063–1069
Nash MA, Hoffmann AA, Thomson LJ (2010) Identifying signature of chemical applications on indigenous and invasive nontarget arthropod communities in vineyards. Ecol Appl 20:1693–1703. https://doi.org/10.1890/09-1065.1
Negoita L, Fridle JD, Lomolino MV, Mittelhauser G, Craine JM, Weiher E (2016) Isolation-driven functional assembly of plant communities on islands. Ecography 39:1066–1077. https://doi.org/10.1111/ecog.01551
Nentwig W, Blick T, Gloor D, Hänggi A, Kropf C (2019) Spinnen Europas. Version 11.2018. https://www.araneae.nmbe.ch. Accessed 15 Apr 2019
Norris RF, Kogan M (2000) Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sci 48:94–158
Paoletti MG, Hassall M (1999) Woodlice (Isopoda: Oniscidea): their potential for assessing sustainability and use as bioindicators. Agric Ecosyst Environ 74:157–165. https://doi.org/10.1016/S0167-8809(99)00035-3
Peverieri GS, Simoni S, Goggioli D, Liguori M, Castagnoli M (2009) Effects of variety and management practices on mite species diversity in Italian vineyards. Bull Insectol 62:53–60
Pfingstmann A, Paredes D, Buchholz J, Querner P, Bauer T, Strauss P, Kratschmer S, Winter S, Zaller JG (2019) Contrasting effects of tillage and landscape structure on spiders and springtails in vineyards. Sustainability 11:2095
Puig-Montserrat X, Stefanescu C, Torre I, Palet J, Fàbregas E, Dantart J, Arrizabalaga A, Flaquer C (2017) Effects of organic farming and conventional crop management on vineyard biodiversity. Agric Ecosyst Environ 243:19–26. https://doi.org/10.1016/j.agee.2017.04.005
R Core Team (2019) R—a language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/, Vienna, Austria
Ruiz-Colmenero M, Bienes R, Eldridge DJ, Marques MJ (2013) Vegetation cover reduces erosion and enhances soil organic carbon in a vineyard in the central Spain. CATENA 104:153–160. https://doi.org/10.1016/j.catena.2012.11.007
Rusch A, Delbac L, Thiéry D (2017) Grape moth density in Bordeaux vineyards depends on local habitat management despite effects of landscape heterogeneity on their biological control. J Appl Ecol. https://doi.org/10.1111/1365-2664.12858
Saska P, Němeček J, Koprdová S, Skuhrovec J, Káš M (2014) Weeds determine the composition of carabid assemblage in maize at a fine scale. Sci Agric Bohem 45:85–92. https://doi.org/10.7160/sab.2014.450202
Schmidt MH, Thies C, Nentwig W, Tscharntke T (2008) Contrasting responses of arable spiders to the landscape matrix at different spatial scales. J Biogeogr 35:157–166
Schmitt T, Augenstein B, Finger A (2008) The influence of changes in viticulture management on the butterfly (Lepidoptera) diversity in a wine growing region of southwestern Germany. Eur J Entomol 105:249–255. https://doi.org/10.14411/eje.2008.035
Seniczak A, Seniczak S, García-Parra I, Ferragut F, Xamaní P, Graczyk R, Messeguer E, Laborda R, Rodrigo E (2018) Oribatid mites of conventional and organic vineyards in the Valencian Community, Spain. Acarologia 58:119–133. https://doi.org/10.24349/acarologia/20184281
Sharley DJ, Hoffman AA, Thomson LJ (2008) The effect of soil tillage on beneficial invertebrates within the vineyard. Agric For Entomol 10:233–243
Shearin AF, Reberg-Horton SC, Gallandt ER (2008) Cover crop effects on the activity-density of the weed seed predator Harpalus rufipes (Coleoptera: Carabidae). Weed Sci 56:442–450. https://doi.org/10.1614/WS-07-137.1
Siemann E (1998) Experimental tests of effects of plant productivity and diversity on grassland arthropod diversity. Ecology 79:2057–2070
Simoni S, Castagnoli M (2007) The effects of varieties and agronomic practices on acarine populations in Italian vineyards. IOBC-WPRS Bull 30:95–100
Speight MR, Lawton JH (1976) The influence of weed cover on the mortality imposed on artificial prey by predatory ground beetles in cereal fields. Oecologia 23:211–223
Thomson LJ, Hoffmann AA (2007) Effects of ground cover (straw and compost) on the abundance of natural enemies and soil macro invertebrates in vineyards. Agric For Entomol 9:173–179. https://doi.org/10.1111/j.1461-9563.2007.00322.x
Thornhill WA (1983) The distribution and probable importance of linyphiid spiders living on the soil surface of sugar-beet fields. Bull Br Arachnol Soc 6:127–136
Topping CJ, Sunderland KD (1992) Limitations to the use of pitfall traps in ecological studies exemplified by a study of spiders in a field of winter wheat. J Appl Ecol 29:485–491. https://doi.org/10.2307/2404516
White WH, Viator RP, White PM (2011) Effect of post harvest residue and methods of residue removal on ground inhabiting arthropod predators in sugarcane. Am Soc Sugar Cane Technol 31:39–50
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Winter S, Bauer T, Strauss P, Kratschmer S, Paredes D, Popescu D, Landa B, Guzmán G, Gómez JA, Guernion M, Zaller JG, Batáry P (2018) Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: a meta-analysis. J Appl Ecol 55:2484–2495. https://doi.org/10.1111/1365-2664.13124
Woodcock BA, Westburya DB, Tscheulin T, Harrison-Cripps J, Harris SJ, Ramsey AJ, Brown VK, Potts SG (2008) Effects of seed mixture and management on beetle assemblages of arable field margins. Agr Ecosyst Environ 125:246–254. https://doi.org/10.1016/j.agee.2008.01.004
Zulka KP, Milasowszky N, Lethmayer C (1997) Spider biodiversity potential of an ungrazed and grazed inland salt meadow in the National Park ‘Neusiedler See- Seewinkel’ (Austria): implications for management (Arachnida: Araneae). Biodiv Conserv 6:75–88
Acknowledgements
Open access funding provided by Austrian Science Fund (FWF). This research is part of the BiodivERsA project VineDivers (www.vinedivers.eu) and was funded through the BiodivERsA/FACCE JPI (2013–2014 joint call) for research proposals, with the national funders: Austrian Science Fund (FWF grant number I 2044-B25), French National Research Agency (ANR), Spanish Ministry of Economy and Competitiveness (MINECO), Romanian Executive Agency for Higher Education, Research, Development and Innovation Funding (UEFISCDI) and Federal Ministry of Education and Research (BMBF/Germany). W.U. acknowledges funding by an institutional grant of the Nicolaus Copernicus University. The study has been supported by the TRY initiative on plant traits (https://www.try-db.org). The TRY initiative and database is hosted, developed and maintained by J. Kattge and G. Bönisch (Max Planck Institute for Biogeochemistry, Jena, Germany). TRY is currently supported by DIVERSITAS/Future Earth and the German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig. We kindly acknowledge that wine growers provided their vineyards as study sites and information on vineyard management. Thanks to Claudiu Avramescu (IBB) for help with sorting the invertebrate groups. We are very grateful to Prof. Ernest C. Bernard (University of Tennessee, Entomology and Plant Pathology, USA) for comments on an earlier version of this text.
Author information
Authors and Affiliations
Contributions
DP, C-IB, SW, and JGZ designed the experiment. DP, C-IB, NP, SW, SK, JB, PQ and JGZ conducted the experiments and field sampling. WU performed statistical analyses. CF, MM, IN, MS, BM, IU, AG, NP, SW identified species, CF, WU, DP, C-IB, SW, SK, PQ, and JGZ prepared and reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval
The ethics committee of the Institute of Biology Bucharest, Romanian Academy, under the head of Enache Mădălin Iancu PhD, confirms that the interviews included in this manuscript were conducted under informed consent of the interviewed persons and that all data acquired have been anonymized.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fiera, C., Ulrich, W., Popescu, D. et al. Effects of vineyard inter-row management on the diversity and abundance of plants and surface-dwelling invertebrates in Central Romania. J Insect Conserv 24, 175–185 (2020). https://doi.org/10.1007/s10841-019-00215-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-019-00215-0